
Effect of Chia (Salvia hispanica L.) Associated with High-Fat Diet on the Intestinal Health of Wistar Rats
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Material
2.2. Animals and Diets
2.3. Fecal Moisture
2.4. Cecal Content pH
2.5. IgA Quantification
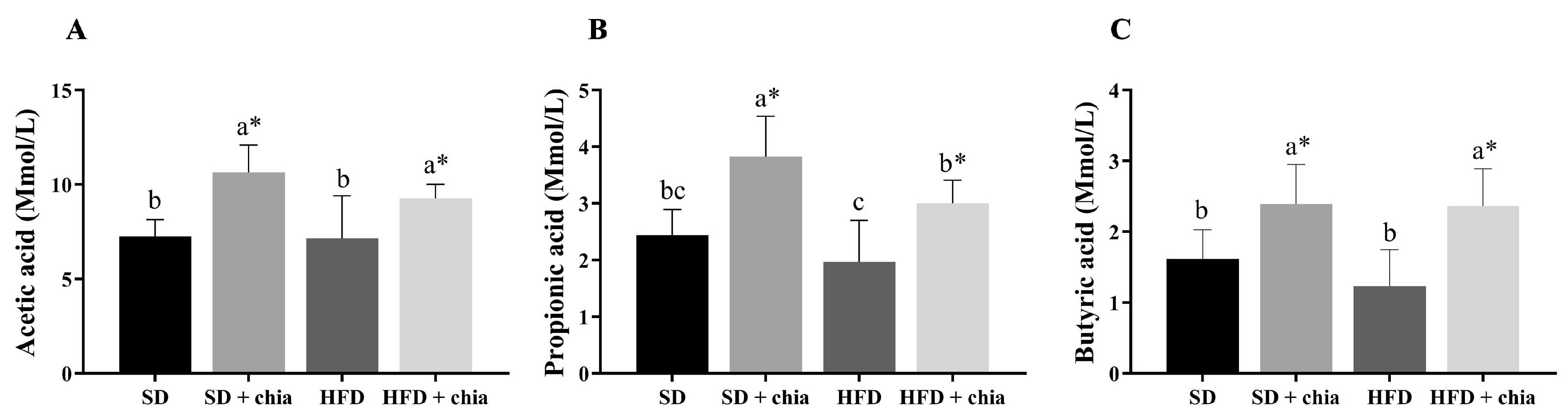
2.6. Short Chain Fatty Acids (SCFA) Content
2.7. Colon Histomorphometry
2.8. DNA Extraction and Sequencing
2.9. Determination of Gene Expression of Proteins Involved in Intestinal Health by Quantitative Reverse Transcriptase Polymerase Chain Reaction (RT-qPCR)
2.10. Statistical Analysis
3. Results
3.1. Data of Animals
3.2. Colonic Histomorphometry Characteristics
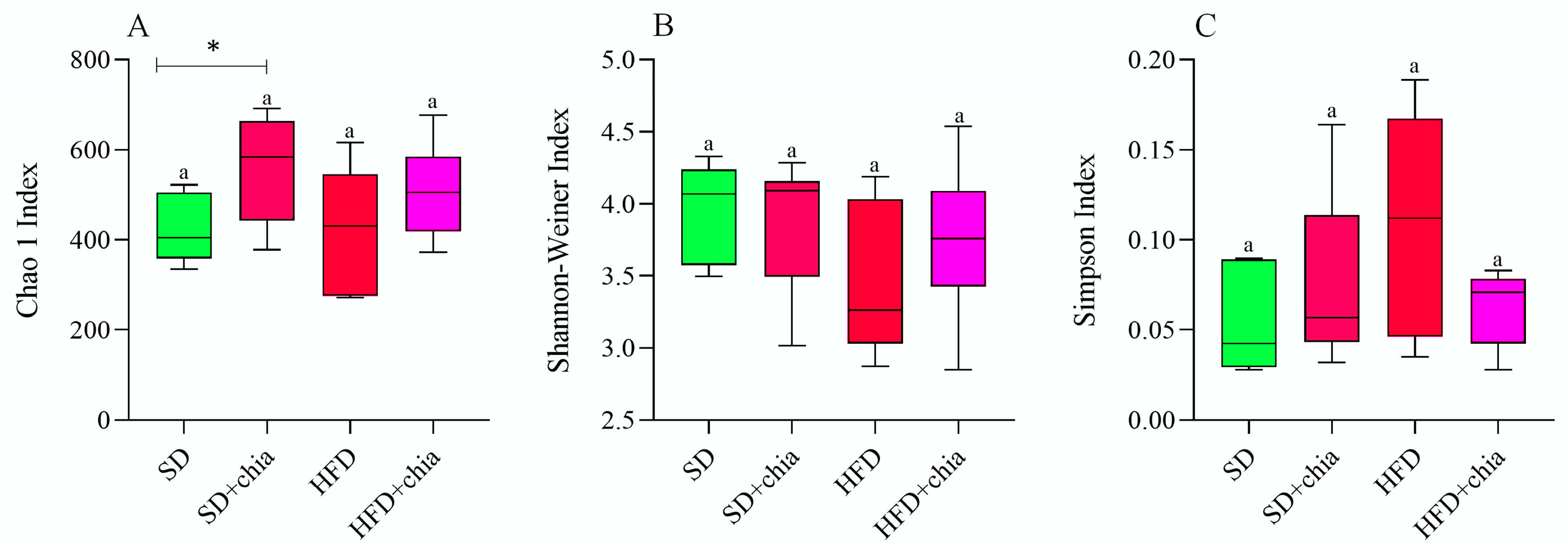
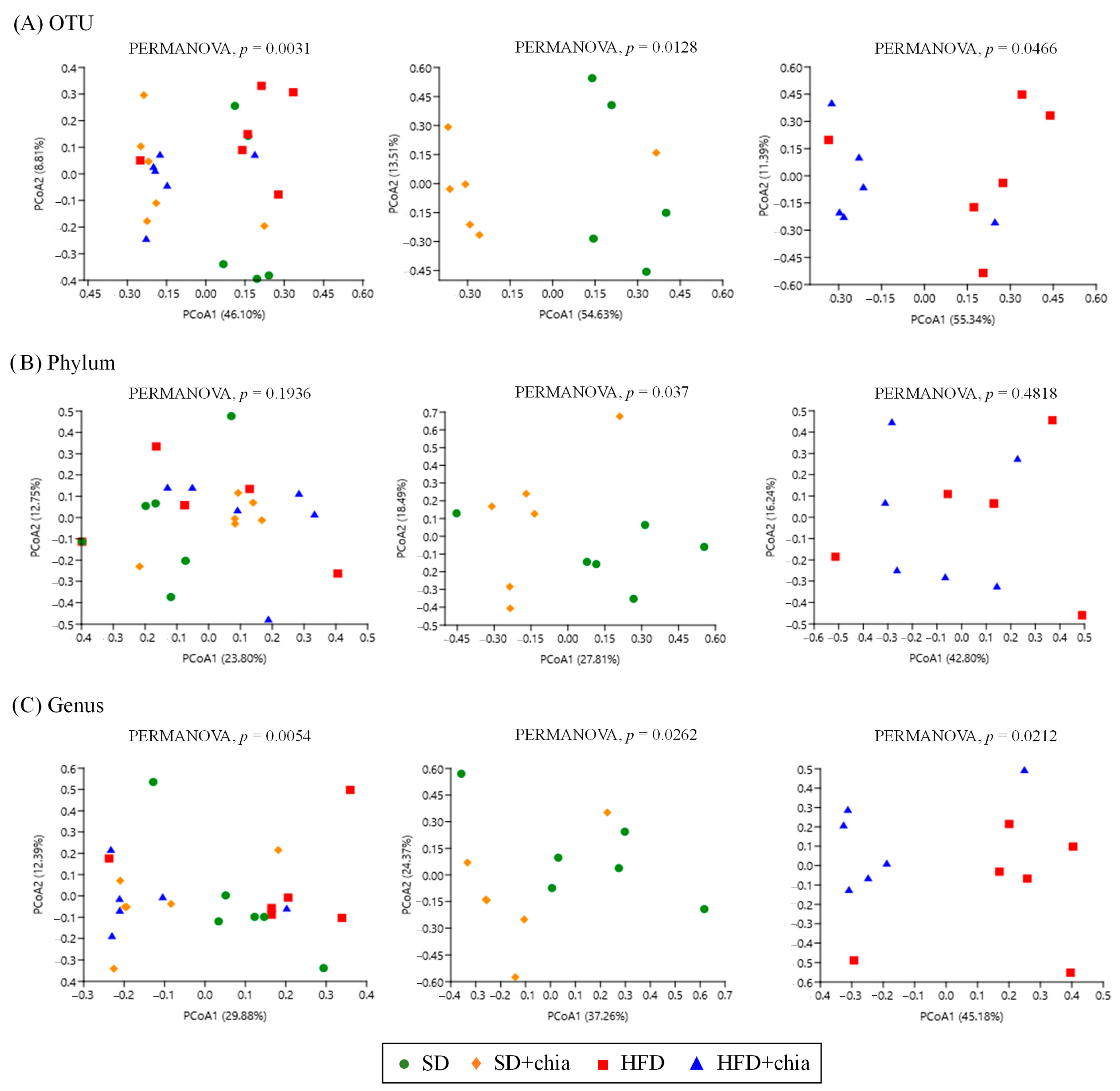
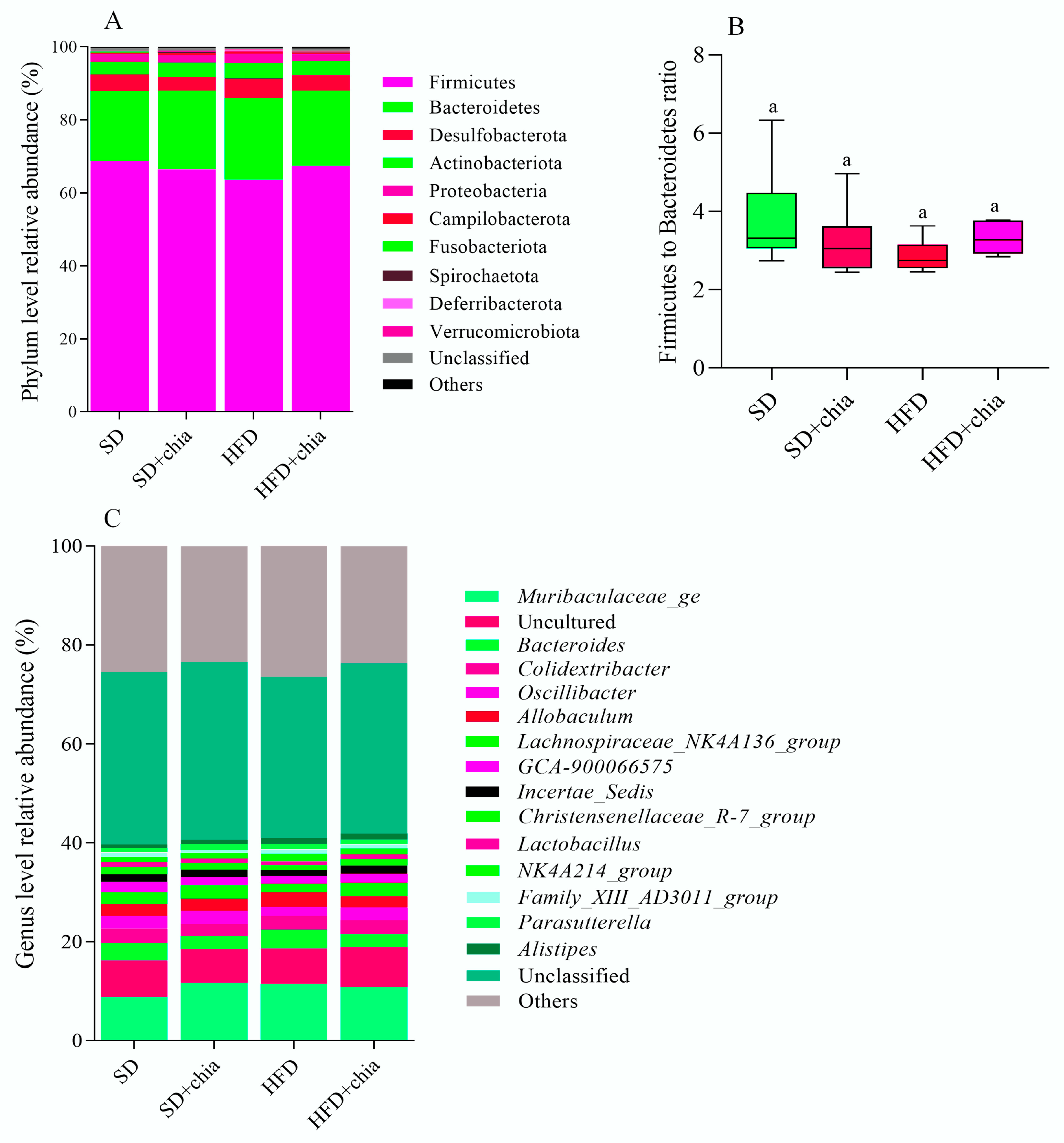
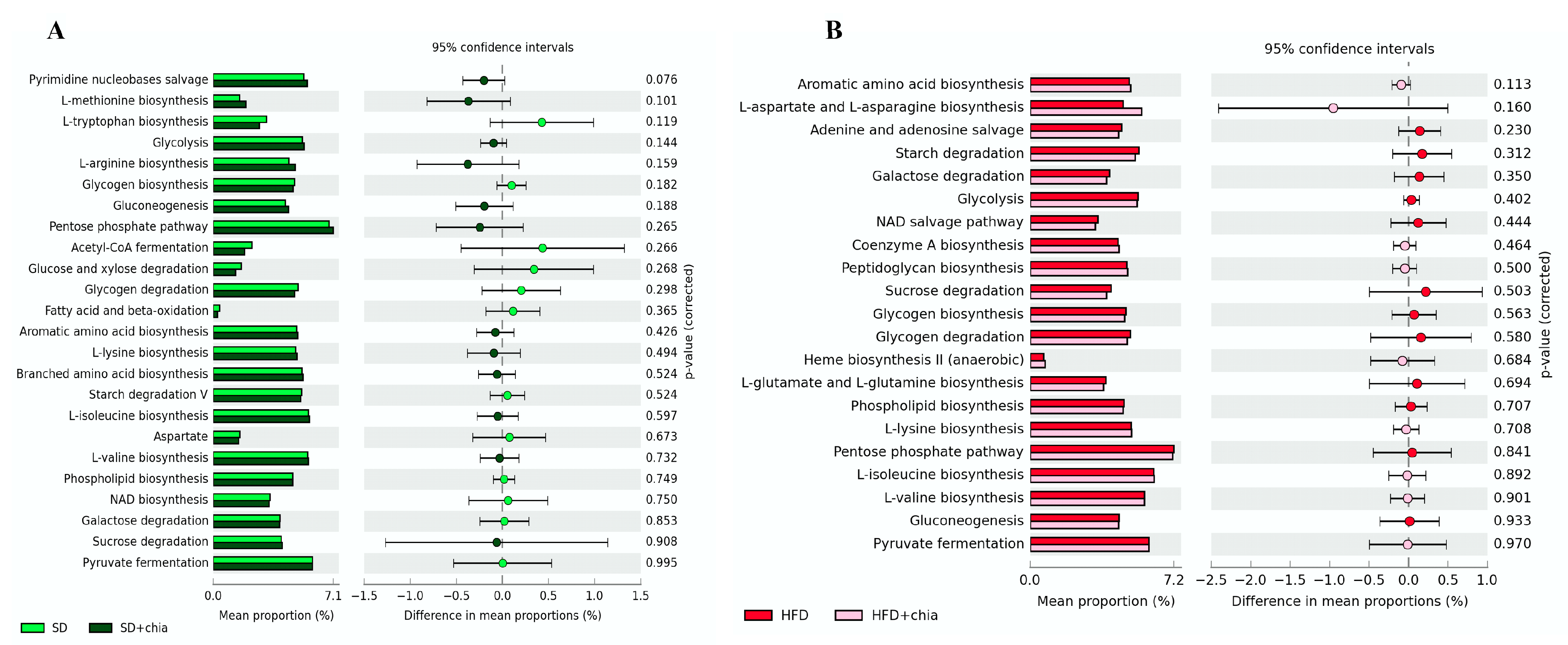
3.3. Intestinal Microbiota Analysis
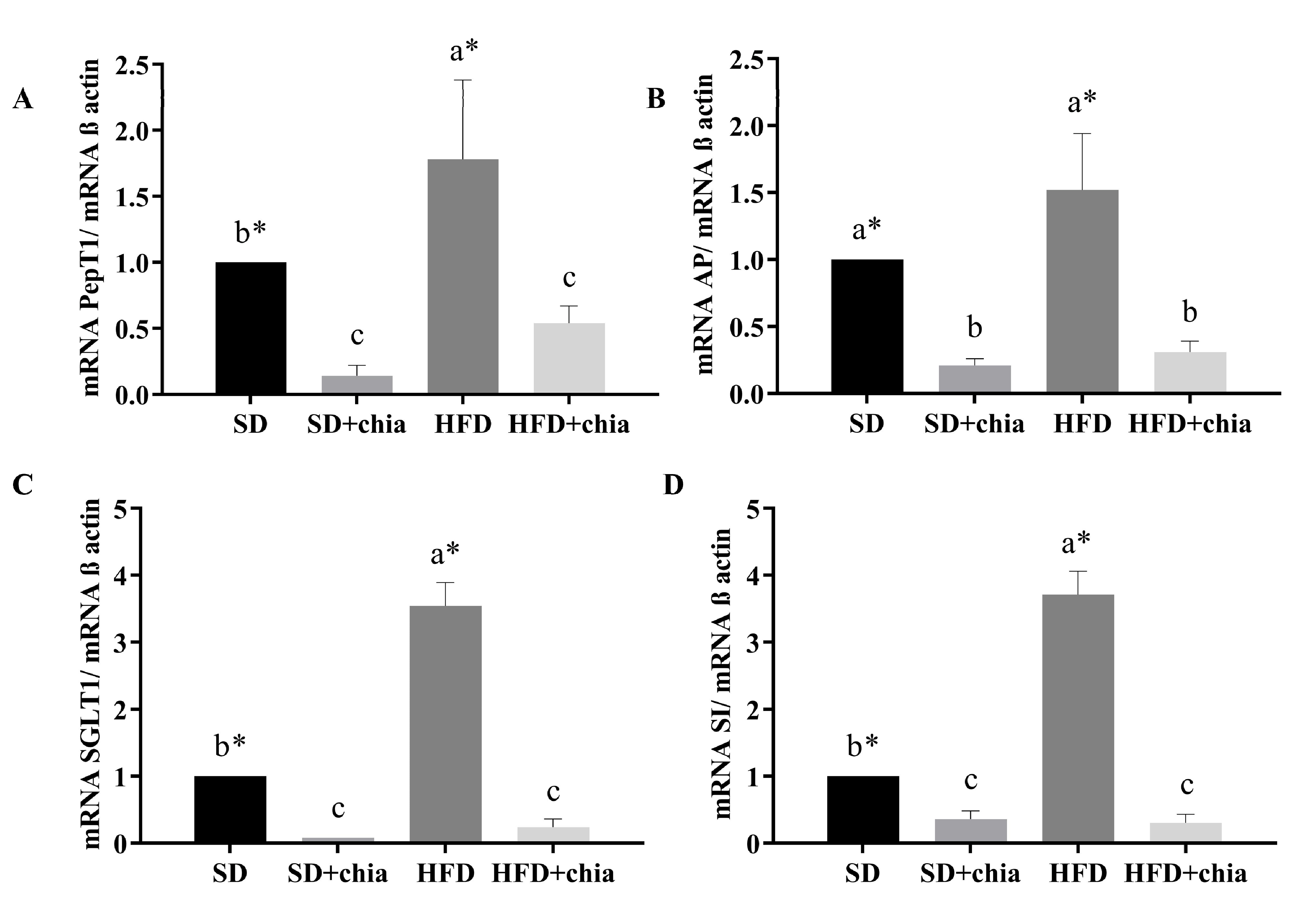
3.4. BBM Functional Proteins
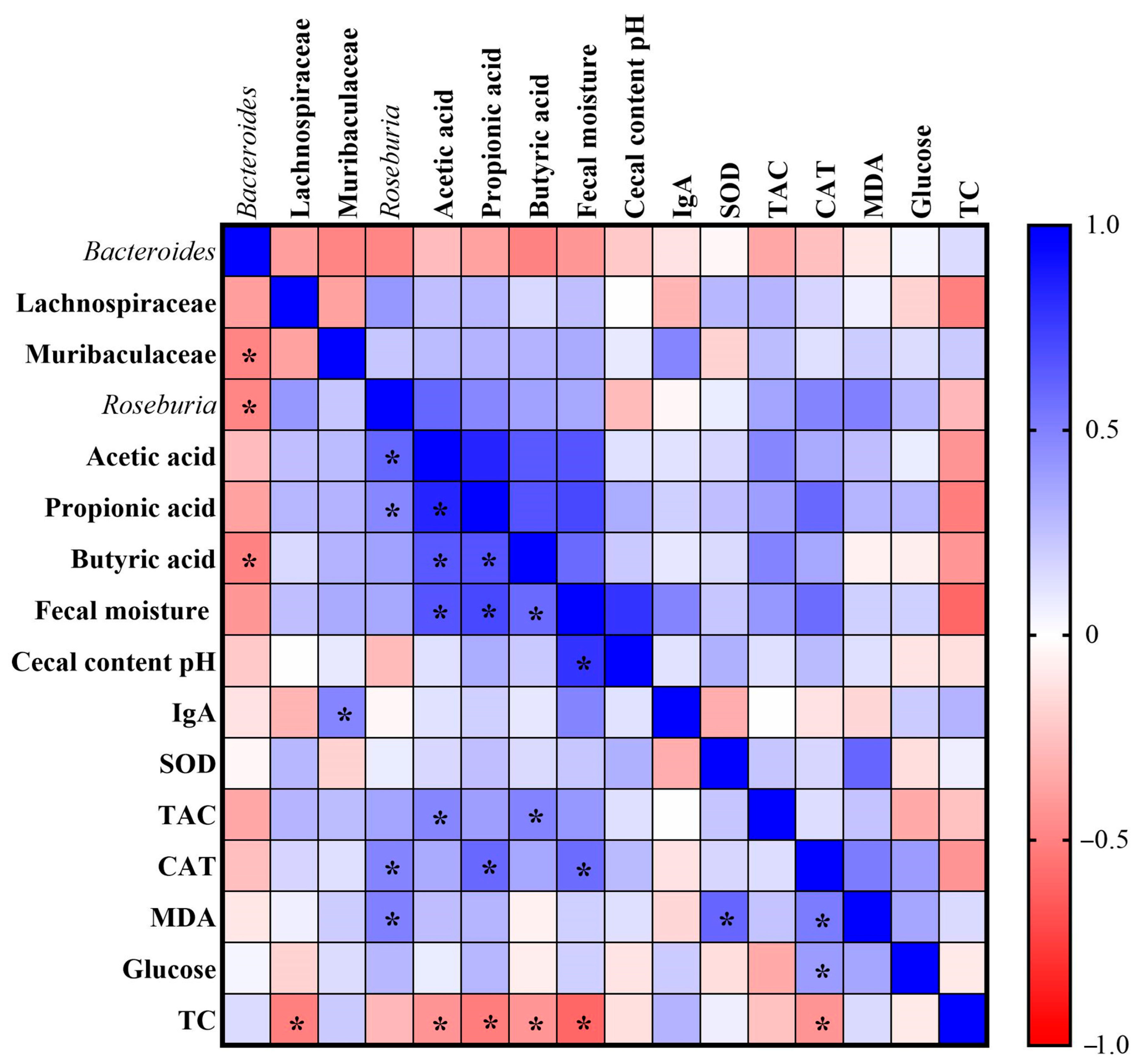
3.5. Pearson Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wali, J.A.; Jarzebska, N.; Raubenheimer, D.; Simpson, S.J.; Rodionov, R.N.; Sullivan, J.F.O. Cardio-Metabolic Effects of High-Fat Diets and Their Underlying Mechanisms—A Narrative Review. Nutrients 2020, 12, 1505. [Google Scholar] [CrossRef] [PubMed]
- Sferra, R.; Pompili, S.; Cappariello, A.; Gaudio, E.; Latella, G.; Vetuschi, A. Prolonged Chronic Consumption of a High Fat with Sucrose Diet Alters the Morphology of the Small Intestine. Int. J. Mol. Sci. 2021, 22, 7280. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P.J. Impact of High-Fat Diet on the Intestinal Microbiota and Small Intestinal Physiology before and after the Onset of Obesity. Biochimie 2017, 141, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, C.L.; Weir, T.L. The Gut Microbiota at the Intersection of Diet and Human Health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Van