
Integrated Metabolomics and Network Pharmacology Investigation of Cardioprotective Effects of Myricetin after 1-Week High-Intensity Exercise
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Databases and Software
2.2. Animal Model Establishment and Sample Collection
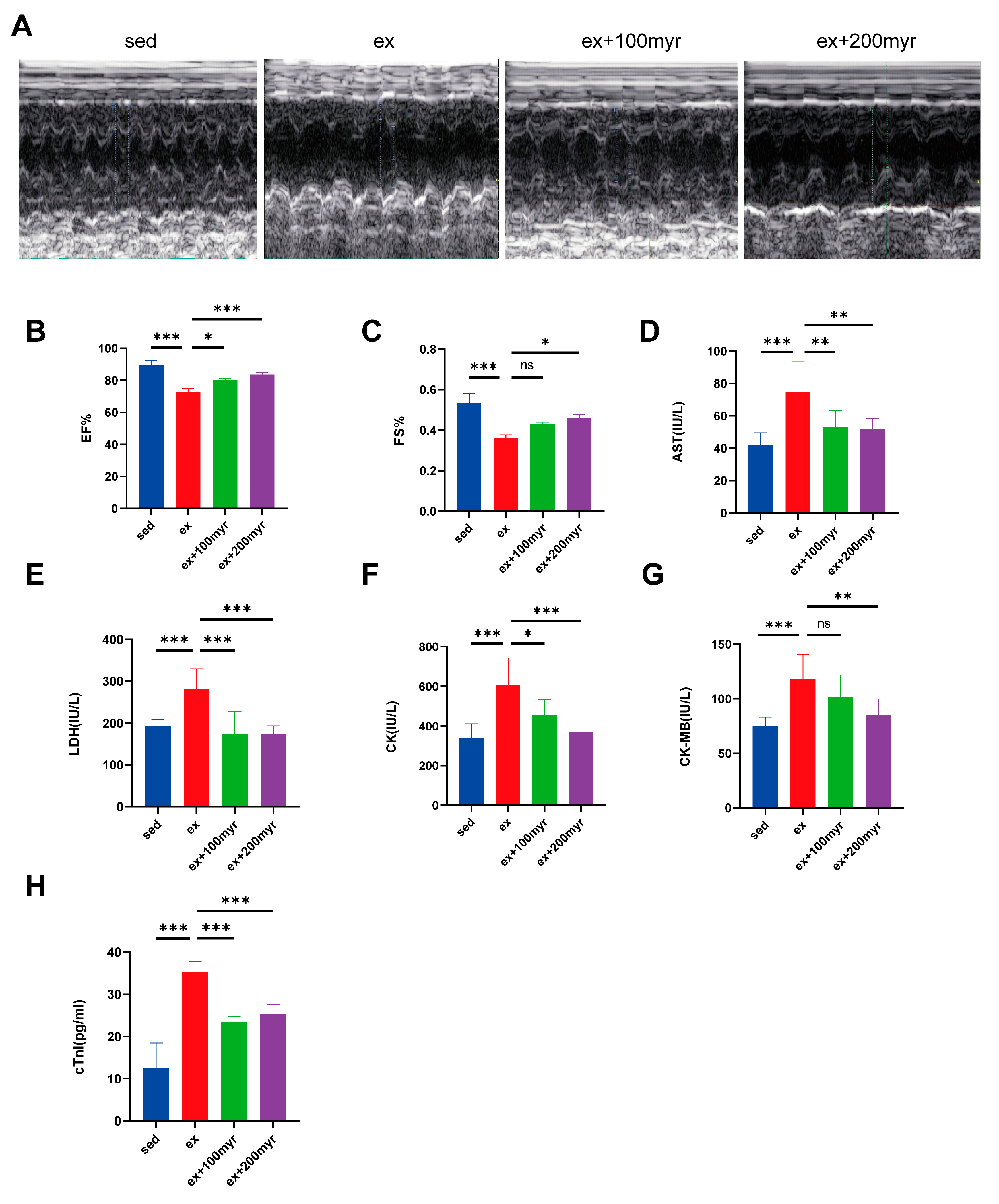
2.3. Echocardiography Analysis
2.4. Serological Analysis and ELISA
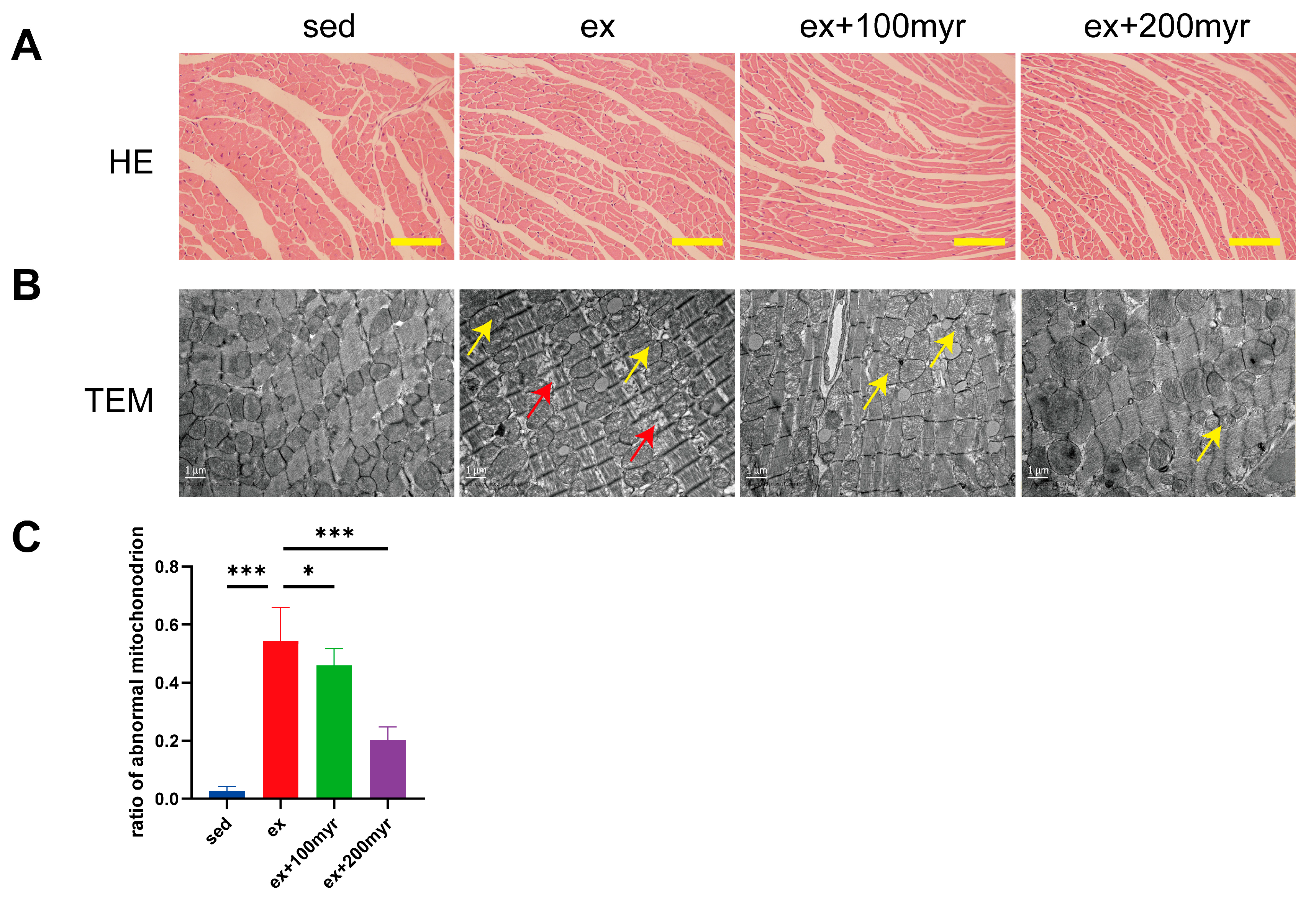
2.5. Hematoxylin and Eosin (HE) Staining
2.6. Transmission Electron Microscopy
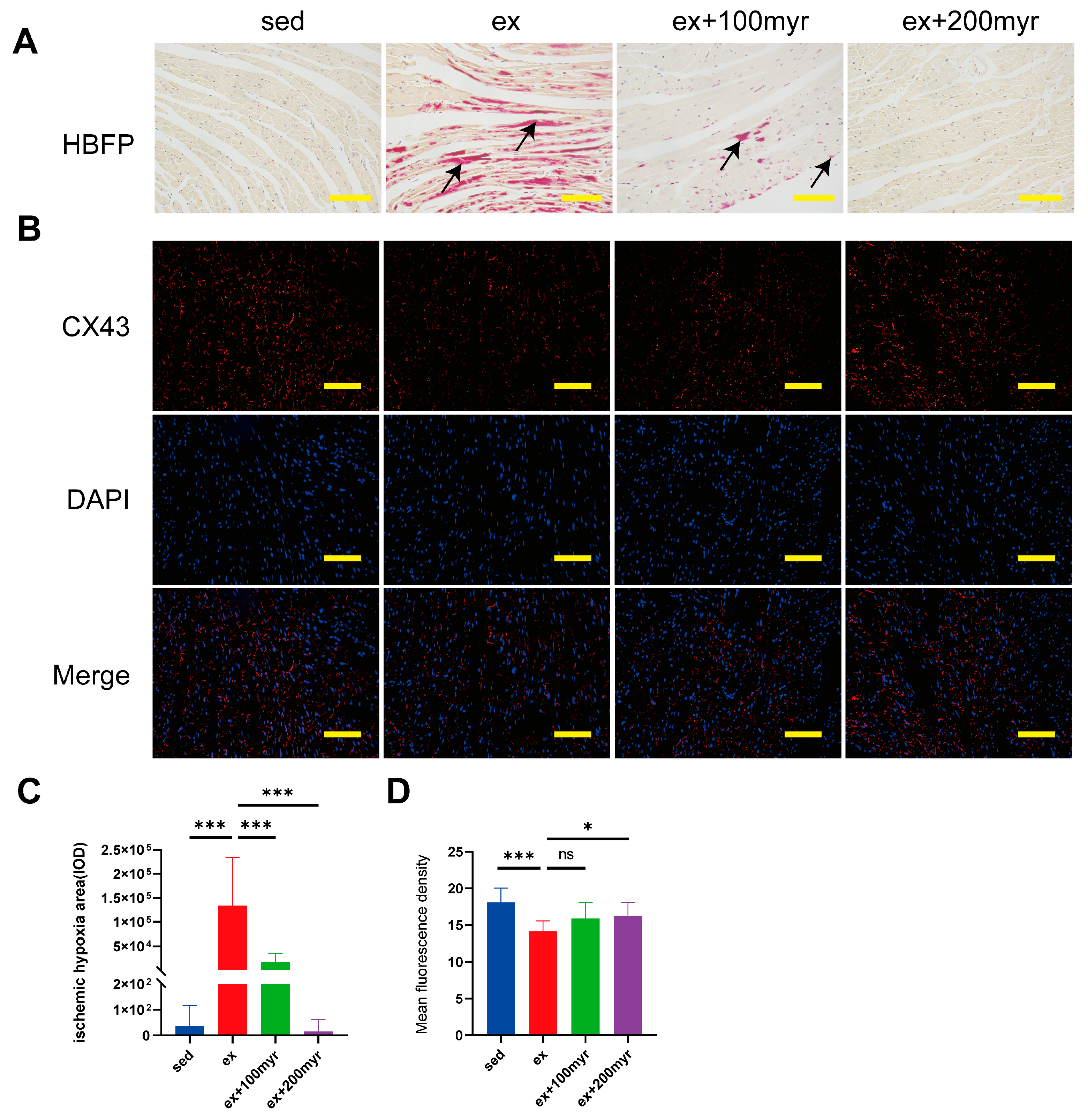
2.7. Hematoxylin-Basic Fuchsin-Picric Acid (HBFP) Staining
2.8. Immunofluorescence Staining
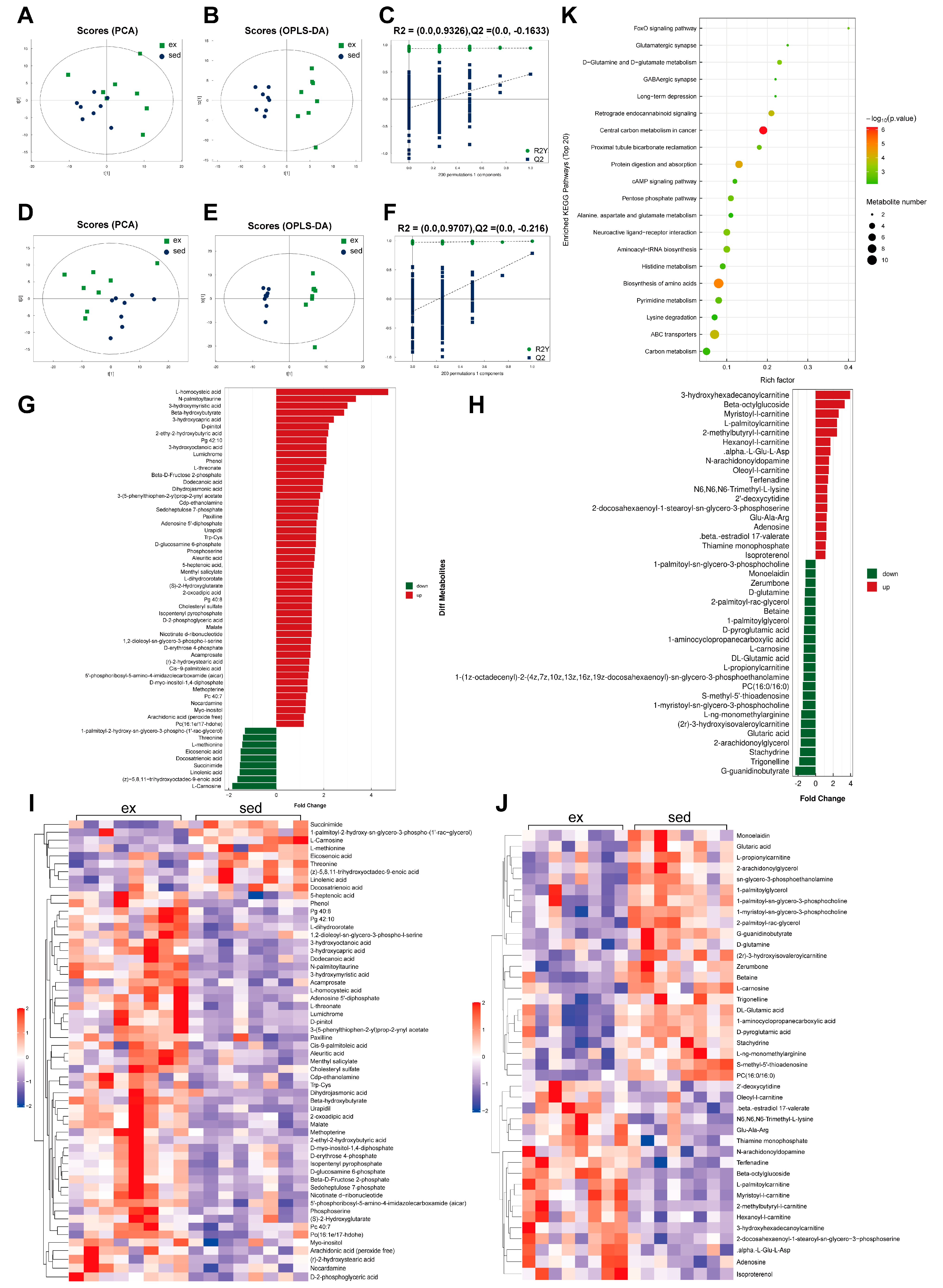
2.9. Bioinformatics Analysis for Metabolomics
2.10. Network Pharmacology Analysis
2.11. Molecular Docking
2.12. Quantitative Real-Time PCR
2.13. Statistical Analysis
3. Results
3.1. Myricetin Improved Cardiac Function and Reduced Myocardial Injury Marker Levels
3.2. Myricetin Alleviated Myocardial Pathological Changes Induced by HIE
3.3. Myricetin Decreased Myocardial Ischemia and Hypoxia Areas and Restored CX43 Content after HIE
3.4. Dramatic Changes in Myocardial Metabolism after HIE
3.5. Network Pharmacology Analysis of Myricetin
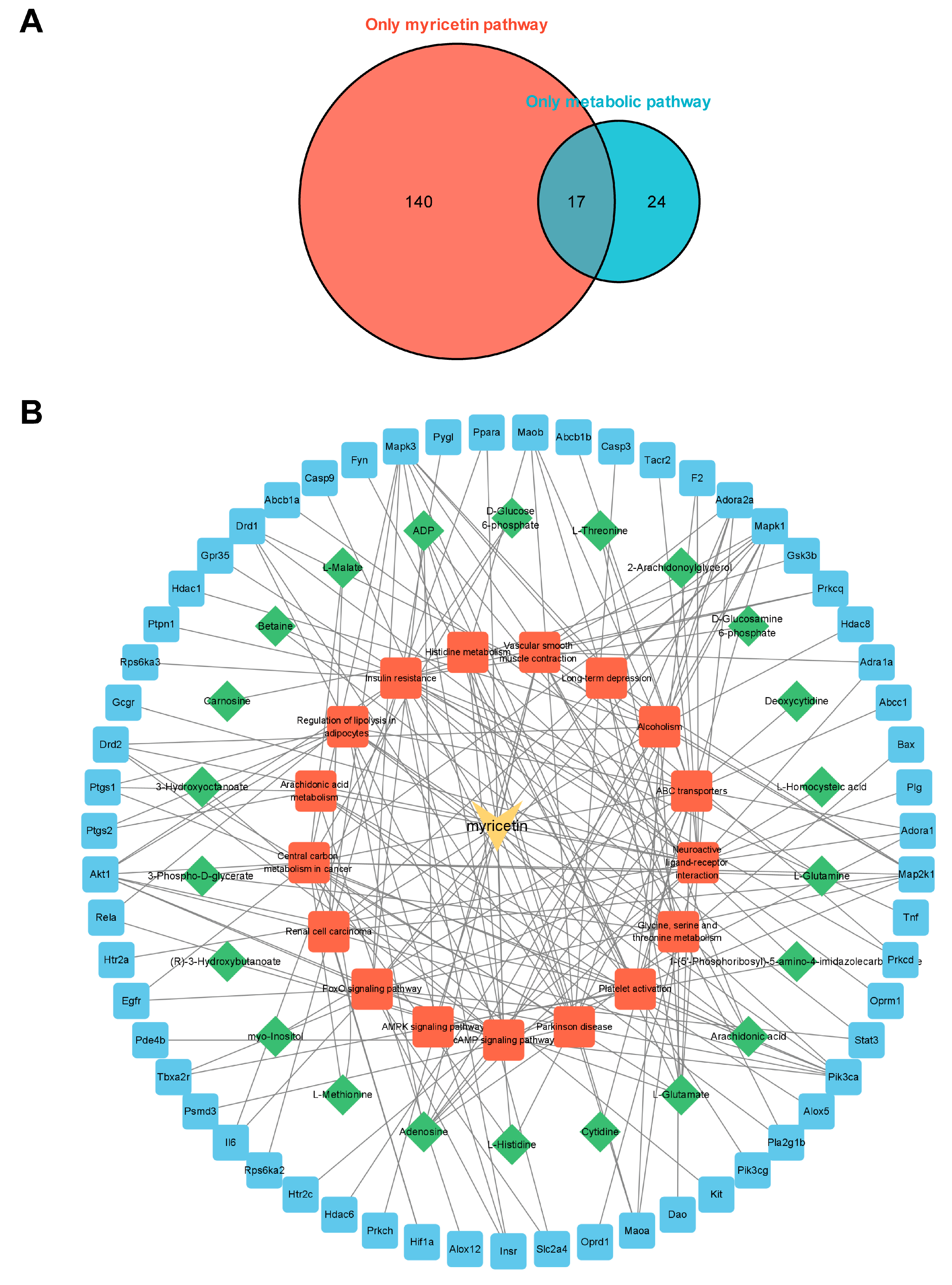
3.6. Integrated Analysis of Metabolomics and Network Pharmacology
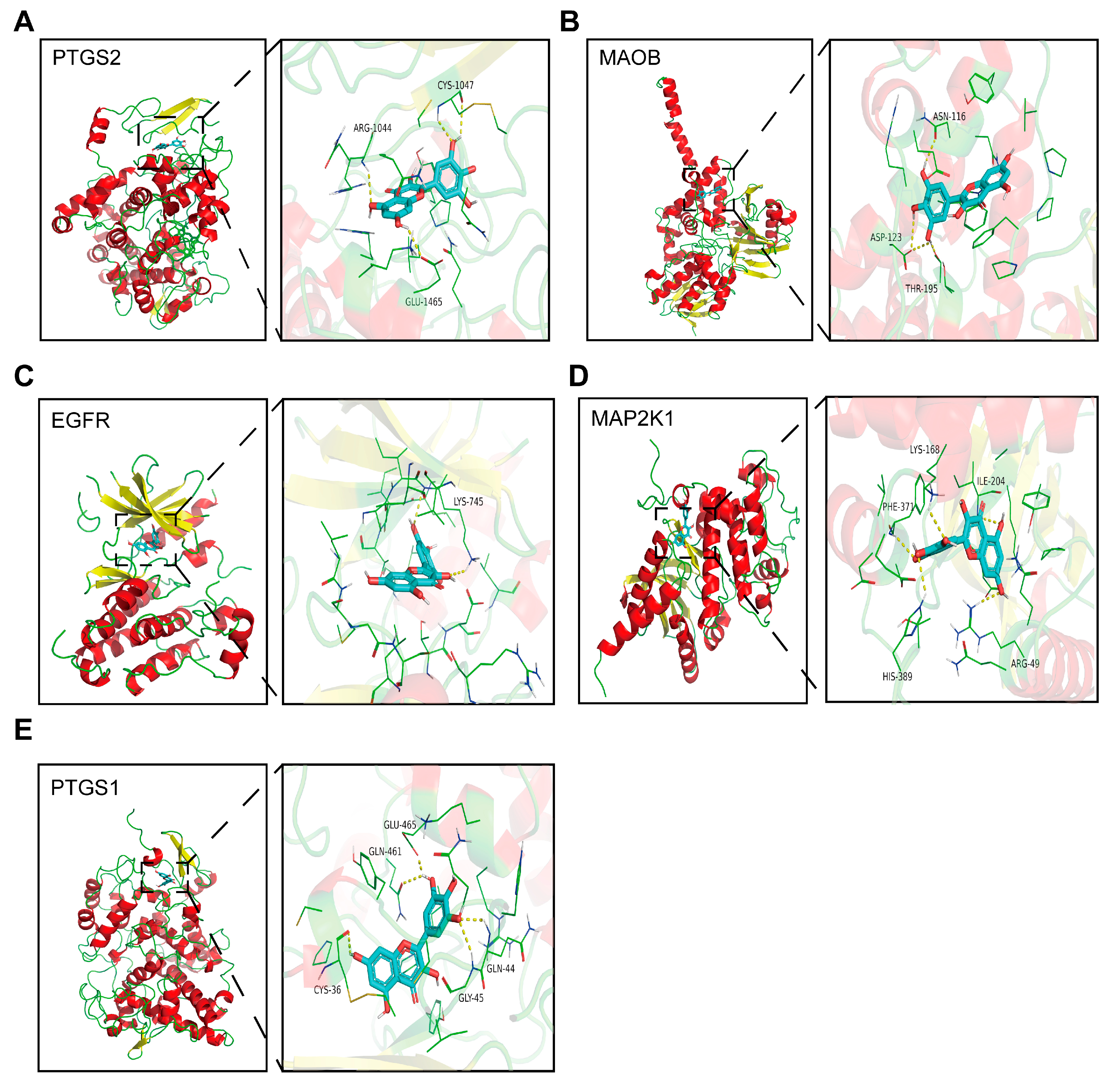
3.7. Molecular Docking
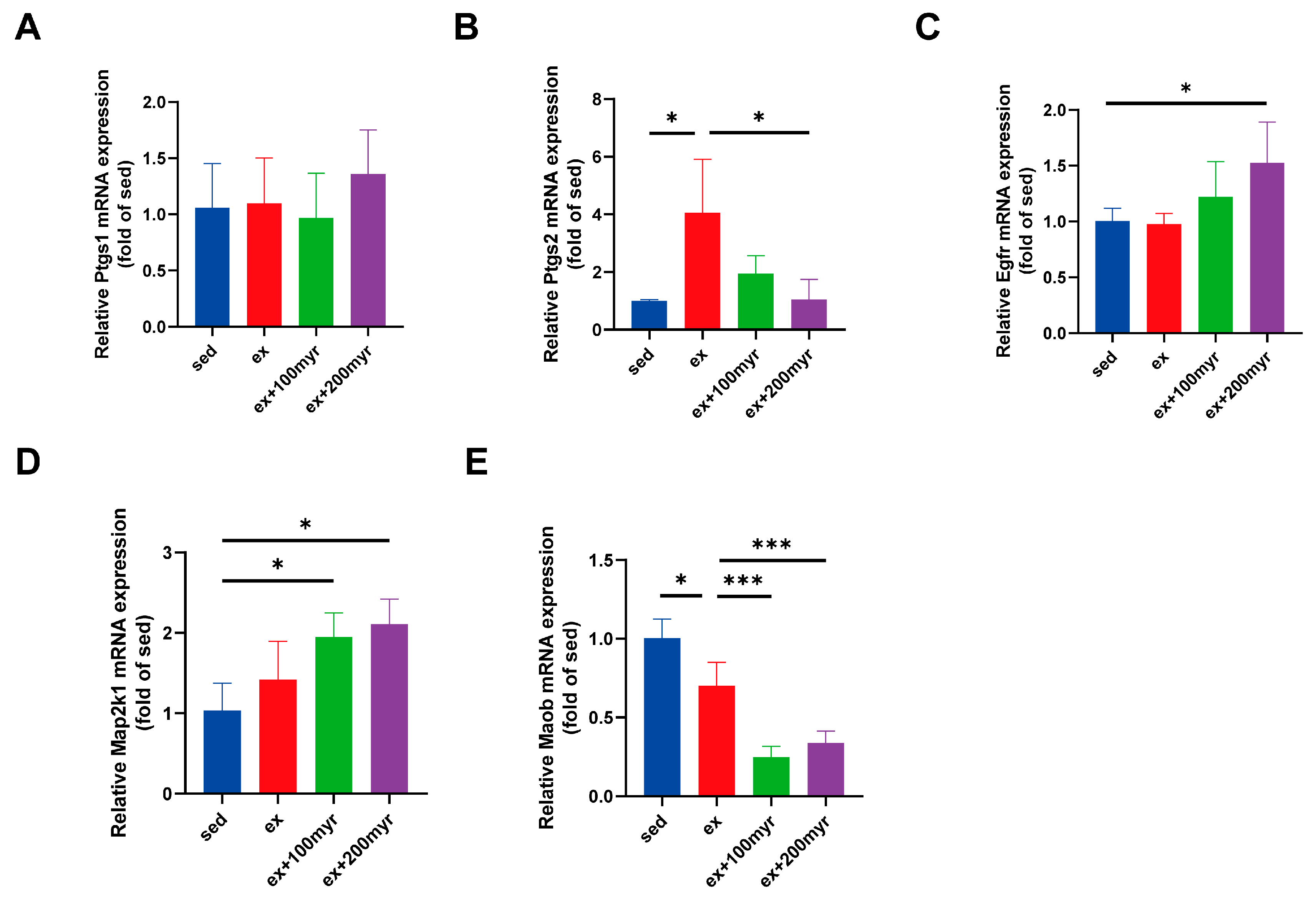
3.8. RT-qPCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ainsworth, B.E.; Haskell, W.L.; Herrmann, S.D.; Meckes, N.; Bassett, D.R., Jr.; Tudor-Locke, C.; Greer, J.L.; Vezina, J.; Whitt-Glover, M.C.; Leon, A.S. 2011 Compendium of Physical Activities: A second update of codes and MET values. Med. Sci. Sports Exerc. 