
d-Allulose Improves Endurance and Recovery from Exhaustion in Male C57BL/6J Mice

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Experimental Protocols
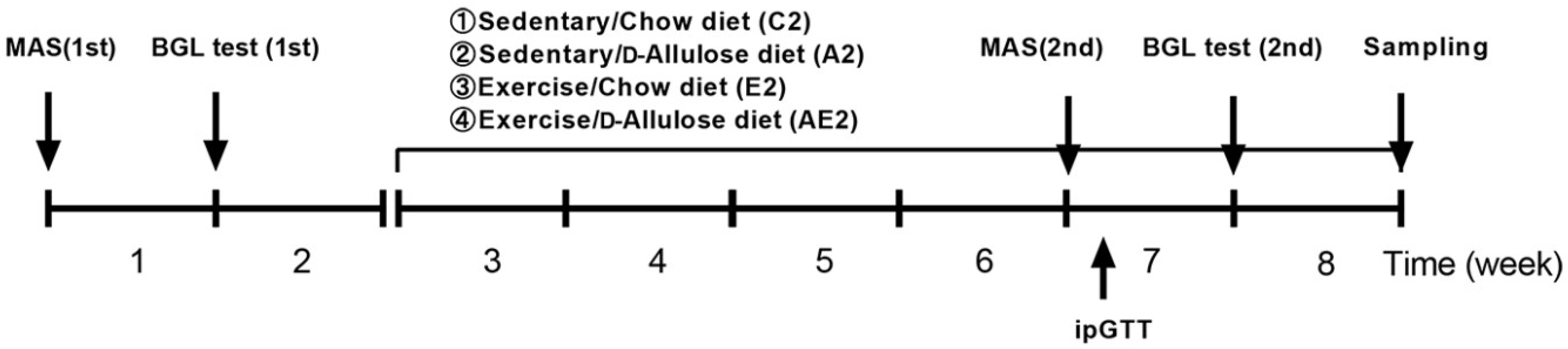
2.2.1. Experiment 1: Effect of Long-Term d-Allulose Administration on Aerobic Performance
2.2.2. Experiment 2: Effect of Long-Term d-Allulose Administration on Maximal Aerobic Speed and Physiological Indicators Related to Aerobic Performance
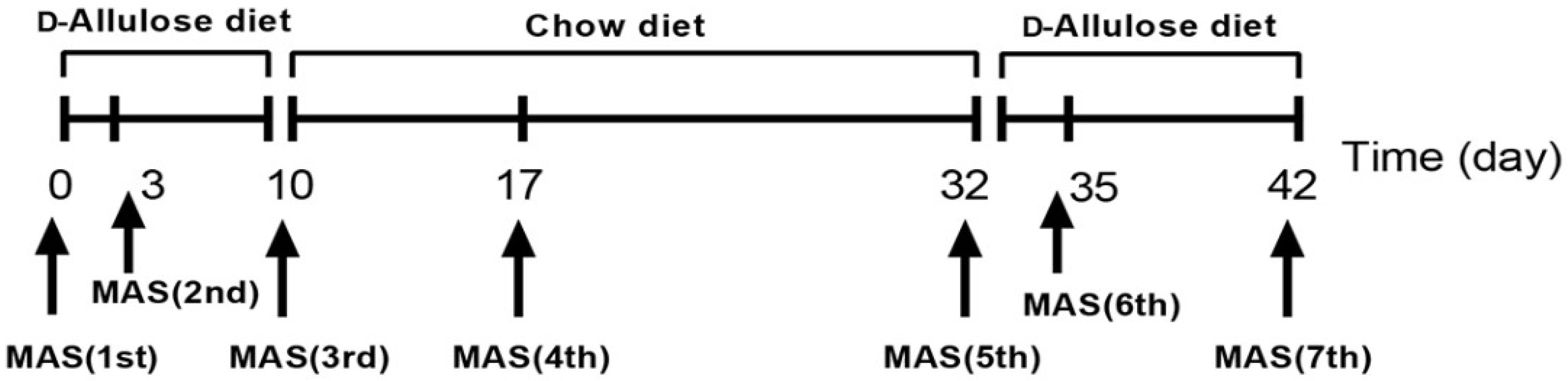
2.2.3. Experiment 3: Effect of Short-Term d-Allulose Administration on MAS, Blood Glucose, and Blood Lactate Levels
2.3. Treadmill Acclimatization and Aerobic Performance Tests
2.3.1. Treadmill and Running Wheel Acclimatization
2.3.2. MAS Test
2.3.3. Endurance Test
2.3.4. Recovery Test
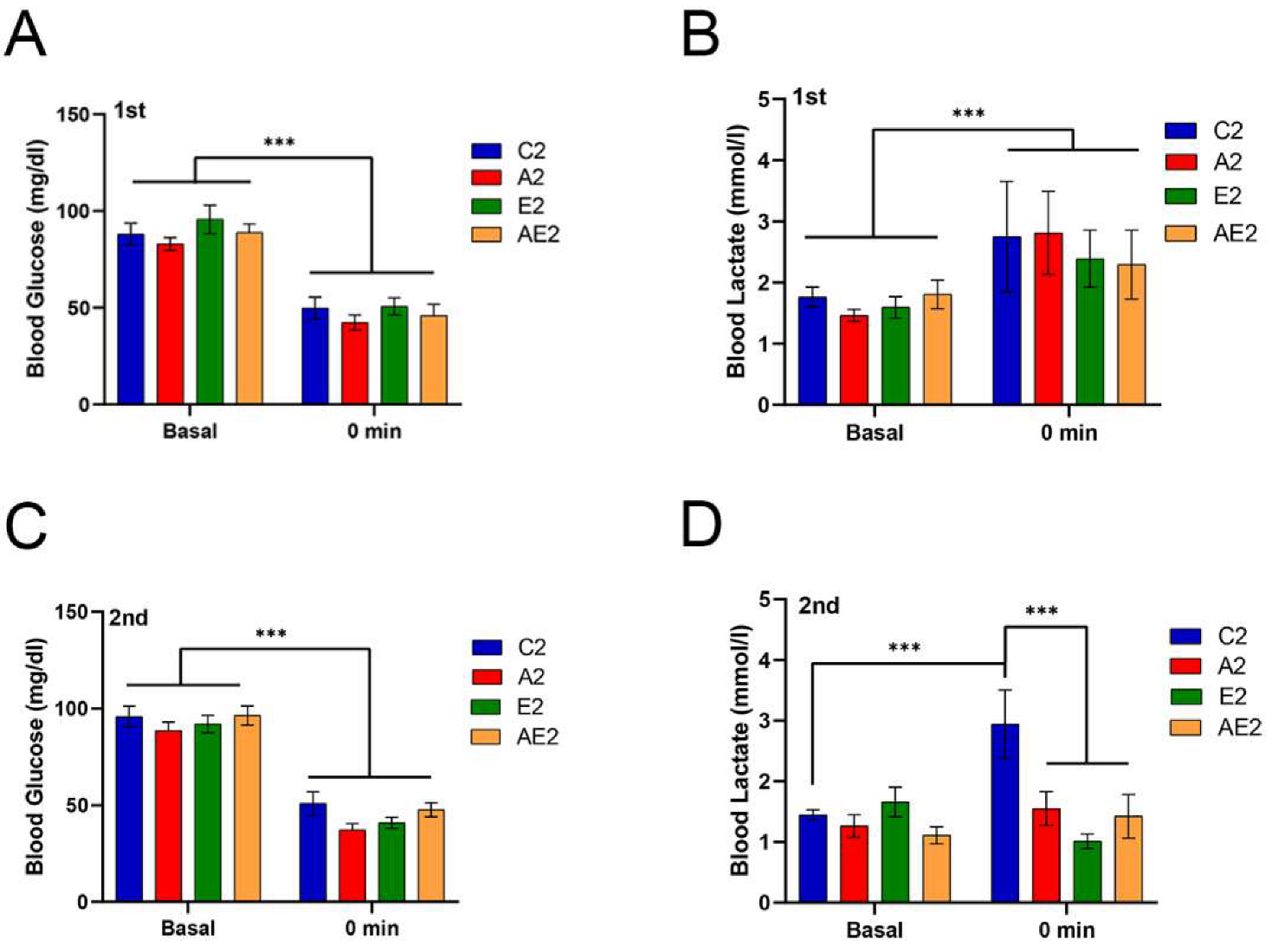
2.3.5. Blood Glucose and Lactate Measurement before and after Running for 2 h at 20 m/min (BGL Test)
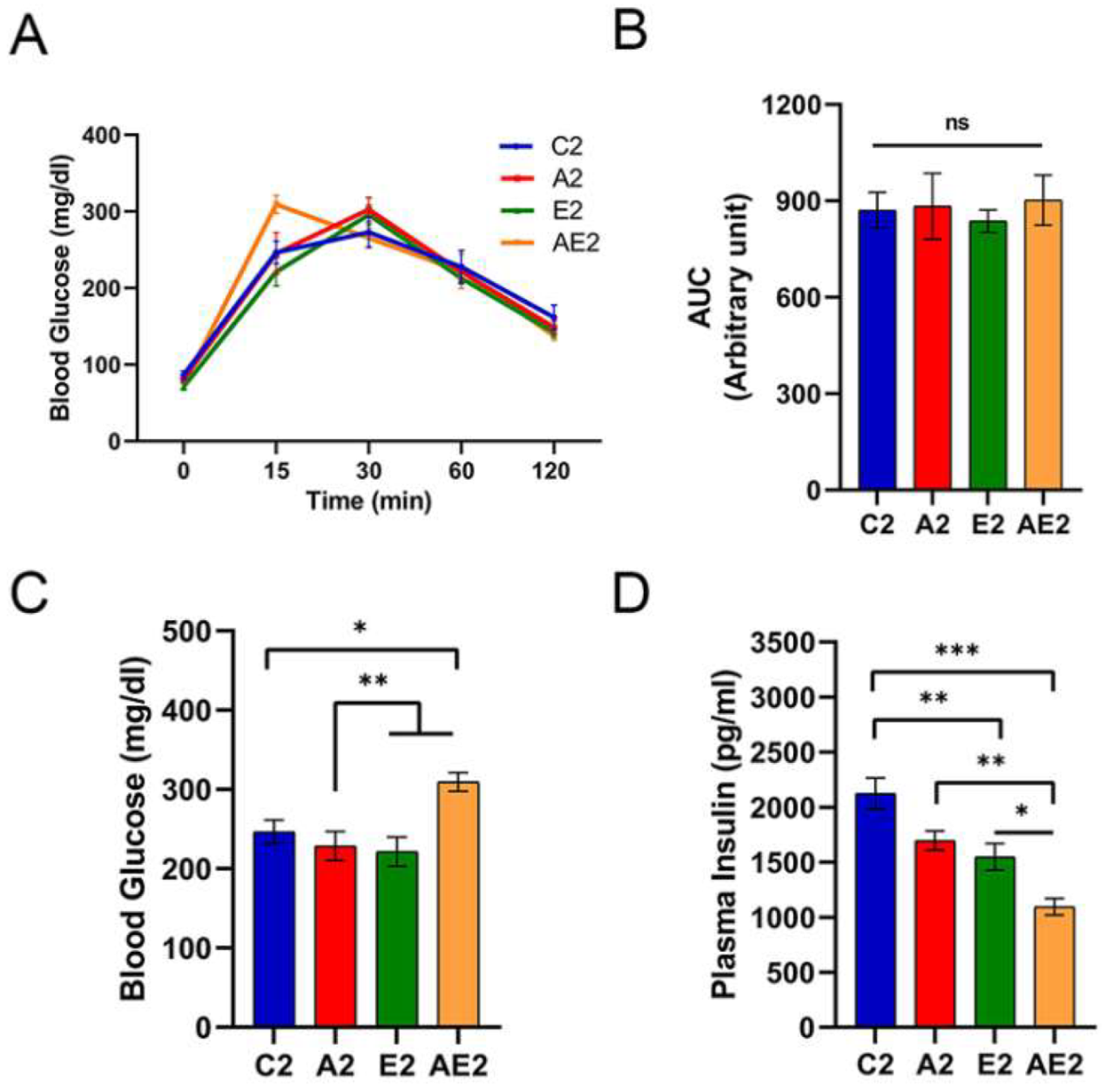
2.4. ipGTT
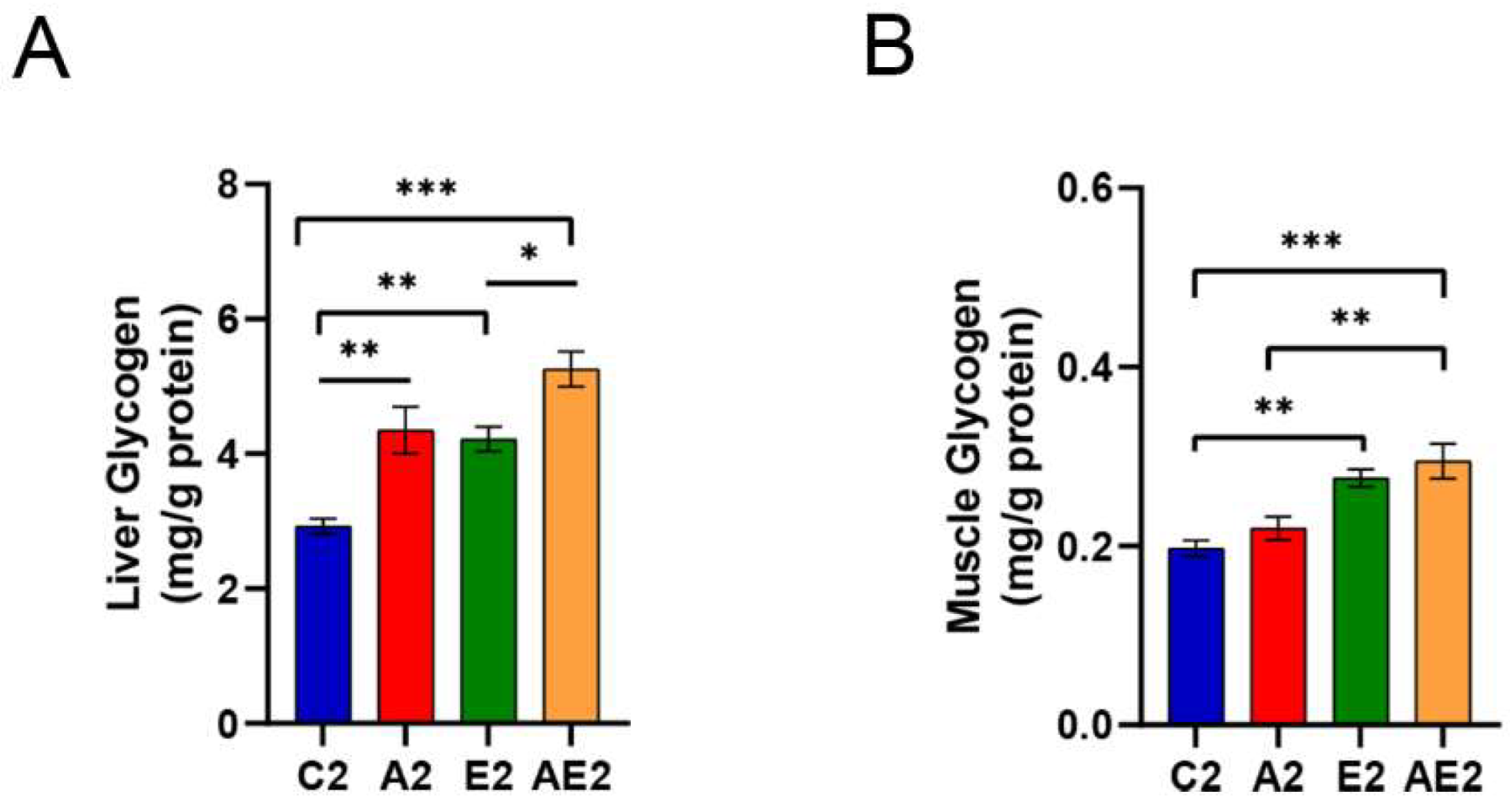
2.5. Liver and Muscle Glycogen Measurement
2.6. Western Blotting
2.7. Statistical Analyses
3. Results
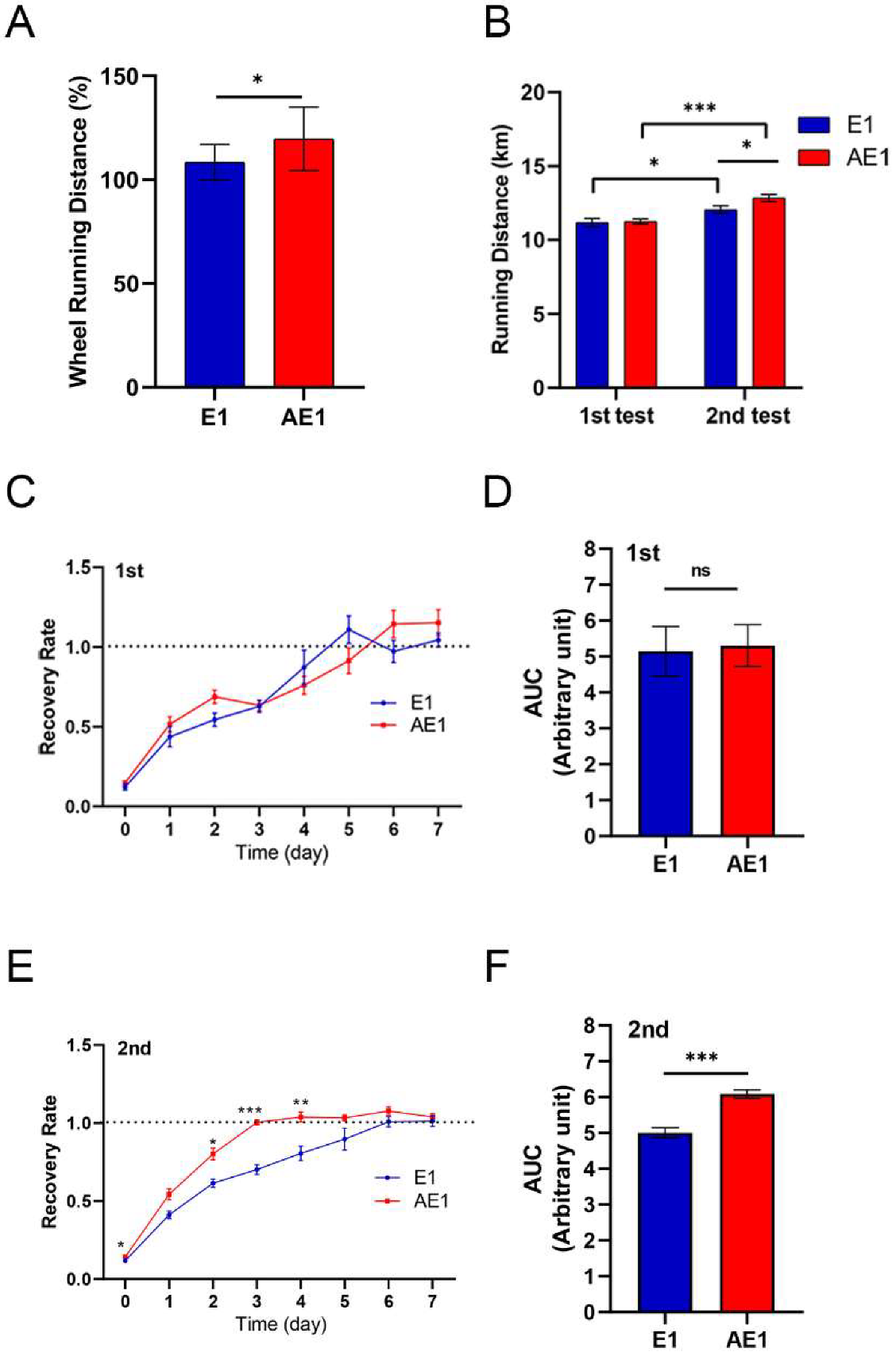
3.1. d-Allulose Improves Endurance and Recovery (Experiment 1)
3.2. Effect of Long-Term Administration of d-Allulose, Exercise, and Their Combination (Experiment 2)
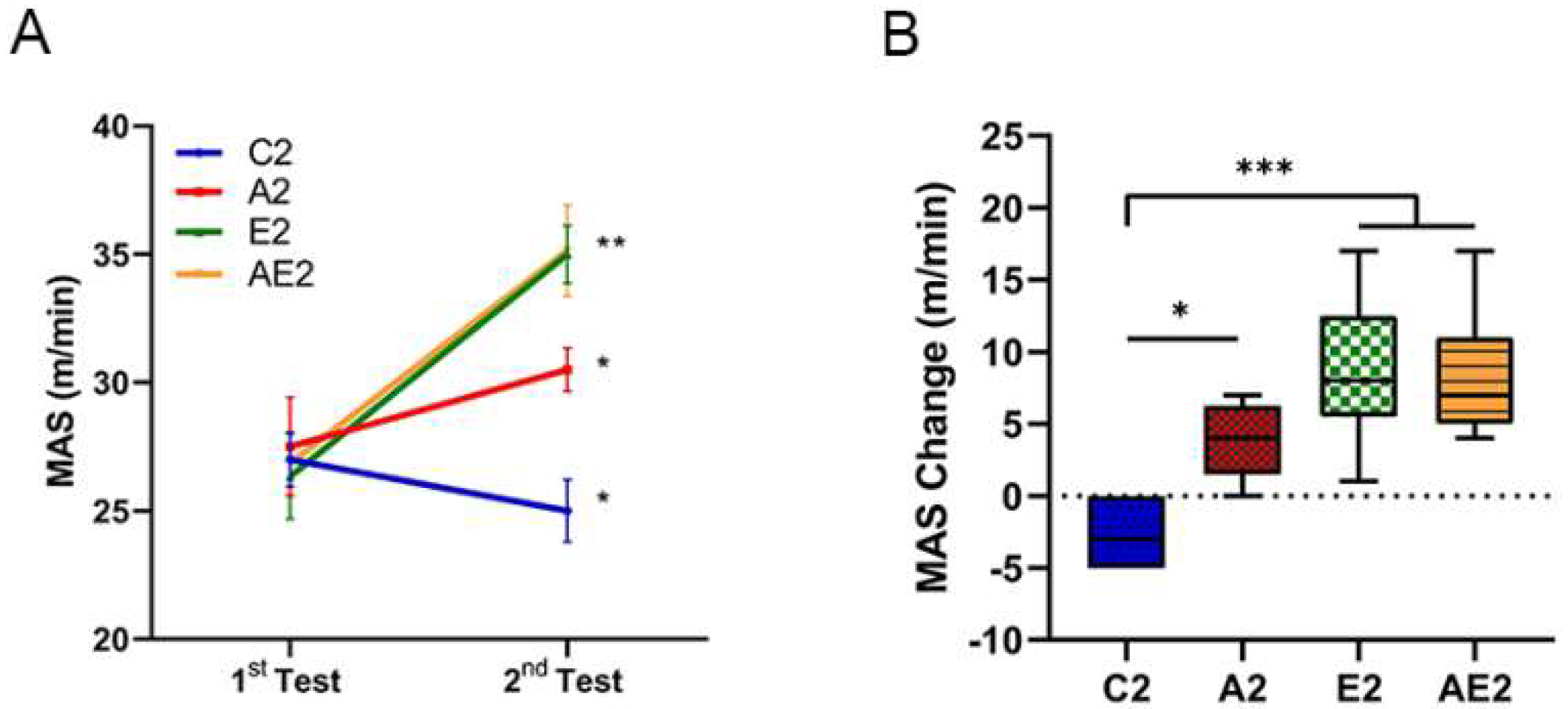
3.2.1. d-Allulose Improves MAS
3.2.2. d-Allulose Suppresses Blood Lactate Increase after Running
3.2.3. d-Allulose Improves Insulin Sensitivity
3.2.4. d-Allulose Prevents Increase in Body Weight and White Adipose Tissue Weights
3.2.5. d-Allulose Increases Liver Glycogen But Not Muscle Glycogen Levels
3.2.6. Effect of d-Allulose on AMPK, ACC, and PGC-1α in Skeletal Muscle
