
Effects of Long-Term Supplementation of Bovine Colostrum on Iron Homeostasis, Oxidative Stress, and Inflammation in Female Athletes: A Placebo-Controlled Clinical Trial
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
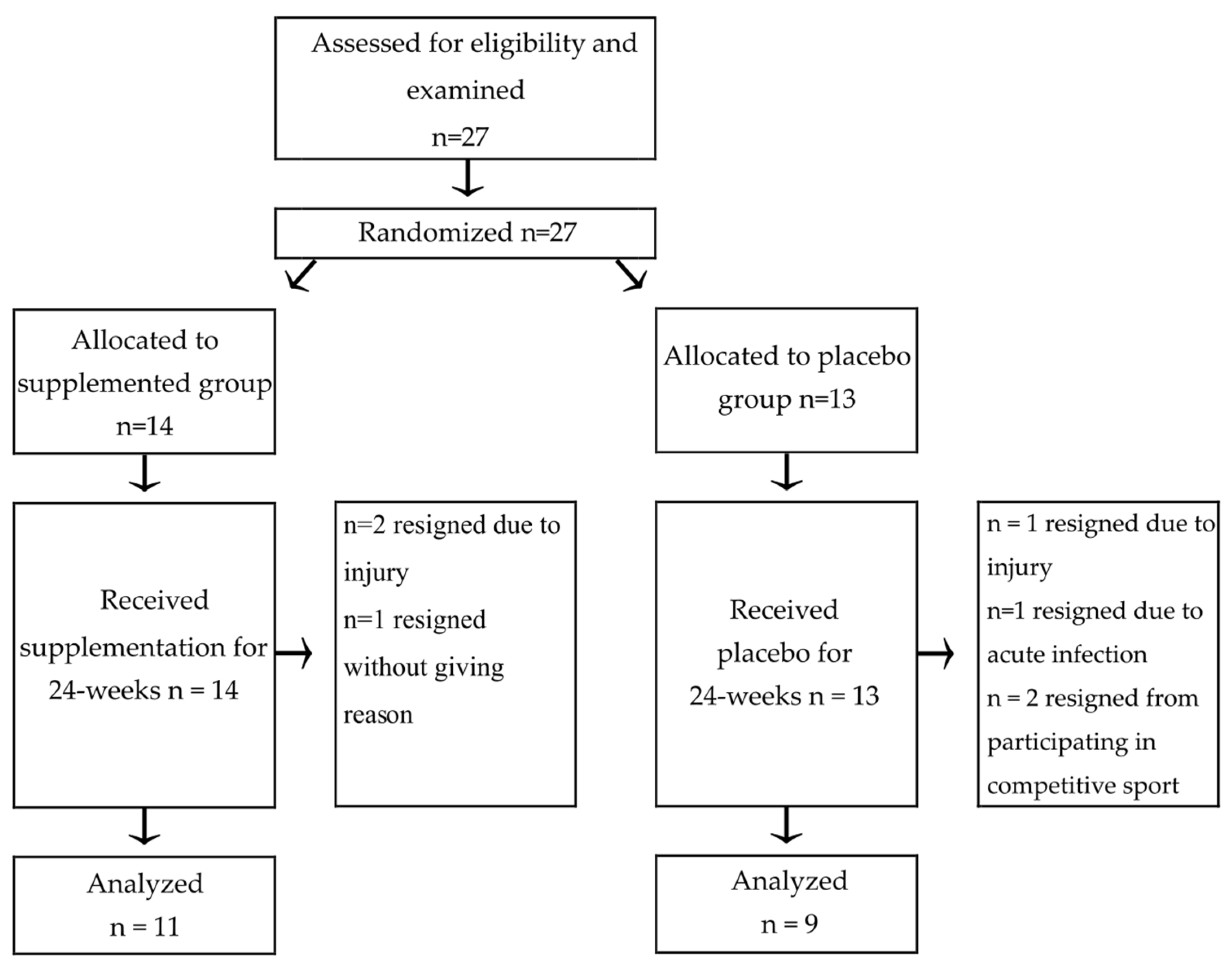
2.1. Participants
2.2. Diet Supplementation
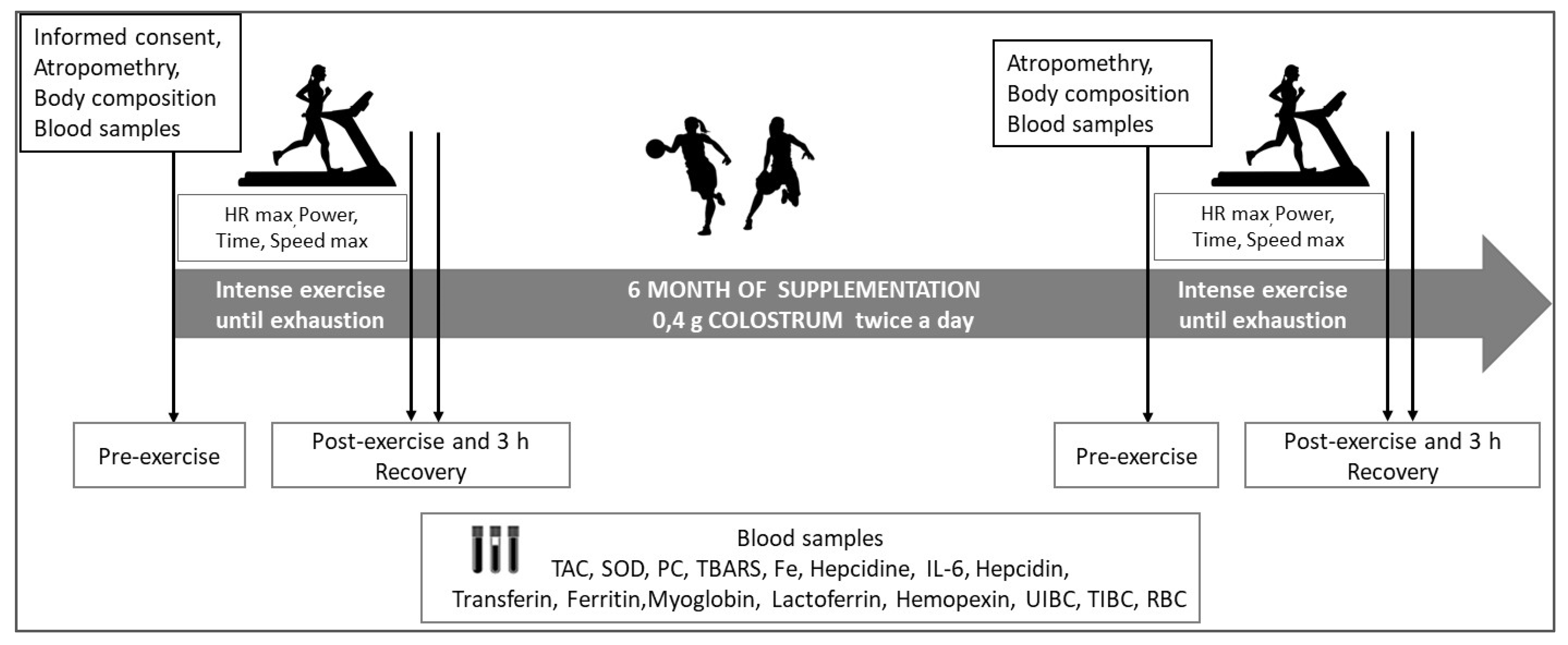
2.3. Physical Performance Examination
2.4. Data Collection and Examination
2.5. Statistical Analysis
3. Results
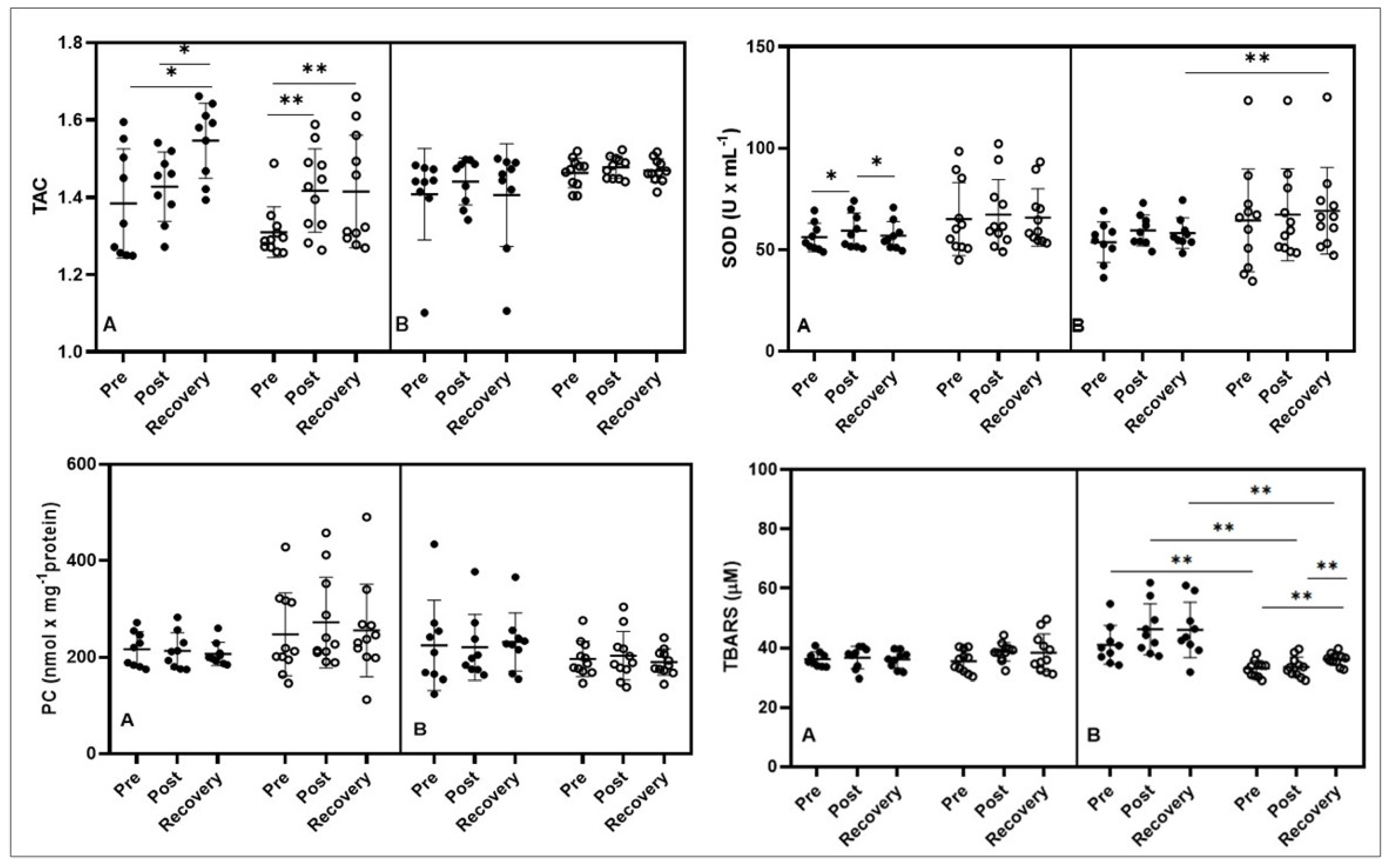
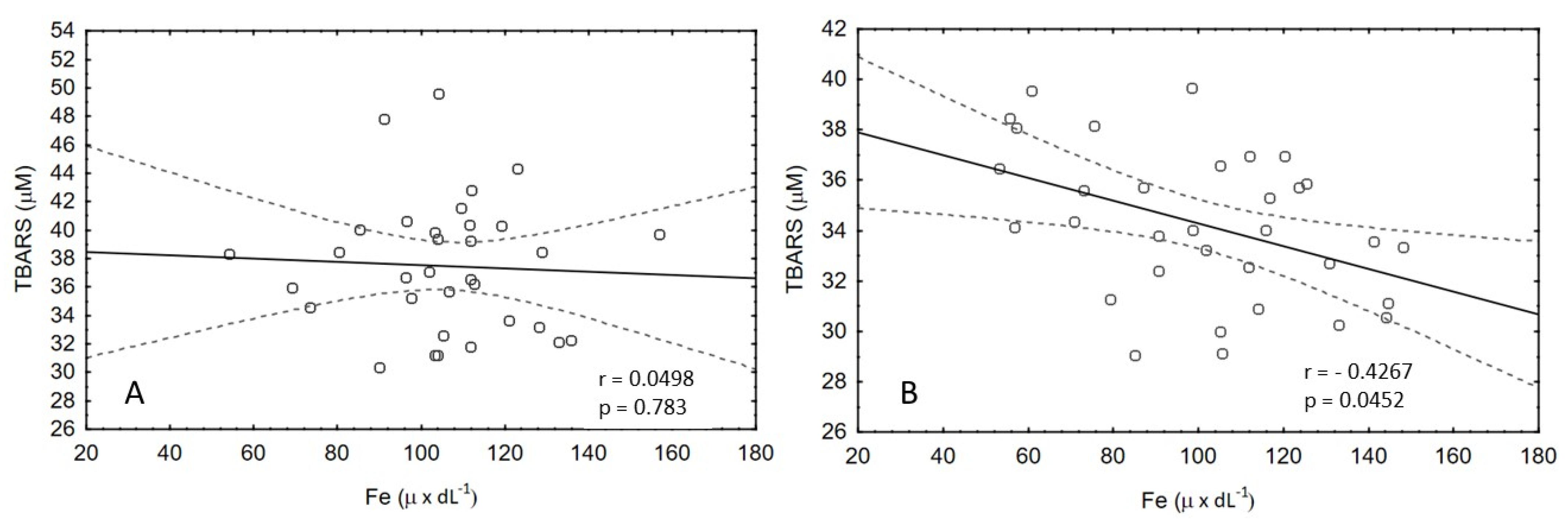
3.1. Oxidative Stress
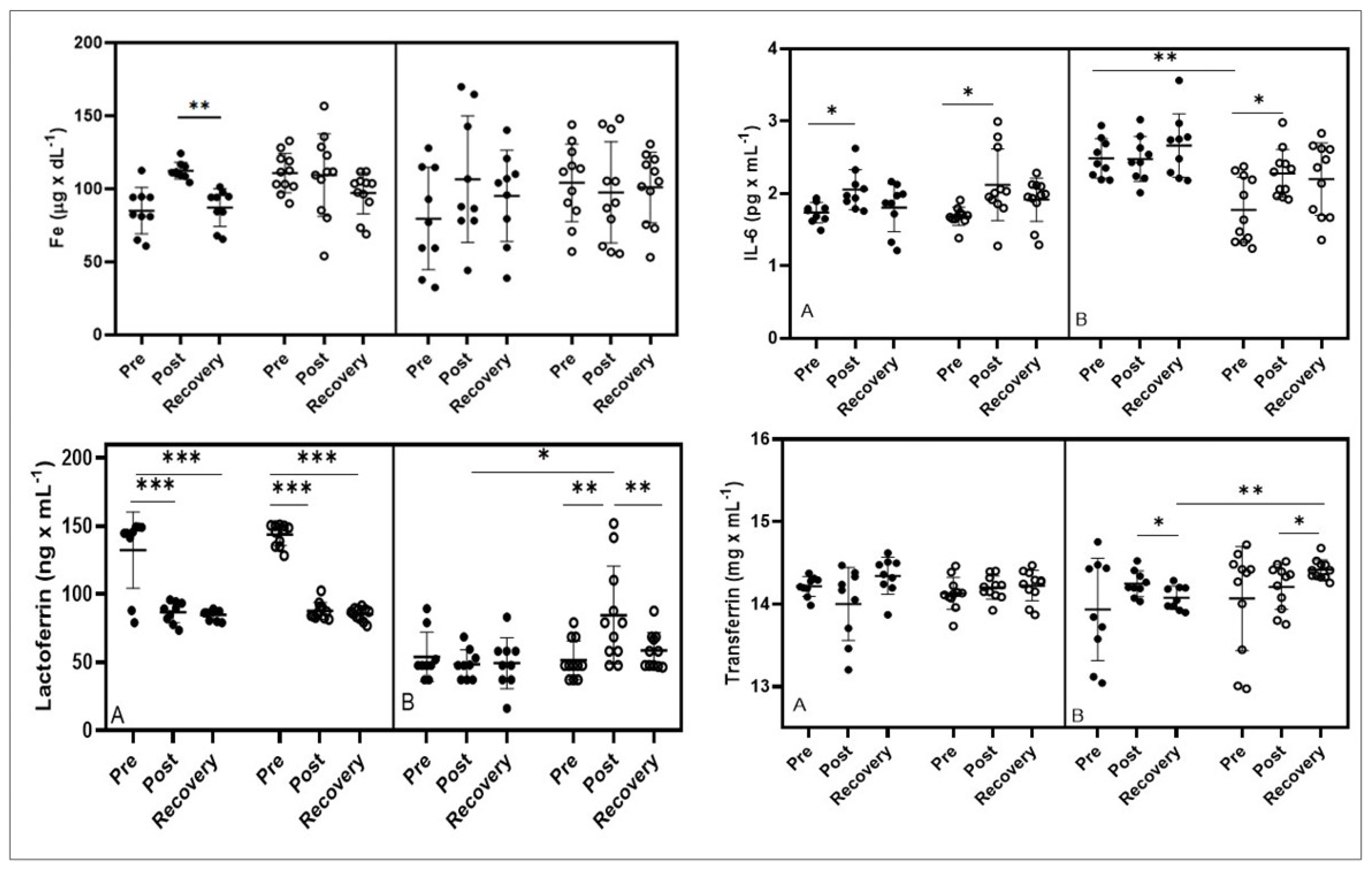
3.2. IL-6 and Iron Homeostasis
3.3. RBC, Hb, HTC, and Myoglobin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BC | bovine colostrum |
| Fe | iron |
| Hb | hemoglobin |
| HR | heart rate |
| HTC | hematocrit |
| IgG | immunoglobulin G |
| IGF | insulin growth factor |
| IL-6 | interleukin 6 |
| αLA | alpha-lactalbumin |
| LZM | lysozyme |
| OH | hydroxyl radical |
| PC | Protein carbonyl |
| Pro | proline |
| PRP | proline-rich polypeptide |
| RBC | red blood cells |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| TAC | total antioxidant capacity |
| TBARS | thiobarbituric acid reactive substances |
| TIBC | total iron-binding capacity |
| UIBC | unsaturated iron-binding capacity |
| Val | valine |
References
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolaou, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K.; et al. Oxidative Stress Biomarkers Responses to Physical Overtraining: Implications for Diagnosis. Free Radic. Biol. Med. 2007, 43, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, J.E.; Amorim, M.R.; Pinto, A.P.; da Rocha, A.L.; da Silva, A.S.R.; Branco, L.G.S. Molecular Hydrogen Downregulates Acute Exhaustive Exercise-Induced Skeletal Muscle Damage. Can. J. Physiol. Pharmacol. 