2011, 43, 1575–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merghani, A.; Malhotra, A.; Sharma, S. The U-shaped relationship between exercise and cardiac morbidity. Trends Cardiovasc. Med. 2016, 26, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Harmon, K.G.; Asif, I.M.; Maleszewski, J.J.; Owens, D.S.; Prutkin, J.M.; Salerno, J.C.; Zigman, M.L.; Ellenbogen, R.; Rao, A.L.; Ackerman, M.J. Incidence, Cause, and Comparative Frequency of Sudden Cardiac Death in National Collegiate Athletic Association Athletes: A Decade in Review. Circulation 2015, 132, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa, R. Cardiovascular disease among United States military veterans: Evidence of a waning healthy soldier effect using the National Health Interview Survey. Chronic Illn. 2020, 16, 55–68. [Google Scholar] [CrossRef]
- Ping, Z.; Qiu, W.-J.; Yang, M.; Zhang, X.-L.; Wang, D.-Y.; Xu, P.; Wang, Z.-W.; Cao, X.-B. Optimization of different intensities of exercise preconditioning in protecting exhausted exercise induced heart injury in rats. Sports Med. Health Sci. 2021, 3, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Vega, R.B.; Konhilas, J.P.; Kelly, D.P.; Leinwand, L.A. Molecular Mechanisms Underlying Cardiac Adaptation to Exercise. Cell Metab. 2017, 25, 1012–1026. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, Z.; Zhang, Z.; Zhao, M.; Tong, C.; Cong, P.; Mao, S.; Zhao, Y.; Hou, M.; Piao, Y.; et al. Protective effect and mechanism of cannabidiol on myocardial injury in exhaustive exercise training mice. Chem. Biol. Interact. 2022, 365, 110079. [Google Scholar] [CrossRef]
- Wu, L.; Wang, J.; Cao, X.; Tian, Y.; Li, J. Effect of acute high-intensity exercise on myocardium metabolic profiles in rat and human study via metabolomics approach. Sci. Rep. 2022, 12, 6791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, M.; Zhang, Y.; Li, X. Trimetazidine Attenuates Exhaustive Exercise-Induced Myocardial Injury in Rats via Regulation of the Nrf2/NF-kappaB Signaling Pathway. Front. Pharmacol. 2019, 10, 175. [Google Scholar] [CrossRef]
