
Isatis tinctoria L. Leaf Extract Inhibits Replicative Senescence in Dermal Fibroblasts by Regulating mTOR-NF-κB-SASP Signaling

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Preparation and Extraction
2.2. Cell Culture
2.3. Lactate Dehydrogenase Assay
2.4. SA-β-Gal Staining Assay
2.5. Luminex Cytokine Assay
2.6. Immunofluorescence Staining
2.7. Quantitative Polymerase Chain Reaction (qRT-PCR)
2.8. Western Blots
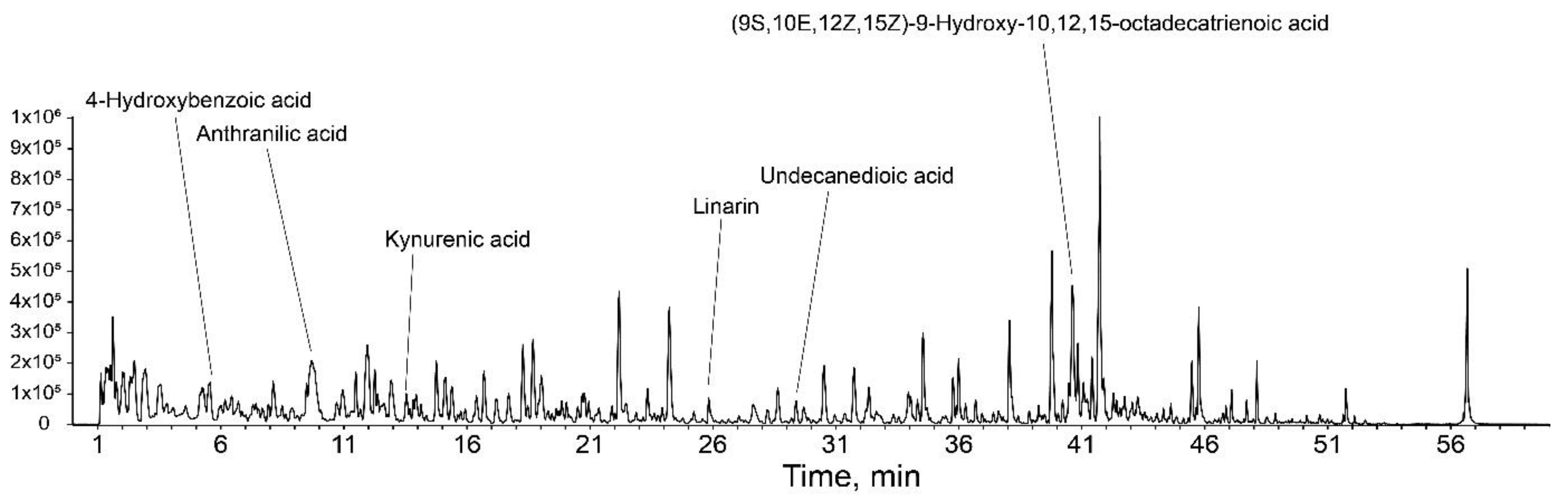
2.9. Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS)
2.10. Statistical Analysis
3. Results
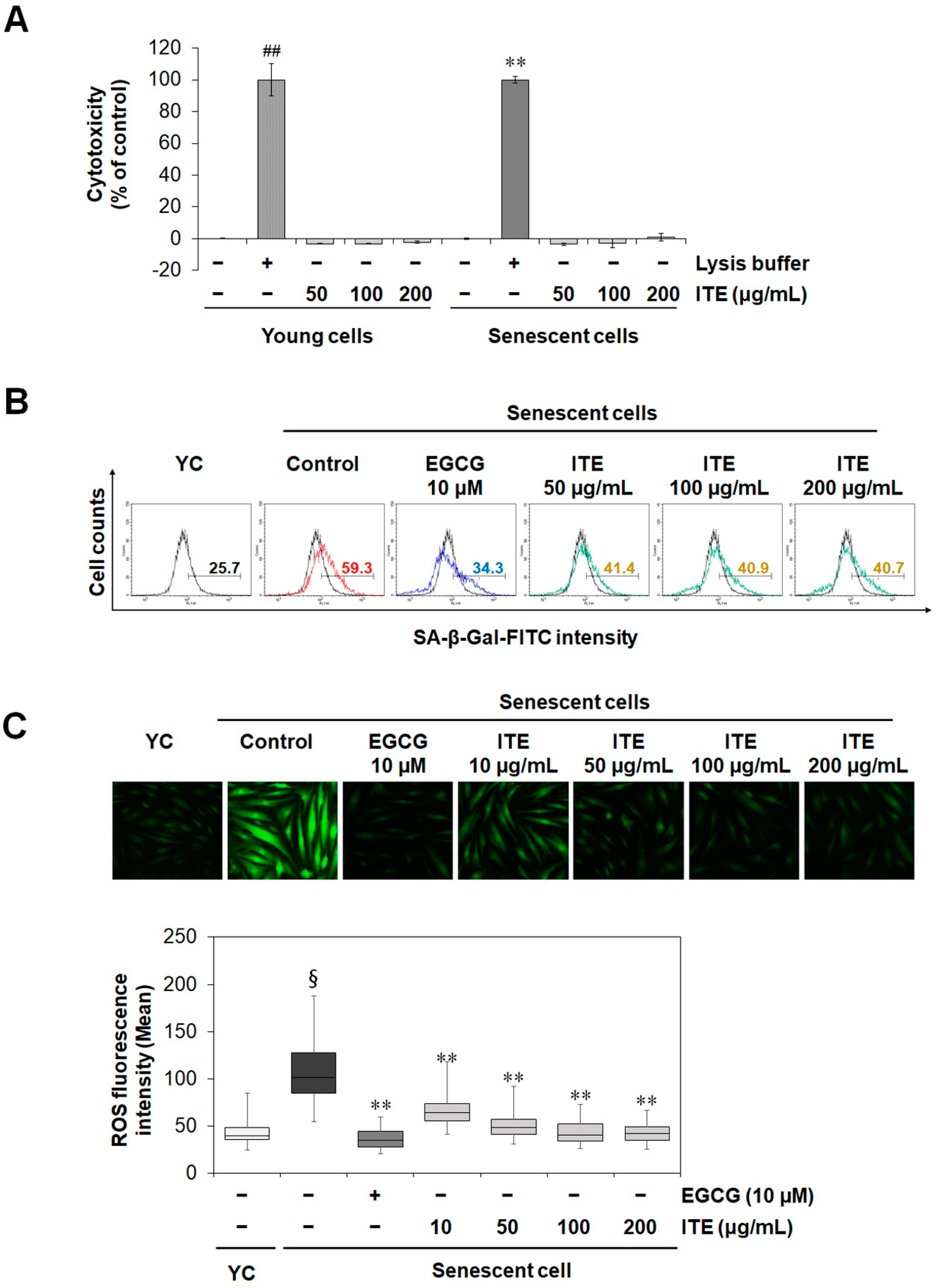
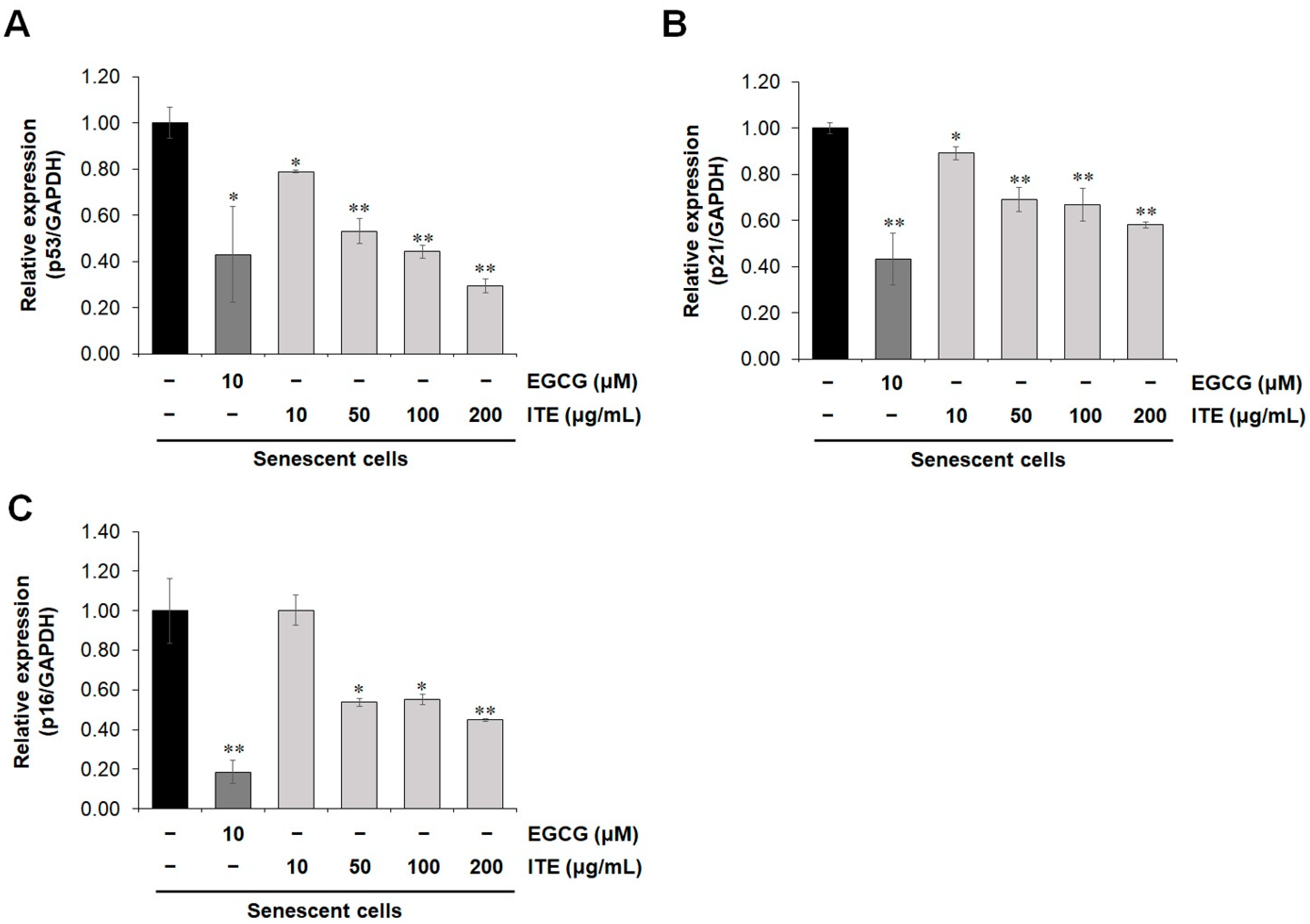
3.1. ITE Inhibits Replicative Senescence and Oxidative Stress
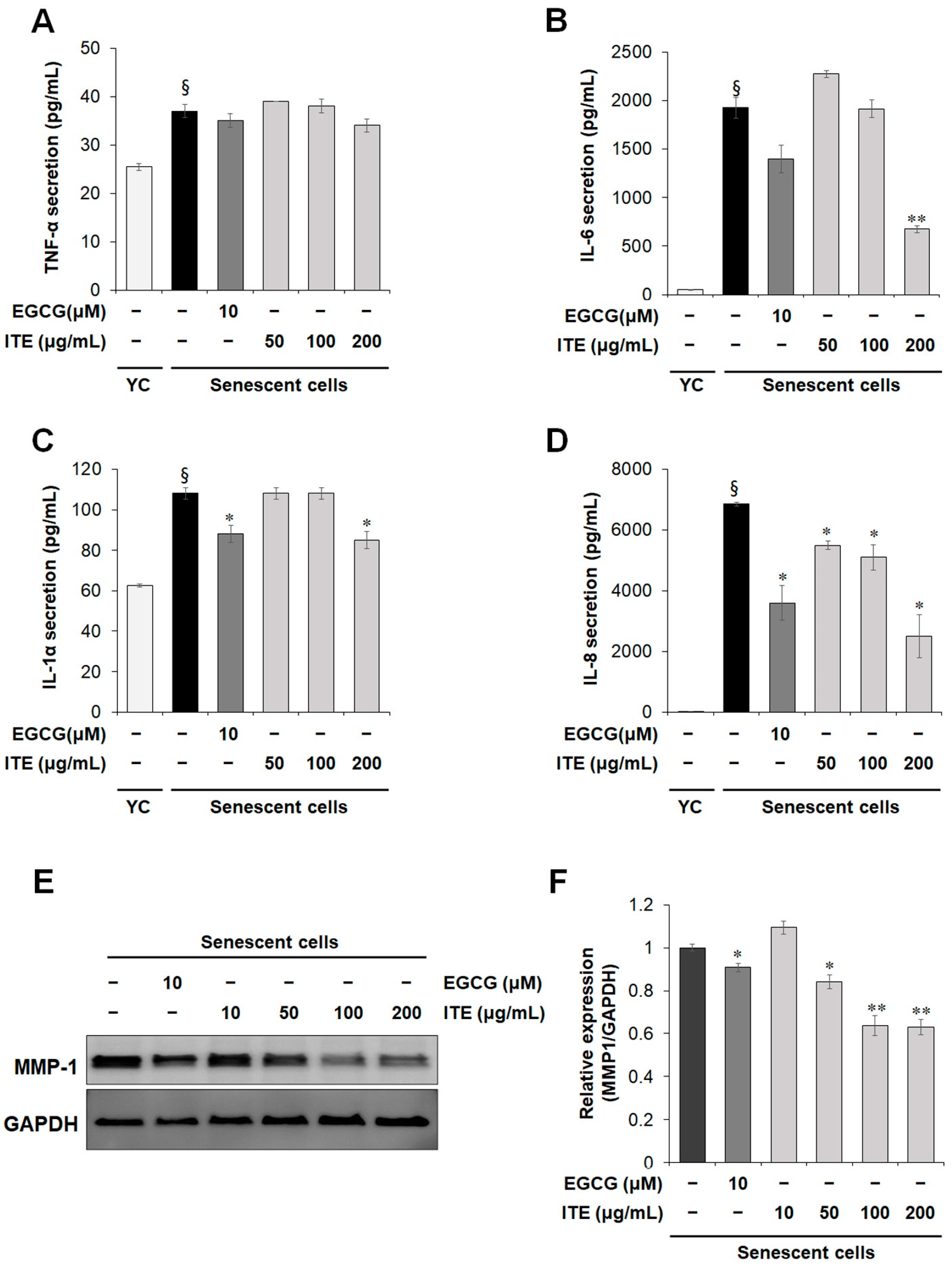
3.2. ITE Shows a Senomorphic Effect by Inhibiting Senescence-Associated Secretory Phenotype (SASP) Factor Production in Senescent Fibroblasts
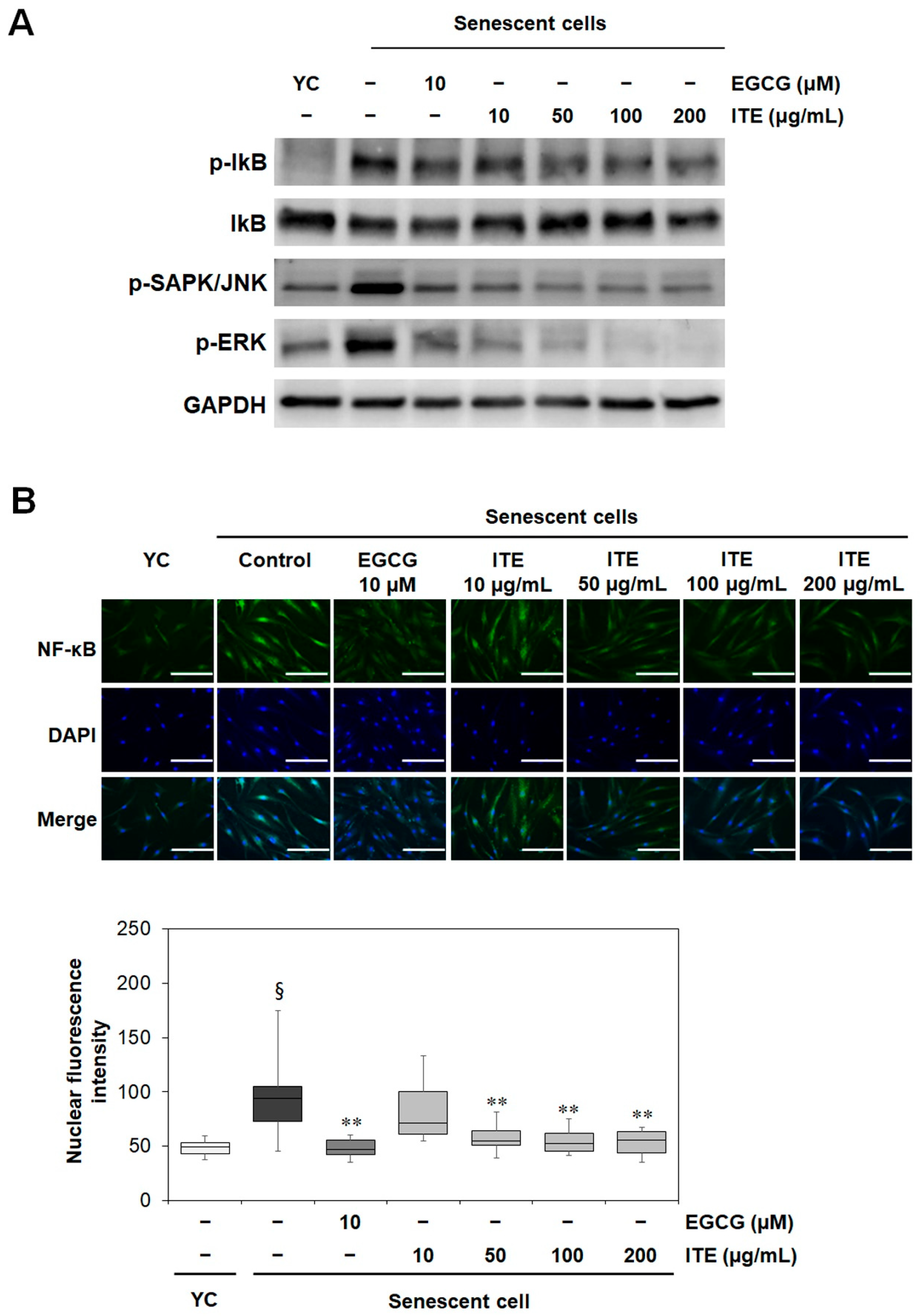
3.3. ITE Suppresses the Activation of MAPK (JNK and ERK), IκB, and NF-κB in RS-HDFs
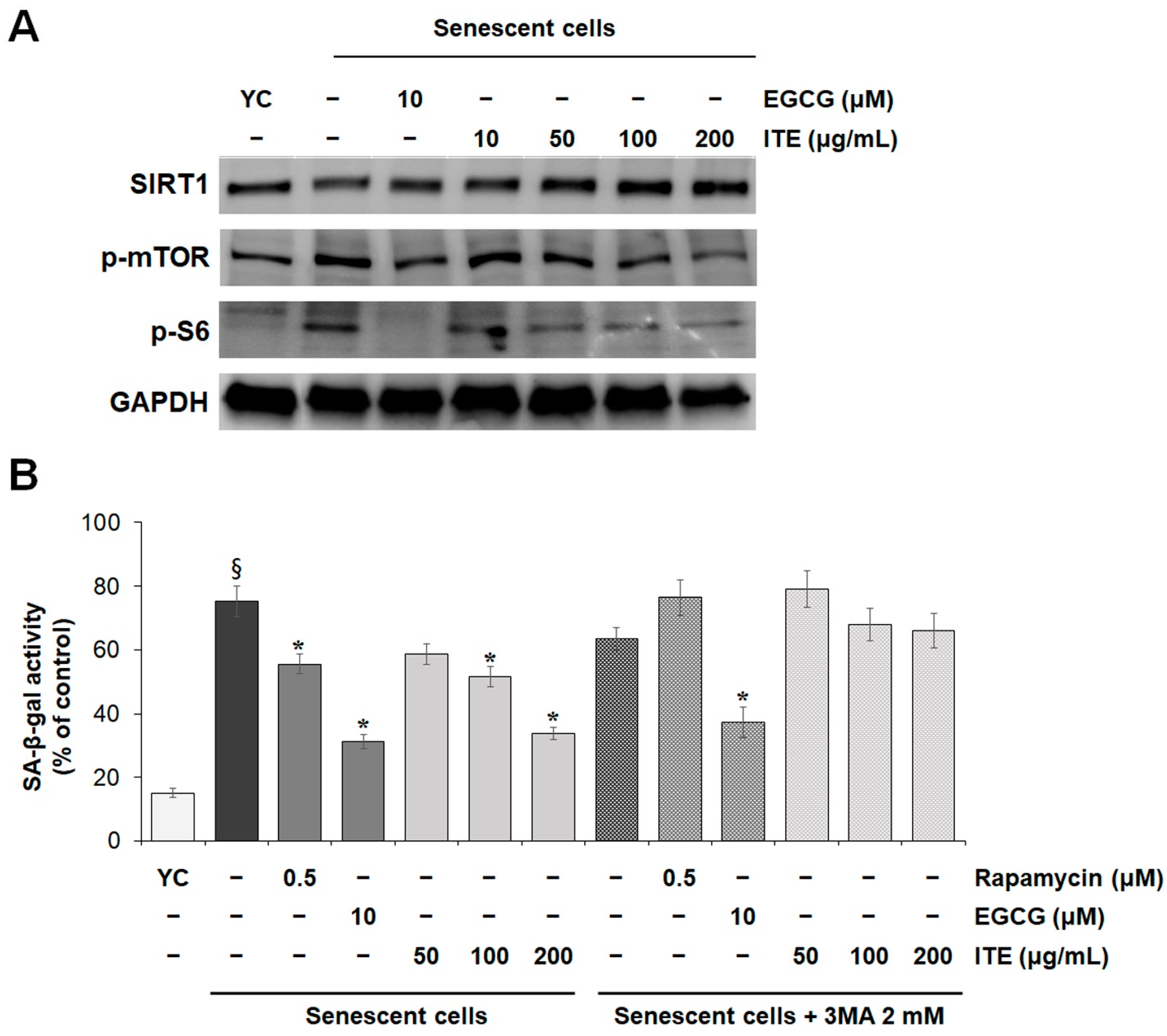
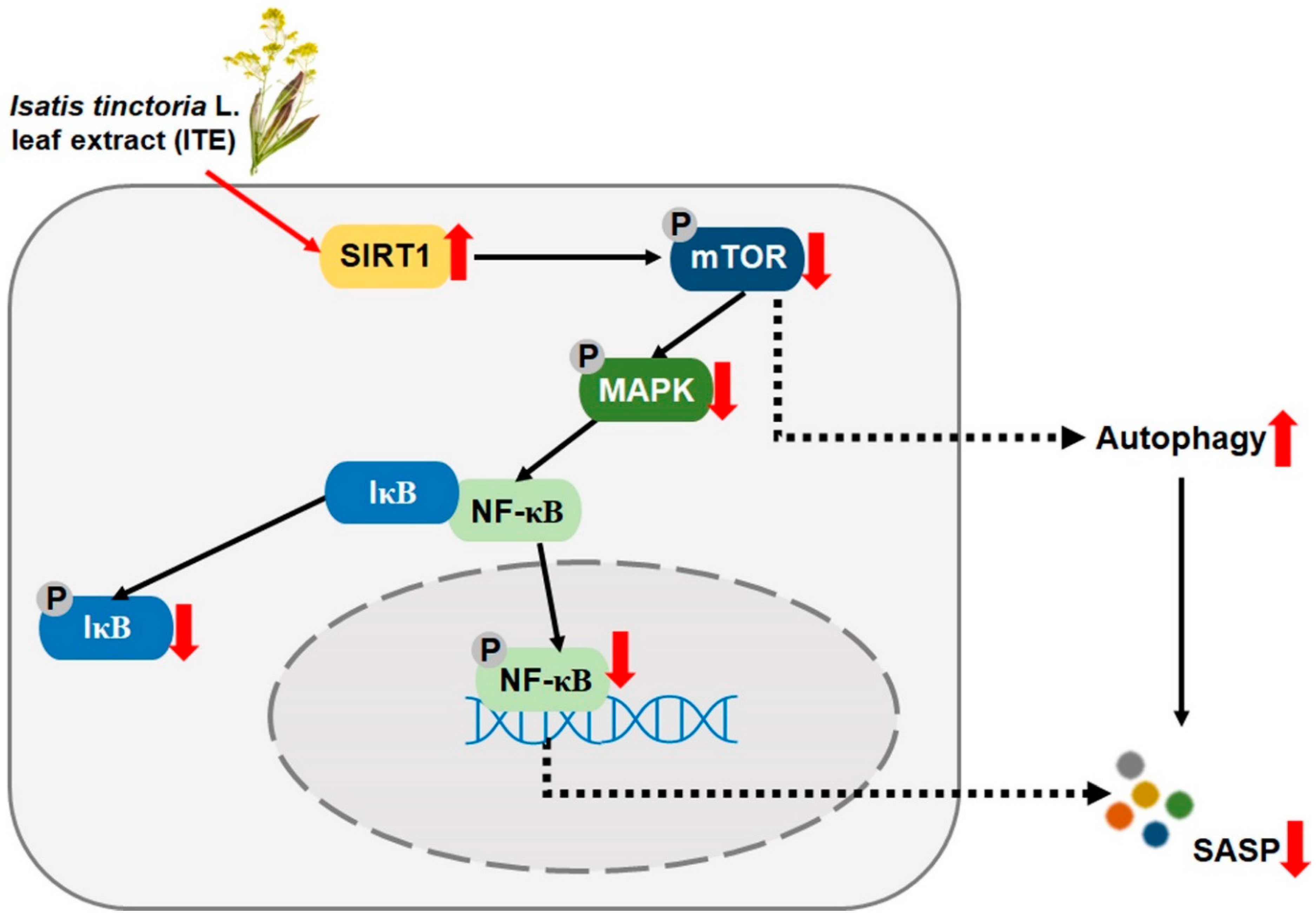