Der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-Chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, B.P.; Kolba, N.; Martino, H.S.D.; Hart, J.; Tako, E. Soluble Extracts from Chia Seed (Salvia Hispanica L.) Affect Brush Border Membrane Functionality, Morphology and Intestinal Bacterial Populations in Vivo (Gallus gallus). Nutrients 2019, 11, 2457. [Google Scholar] [CrossRef] [Green Version]
- Martino, H.S.D.; Kolba, N.; Tako, E. Yacon (Smallanthus Sonchifolius) Flour Soluble Extract Improve Intestinal Bacterial Populations, Brush Border Membrane Functionality and Morphology in Vivo (Gallus gallus). Food Res. Int. 2020, 137, 109705. [Google Scholar] [CrossRef]
- Melo, D.; Machado, T.B.; Oliveira, M.B.P.P. Chia Seeds: An Ancient Grain Trending in Modern Human Diets. Food Funct. 2019, 10, 3068–3089. [Google Scholar] [CrossRef]
- Enes, B.N.; Moreira, L.P.D.; Silva, B.P.; Grancieri, M.; Lúcio, H.G.; Venâncio, V.P.; Mertens-Talcott, S.U.; Rosa, C.O.B.; Martino, H.S.D. Chia Seed (Salvia Hispanica L.) Effects and Their Molecular Mechanisms on Unbalanced Diet Experimental Studies: A Systematic Review. J. Food Sci. 2020, 85, 226–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishima, M.D.V.; Ladeira, L.C.M.; da Silva, B.P.; Toledo, R.C.L.; de Oliveira, T.V.; Costa, N.M.B.; Martino, H.S.D. Cardioprotective Action of Chia (Salvia Hispanica L.) in Ovariectomized Rats Fed a High Fat Diet. Food Funct. 2021, 12, 3069–3082. [Google Scholar] [CrossRef]
- Da Silva, B.P.; Anunciação, P.C.; Matyelka, J.C.D.S.; Della Lucia, C.M.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Chemical Composition of Brazilian Chia Seeds Grown in Different Places. Food Chem. 2017, 221, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Grancieri, M.; Martino, H.S.D.; de Mejia, E.G. Protein Digests and Pure Peptides from Chia Seed Prevented Adipogenesis and Inflammation by Inhibiting Pparγ and Nf-Κb Pathways in 3t3l-1 Adipocytes. Nutrients 2021, 13, 176. [Google Scholar] [CrossRef]