2021, 99, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, T.; Ikemura, T.; Yada, K.; Kanda, K.; Sugama, K.; Ma, S.; Choi, W.; Araya, M.; Huang, J.; Nakamura, N.; et al. The Effects of Beverage Intake after Exhaustive Exercise on Organ Damage, Inflammation and Oxidative Stress in Healthy Males. Antioxidants 2021, 10, 866. [Google Scholar] [CrossRef] [PubMed]
- Bresgen, N.; Eckl, P. Oxidative Stress and Cell Death: The Role of Iron. Free. Radic. Biol. Med. 2018, 124, 558. [Google Scholar] [CrossRef]
- Di Santolo, M.; Stel, G.; Banfi, G.; Gonano, F.; Cauci, S. Anemia and Iron Status in Young Fertile Non-Professional Female Athletes. Eur. J. Appl. Physiol. 2008, 102, 703–709. [Google Scholar] [CrossRef]
- Ponorac, N.; Popović, M.; Karaba-Jakovljević, D.; Bajić, Z.; Scanlan, A.; Stojanović, E.; Radovanović, D. Professional Female Athletes Are at a Heightened Risk of Iron-Deficient Erythropoiesis Compared with Nonathletes. Int. J. Sport Nutr. Exerc. Metab. 2020, 30, 48–53. [Google Scholar] [CrossRef]
- Castell, L.M.; Nieman, D.C.; Bermon, S.; Peeling, P. Exercise-Induced Illness and Inflammation: Can Immunonutrition and Iron Help? Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Albera, E.; Kankofer, M. Antioxidants in Colostrum and Milk of Sows and Cows. Reprod. Domest. Anim. 2009, 44, 606–611. [Google Scholar] [CrossRef]
- Przybylska, J.; Albera, E.; Kankofer, M. Antioxidants in Bovine Colostrum. Reprod. Domest. Anim. 2007, 42, 402–409. [Google Scholar] [CrossRef]
- Chiang, S.-H.; Chang, C.-Y. Antioxidant Properties of Caseins and Whey Proteins from Colostrums. J. Food Drug Anal. 2005, 13, 6. [Google Scholar] [CrossRef]