- Wang, X.; Wu, M.; Lai, X.; Zheng, J.; Hu, M.; Li, Y.; Li, S. Network Pharmacology to Uncover the Biological Basis of Spleen Qi Deficiency Syndrome and Herbal Treatment. Oxid. Med. Cell. Longev. 2020, 2020, 2974268. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Tomas, M.; Ozdal, T.; Capanoglu, E. Effect of food matrix on the content and bioavailability of flavonoids. Trends Food Sci. Technol. 2020, 117, 15–33. [Google Scholar] [CrossRef]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A review of the most recent research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef] [PubMed]
- Bertin, R.; Chen, Z.; Marin, R.; Donati, M.; Feltrinelli, A.; Montopoli, M.; Zambon, S.; Manzato, E.; Froldi, G. Activity of myricetin and other plant-derived polyhydroxyl compounds in human LDL and human vascular endothelial cells against oxidative stress. Biom. Pharmacoth. 2016, 82, 472–478. [Google Scholar] [CrossRef]
- Hassan, S.M.; Khalaf, M.M.; Sadek, S.A.; Abo-Youssef, A.M. Protective effects of apigenin and myricetin against cisplatin-induced nephrotoxicity in mice. Phar. Biol. 2017, 55, 766–774. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.M.; Wang, L.; Yang, D.; Li, C.; Pang, S.T.; Li, X.H.; Li, R.; Yang, B.; Lian, Y.P.; Ma, L.; et al. Wedelolactone alleviates doxorubicin-induced inflammation and oxidative stress damage of podocytes by IkappaK/IkappaB/NF-kappaB pathway. Biomed. Pharmacother. 2019, 117, 109088. [Google Scholar] [CrossRef]
- Zhang, N.; Feng, H.; Liao, H.H.; Chen, S.; Yang, Z.; Deng, W.; Tang, Q.Z. Myricetin attenuated LPS induced cardiac injury in vivo and in vitro. Phytother. Res. 2018, 32, 459–470. [Google Scholar] [CrossRef]
- Chen, S.; Fan, B. Myricetin protects cardiomyocytes from LPS-induced injury. Herz 2018, 43, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Cong, N.; Liang, M.; Wang, Y.; Wang, J. Systems Pharmacology Dissection of the Protective Effect of Myricetin Against Acute Ischemia/Reperfusion-Induced Myocardial Injury in Isolated Rat Heart. Cardiovasc. Toxicol. 2017, 17, 277–286. [Google Scholar] [CrossRef]