- Abramson, J.H. WINPEPI Updated: Computer Programs for Epidemiologists, and Their Teaching Potential. Epidemiol. Perspect. Innov. 2011, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.S.; Teixeira, L.G.; Aguilar, E.C.; Matoso, R.O.; Soares, F.L.P.; Ferreira, A.V.M.; Alvarez-Leite, J.I. Differences in Adipose Tissue Inflammation and Oxidative Status in C57BL/6 and ApoE-/- Mice Fed High Fat Diet. Anim. Sci. J. 2012, 83, 549–555. [Google Scholar] [CrossRef]
- AOAC Official Methods of Analysis of AOAC International, 20th ed.; Association of Official Analytical Chemistry: Gaithersburg, MD, USA, 2016.
- Grancieri, M.; Costa, N.M.B.; Vaz Tostes, M.D.G.; de Oliveira, D.S.; Nunes, L.D.C.; Marcon, L.D.N.; Veridiano, T.A.; Viana, M.L. Yacon Flour (Smallanthus Sonchifolius) Attenuates Intestinal Morbidity in Rats with Colon Cancer. J. Funct. Foods 2017, 37, 666–675. [Google Scholar] [CrossRef]
- das Graças Vaz-Tostes, M.; Viana, M.L.; Grancieri, M.; dos Santos Luz, T.C.; de Paula, H.; Pedrosa, R.G.; Costa, N.M.B. Yacon Effects in Immune Response and Nutritional Status of Iron and Zinc in Preschool Children. Nutrition 2014, 30, 666–672. [Google Scholar] [CrossRef]
- Siegfried, V.R.; Ruckemmann, H.; Stumpf, G. Method for the Determination of Organic Acids in Silage by High Performance Liquid Chromatography. Landwirtsch. Forsch. 1984, 37, 298–304. [Google Scholar]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and Low Abundance of Classical Ruminal Bacterial Species in the Bovine Rumen Revealed by Relative Quantification Real-Time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Caicedo, H.H.; Hashimoto, D.A.; Caicedo, J.C.; Pentland, A.; Pisano, G.P. Overcoming Barriers to Early Disease Intervention. Nat. Biotechnol. 2020, 38, 669–673. [Google Scholar] [CrossRef]