3.4. ITE Suppresses SASP Secretions and Cellular Senescence by Modulating SIRT1-mTOR-MAPK-NF-κB Signaling in RS-HDFs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, A.S.; Dreesen, O. Biomarkers of Cellular Senescence and Skin Aging. Front. Genet. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Herranz, N.; Gil, J. Mechanisms and Functions of Cellular Senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, T.; Zhang, X.; Zhang, R.; Lao, K.; Mi, Y.; Gou, X. AMPK Alleviates Oxidative Stress-induced Premature Senescence via Inhibition of NF-ΚB/STAT3 Axis-Mediated Positive Feedback Loop. Mech. Ageing Dev. 2020, 191, 111347. [Google Scholar] [CrossRef] [PubMed]
- Lagoumtzi, S.M.; Chondrogianni, N. Senolytics and Senomorphics: Natural and Synthetic Therapeutics in the Treatment of Aging and Chronic Diseases. Free Radic. Biol. Med. 2021, 171, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.S.; Lee, D.Y.; Kim, H.S.; Park, S.C.; Park, J.T.; Kim, H.S.; Oh, W.K.; Cho, K.A. Identification of a Novel Senomorphic Agent, Avenanthramide C, via the Suppression of the Senescence-Associated Secretory Phenotype. Mech. Ageing Dev. 2020, 192, 111355. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Lin, S.; Copland, D.A.; Dick, A.D.; Liu, J. Cellular Senescence in the Aging Retina and Developments of Senotherapies for Age-Related Macular Degeneration. J. Neuroinflammation 2021, 18, 32. [Google Scholar] [CrossRef]