3.3. Effect of Short-Term d-Allulose Administration on MAS, Blood Glucose, and Blood Lactate Levels (Experiment 3)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Nimptsch, K.; Konigorski, S.; Pischon, T. Diagnosis of obesity and use of obesity biomarkers in science and clinical medicine. Metabolism 2019, 92, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, J.P.; Sacks, J.; Nieuwoudt, S. The essential role of exercise in the management of type 2 diabetes. Cleve Clin. J. Med. 2017, 84 (Suppl. 1), S15–S21. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Matsuo, T.; Tsukamoto, I.; Toyoda, Y.; Ogawa, M.; Nagata, Y.; Tokuda, M. Rare sugar d-allulose: Potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2015, 155, 49–59. [Google Scholar] [CrossRef]
- Mu, W.M.; Zhang, W.L.; Feng, Y.H.; Jiang, B.; Zhou, L. Recent advances on applications and biotechnological production of d-psicose. Appl. Microbiol. Biot. 2012, 94, 1461–1467. [Google Scholar] [CrossRef]
- Natsume, Y.; Yamada, T.; Iida, T.; Ozaki, N.; Gou, Y.; Oshida, Y.; Koike, T. Investigation of d-allulose effects on high-sucrose diet-induced insulin resistance via hyperinsulinemic-euglycemic clamps in rats. Heliyon 2021, 7, e08013. [Google Scholar] [CrossRef]
- Gou, Y.; Liu, B.Y.; Cheng, M.Y.; Yamada, T.; Iida, T.; Wang, S.X.; Banno, R.; Koike, T. d-Allulose Ameliorates Skeletal Muscle Insulin Resistance in High-Fat Diet-Fed Rats. Molecules 2021, 26, 6310. [Google Scholar] [CrossRef]
- Shintani, T.; Yamada, T.; Hayashi, N.; Iida, T.; Nagata, Y.; Ozaki, N.; Toyoda, Y. Rare Sugar Syrup Containing d-Allulose but Not High-Fructose Corn Syrup Maintains Glucose Tolerance and Insulin Sensitivity Partly via Hepatic Glucokinase Translocation in Wistar Rats. J. Agric. Food Chem. 2017, 65, 2888–2894. [Google Scholar] [CrossRef]