- Da Silva, B.P.; Toledo, R.C.L.; Grancieri, M.; de Castro Moreira, M.E.; Medina, N.R.; Silva, R.R.; Costa, N.M.B.; Martino, H.S.D. Effects of Chia (Salvia Hispanica L.) on Calcium Bioavailability and Inflammation in Wistar Rats. Food Res. Int. 2019, 116, 592–599. [Google Scholar] [CrossRef]
- Grancieri, M.; Stampini, H.; Martino, D.; De Mejia, E.G. Chia Seed (Salvia Hispanica L.) as a Source of Proteins and Bioactive Peptides with Health Benefits : A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 480–499. [Google Scholar] [CrossRef] [Green Version]
- Grancieri, M.; Verediano, T.A.; Sant’Ana, C.T.; de Assis, A.; Toledo, R.L.; de Mejia, E.G.; Martino, H.S.D. Digested Protein from Chia Seed (Salvia Hispanica L) Prevents Obesity and Associated Inflammation of Adipose Tissue in Mice Fed a High-Fat Diet. Pharma Nutr. 2022, 21, 100298. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Hung, T.H.; Chiang, W.D.; Lin, W.T. Lipolysis-Stimulating Peptide from Soybean Protects Against High Fat Diet-Induced Apoptosis in Skeletal Muscles. J. Med. Food 2018, 21, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Li, R.; Zhang, T.; Li, M.; Mou, H. Study on the Ability of Partially Hydrolyzed Guar Gum to Modulate the Gut Microbiota and Relieve Constipation. J. Food Biochem. 2019, 43, e12715. [Google Scholar] [CrossRef] [PubMed]
- Mishima, M.D.V.; da Silva, B.P.; Gomes, M.J.C.; Toledo, R.C.L.; Pereira, C.E.R.; Costa, N.M.B.; Martino, H.S.D. Effect of Chia Flour Associated with High Fat Diet on Intestinal Health in Female Ovariectomized Wistar Rats. Eur. J. Nutr. 2022. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, B.P.; Matyelka, J.C.D.S.; Moreira, M.E.D.C.; Toledo, R.C.L.; Della Lucia, C.M.; Pinheiro-Sant’Ana, H.M.; Martino, H.S.D. A High Fat Diet Does Not Affect the Iron Bioavailability in Wistar Rats Fed with Chia and Increases Gene Expression of Iron Metabolism Proteins. Food Funct. 