- Kucheryavenko, O.; Nelson, G.; von Zglinicki, T.; Korolchuk, V.I.; Carroll, B. The MTORC1-Autophagy Pathway Is a Target for Senescent Cell Elimination. Biogerontology 2019, 20, 331–335. [Google Scholar] [CrossRef] [Green Version]
- European Directorate for the Quality of Medicines. Isatis root (Isatidis radix). In European Pharmacopoeia, 7th ed.; Suppl. 7.3, European Directorate for the Quality of Medicines: Strasburg, Germany, 2011; pp. 3866–3867. [Google Scholar]
- Speranza, J.; Miceli, N.; Taviano, M.F.; Ragusa, S.; Kwiecień, I.; Szopa, A.; Ekiert, H. Isatis tinctoria L. (Woad): A Review of Its Botany, Ethnobotanical Uses, Phytochemistry, Biological Activities, and Biotechnological Studies. Plants 2020, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Gao, D.; Jeong, W.S.; Kim, C.T.; Cho, C.W.; Kim, H.M.; Kang, J.S. Anti-Wrinkle Effect of Isatis indigotica Leaf Extract: Evaluation of Antioxidant, Anti-Inflammation, and Clinical Activity. Antioxidants 2021, 10, 1339. [Google Scholar] [CrossRef]
- Heinemann, C.; Schliemann-Willers, S.; Oberthür, C.; Hamburger, M.; Elsner, P. Prevention of experimentally induced irritant contact dermatitis by extracts of Isatis tinctoria compared to pure