- Hossain, M.A.; Kitagaki, S.; Nakano, D.; Nishiyama, A.; Funamoto, Y.; Matsunaga, T.; Tsukamoto, I.; Yamaguchi, F.; Kamitori, K.; Dong, Y.; et al. Rare sugar d-psicose improves insulin sensitivity and glucose tolerance in type 2 diabetes Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Biochem. Biophys. Res. Commun. 2011, 405, 7–12. [Google Scholar] [CrossRef]
- Lee, D.; Han, Y.; Kwon, E.Y.; Choi, M.S. d-allulose Ameliorates Metabolic Dysfunction in C57BL/KsJ-db/db Mice. Molecules 2020, 25, 3656. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Kwon, E.Y.; Choi, M.S. Anti-Diabetic Effects of Allulose in Diet-Induced Obese Mice via Regulation of mRNA Expression and Alteration of the Microbiome Composition. Nutrients 2020, 12, 2113. [Google Scholar] [CrossRef]
- Hayashi, N.; Iida, T.; Yamada, T.; Okuma, K.; Takehara, I.; Yamamoto, T.; Yamada, T.; Tokuda, M. Study on the postprandial blood glucose suppression effect of d-psicose in borderline diabetes and the safety of long-term ingestion by normal human subjects. Biosci. Biotechnol. Biochem. 2010, 74, 510–519. [Google Scholar] [CrossRef] [Green Version]
- Iida, T.; Yamada, T.; Hayashi, N.; Okuma, K.; Izumori, K.; Ishii, R.; Matsuo, T. Reduction of abdominal fat accumulation in rats by 8-week ingestion of a newly developed sweetener made from high fructose corn syrup. Food Chem. 2013, 138, 781–785. [Google Scholar] [CrossRef]