- Appukutty, M.; Radhakrishnan, A.K.; Ramasamy, K.; Ramasamy, R.; Abdul Majeed, A.B.; Noor, M.I.; Safii, N.S.; Koon, P.B.; Chinna, K.; Haleagrahara, N. Colostrum Supplementation Protects against Exercise-Induced Oxidative Stress in Skeletal Muscle in Mice. BMC Res. Notes 2012, 5, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruzel, M.L.; Polanowski, A.; Wilusz, T.; Sokołowska, A.; Pacewicz, M.; Bednarz, R.; Georgiades, J.A. The Alcohol-Induced Conformational Changes in Casein Micelles: A New Challenge for the Purification of Colostrinin. Protein J. 2004, 23, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Dill, D.B.; Costill, D.L. Calculation of Percentage Changes in Volumes of Blood, Plasma, and Red Cells in Dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Kwak, Y.-S. Impact of Aerobic and Anaerobic Exercise Training on Oxidative Stress and Antioxidant Defense in Athletes. J. Exerc. Rehabil. 2016, 12, 113–117. [Google Scholar] [CrossRef]
- Radak, Z.; Chung, H.Y.; Koltai, E.; Taylor, A.W.; Goto, S. Exercise, Oxidative Stress and Hormesis. Ageing Res. Rev. 2008, 7, 34–42. [Google Scholar] [CrossRef]
- Souissi, W.; Bouzid, M.A.; Farjallah, M.A.; Ben Mahmoud, L.; Boudaya, M.; Engel, F.A.; Sahnoun, Z. Effect of Different Running Exercise Modalities on Post-Exercise Oxidative Stress Markers in Trained Athletes. Int. J. Environ. Res. Public Health 2020, 17, 3729. [Google Scholar] [CrossRef]
- Magherini, F.; Fiaschi, T.; Marzocchini, R.; Mannelli, M.; Gamberi, T.; Modesti, P.A.; Modesti, A. Oxidative Stress in Exercise Training: The Involvement of Inflammation and Peripheral Signals. Free Radic. Res. 2019, 53, 1155–1165. [Google Scholar] [CrossRef]