tryptanthrin and its impact on UVB-induced erythema. Planta Med. 2004, 70, 385–390. [Google Scholar]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot095497. [Google Scholar] [CrossRef] [PubMed]
- Hütter, E.; Unterluggauer, H.; Überall, F.; Schramek, H.; Jansen-Dürr, P. Replicative Senescence of Human Fibroblasts: The Role of Ras-Dependent Signaling and Oxidative Stress. Exp. Gerontol. 2002, 37, 1165–1174. [Google Scholar] [CrossRef]
- Giovannini, L.; Bianchi, S. Role of Nutraceutical SIRT1 Modulators in AMPK and MTOR Pathway: Evidence of a Synergistic Effect. Nutrition 2017, 34, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Young, M.C.; Seung, B.L.; Hyung, J.K.; Seon, H.P.; Jung, J.K.; Jin, S.C.; Young, D.Y. Replicative Senescence Induced by Romo1-Derived Reactive Oxygen Species. J. Biol. Chem. 2008, 283, 33763–33771. [Google Scholar]
- Schosserer, M.; Grillari, J.; Breitenbach, M. The Dual Role of Cellular Senescence in Developing Tumors and Their Response to Cancer Therapy. Front. Oncol. 2017, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Gasparrini, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Afrin, S.; Cianciosi, D.; Reboredo-Rodriguez, P.; Varela-Lopez, A.; Zhang, J.J.; Quiles, J.L.; Mezzetti, B.; et al. Strawberry Extracts Efficiently Counteract Inflammatory Stress Induced by the Endotoxin Lipopolysaccharide in Human Dermal Fibroblast. Food Chem. Toxicol. 