- Ochiai, M.; Nakanishi, Y.; Yamada, T.; Iida, T.; Matsuo, T. Inhibition by dietary d-psicose of body fat accumulation in adult rats fed a high-sucrose diet. Biosci. Biotechnol. Biochem. 2013, 77, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.R.; Kwon, E.Y.; Kim, H.J.; Choi, M.S. Role of Synbiotics Containing d-Allulose in the Alteration of Body Fat and Hepatic Lipids in Diet-Induced Obese Mice. Nutrients 2018, 10, 1797. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, N.; Yamada, T.; Takamine, S.; Iida, T.; Okuma, K.; Tokuda, M. Weight reducing effect and safety evaluation of rare sugar syrup by a randomized double-blind.; parallel-group study in human. J. Funct. Foods 2014, 11, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Kwon, E.Y.; Yu, M.K.; Lee, S.J.; Kim, H.J.; Kim, S.B.; Kim, Y.H.; Choi, M.S. A Preliminary Study for Evaluating the Dose-Dependent Effect of d-Allulose for Fat Mass Reduction in Adult Humans: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2018, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.; Spriet, L.L. Exercise Metabolism: Fuels for the Fire. Cold Spring Harb. Perspect. Med. 2018, 8, a029744. [Google Scholar] [CrossRef]
- Hearris, M.A.; Hammond, K.M.; Fell, J.M.; Morton, J.P. Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients 2018, 10, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, E.R.; Allen, D.G. Effects of reduced muscle glycogen concentration on force; Ca2+ release and contractile protein function in intact mouse skeletal muscle. J. Physiol. 1997, 498 Pt 1, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Taniguchi, H. Fat max as an index of aerobic exercise performance in mice during uphill running. PLoS ONE 2018, 13, e0193470. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.J.; Spinelli, J.B.; Zaganjor, E.; Wong, S.J.; German, N.J.; Randall, E.C.; Dean, A.; Clermont, A.; Paulo, J.A.; Garcia, D.; et al. PHD3 Loss Promotes Exercise Capacity and Fat Oxidation in Skeletal Muscle. Cell Metab. 