- Teixeira, V.; Valente, H.; Casal, S.; Marques, F.; Moreira, P. Antioxidant Status, Oxidative Stress, and Damage in Elite Trained Kayakers and Canoeists and Sedentary Controls. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 443–456. [Google Scholar] [CrossRef]
- Ammar, A.; Trabelsi, K.; Boukhris, O.; Glenn, J.M.; Bott, N.; Masmoudi, L.; Hakim, A.; Chtourou, H.; Driss, T.; Hoekelmann, A.; et al. Effects of Aerobic-, Anaerobic- and Combined-Based Exercises on Plasma Oxidative Stress Biomarkers in Healthy Untrained Young Adults. Int. J. Environ. Res. Public Health 2020, 17, 2601. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.P.; Bolin, A.P.; Campoio, T.R.; Guerra, B.A.; Otton, R. Oxidative Stress and Antioxidant Status Response of Handball Athletes: Implications for Sport Training Monitoring. Int. Immunopharmacol. 2013, 17, 462–470. [Google Scholar] [CrossRef]
- Slattery, K.; Bentley, D.; Coutts, A.J. The Role of Oxidative, Inflammatory and Neuroendocrinological Systems during Exercise Stress in Athletes: Implications of Antioxidant Supplementation on Physiological Adaptation during Intensified Physical Training. Sports Med. 2015, 45, 453–471. [Google Scholar] [CrossRef]
- Kruzel, M.L.; Zimecki, M.; Actor, J.K. Lactoferrin in a Context of Inflammation-Induced Pathology. Front. Immunol. 2017, 8, 1438. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-Induced Oxidative Stress: Cellular Mechanisms and Impact on Muscle Force Production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derosa, G.; D’Angelo, A.; Maffioli, P. Change of Some Oxidative Stress Parameters after Supplementation with Whey Protein Isolate in Patients with Type 2 Diabetes. Nutrition 2020, 73, 110700. [Google Scholar] [CrossRef] [PubMed]