2018, 114, 128–140. [Google Scholar] [CrossRef]
- Zhu, Y.; Tchkonia, T.; Fuhrmann-Stroissnigg, H.; Dai, H.M.; Ling, Y.Y.; Stout, M.B.; Pirtskhalava, T.; Giorgadze, N.; Johnson, K.O.; Giles, C.B.; et al. Identification of a Novel Senolytic Agent, Navitoclax, Targeting the Bcl-2 Family of Anti-Apoptotic Factors. Aging Cell 2016, 15, 428–435. [Google Scholar] [CrossRef]
- Bahrami, B.F.; Ataie-Kachoie, P.; Pourgholami, M.H.; Morris, D.L. P70 Ribosomal Protein S6 Kinase (Rps6kb1): An Update. J. Clin. Pathol. 2014, 67, 1019–1025. [Google Scholar] [CrossRef]
- Liu, Z.; Gan, L.; Liu, G.; Chen, Y.; Wu, T.; Feng, F.; Sun, C. Sirt1 Decreased Adipose Inflammation by Interacting with Akt2 and Inhibiting MTOR/S6K1 Pathway in Mice. J. Lipid Res. 2016, 57, 1373–1381. [Google Scholar] [CrossRef] [Green Version]
- Sung, J.Y.; Kim, S.G.; Kim, J.R.; Choi, H.C. SIRT1 Suppresses Cellular Senescence and Inflammatory Cytokine Release in Human Dermal Fibroblasts by Promoting the Deacetylation of NF-ΚB and Activating Autophagy. Exp. Gerontol. 2021, 150, 111394. [Google Scholar] [CrossRef]