2020, 32, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T. Effects of daily intake of the rare sugar d-psicose on liver and muscle glycogen repletion with d-fructose administration after exhaustive swimming. BioTechnology 2011, 5, 290–292. [Google Scholar]
- Nagata, Y.; Kanasaki, A.; Tamaru, S.; Tanaka, K. d-psicose, an epimer of d-fructose. favorably alters lipid metabolism in Sprague-Dawley rats. J. Agric. Food Chem. 2015, 63, 3168–3176. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kanasaki, A.; Hayashi, N.; Yamada, T.; Iida, T.; Nagata, Y.; Okuma, K. d-Allulose enhances postprandial fat oxidation in healthy humans. Nutrition 2017, 43–44, 16–20. [Google Scholar] [CrossRef]
- Goh, J.; Ladiges, W. Voluntary Wheel Running in Mice. Curr. Protoc. Mouse Biol. 2015, 5, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Pauly, M.; Assense, A.; Rondon, A.; Thomas, A.; Dubouchaud, H.; Freyssenet, D.; Benoit, H.; Castells, J.; Flore, P. High intensity aerobic exercise training improves chronic intermittent hypoxia-induced insulin resistance without basal autophagy modulation. Sci. Rep. 2017, 7, 43663. [Google Scholar] [CrossRef] [Green Version]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. J. Cachexia Sarcopeni. 2020, 11, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Tappe-Theodor, A.; King, T.; Morgan, M.M. Pros and Cons of Clinically Relevant Methods to Assess Pain in Rodents. Neurosci. Biobehav. Rev. 2019, 100, 335–343. [Google Scholar] [CrossRef]