- Beneke, R.; Bihn, D.; Hütler, M.; Leithäuser, R.M. Haemolysis Caused by Alterations of Alpha- and Beta-Spectrin after 10 to 35 Min of Severe Exercise. Eur. J. Appl. Physiol. 2005, 95, 307–312. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Nakatsuji, A.; Aoi, W.; Wada, S.; Kuwahata, M.; Kido, Y. Intense Exercise Increases Protein Oxidation in Spleen and Liver of Mice. Nutr. Metab. Insights 2014, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tolosano, E.; Fagoonee, S.; Morello, N.; Vinchi, F.; Fiorito, V. Heme Scavenging and the Other Facets of Hemopexin. Antioxid. Redox Signal. 2010, 12, 305–320. [Google Scholar] [CrossRef]
- Reinke, S.; Taylor, W.R.; Duda, G.N.; von Haehling, S.; Reinke, P.; Volk, H.-D.; Anker, S.D.; Doehner, W. Absolute and Functional Iron Deficiency in Professional Athletes during Training and Recovery. Int. J. Cardiol. 2012, 156, 186–191. [Google Scholar] [CrossRef]
- Zügel, M.; Treff, G.; Steinacker, J.M.; Mayer, B.; Winkert, K.; Schumann, U. Increased Hepcidin Levels During a Period of High Training Load Do Not Alter Iron Status in Male Elite Junior Rowers. Front. Physiol. 2019, 10, 1577. [Google Scholar] [CrossRef]
- Domínguez, R.; Sánchez-Oliver, A.J.; Mata-Ordoñez, F.; Feria-Madueño, A.; Grimaldi-Puyana, M.; López-Samanes, Á.; Pérez-López, A. Effects of an Acute Exercise Bout on Serum Hepcidin Levels. Nutrients 2018, 10, 209. [Google Scholar] [CrossRef]
- Larsuphrom, P.; Latunde-Dada, G.O. Association of Serum Hepcidin Levels with Aerobic and Resistance Exercise: A Systematic Review. Nutrients 2021, 13, 393. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | BC Supplemented Group (n = 11) | Control Group (n = 9) |
|---|---|---|
| Age [years] | 17.09 ± 1.24 | 16.0 ± 0.67 |
| Body mass [kg] Body height [cm] | 67.2 ± 6.66 177.4 ± 5.91 | 65.6 ± 7.02 169.5 ± 4.36 |