- Meng, Z.; Wang, M.; Xing, J.; Liu, Y.; Li, H. Myricetin ameliorates atherosclerosis in the low-density-lipoprotein receptor knockout mice by suppression of cholesterol accumulation in macrophage foam cells. Nutr. Metab. 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, K.A.; da Conceição, A.R.; Pereira, S.M.S.; Oliveira, L.A.; Rodrigues, J.V.D.S.; Dias, R.S.; de Paula, S.O.; Natali, A.J.; da Matta, S.L.P.; Gonçalves, R.V.; et al. Curcumin-Added Whey Protein Positively Modulates Skeletal Muscle Inflammation and Oxidative Damage after Exhaustive Exercise. Nutrients 2022, 14, 4905. [Google Scholar] [CrossRef]
- Wu, Q.; Tao, J.; Wu, L.; Wei, W.; Zhang, X.; Zhang, Z.; Zhang, X.; Jiang, X.; Chen, N. Corn peptide enhances exercise performance and prevents myocardial damage of mice caused by overtraining through activating autophagy. J. Funct. Foods 2022, 97, 105227. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, P.; Zhao, F.; Sun, X.; Ma, W.; Tang, J.; Zhang, C.; Ji, H.; Wang, X. Uncovering the molecular mechanisms of Curcumae Rhizoma against myocardial fibrosis using network pharmacology and experimental validation. J. Ethnopharmacol. 2022, 300, 115751. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Yao, J.; Ni, M.; Wei, J.; Zhong, X.; Guo, W.; Zhang, L.; Wang, R.; Belke, D.; Chen, Y.X. Cardiac ryanodine receptor calcium release deficiency syndrome. Sci. Transl. Med. 2021, 13, eaba7287. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Lavoie, H.; Gagnon, J.; Therrien, M. ERK signalling: A master regulator of cell behaviour, life and fate. Nat. Rev. Mol. Cell Biol. 2020, 21, 607–632. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, E.V.; Tesmer, J.J.; Mushegian, A.; Gurevich, V.V. G protein-coupled receptor kinases: More than just kinases and not only for GPCRs. Pharma. Ther. 2012, 133, 40–69. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, J.; Fontes, S.K.; Bautista, E.N.; Cheng, Z. Physiological and pathological roles of protein kinase A in the heart. Cardiovasc. Res. 2022, 118, 386–398. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef]
- Doenst, T.; Nguyen, T.D.; Abel, E.D. Cardiac metabolism in heart failure: Implications beyond ATP production. Circ. Res. 2013, 113, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.W.; Dai, C.; Liu, W.Z.; Zhang, C.; Zhang, Y.; Yang, G.S.; Guo, Q.H.; Li, S.; Yang, H.X.; Li, A.Y. Gentianella acuta improves TAC-induced cardiac remodelling by regulating the Notch and PI3K/Akt/FOXO1/3 pathways. Biomed. Pharmacother. 2022, 154, 113564. [Google Scholar] [CrossRef]
- Xie, D.; Xiong, K.; Su, X.; Wang, G.; Zou, Q.; Wang, L.; Zhang, C.; Cao, Y.; Shao, B.; Zhang, Y.; et al. Glutamate drives ‘local Ca2+ release’ in cardiac pacemaker cells. Cell Res. 2022, 32, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Zhang, Y.-H.; Zhang, L.; Yang, M.; Chen, C.; Yu, H.; Li, T. Ketogenic diet attenuates post-cardiac arrest brain injury by upregulation of pentose phosphate pathway-mediated antioxidant defense in a mouse model of cardiac arrest. Nutrition 2022, 103–104, 111814. [Google Scholar] [CrossRef] [PubMed]
- Graham, G.G.; Davies, M.J.; Day, R.O.; Mohamudally, A.; Scott, K.F. The modern pharmacology of paracetamol: Therapeutic actions, mechanism of action, metabolism, toxicity and recent pharmacological findings. Inflammopharmacology 2013, 21, 201–232. [Google Scholar] [CrossRef]
- Yang, C.; Li, P.; Wang, P.; Zhu, B.T. Mechanism of reactivation of the peroxidase catalytic activity of human cyclooxygenases by reducing cosubstrate quercetin. J. Mol. Graph Model. 2021, 107, 107941. [Google Scholar] [CrossRef]