- Li, P.; Koike, T.; Qin, B.; Kubota, M.; Kawata, Y.; Jia, Y.J.; Oshida, Y. A high-fructose diet impairs Akt and PKCzeta phosphorylation and GLUT4 translocation in rat skeletal muscle. Horm. Metab. Res. 2008, 40, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Vigh-Larsen, J.F.; Ortenblad, N.; Spriet, L.L.; Overgaard, K.; Mohr, M. Muscle Glycogen Metabolism and High-Intensity Exercise Performance: A Narrative Review. Sports Med. 2021, 51, 1855–1874. [Google Scholar] [CrossRef]
- Kasuga, M.; Ogawa, W.; Ohara, T. Tissue glycogen content and glucose intolerance. J. Clin. Investig. 2003, 111, 1282–1284. [Google Scholar] [CrossRef]
- Pederson, B.A.; Cope, C.R.; Schroeder, J.M.; Smith, M.W.; Irimia, J.M.; Thurberg, B.L.; DePaoli-Roach, A.A.; Roach, P.J. Exercise capacity of mice genetically lacking muscle glycogen synthase: In mice; muscle glycogen is not essential for exercise. J. Biol. Chem. 2005, 280, 17260–17265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Soldado, I.; Guinovart, J.J.; Duran, J. Increased liver glycogen levels enhance exercise capacity in mice. J. Biol. Chem. 2021, 297, 100976. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, H.M.; Lally, J.S.; Galic, S.; Thomas, M.; Azizi, P.D.; Fullerton, M.D.; Smith, B.K.; Pulinilkunnil, T.; Chen, J.P.; Samaan, M.C.; et al. AMPK phosphorylation of ACC2 is required for skeletal muscle fatty acid oxidation and insulin sensitivity in mice. Diabetologia 2014, 57, 1693–1702. [Google Scholar] [CrossRef]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcinko, K.; Steinberg, G.R. The role of AMPK in controlling metabolism and mitochondrial biogenesis during exercise. Exp. Physiol. 