- Birch, J.; Gil, J. Senescence and the SASP: Many Therapeutic Avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- von Kobbe, C. Targeting Senescent Cells: Approaches, Opportunities, Challenges. Aging 2019, 1, 12844–12861. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of Aging: An Autophagic Perspective. Front. Endocrinol. 2019, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Recio, M.C.; Cerdá-Nicolás, M.; Hamburger, M.; Ríos, J.L. Anti-Arthritic Activity of a Lipophilic Woad (Isatis tinctoria) Extract. Planta Med. 2006, 72, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Luo, A.; Lu, X.; Liu, M.; Wang, H.; Song, H.; Wei, C.; Wang, Y.; Duan, X. P-Hydroxybenzoic Acid Alleviates Inflammatory Responses and Intestinal Mucosal Damage in DSS-Induced Colitis by Activating ERβ Signaling. J. Funct. Foods 2021, 87, 104835. [Google Scholar] [CrossRef]
- Han, X.; Wu, Y.C.; Meng, M.; Sun, Q.S.; Gao, S.M.; Sun, H. Linarin Prevents LPS-Induced Acute Lung Injury by Suppressing Oxidative Stress and Inflammation via Inhibition of TXNIP/NLRP3 and NF-ΚB Pathways. Int. J. Mol. Med. 2018, 42, 1460–1472. [Google Scholar] [CrossRef] [Green Version]
