
Therapeutic Effects of a Novel Form of Biotin on Propionic Acid-Induced Autistic Features in Rats

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rats, Study Protocol
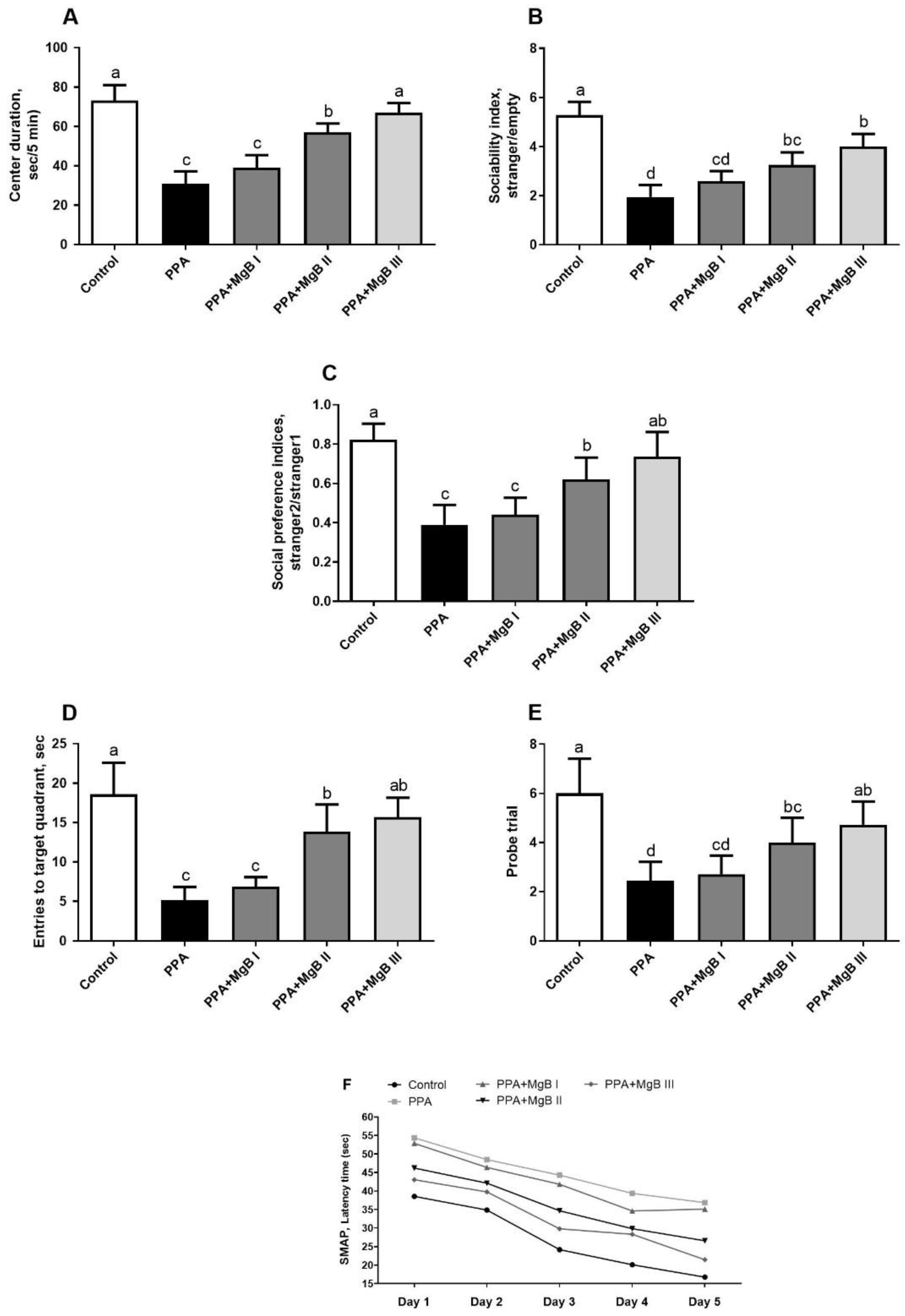
2.2. Behavioral Tests and Social Interaction Test
2.3. Morris Water Maze Task
2.4. Laboratory Analyses
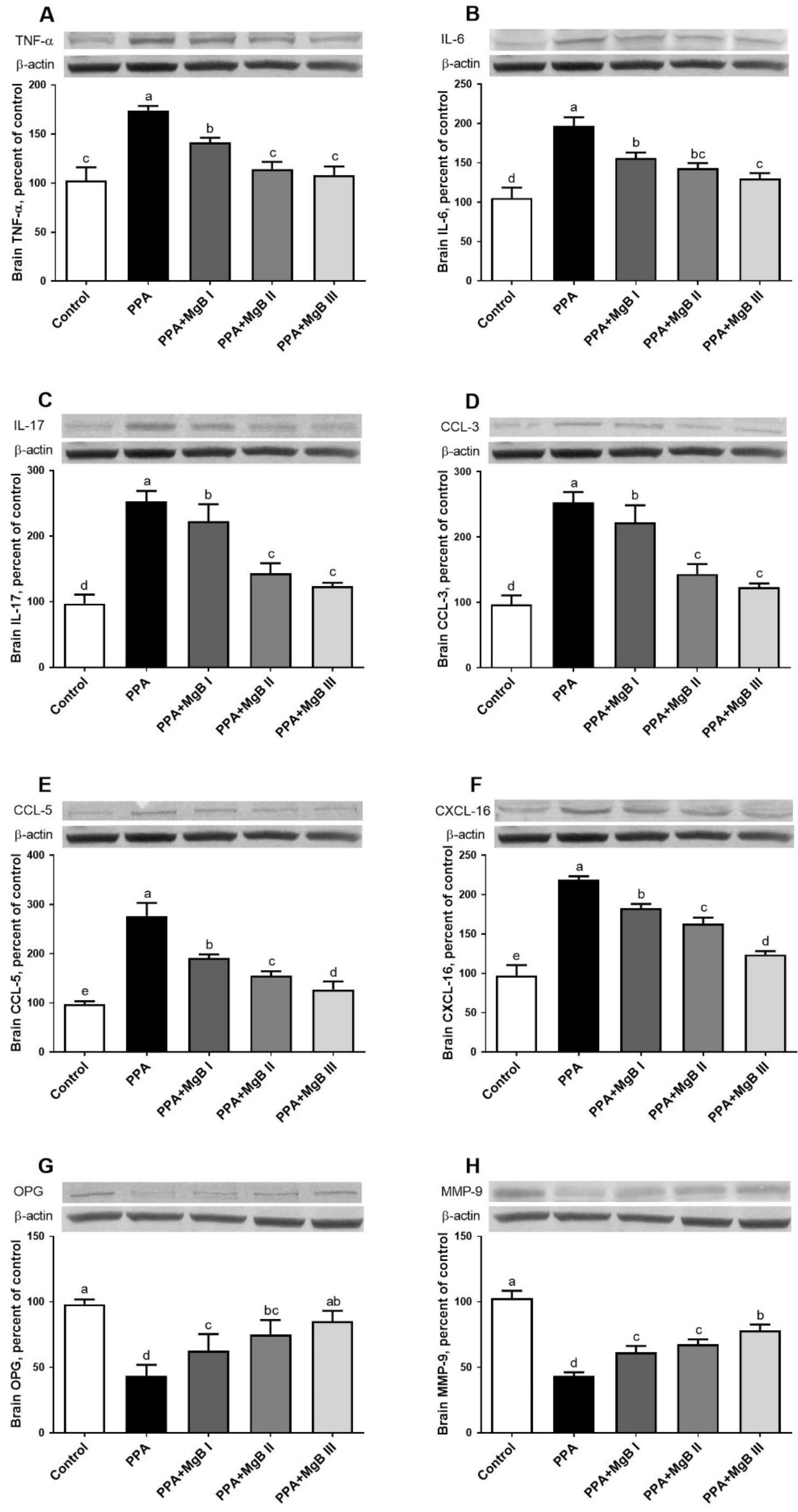
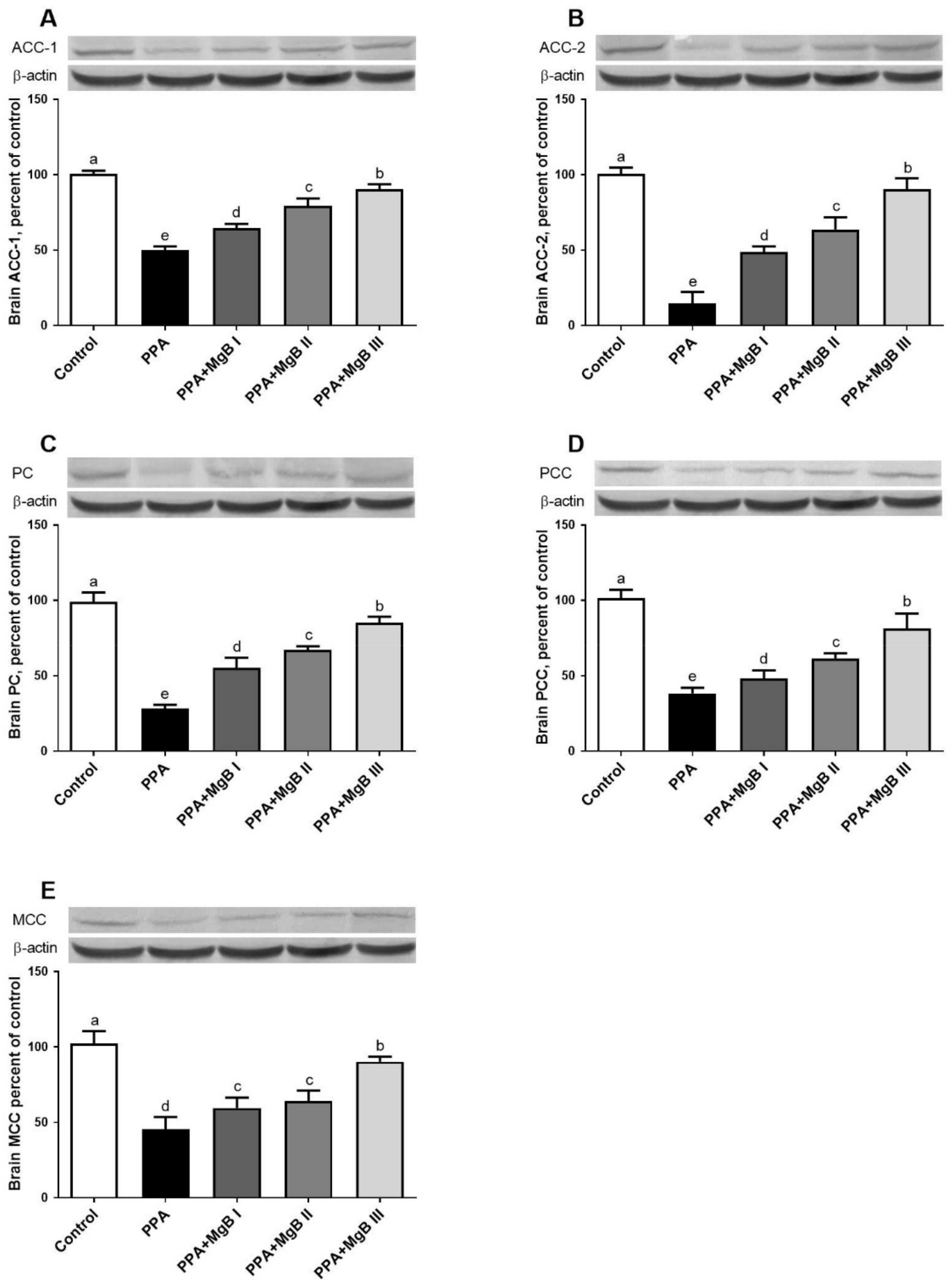
2.5. Western Blot Analyses
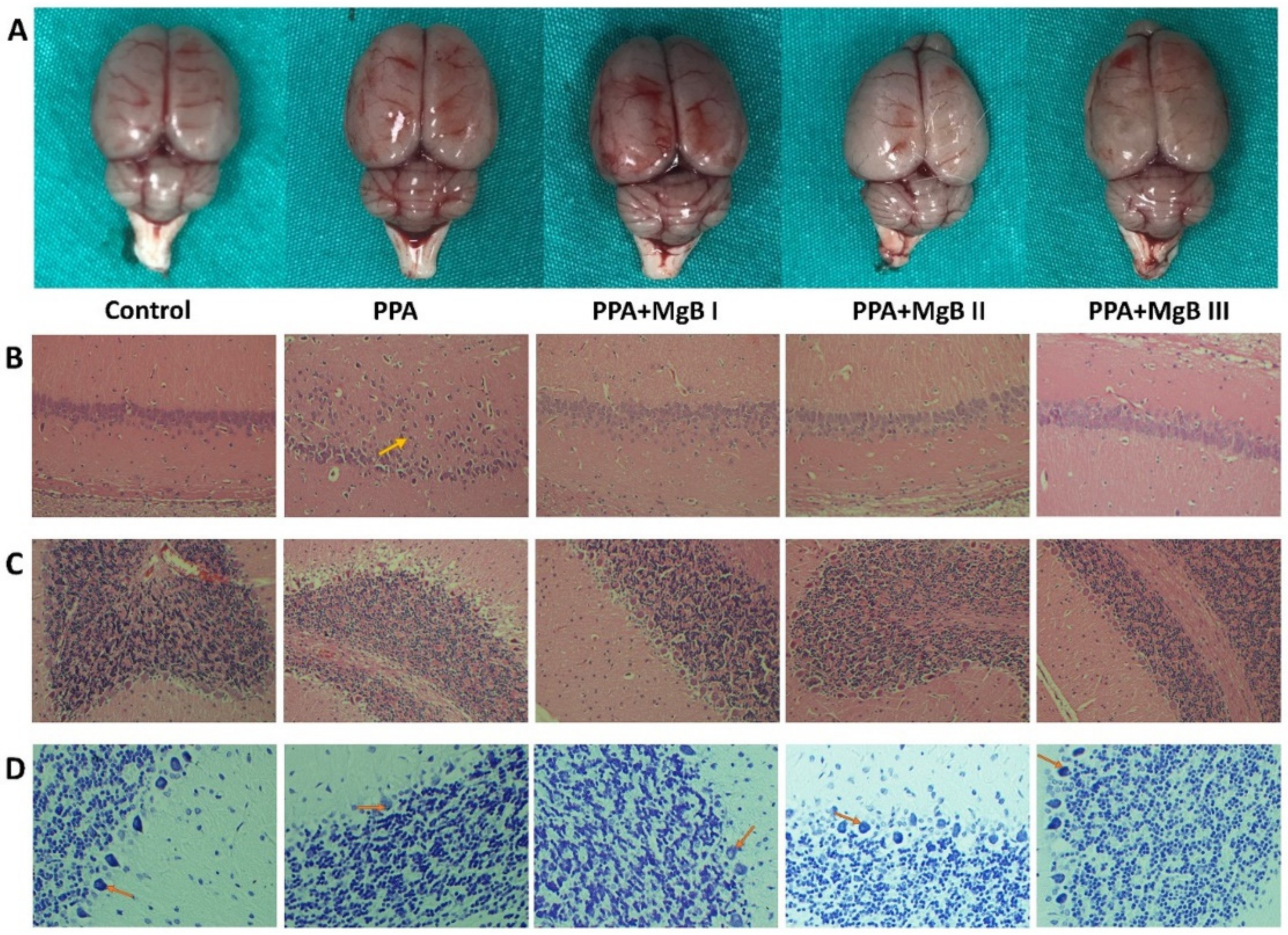
2.6. Histological Evaluation
2.7. Statistical Analyses
3. Results
3.1. Effects of MgB Supplementation on Mg, Biotin, Serotonin and Dopamine Concentrations
3.2. Effects of MgB Supplementation on Oxidative Stress-Related Biochemical Parameters
3.3. Effects of MgB Supplementation on Brain Histology
3.4. Effects of MgB Supplementation on Social Behavior, Learning, and Memory
3.5. Effects of MgB Supplementation on Neurotoxicity-Related Cytokines
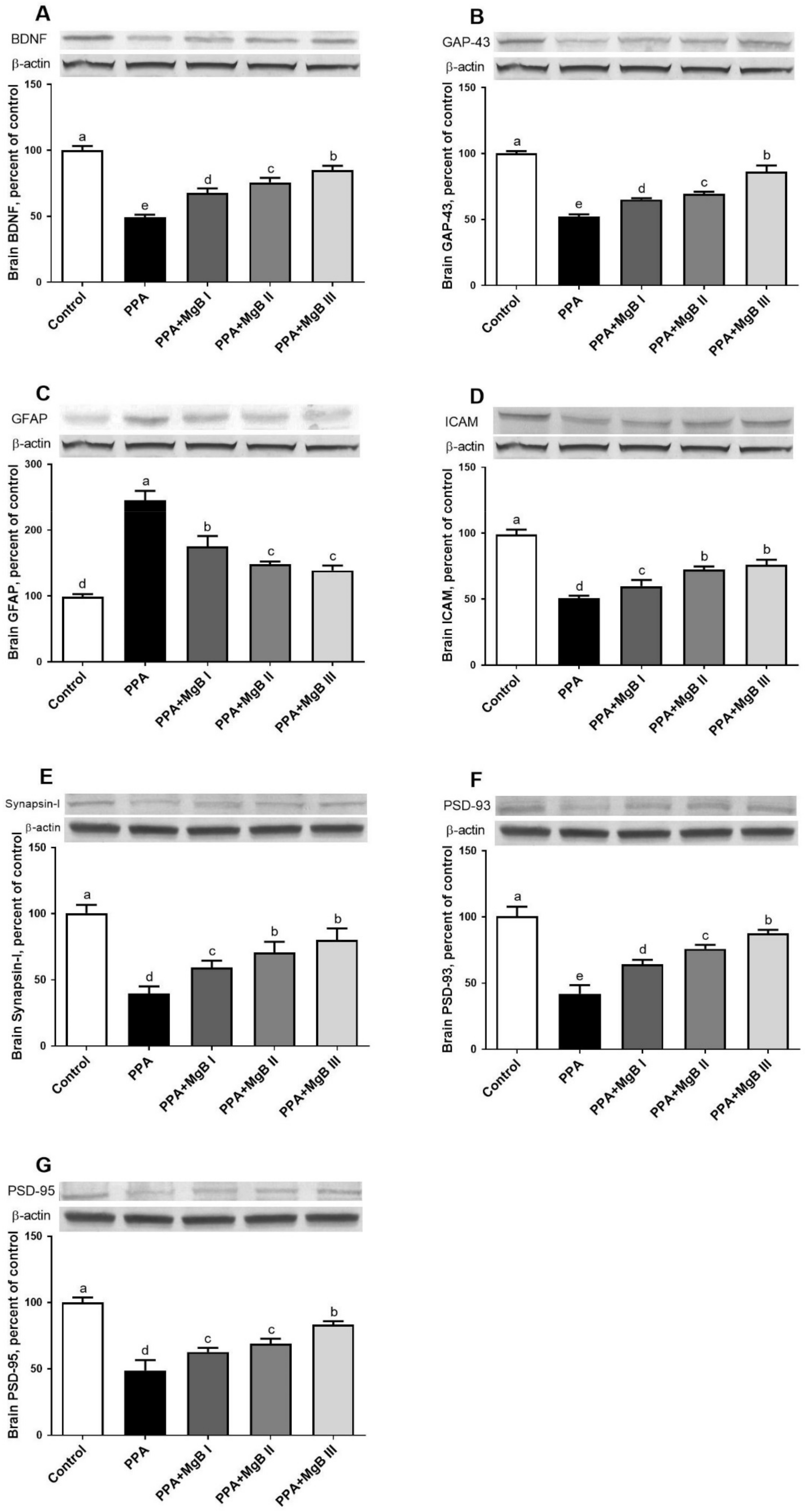
3.6. Effects of MgB Supplementation on Neurodevelopment Markers
3.7. Effects of MgB Supplementation on Neurotransmission-Related Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geschwind, D.H.; State, M.W. Gene hunting in autism spectrum disorder: On the path to precision medicine. Lancet Neurol. 2015, 14, 1109–1120. [Google Scholar] [CrossRef] [Green Version]
- Troyb, E.; Knoch, K.; Barton, M.L. Phenomenology of ASD: Definition, syndromes, and major features. In The Neuropsychology of Autism; Oxford University Press: Oxford, UK, 2011; pp. 9–34. [Google Scholar]
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current enlightenment about etiology and pharmacological treatment of autism spectrum disorder. Front. Neurosci. 2018, 12, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagnozzi, A.M.; Conti, E.; Calderoni, S.; Fripp, J.; Rose, S.E. A systematic review of structural MRI biomarkers in autism spectrum disorder: A machine learning perspective. Int. J. Dev. Neurosci. 2018, 71, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Roeyers, H. Exploring brain functions in autism spectrum disorder: A systematic review on functional near-infrared spectroscopy (fNIRS) studies. Int. J. Psychophysiol. 2019, 137, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Lee, S.; Won, J.; Jin, Y.; Hong, Y.; Hur, T.-Y.; Kim, J.-H.; Lee, S.-R.; Hong, Y. Pathophysiological and neurobehavioral characteristics of a propionic acid-mediated autism-like rat model. PLoS ONE 2018, 13, e0192925. [Google Scholar] [CrossRef] [Green Version]
- El-Ansary, A.K.; Bacha, A.B.; Kotb, M. Etiology of autistic features: The persisting neurotoxic effects of propionic acid. J. Neuroinflamm. 2012, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Macfabe, D.; Caın, D.; Rodrıguezcapote, K.; Franklın, A.; Hoffman, J.; Boon, F.; Taylor, A.; Kavalıers, M.; Ossenkopp, K. Neurobiological effects of intraventricular propionic acid in rats: Possible role of short chain fatty acids on the pathogenesis and characteristics of autism spectrum disorders. Behav. Brain Res. 2007, 176, 149–169. [Google Scholar] [CrossRef]
- Khalil, S.R.; Abd-Elhakim, Y.M.; Selim, M.E.; Al-Ayadhi, L.Y. Apitoxin protects rat pups brain from propionic acid-induced oxidative stress: The expression pattern of Bcl-2 and Caspase-3 apoptotic genes. Neurotoxicology 2015, 49, 121–131. [Google Scholar] [CrossRef]