- Mayerhofer, A. Peritubular cells of the human testis: Prostaglandin E(2) and more. Andrology 2020, 8, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; An, Q.; Le, L.; Geng, F.; Jiang, L.; Yan, J.; Xiang, D.; Peng, L.; Zou, L.; Zhao, G.; et al. Prospects of cereal protein-derived bioactive peptides: Sources, bioactivities diversity, and production. Crit. Rev. Food Sci. Nutr. 2020, 62, 2855–2871. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Hart, A.; Jatis, A.; Karlan, N.; Akhter, S.; Shih, J.C.; Smith, R.M.; Song, L.-S.; Anderson, E.J. Abstract 12954: A Genotype-Phenotype Link Between Cardiac Monoamine Oxidase Activity and Arrhythmogenic Potential Using a Translational Approach. Circulation 2021, 144, 12954. [Google Scholar] [CrossRef]
- Holecek, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef] [Green Version]
- Lai, Q.; Yuan, G.-Y.; Wang, H.; Liu, Z.-L.; Kou, J.-P.; Yu, B.-Y.; Li, F. Exploring the protective effects of schizandrol A in acute myocardial ischemia mice by comprehensive metabolomics profiling integrated with molecular mechanism studies. Acta Pharmacol. Sin. 2020, 41, 1058–1072. [Google Scholar] [CrossRef]
- Lee, J.K.; Zaidi, S.H.E.; Liu, P.; Dawood, F.; Cheah, A.Y.L.; Wen, W.-H.; Saiki, Y.; Rabinovitch, M. A serine elastase inhibitor reduces inflammation and fibrosis and preserves cardiac function after experimentally-induced murine myocarditis. Nat. Med. 1998, 4, 1383–1391. [Google Scholar] [CrossRef]
- Yang, C.M.; Yang, C.C.; Hsu, W.H.; Hsiao, L.D.; Tseng, H.C.; Shih, Y.F. Tumor Necrosis Factor-alpha-Induced C-C Motif Chemokine Ligand 20 Expression through TNF Receptor 1-Dependent Activation of EGFR/p38 MAPK and JNK1/2/FoxO1 or the NF-kappaB Pathway in Human Cardiac Fibroblasts. Int. J. Mol. Sci. 2022, 23, 9086. [Google Scholar] [CrossRef] [PubMed]