- Francisco-Marquez, M.; Aguilar-Fernández, M.; Galano, A. Anthranilic Acid as a Secondary Antioxidant: Implications to the Inhibition of ·OH Production and the Associated Oxidative Stress. Comput. Theor. Chem. 2016, 1077, 18–24. [Google Scholar] [CrossRef]
- Lugo-Huitrón, R.; Blanco-Ayala, T.; Ugalde-Muñiz, P.; Carrillo-Mora, P.; Pedraza-Chaverrí, J.; Silva-Adaya, D.; Maldonado, P.D.; Torres, I.; Pinzón, E.; Ortiz-Islas, E.; et al. On the Antioxidant Properties of Kynurenic Acid: Free Radical Scavenging Activity and Inhibition of Oxidative Stress. Neurotoxicol. Teratol. 2011, 33, 538–547. [Google Scholar] [CrossRef]
- Lamming, D.W.; Ye, L.; Sabatini, D.M.; Baur, J.A. Rapalogs and MTOR Inhibitors as Anti-Aging Therapeutics. J. Clin. Investig. 2013, 123, 980–989. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.T.; Tan, H.L.; Shui, G.; Bauvy, C.; Huang, Q.; Wenk, M.R.; Ong, C.N.; Codogno, P.; Shen, H.M. Dual Role of 3-Methyladenine in Modulation of Autophagy via Different Temporal Patterns of Inhibition on Class I and III Phosphoinositide 3-Kinase. J. Biol. Chem. 2010, 285, 10850–10861. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bi, Y.; Chen, X.; Li, C.; Li, Y.; Zhang, Z.; Wang, J.; Lu, Y.; Yu, Q.; Su, H.; et al. Histone Deacetylase SIRT1 Negatively Regulates the Differentiation of Interleukin-9-Producing CD4+ T Cells. Immunity 2016, 44, 1337–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| p16INK4A | CTCGTGCTGATGCTACTGAGGA | GGTCGGCGCAGTTGGGCTCC |