- Kałużna-Czaplińska, J.; Jóźwik-Pruska, J. Chromatographic and mass spectrometric techniques in studies on oxidative stress in autism. J. Chromatogr. B 2016, 1019, 4–14. [Google Scholar] [CrossRef]
- Abdallah, M.W.; Pearce, B.D.; Larsen, N.; Greaves-Lord, K.; Nørgaard-Pedersen, B.; Hougaard, D.M.; Mortensen, E.L.; Grove, J. Amniotic fluid MMP-9 and neurotrophins in autism spectrum disorders: An exploratory study. Autism Res. 2012, 5, 428–433. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Asadi, S.; Patel, A.B. Focal brain inflammation and autism. J. Neuroinflamm. 2013, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.W.; Michel, T.M. Matrix metalloproteinases in autism spectrum disorders. J. Mol. Psychiatry 2013, 1, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Chadman, K.K.; McCloskey, D.P.; Sheikh, A.M.; Malik, M.; Brown, W.T.; Li, X. Brain IL-6 elevation causes neuronal circuitry imbalances and mediates autism-like behaviors. Biochim. Biophys. Acta-Mol. Basis Dis. 2012, 1822, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Stuart, M.J.; Singhal, G.; Baune, B.T. Systematic review of the neurobiological relevance of chemokines to psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Zoheir, K.M.A.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; AL-Ayadhi, L.Y.; Alzahrani, M.Z.; Al-Shabanah, O.A.; Al-Harbi, M.M.; Attia, S.M. Dysregulation of Th1, Th2, Th17, and T regulatory cell-related transcription factor signaling in children with autism. Mol. Neurobiol. 2017, 54, 4390–4400. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Chin, L.S.; Li, L.; Lanier, L.M.; Kosik, K.S.; Greengard, P. Distinct roles of synapsin I and synapsin II during neuronal development. Mol. Med. 1998, 4, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Xu, W. PSD-95-like membrane associated guanylate kinases (PSD-MAGUKs) and synaptic plasticity. Curr. Opin. Neurobiol. 2011, 21, 306–312. [Google Scholar] [CrossRef] [Green Version]
- McAllister, A.K.; Katz, L.C.; Lo, D.C. Neurotrophins and synaptic plasticity. Annu. Rev. Neurosci. 1999, 22, 295–318. [Google Scholar] [CrossRef] [Green Version]
- Nye, C.; Brice, A. Combined vitamin B6-magnesium treatment in autism spectrum disorder. Cochrane Database Syst. Rev. 2005, 2005, CD003497. [Google Scholar] [CrossRef] [Green Version]
- Levy, S.E.; Hyman, S.L. Complementary and alternative medicine treatments for children with autism spectrum disorders. Child Adolesc. Psychiatr. Clin. N. Am. 2008, 17, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Dakshinamurti, K. Biotin—A regulator of gene expression. J. Nutr. Biochem. 2005, 16, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Mejia, C. Pharmacological effects of biotin. J. Nutr. Biochem. 2005, 16, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, J.; Wijeratne, S.S.K.; Hassan, Y.I. Biotin. BioFactors 2009, 35, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Said, H.M. Biotin: Biochemical, physiological and clinical aspects. Subcell. Biochem. 2012, 56, 1–19. [Google Scholar] [CrossRef]
- Xu, M.; Dai, W.; Deng, X. Effects of magnesium sulfate on brain mitochondrial respiratory function in rats after experimental traumatic brain injury. Chin. J. Traumatol. = Zhonghua Chuang Shang Za Zhi 2002, 5, 361–364. [Google Scholar]