2014, 99, 1581–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, P.; Mendes, S.V.D.; Leal-Cardoso, J.H.; Ceccatto, V.M. Anaerobic threshold employed on exercise training prescription and performance assessment for laboratory rodents: A short review. Life Sci. 2016, 151, 1–6. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Jain, S.S.; McFarlan, J.T.; Snook, L.A.; Chabowski, A.; Bonen, A. Exercise- and training-induced upregulation of skeletal muscle fatty acid oxidation are not solely dependent on mitochondrial machinery and biogenesis. J. Physiol. 2013, 591, 4415–4426. [Google Scholar] [CrossRef]
- Green, H.J.; Helyar, R.; Ball-Burnett, M.; Kowalchuk, N.; Symon, S.; Farrance, B. Metabolic adaptations to training precede changes in muscle mitochondrial capacity. J. Appl. Physiol. 1992, 72, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, Y.; Sendo, M.; Dezaki, K.; Hira, T.; Sato, T.; Nakata, M.; Goswami, C.; Aoki, R.; Arai, T.; Kumari, P.; et al. GLP-1 release and vagal afferent activation mediate the beneficial metabolic and chronotherapeutic effects of d-allulose. Nat. Commun. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, M.; Onishi, K.; Yamada, T.; Iida, T.; Matsuo, T. d-psicose increases energy expenditure and decreases body fat accumulation in rats fed a high-sucrose diet. Int. J. Food Sci. Nutr. 2014, 65, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Shintani, H.; Shintani, T.; Ashida, H.; Sato, M. Calorie Restriction Mimetics: Upstream-Type Compounds for Modulating Glucose Metabolism. Nutrients 2018, 10, 1821. [Google Scholar] [CrossRef] [Green Version]
- Hofer, S.J.; Davinelli, S.; Bergmann, M.; Scapagnini, G.; Madeo, F. Caloric Restriction Mimetics in Nutrition and Clinical Trials. Front. Nutr. 2021, 6, 717343. [Google Scholar] [CrossRef]