- Eid, R.A.; Alkhateeb, M.A.; Al-Shraim, M.; Eleawa, S.M.; Shatoor, A.S.; El-Kott, A.F.; Zaki, M.S.A.; Shatoor, K.A.; Bin-Jaliah, I.; Al-Hashem, F.H. Ghrelin prevents cardiac cell apoptosis during cardiac remodelling post experimentally induced myocardial infarction in rats via activation of Raf-MEK1/2-ERK1/2 signalling. Arch. Physiol. Biochem. 2019, 125, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ran, L.; Lang, H.; Zhou, M.; Yu, L.; Yi, L.; Zhu, J.; Liu, L.; Mi, M. Myricetin improves endurance capacity by inducing muscle fiber type conversion via miR-499. Nutr. Metab. 2019, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Alam, F.; Mohammadin, K.; Shafique, Z.; Amjad, S.T.; bin Asad, M.H.H. Citrus flavonoids as potential therapeutic agents: A review. Phytother. Res. 2021, 36, 1417–1441. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database | Function | Website |
|---|---|---|
| MetaboLights | Metabolomics data | https://www.ebi.ac.uk/metabolights (accessed on 30 January 2023) |
| SwissTarget prediction | Myricetin intervention target prediction | http://www.swisstargetprediction.ch (accessed on 3 August 2022) |
| String | Protein-protein interaction analysis | https://string-db.org (accessed on 10 August 2022) |
| Metascape | MCODE analysis | https://www.metascape.org (accessed on 10 August 2022) |
| Uniprot | Target protein crystal structure | https://www.uniprot.org (accessed on 25 August 2022) |
| PubChem | 3D structure of myricetin | https://pubchem.ncbi.nlm.nih.gov (accessed on 25 August 2022) |
| Software | Function | Version |
|---|---|---|
| R | GO and KEGG analyses and visualization | 4.2.1 |
| ImageJ | Image analysis | 2.0.0 |
| Cytoscape | Metabolic network visualization | 3.9.1 |
| Chem3D | Analysis of the 3D structure of myricetin | v20 |
| PyMol | Removal of solvent molecules in protein molecules and visualization of molecular docking results | 2.5.2 |
| ADFRsuite | Converting file formats | 1.0 |
| AutoDock Vina | Molecular docking | 1.1.2 |