- Barbagallo, M.; Dominguez, L. Magnesium and aging. Curr. Pharm. Des. 2010, 16, 832–839. [Google Scholar] [CrossRef]
- Colamaria, V.; Burlina, A.B.; Gaburro, D.; Pajno-Ferrara, F.; Saudubray, J.M.; Merino, R.G.; Bemardina, B.D. Biotin-Responsive infantile encephalopathy: EEG-polygraphic study of a case. Epilepsia 1989, 30, 573–578. [Google Scholar] [CrossRef]
- Benke, P.J.; Duchowny, M.; McKnight, D. Biotin and acetazolamide for treatment of an unusual child with autism plus lack of nail and hair growth. Pediatr. Neurol. 2018, 79, 61–64. [Google Scholar] [CrossRef]
- Zaffanello, M.; Zamboni, G.; Fontana, E.; Zoccante, L.; Tatò, L. A case of partial biotinidase deficiency associated with autism. Child Neuropsychol. 2003, 9, 184–188. [Google Scholar] [CrossRef]
- Findling, R.L.; Maxwell, K.; Scotese-Wojtila, L.; Huang, J.; Yamashita, T.; Wiznitzer, M. High-dose pyridoxine and magnesium administration in children with autistic disorder: An absence of salutary effects in a double-blind, placebo-controlled study. J. Autism Dev. Disord. 1997, 27, 467–478. [Google Scholar] [CrossRef]
- Lelord, G.; Muh, J.P.; Barthelemy, C.; Martineau, J.; Garreau, B.; Callaway, E. Effects of pyridoxine and magnesium on autistic symptoms?Initial observations. J. Autism Dev. Disord. 1981, 11, 219–230. [Google Scholar] [CrossRef]
- Martineau, J.; Barthelemy, C.; Garreau, B.; Lelord, G. Vitamin B6, magnesium, and combined B6-Mg: Therapeutic effects in childhood autism. Biol. Psychiatry 1985, 20, 467–478. [Google Scholar] [CrossRef]
- Tolbert, L.; Haigler, T.; Waits, M.M.; Dennis, T. Brief report: Lack of response in an autistic population to a low dose clinical trial of pyridoxine plus magnesium. J. Autism Dev. Disord. 1993, 23, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.; Buckel, W. On the mechanism of action of the antifungal agent propionate. Eur. J. Biochem. 2004, 271, 3227–3241. [Google Scholar] [CrossRef] [PubMed]
- Mepham, J.R.; Boon, F.H.; Foley, K.A.; Cain, D.P.; MacFabe, D.F.; Ossenkopp, K.-P. Impaired spatial cognition in adult rats treated with multiple intracerebroventricular (ICV) infusions of the enteric bacterial metabolite, propionic acid, and return to baseline after 1 week of no treatment: Contribution to a rodent model of ASD. Neurotox. Res. 2019, 35, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Feliz, B.; Witt, D.R.; Harris, B.T. Propionic acidemia: A neuropathology case report and review of prior cases. Arch. Pathol. Lab. Med. 2003, 127, e325–e328. [Google Scholar] [CrossRef]
- Gupta, R.; Deshpande, S.B. 3-Nitropropionic acid depresses spinal reflexes involving GABAergic and glycinergic transmission in neonatal rat spinal cord in vitro. Life Sci. 2008, 83, 756–760. [Google Scholar] [CrossRef]
- Wajner, M.; Santos, K.D.; Schlottfeldt, J.L.; Rocha, M.P.; Wannmacher, C.M. Inhibition of mitogen-activated proliferation of human peripheral lymphocytes in vitro by propionic acid. Clin. Sci. 1999, 96, 99–103. [Google Scholar] [CrossRef]
- Shultz, S.R.; MacFabe, D.F.; Ossenkopp, K.-P.; Scratch, S.; Whelan, J.; Taylor, R.; Cain, D.P. Intracerebroventricular injection of propionic acid, an enteric bacterial metabolic end-product, impairs social behavior in the rat: Implications for an animal model of autism. Neuropharmacology 2008, 54, 901–911. [Google Scholar] [CrossRef]