| Parameter | Group | Time Point I (mean ± SD) | Time Point II (mean ± SD) |
|---|---|---|---|
| HR max (bpm) | Control | 192.13 (12.29) | 190.9 (8.76) |
| Supplemented | 189.0 (18.46) | 188.5 (7.32) | |
| Power (watt) | Control | 166.3 (21.29) | 162.3 (18.94) |
| Supplemented | 187.4 (17.88) | 187.4 (18.97) | |
| Watt/kg | Control | 2.51 (0.17) | 2.46 (0.16) |
| Supplemented | 2.78 (0.21) | 2.76 (0.15) | |
| Time (s) | Control | 643.33 (96.18) | 596.67 (125.0) |
| Supplemented | 781.82 (126.63) | 774.55 (88.81) | |
| Speed max (km/h) | Control | 12.6 (0.87) | 12.3 (0.80) |
| Supplemented | 13.9 (1.11) | 13.78 (0.84) |
| Variables | Before Supplementation | p Value | After Supplementation | p Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Exercise x ± SD | Post-Exercise x ± SD | Recovery x ± SD | Pre CON vs. SUPL | Post CON vs. SUPL | Recovery CON vs. SUPL | Pre-Exercise x ± SD | Post-Exercise x ± SD | Recovery x ± SD | Pre CON vs. SUPL | Post CON vs. SUPL | Recovery CON vs. SUPL | |
| Hepcidin (ng × dL−1) | ||||||||||||
| PLACEBO | 15.91 ± 1.29 | 14.81 ± 1.95 | 15.25 ± 2.70 | 0.213 | 0.890 | 0.979 | 19.10 ± 3.54 | 22.83 ± 2.60 a | 22.50 ± 3.00 b | 0.460 | 0.402 | 0.149 |
| COLOSTRUM | 14.91 ± 2.29 | 14.55 ± 1.82 | 14.89 ± 2.04 | 17.27 ± 3.83 | 21.19 ± 2.75 a | 18.98 ± 3.33 | ||||||
| Hemopexin (µg × mL−1) | ||||||||||||
| PLACEBO | 614.64 ± 156.38 | 607.97 ± 138.93 | 672.14 ± 181.18 | 0.524 | 0.773 | 0.750 | 613.00 ± 108.27 | 685.78 ± 150.34 | 600.67 ± 179.83 | 0.722 | 0.551 | 0.894 |
| COLOSTRUM | 653.02 ± 144.18 | 614.61 ± 126.08 | 676.20 ± 150.73 | 616.33 ± 140.10 | 714.00 ± 152.41 a | 655.85 ± 114.07 b | ||||||
| Ferritin (ng × mL−1) | ||||||||||||
| PLACEBO | 88.55 ± 11.95 | 91.84 ± 15.84 | 86.43 ± 10.81 | 0.732 | 0.508 | 0.802 | 99.71 ± 43.50 | 90.84 ± 9.13 | 89.27 ± 6.96 | 0.638 | 0.431 | 0.313 |