2016, 7, 4861–4868. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, A.; Cueva, C.; Laguna, L.; Moreno-Arribas, M.V.; Muñoz, L.A. Understanding the Impact of Chia Seed Mucilage on Human Gut Microbiota by Using the Dynamic Gastrointestinal Model Simgi®. J. Funct. Foods 2018, 50, 104–111. [Google Scholar] [CrossRef]
- Ghosh, S.; Pramanik, S. Structural Diversity, Functional Aspects and Future Therapeutic Applications of Human Gut Microbiome. Arch. Microbiol. 2021, 203, 5281–5308. [Google Scholar] [CrossRef]
- Hernández, M.A.G.; Canfora, E.E.; Jocken, J.W.E.; Blaak, E.E. The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11, 1943. [Google Scholar] [CrossRef] [Green Version]
- Patterson, J.K.; Lei, X.G.; Miller, D.D. The Pig as an Experimental Model for Elucidating the Mechanisms Governing Dietary Influence on Mineral Absorption. Exp. Biol. 2008, 233, 651–664. [Google Scholar] [CrossRef]
- Geva-zatorsky, N.; Sefik, E.; Kua, L.; Pasman, L.; Tan, T.G.; Ortiz-lopez, A.; Yanortsang, T.B.; Yang, L.; Jupp, R.; Mathis, D.; et al. Mining the Human Gut Microbiota for Immunomodulatory Organisms. Cell 2017, 168, 928–943. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Ohno, H. Reciprocal Regulation of IgA and the Gut Microbiota: A Key Mutualism in the Intestine. Int. Immunol. 2021, 33, 781–786. [Google Scholar] [CrossRef]
- Nakajima, A.; Sasaki, T.; Itoh, K.; Kitahara, T.; Takema, Y.; Hiramatsu, K.; Ishikawa, D. A Soluble Fiber Diet Increases Bacteroides Fragilis Group Abundance and Immunoglobulin A Production in the Gut. Appl. Environ. Microbiol. 2020, 86, e00405-20. [Google Scholar] [CrossRef]
- Covasa, M.; Stephens, R.W.; Toderean, R.; Cobuz, C. Intestinal Sensing by Gut Microbiota: Targeting Gut Peptides. Front. Endocrinol. 2019, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwma, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 12. [Google Scholar] [CrossRef]