- Shultz, S.R.; MacFabe, D.F.; Martin, S.; Jackson, J.; Taylor, R.; Boon, F.; Ossenkopp, K.-P.; Cain, D.P. Intracerebroventricular injections of the enteric bacterial metabolic product propionic acid impair cognition and sensorimotor ability in the Long–Evans rat: Further development of a rodent model of autism. Behav. Brain Res. 2009, 200, 33–41. [Google Scholar] [CrossRef]
- Thomas, R.H.; Foley, K.A.; Mepham, J.R.; Tichenoff, L.J.; Possmayer, F.; MacFabe, D.F. Altered brain phospholipid and acylcarnitine profiles in propionic acid infused rodents: Further development of a potential model of autism spectrum disorders. J. Neurochem. 2010, 113, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.E.; Melnyk, S.; MacFabe, D.F. Unique acyl-carnitine profiles are potential biomarkers for acquired mitochondrial disease in autism spectrum disorder. Transl. Psychiatry 2013, 3, e220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojalvo, S.P.; Sylla, S.; Komorowski, J.; Orhan, C.; Tuzcu, M.; Sahin, N.; Sahin, K. The safety and absorption of magnesium biotinate in rats (P06-029-19). Curr. Dev. Nutr. 2019, 3, nzz031.P06-029-19. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.W.; Seol, I.C.; Son, C.G. Interpretation of animal dose and human equivalent dose for drug development. J. Korean Med. 2010, 31, 1–7. [Google Scholar]
- Nair, A.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.; Rathman, S.; McMahon, R. Dietary biotin intake modulates the pool of free and protein-bound biotin in rat liver. J. Nutr. 2001, 131, 2310–2315. [Google Scholar] [CrossRef] [Green Version]
- Olszewska-Słonina, D.M.; Mątewski, D.; Czajkowski, R.; Olszewski, K.J.; Woźniak, A.; Odrowąż-Sypniewska, G.; Lis, K.; Musiałkiewicz, D.; Kowaliszyn, B. The concentration of thiobarbituric acid reactive substances (TBARS) and paraoxonase activity in blood of patients with osteoarthrosis after endoprosthesis implantation. Med. Sci. Monit. 2011, 17, CR498–CR504. [Google Scholar] [CrossRef] [Green Version]
- Giustarini, D.; Fanti, P.; Matteucci, E.; Rossi, R. Micro-method for the determination of glutathione in human blood. J. Chromatogr. B 2014, 964, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Serra, J.A.; Marschoff, E.R.; Domínguez, R.O.; de Lustig, E.S.; Famulari, A.L.; Bartolomé, E.L.; Guareschi, E.M. Comparison of the determination of superoxide dismutase and antioxidant capacity in neurological patients using two different procedures. Clin. Chim. Acta 2000, 301, 87–102. [Google Scholar] [CrossRef]
- Hadwan, M.H. New method for assessment of serum catalase activity. Indian J. Sci. Technol. 2016, 9. [Google Scholar] [CrossRef]
- Foley, K.A.; Ossenkopp, K.-P.; Kavaliers, M.; MacFabe, D.F. Pre- and neonatal exposure to lipopolysaccharide or the enteric metabolite, propionic acid, alters development and behavior in adolescent rats in a sexually dimorphic manner. PLoS ONE 2014, 9, e87072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharman Moser, S.; Davidovitch, M.; Rotem, R.S.; Chodick, G.; Shalev, V.; Koren, G. High dose folic acid during pregnancy and the risk of autism; The birth order bias: A nested case-control study. Reprod. Toxicol. 