| p21CDKN1A | AGGTGGACCTGGAGACTCTCAG | TCCTCTTGGAGAAGATCAGCCG |
| p53 | CCTCAGCATCTTATCCGAGTGG | TGGATGGTGGTACAGTCAGAGC |
| GAPDH | CATCAAGAAGGTGGTGAAGCAGG | AGTGGTCGTTGAGGGCAATGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, J.; Shin, S.; Ji, H.; Ryu, D.; Cho, E.; Kim, Y.; Kim, J.; Park, D.; Jung, E. Isatis tinctoria L. Leaf Extract Inhibits Replicative Senescence in Dermal Fibroblasts by Regulating mTOR-NF-κB-SASP Signaling. Nutrients 2022, 14, 1979. https://doi.org/10.3390/nu14091979
Woo J, Shin S, Ji H, Ryu D, Cho E, Kim Y, Kim J, Park D, Jung E. Isatis tinctoria L. Leaf Extract Inhibits Replicative Senescence in Dermal Fibroblasts by Regulating mTOR-NF-κB-SASP Signaling. Nutrients. 2022; 14(9):1979. https://doi.org/10.3390/nu14091979
Chicago/Turabian StyleWoo, Jieun, Seoungwoo Shin, Hyanggi Ji, Dehun Ryu, Eunae Cho, Youngseok Kim, Junoh Kim, Deokhoon Park, and Eunsun Jung. 2022. "Isatis tinctoria L. Leaf Extract Inhibits Replicative Senescence in Dermal Fibroblasts by Regulating mTOR-NF-κB-SASP Signaling" Nutrients 14, no. 9: 1979. https://doi.org/10.3390/nu14091979