| COLOSTRUM | 92.51 ± 10.47 | 90.08 ± 11.97 | 89.67 ± 10.74 | 93.84 ± 11.07 | 89.50 ± 8.50 | 86.92 ± 8.02 | ||||||
| UIBC (µg × dL−1) | ||||||||||||
| PLACEBO | 239.71 ± 86.10 | 259.76 ± 86.33 | 182.27 ± 49.26 b | 0.777 | 0.934 | 0.525 | 255.52 ± 112.27 | 249.84 ± 71.29 | 200.45 ± 69.98 | 0.631 | 0.909 | 0.054 |
| COLOSTRUM | 224.33 ± 55.50 | 228.22 ± 93.67 | 245.15 ± 85.92 | 218.29 ± 58.55 | 245.97 ± 52.77 | 261.89 ± 55.26 | ||||||
| TIBC (µg × dL−1) | ||||||||||||
| PLACEBO | 325.59 ± 59.78 | 374.75 ± 66.40 a | 297.77 ± 57.89 b | 0.614 | 0.658 | 0.254 | 332.98 ± 83.48 | 367.55 ± 112.86 | 317.85 ± 83.98 | 0.995 | 0.582 | 0.327 |
| COLOSTRUM | 337.68 ± 50.22 | 360.19 ± 116.69 | 346.94 ± 83.82 | 322.51 ± 55.26 | 332.25 ± 65.02 | 353.76 ± 51.53 | ||||||
| Variables | Before Supplementation | p Value | After Supplementation | p Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Exercise x ± SD | Post-Exercise x ± SD | Recovery x ± SD | Pre CON vs. SUPL | Post CON vs. SUPL | Recovery CON vs. SUPL | Pre-Exercise x ± SD | Post-Exercise x ± SD | Recovery x ± SD | Pre CON vs. SUPL | Post CON vs. SUPL | Recovery CON vs. SUPL | |
| RBC (109 × mL−1) | ||||||||||||
| PLACEBO | 4.36 ± 0.37 | 4.52 ± 0.37 | 4.19 ± 0.34 c | 0.929 | 0.936 | 0.094 | 4.63 ± 0.45 | 4.88 ± 0.45 | 4.31 ± 0.27 c | 0.998 | 0.019 | 0.873 |
| COLOSTRUM | 4.22 ± 0.25 | 4.35 ± 0.27 | 3.83 ± 0.37 b | 4.50 ± 0.45 | 4.35 ± 0.27 | 4.42 ± 0.21 | ||||||
| Hb (g × L−1) | ||||||||||||
| PLACEBO | 13.51 ± 1.20 | 13.89 ± 1.13 a | 12.62 ± 1.16 b | 0.962 | 0.994 | 0.540 | 13.93 ± 1.07 | 14.80 ± 1.75 | 12.95 ± 0.78 b | 0.845 | 0.579 | 0.999 |
| COLOSTRUM | 13.21 ± 0.78 | 13.54 ± 0.75 a | 11.95 ± 0.91 b | 13.27 ± 0.44 | 13.99 ± 0.73 | 12.80 ± 1.01 b | ||||||