2019, 89, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Tsugiyama, L.E.; Ida-Eto, M.; Ohkawara, T.; Noro, Y.; Narita, M. Altered neuronal activity in the auditory brainstem following sound stimulation in thalidomide-induced autism model rats. Congenit. Anom. 2020, 60, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Hu, Y.; Chen, D.; Li, Y.; Wang, S. Improvement of a mouse model of valproic acid-induced autism. Nan Fang Yi Ke Da Xue Xue Bao 2019, 39, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, L.S.; Samsam, A.; Naser, S.A. Propionic acid induces gliosis and neuro-inflammation through modulation of PTEN/AKT pathway in autism spectrum disorder. Sci. Rep. 2019, 9, 8824. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Han, P.-L. Early-life stress in D2 heterozygous mice promotes autistic-like behaviors through the downregulation of the BDNF-TrkB pathway in the dorsal striatum. Exp. Neurobiol. 2019, 28, 337–351. [Google Scholar] [CrossRef]
- Qi, Z.; Lyu, M.; Yang, L.; Yuan, H.; Cao, Y.; Zhai, L.; Dang, W.; Liu, J.; Yang, F.; Li, Y. A novel and reliable rat model of autism. Front. Psychiatry 2021, 12, 549810. [Google Scholar] [CrossRef]
- Nicolini, C.; Fahnestock, M. The valproic acid-induced rodent model of autism. Exp. Neurol. 2018, 299, 217–227. [Google Scholar] [CrossRef]
- MacFabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2012, 23, 19260. [Google Scholar] [CrossRef]
- Aldbass, A.M.; Bhat, R.S.; El-Ansary, A. Protective and therapeutic potency of N-acetyl-cysteine on propionic acid-induced biochemical autistic features in rats. J. Neuroinflamm. 2013, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, U. Intracellular pH modulates spontaneous and epileptiform bioelectric activity of hippocampal CA3-neurones. Eur. Neuropsychopharmacol. 2000, 10, 97–103. [Google Scholar] [CrossRef]
- Severson, C.A.; Wang, W.; Pieribone, V.A.; Dohle, C.I.; Richerson, G.B. Midbrain serotonergic neurons are central pH chemoreceptors. Nat. Neurosci. 2003, 6, 1139–1140. [Google Scholar] [CrossRef] [PubMed]
- Lackey, E.P.; Heck, D.H.; Sillitoe, R.V. Recent advances in understanding the mechanisms of cerebellar granule cell development and function and their contribution to behavior. F1000Research 2018, 7, 1142. [Google Scholar] [CrossRef] [PubMed]
- Reeber, S.L.; Otis, T.S.; Sillitoe, R.V. New roles for the cerebellum in health and disease. Front. Syst. Neurosci. 2013, 7, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banker, S.M.; Gu, X.; Schiller, D.; Foss-Feig, J.H. Hippocampal contributions to social and cognitive deficits in autism spectrum disorder. Trends Neurosci. 2021, 44, 793–807. [Google Scholar] [CrossRef]
- Barnea-Goraly, N.; Frazier, T.W.; Piacenza, L.; Minshew, N.J.; Keshavan, M.S.; Reiss, A.L.; Hardan, A.Y. A preliminary longitudinal volumetric MRI study of amygdala and hippocampal volumes in autism. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 124–128. [Google Scholar] [CrossRef]
- Schumann, C.M. The amygdala is enlarged in children but not adolescents with autism; the hippocampus is enlarged at all ages. J. Neurosci. 2004, 24, 6392–6401. [Google Scholar] [CrossRef]
- Groen, W.; Teluij, M.; Buitelaar, J.; Tendolkar, I. Amygdala and hippocampus enlargement during adolescence in autism. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 552–560. [Google Scholar] [CrossRef]
- Aylward, E.H.; Minshew, N.J.; Goldstein, G.; Honeycutt, N.A.; Augustine, A.M.; Yates, K.O.; Barta, P.E.; Pearlson, G.D. MRI volumes of amygdala and hippocampus in non-mentally retarded autistic adolescents and adults. Neurology 1999, 53, 2145. [Google Scholar] [CrossRef]