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Actb | GTGCTATGTTGCTCTAGACTTCG | ATGCCACAGGATTCCATACC |
| Ptgs1 | ATGAGTCGAAGGAGTCTCTCG | GCACGGATAGTAACAACAGGGA |
| Ptgs2 | TTCAACACACTCTATCACTGGC | AGAAGCGTTTGCGGTACTCAT |
| Egfr | GCCATCTGGGCCAAAGATACC | GTCTTCGCATGAATAGGCCAAT |
| Maob | ATGAGCAACAAAAGCGATGTGA | TCCTAATTGTGTAAGTCCTGCCT |
| Map2k1 | AAGGTGGGGGAACTGAAGGAT | CGGATTGCGGGTTTGATCTC |
| Protein | Binding Energy (kcal/mol) | Metabolic Pathway |
|---|---|---|
| PTGS2 | −9.4 | Arachidonic acid metabolism |
| Regulation of lipolysis in adipocytes | ||
| MAOB | −9.3 | Glycine, serine, and threonine metabolism |
| Histidine metabolism | ||
| EGFR | −9.1 | Central carbon metabolism in cancer |
| FoxO signaling pathway | ||
| MAP2K1 | −9.1 | Central carbon metabolism in cancer |
| FoxO signaling pathway | ||
| cAMP signaling pathway | ||
| Vascular smooth muscle contraction | ||
| PTGS1 | −9.1 | Regulation of lipolysis in adipocytes |
| Platelet activation | ||
| Arachidonic acid metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Wang, L.; Wu, L.; Xie, Y.; Chang, M.; Wang, D.; Yi, L.; Zhu, X.; Mi, M. Integrated Metabolomics and Network Pharmacology Investigation of Cardioprotective Effects of Myricetin after 1-Week High-Intensity Exercise. Nutrients 2023, 15, 1336. https://doi.org/10.3390/nu15061336
Li T, Wang L, Wu L, Xie Y, Chang M, Wang D, Yi L, Zhu X, Mi M. Integrated Metabolomics and Network Pharmacology Investigation of Cardioprotective Effects of Myricetin after 1-Week High-Intensity Exercise. Nutrients. 2023; 15(6):1336. https://doi.org/10.3390/nu15061336
Chicago/Turabian StyleLi, Tianyou, Le Wang, Luting Wu, Yingquan Xie, Mengyun Chang, Dawei Wang, Long Yi, Xiaohui Zhu, and Mantian Mi. 2023. "Integrated Metabolomics and Network Pharmacology Investigation of Cardioprotective Effects of Myricetin after 1-Week High-Intensity Exercise" Nutrients 15, no. 6: 1336. https://doi.org/10.3390/nu15061336