| HTC (%) | ||||||||||||
| PLACEBO | 37.84 ± 3.05 | 39.13 ± 2.93 a | 35.82 ± 2.83 b | 0.823 | 0.788 | 0.023 | 40.95 ± 3.27 | 43.73 ± 4.86 | 37.94 ± 2.30 b | 0.772 | 0.376 | 0.999 |
| COLOSTRUM | 36.74 ± 2.01 | 37.6 ± 1.87 a | 32.76 ± 2.57 b | 38.89 ± 1.35 | 41.01 ± 1.87 a | 37.70 ± 2.67 b | ||||||
| Mioglobin (µg × L−1) | ||||||||||||
| PLACEBO | 17.11 ± 5.03 | 19.06 ± 3.04 a | 29.69 ± 3.85 b | 0.309 | 0.203 | 0.994 | 25.69 ± 8.31 | 27.27 ± 6.60 | 29.60 ± 7.25 | 0.964 | 0.671 | 0.716 |
| COLOSTRUM | 19.27 ± 3.50 | 21.21 ± 2.13 a | 29.41 ± 2.92 b | 26.51 ± 10.87 | 28.25 ± 8.43 | 29.82 ± 7.47 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cieślicka, M.; Ostapiuk-Karolczuk, J.; Buttar, H.S.; Dziewiecka, H.; Kasperska, A.; Skarpańska-Stejnborn, A. Effects of Long-Term Supplementation of Bovine Colostrum on Iron Homeostasis, Oxidative Stress, and Inflammation in Female Athletes: A Placebo-Controlled Clinical Trial. Nutrients 2023, 15, 186. https://doi.org/10.3390/nu15010186
Cieślicka M, Ostapiuk-Karolczuk J, Buttar HS, Dziewiecka H, Kasperska A, Skarpańska-Stejnborn A. Effects of Long-Term Supplementation of Bovine Colostrum on Iron Homeostasis, Oxidative Stress, and Inflammation in Female Athletes: A Placebo-Controlled Clinical Trial. Nutrients. 2023; 15(1):186. https://doi.org/10.3390/nu15010186
Chicago/Turabian StyleCieślicka, Mirosława, Joanna Ostapiuk-Karolczuk, Harpal S. Buttar, Hanna Dziewiecka, Anna Kasperska, and Anna Skarpańska-Stejnborn. 2023. "Effects of Long-Term Supplementation of Bovine Colostrum on Iron Homeostasis, Oxidative Stress, and Inflammation in Female Athletes: A Placebo-Controlled Clinical Trial" Nutrients 15, no. 1: 186. https://doi.org/10.3390/nu15010186