- Nicolson, R.; DeVito, T.J.; Vidal, C.N.; Sui, Y.; Hayashi, K.M.; Drost, D.J.; Williamson, P.C.; Rajakumar, N.; Toga, A.W.; Thompson, P.M. Detection and mapping of hippocampal abnormalities in autism. Psychiatry Res. Neuroimaging 2006, 148, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Uppal, N.; Puri, R.; Yuk, F.; Janssen, W.G.M.; Bozdagi-Gunal, O.; Harony-Nicolas, H.; Dickstein, D.L.; Buxbaum, J.D.; Hof, P.R. Ultrastructural analyses in the hippocampus CA1 field in Shank3-deficient mice. Mol. Autism 2015, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; McCoy, P.A.; Rodriguiz, R.M.; Pan, Y.; Je, H.S.; Roberts, A.C.; Kim, C.J.; Berrios, J.; Colvin, J.S.; Bousquet-Moore, D.; et al. Synaptic dysfunction and abnormal behaviors in mice lacking major isoforms of Shank3. Hum. Mol. Genet. 2011, 20, 3093–3108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The neurochemistry of autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, C.L.; Anacker, A.M.J.; Veenstra-VanderWeele, J. The serotonin system in autism spectrum disorder: From biomarker to animal models. Neuroscience 2016, 321, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Siemann, J.K.; Muller, C.L.; Forsberg, C.G.; Blakely, R.D.; Veenstra-VanderWeele, J.; Wallace, M.T. An autism-associated serotonin transporter variant disrupts multisensory processing. Transl. Psychiatry 2017, 7, e1067. [Google Scholar] [CrossRef]
- Abdulamir, H.A.; Abdul-Rasheed, O.F.; Abdulghani, E.A. Serotonin and serotonin transporter levels in autistic children. Saudi Med. J. 2018, 39, 487–494. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Persico, A.M. Blood serotonin levels in autism spectrum disorder: A systematic review and meta-analysis. Eur. Neuropsychopharmacol. 2014, 24, 919–929. [Google Scholar] [CrossRef]
- Narita, N.; Kato, M.; Tazoe, M.; Miyazaki, K.; Narita, M.; Okado, N. Increased monoamine concentration in the brain and blood of fetal thalidomide- and valproic acid–exposed rat: Putative animal models for autism. Pediatr. Res. 2002, 52, 576–579. [Google Scholar] [CrossRef] [Green Version]
- Azmitia, E.C.; Singh, J.S.; Hou, X.P.; Wegiel, J. Dystrophic serotonin axons in postmortem brains from young autism patients. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2011, 294, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.; Reutiman, T.J.; Folsom, T.D.; Sohr, R.; Wolf, R.J.; Juckel, G.; Fatemi, S.H. Dopamine and serotonin levels following prenatal viral infection in mouse—Implications for psychiatric disorders such as schizophrenia and autism. Eur. Neuropsychopharmacol. 2008, 18, 712–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dichter, G.S.; Felder, J.N.; Green, S.R.; Rittenberg, A.M.; Sasson, N.J.; Bodfish, J.W. Reward circuitry function in autism spectrum disorders. Soc. Cogn. Affect. Neurosci. 2012, 7, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dichter, G.S.; Damiano, C.A.; Allen, J.A. Reward circuitry dysfunction in psychiatric and neurodevelopmental disorders and genetic syndromes: Animal models and clinical findings. J. Neurodev. Disord. 2012, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polšek, D.; Jagatic, T.; Cepanec, M.; Hof, P.; Šimić, G. Recent developments in neuropathology of autism spectrum disorders. Transl. Neurosci. 