- Siddiqui, M.T.; Cresci, G.A.M. The Immunomodulatory Functions of Butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and Cultivation Study of Muribaculaceae Reveals Novel Species, Host Preference, and Functional Potential of This yet Undescribed Family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.J.; Miller, R.A.; Ericsson, A.C.; Harrison, D.C.; Strong, R.; Schmidt, T.M. Changes in the Gut Microbiome and Fermentation Products Concurrent with Enhanced Longevity in Acarbose-Treated Mice. BMC Microbiol. 2019, 19, 130. [Google Scholar] [CrossRef] [Green Version]
- Barros de Pinho, R.; de Oliveira Silva, M.T.; Brenner, G.; Dié Alves, M.S.; Azevedo, V.; Dias Portela, R.; Borsuk, S. A Novel Approach for an Immunogen against Corynebacterium Pseudotuberculosis Infection: An Escherichia Coli Bacterin Expressing Phospholipase D. Microb. Pathog. 2021, 151, 104746. [Google Scholar] [CrossRef]
- da Silva, B.P.; Toledo, R.C.L.; Mishima, M.D.V.; Moreira, M.E.D.C.; Vasconcelos, C.M.; Pereira, C.E.R.; Favarato, L.S.C.; Costa, N.M.B.; Martino, H.S.D. Effects of Chia (Salvia Hispanica L.) on Oxidative Stress and Inflammation in Ovariectomized Adult Female Wistar Rats. Food Funct. 2019, 10, 4036–4045. [Google Scholar] [CrossRef]
- Enes, B.N.; Moreira, L.D.P.D.; Toledo, R.C.L.; Moraes, É.A.; Moreira, M.E.D.C.; Hermsdorff, H.H.M.; Noratto, G.; Mertens-Talcott, S.U.; Talcott, S.; Martino, H.S.D.; et al. Effect of Different Fractions of Chia (Salvia Hispanica L.) on Glucose Metabolism, in Vivo and in Vitro. J. Funct. Foods 2020, 71, 157–164. [Google Scholar] [CrossRef]
- Rosas-Pérez, A.M.; Honma, K.; Goda, T. Sustained Effects of Resistant Starch on the Expression of Genes Related to Carbohydrate Digestion/Absorption in the Small Intestine. Int. J. Food Sci. Nutr. 2020, 71, 572–580. [Google Scholar] [CrossRef] [PubMed]