2011, 2, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Mazur, A.; Maier, J.A.M.; Rock, E.; Gueux, E.; Nowacki, W.; Rayssiguier, Y. Magnesium and the inflammatory response: Potential physiopathological implications. Arch. Biochem. Biophys. 2007, 458, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Elahi, A.; Sabui, S.; Narasappa, N.N.; Agrawal, S.; Lambrecht, N.W.; Agrawal, A.; Said, H.M. Biotin deficiency induces Th1- and Th17-mediated proinflammatory responses in human CD4 + T lymphocytes via activation of the mTOR signaling pathway. J. Immunol. 2018, 200, 2563–2570. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Agrawal, A.; Said, H.M. Biotin deficiency enhances the inflammatory response of human dendritic cells. Am. J. Physiol. Physiol. 2016, 311, C386–C391. [Google Scholar] [CrossRef]
- Zoroglu, S.S.; Armutcu, F.; Ozen, S.; Gurel, A.; Sivasli, E.; Yetkin, O.; Meram, I. Increased oxidative stress and altered activities of erythrocyte free radical scavenging enzymes in autism. Eur. Arch. Psychiatry Clin. Neurosci. 2004, 254, 143–147. [Google Scholar] [CrossRef]
- Gao, J.; Wang, X.; Sun, H.; Cao, Y.; Liang, S.; Wang, H.; Wang, Y.; Yang, F.; Zhang, F.; Wu, L. Neuroprotective effects of docosahexaenoic acid on hippocampal cell death and learning and memory impairments in a valproic acid-induced rat autism model. Int. J. Dev. Neurosci. 2016, 49, 67–78. [Google Scholar] [CrossRef]
- Parker-Athill, E.C.; Tan, J. Maternal immune activation and autism spectrum disorder: Interleukin-6 signaling as a key mechanistic pathway. Neurosignals 2010, 18, 113–128. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, H.G.; Kusek, G.K.; Yang, M.; Phoenix, J.L.; Bolivar, V.J.; Crawley, J.N. Autism-like behavioral phenotypes in BTBR T+tf/J mice. Genes Brain Behav. 2008, 7, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.; Zhang, Y.; Gao, D.; Miller, V.M.; Lawrence, D.A. Aberrant immune responses in a mouse with behavioral disorders. PLoS ONE 2011, 6, e20912. [Google Scholar] [CrossRef] [PubMed]
- Hope, S. Osteoprotegerin levels in patients with severe mental disorders. J. Psychiatry Neurosci. 2010, 35, 304–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveron-Negrete, L.; Fernandez-Mejia, C. Pharmacological effects of biotin in animals. Mini-Rev. Med. Chem. 2017, 17, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, S.M.; Razak, K.; Ethell, I.M. A delicate balance: Role of MMP-9 in brain development and pathophysiology of neurodevelopmental disorders. Front. Cell. Neurosci. 2015, 9, 280. [Google Scholar] [CrossRef] [Green Version]
- Fassio, A.; Patry, L.; Congia, S.; Onofri, F.; Piton, A.; Gauthier, J.; Pozzi, D.; Messa, M.; Defranchi, E.; Fadda, M.; et al. SYN1 loss-of-function mutations in autism and partial epilepsy cause impaired synaptic function. Hum. Mol. Genet. 2011, 20, 2297–2307. [Google Scholar] [CrossRef]
- Carlisle, H.J.; Fink, A.E.; Grant, S.G.N.; O’Dell, T.J. Opposing effects of PSD-93 and PSD-95 on long-term potentiation and spike timing-dependent plasticity. J. Physiol. 2008, 586, 5885–5900. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, K.K.W. Glial fibrillary acidic protein: From intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015, 38, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Esnafoglu, E.; Ayyıldız, S.N.; Cırrık, S.; Erturk, E.Y.; Erdil, A.; Daglı, A.; Noyan, T. Evaluation of serum Neuron-specific enolase, S100B, myelin basic protein and glial fibrilliary acidic protein as brain specific proteins in children with autism spectrum disorder. Int. J. Dev. Neurosci. 2017, 61, 86–91. [Google Scholar] [CrossRef]
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann. Neurol. 2005, 57, 67–81. [Google Scholar] [CrossRef]
- Slutsky, I.; Sadeghpour, S.; Li, B.; Liu, G. Enhancement of synaptic plasticity through chronically reduced Ca2+ flux during uncorrelated activity. Neuron 2004, 44, 835–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.-P.; Li, L.; Bao, J.; Wang, Z.-H.; Zeng, J.; Liu, E.-J.; Li, X.-G.; Huang, R.-X.; Gao, D.; Li, M.-Z.; et al. Magnesium protects cognitive functions and synaptic plasticity in streptozotocin-induced sporadic alzheimer’s model. PLoS ONE 2014, 9, e108645. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Groups | ||||
|---|---|---|---|---|---|
| Control | PPA | PPA+MgB I | PPA+MgB II | PPA+MgB III | |
| Serum Mg | 3.36 ± 0.14 c | 2.20 ± 0.08 e | 2.76 ± 0.05 d | 4.33 ± 0.06 b | 5.88 ± 0.12 a |
| Brain Mg | 143.84 ± 1.97 b | 97.12 ± 2.99 d | 113.24 ± 5.11 c | 144.21 ± 3.58 b | 168.13 ± 4.28 a |
| Serum Biotin | 72.30 ± 1.13 c | 49.16 ± 1.80 d | 55.81 ± 3.40 d | 99.94 ± 2.43 b | 121.50 ± 3.06 a |
| Brain Biotin | 0.159 ± 0.006 bc | 0.099 ± 0.007 d | 0.146 ± 0.007 c | 0.174 ± 0.006 ab | 0.187 ± 0.007 a |
| Serotonin | 54.85 ± 0.94 a | 11.24 ± 0.47 e | 17.07 ± 0.90 d | 27.20 ± 1.24 c | 36.95 ± 0.59 b |
| Dopamine | 8.44 ± 0.88 d | 36.61 ± 2.01 a | 29.03 ± 1.26 b | 18.58 ± 1.04 c | 12.64 ± 0.70 d |
| Items | Groups | ||||
|---|---|---|---|---|---|
| Control | PPA | PPA+MgB I | PPA+MgB II | PPA+MgB III | |
| Serum MDA | 0.57 ± 0.04 e | 1.93 ± 0.03 a | 1.65 ± 0.04 b | 1.41 ± 0.02 c | 0.93 ± 0.03d |
| Brain MDA | 1.84 ± 0.03 e | 5.96 ± 0.17 a | 4.60 ± 0.12 b | 3.80 ± 0.04 c | 2.86 ± 0.08 d |
| CAT | 32.53 ± 1.69 a | 10.12 ± 0.46 d | 13.42 ± 0.87 d | 20.77 ± 0.64 c | 26.56 ± 0.56 b |
| SOD | 145.36 ± 1.96 a | 51.17 ± 2.07 e | 64.10 ± 2.94 d | 93.51 ± 2.38 c | 126.47 ± 2.78 b |
| GPx | 26.29 ± 0.88 a | 7.71 ± 0.31 d | 9.48 ± 0.50 d | 14.19 ± 0.79 c | 19.82 ± 0.99 b |
| GSH | 68.65 ± 2.22 a | 28.03 ± 1.62 d | 31.92 ± 1.83 d | 42.15 ± 1.92 c | 53.19 ± 2.25 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahin, K.; Orhan, C.; Karatoprak, S.; Tuzcu, M.; Deeh, P.B.D.; Ozercan, I.H.; Sahin, N.; Bozoglan, M.Y.; Sylla, S.; Ojalvo, S.P.; et al. Therapeutic Effects of a Novel Form of Biotin on Propionic Acid-Induced Autistic Features in Rats. Nutrients 2022, 14, 1280. https://doi.org/10.3390/nu14061280
Sahin K, Orhan C, Karatoprak S, Tuzcu M, Deeh PBD, Ozercan IH, Sahin N, Bozoglan MY, Sylla S, Ojalvo SP, et al. Therapeutic Effects of a Novel Form of Biotin on Propionic Acid-Induced Autistic Features in Rats. Nutrients. 2022; 14(6):1280. https://doi.org/10.3390/nu14061280
Chicago/Turabian StyleSahin, Kazim, Cemal Orhan, Serdar Karatoprak, Mehmet Tuzcu, Patrick Brice Defo Deeh, Ibrahim Hanifi Ozercan, Nurhan Sahin, Merve Yilmaz Bozoglan, Sarah Sylla, Sara Perez Ojalvo, and et al. 2022. "Therapeutic Effects of a Novel Form of Biotin on Propionic Acid-Induced Autistic Features in Rats" Nutrients 14, no. 6: 1280. https://doi.org/10.3390/nu14061280