- da Silva, B.P.; Martino, H.S.D.; Tako, E. Plant Origin Prebiotics Affect Duodenal Brush Border Membrane Functionality and Morphology, in Vivo (Gallus gallus). Food Funct. 2021, 12, 6157–6166. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Diets | ||||
|---|---|---|---|---|
| Ingredients (g/kg of Diet) | SD | SD+chia | HFD | HFD+chia |
| Albumin * | 217.90 | 117.60 | 217.92 | 117.60 |
| Chia flour | 0.00 | 416.80 | 0.00 | 416.80 |
| Dextrinized starch | 132.00 | 132.00 | 132.00 | 115.10 |
| Sucrose | 100.00 | 100.00 | 100.00 | 100.00 |
| Lard | 0.00 | 0.00 | 200.00 | 200.00 |
| Soybean oil (mL) | 134.20 | 0.00 | 134.20 | 0.00 |
| Microcrystalline cellulose | 139.20 | 0.00 | 139.20 | 0.00 |
| Mineral mix | 35.00 | 35.00 | 35.00 | 35.00 |
| Vitamin mix | 10.00 | 10.00 | 10.00 | 10.00 |
| L-cystine | 3.00 | 3.00 | 3.00 | 3.00 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 | 2.50 |
| Corn starch | 221.20 | 183.10 | 21.20 | 0.00 |
| Total calories (kcal) | 3700.42 b | 3624.75 b | 4700.42 a | 4624.11 a |
| Caloric density (kcal g−1) | 3.70 b | 3.62 b | 4.70 a | 4.62 a |
| Genes | Oligonucleotide (5′-3′) | |
|---|---|---|
| Forward | Reverse | |
| Beta-actin | TTCGTGCCGGTCCACACCC | GCTTTGCACATGCCGGAGCC |
| AP | CTCTCTCCTCAAACACATGAA | AGTTCAGGGCCTTCTCATATTC |
| SI | CCTCCAGAACACAATCCCTATAC | GGAGAGGTGAGATGGATTAGA |
| PEPT1 | CCTGGTCGTCTCATCATATT | TTCTTCTCATCCTCATCGAACTG |
| SGLT1 | CATCCAGTCCATCACATTAC | CAATCAGGAAGCCGAGAATCA |
| SD | SD+chia | HFD | HFD+chia | |
|---|---|---|---|---|
| Food consumption (g/day) | 14.74 ± 0.86 a | 15.46 ± 1.20 a | 10.86 ± 0.49 c | 12.29 ± 0.93 b* |
| Final weight (g) | 221.69 ± 15.25 b | 241.10 ± 15.65 a* | 208.76 ± 11.95 b | 240.11 ± 16.89 a* |
| Adiposity (% of body weight) | 2.32 ± 0.60 a | 2.66 ± 0.39 a | 2.69 ± 0.49 a | 2.96 ± 0.46 a |
| Cecum weight (g) | 4.20 ± 0.71 a | 4.76 ± 0.60 a | 4.25 ± 0.40 a | 4.06 ± 0.83 a |
| Cecal content pH | 7.23 ± 1.28 a | 7.33 ± 0.27 a | 6.70 ± 0.21 a | 7.13 ± 0.45 a |
| Fecal moisture (%) | 16.36 ± 3.86 c | 37.65 ± 5.37 a* | 14.36 ± 6.41 c | 28.01 ± 5.97 b* |
| IgA (ng) | 271.09 ± 95.70 b | 555.50 ± 193.67 a | 751.83 ± 278.88 a | 681.17 ± 155.91 a |
| SD | SD+chia | HFD | HFD+chia | |
|---|---|---|---|---|
| LML (μm) | 49.16 ± 12.48 a | 52.65 ± 26.90 a | 26.02 ± 11.36 b* | 22.68 ± 10.94 b |
| CML (μm) | 111.88 ± 35.84 b | 127.44 ± 70.86 a | 62.32 ± 24.37 c* | 50.01 ± 24.13 d |
| Crypt length (μm) | 180.53 ± 41.33 a* | 163.67 ± 44.52 b | 103.39 ± 13.97 d | 121.51 ± 30.96 c* |
| Crypt thickness (μm) | 45.57 ± 7.64 a* | 36.00 ± 13.53 b | 26.72 ± 5.67 c | 25.41 ± 6.08 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishima, M.D.V.; Da Silva, B.P.; Gomes, M.J.C.; Toledo, R.C.L.; Mantovani, H.C.; José, V.P.B.d.S.; Costa, N.M.B.; Tako, E.; Martino, H.S.D. Effect of Chia (Salvia hispanica L.) Associated with High-Fat Diet on the Intestinal Health of Wistar Rats. Nutrients 2022, 14, 4924. https://doi.org/10.3390/nu14224924
Mishima MDV, Da Silva BP, Gomes MJC, Toledo RCL, Mantovani HC, José VPBdS, Costa NMB, Tako E, Martino HSD. Effect of Chia (Salvia hispanica L.) Associated with High-Fat Diet on the Intestinal Health of Wistar Rats. Nutrients. 2022; 14(22):4924. https://doi.org/10.3390/nu14224924
Chicago/Turabian StyleMishima, Marcella Duarte Villas, Bárbara Pereira Da Silva, Mariana Juste Contin Gomes, Renata Celi Lopes Toledo, Hilário Cuquetto Mantovani, Vinícius Parzanini Brilhante de São José, Neuza Maria Brunoro Costa, Elad Tako, and Hércia Stampini Duarte Martino. 2022. "Effect of Chia (Salvia hispanica L.) Associated with High-Fat Diet on the Intestinal Health of Wistar Rats" Nutrients 14, no. 22: 4924. https://doi.org/10.3390/nu14224924