- Ingram, D.K.; Roth, G.S. Glycolytic inhibition: An effective strategy for developing calorie restriction mimetics. Geroscience 2021, 43, 1159–1169. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | C2 (n = 6) | A2 (n = 6) | E2 (n = 6) | AE2 (n = 7) |
|---|---|---|---|---|
| Initial body weight (g) | 26.0 ± 0.4 | 25.6 ± 0.3 | 26.0 ± 0.2 | 25.9 ± 0.4 |
| Final body weight (g) | 30.4 ± 0.9 | 27.5 ± 0.4 # | 27.5 ± 0.5 # | 26.8 ± 0.4 # |
| Food consumption (g/day) | 4.7 ± 0.2 | 3.8 ± 0.2 # | 4.3 ± 0.2 | 4.1 ± 0.2 |
| Liver (g) | 1.114 ± 0.038 | 1.203 ± 0.024 | 1.075 ± 0.059 | 1.070 ± 0.014 |
| Gastrocnemius (g) | 0.242 ± 0.015 | 0.253 ± 0.006 | 0.285 ± 0.014 | 0.253 ± 0.006 |
| Plantaris (g) | 0.031 ± 0.002 | 0.039 ± 0.004 | 0.033 ± 0.003 | 0.034 ± 0.001 |
| Soleus (g) | 0.021 ± 0.004 | 0.030 ± 0.009 | 0.025 ± 0.006 | 0.016 ± 0.001 |
| Tibialis anterior (g) | 0.089 ± 0.003 | 0.112 ± 0.020 | 0.112 ± 0.013 | 0.097 ± 0.021 |
| Extensor digitorum longus (g) | 0.063 ± 0.013 | 0.045 ± 0.005 | 0.051 ± 0.011 | 0.045 ± 0.003 |
| Epididymal fat (g) | 1.087 ± 0.051 | 0.656 ± 0.035 #, * | 0.512 ± 0.034 #, * | 0.300 ± 0.032 # |
| Perirenal fat (g) | 0.357 ± 0.016 | 0.102 ± 0.017 #, * | 0.106 ± 0.023 #, * | 0.033 ± 0.006 # |
| Mesenteric fat (g) | 0.600 ± 0.013 | 0.341 ± 0.012 #, * | 0.365 ± 0.026 #, * | 0.237 ± 0.039 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Gou, Y.; Tsuzuki, T.; Yamada, T.; Iida, T.; Wang, S.; Banno, R.; Toyoda, Y.; Koike, T. d-Allulose Improves Endurance and Recovery from Exhaustion in Male C57BL/6J Mice. Nutrients 2022, 14, 404. https://doi.org/10.3390/nu14030404
Liu B, Gou Y, Tsuzuki T, Yamada T, Iida T, Wang S, Banno R, Toyoda Y, Koike T. d-Allulose Improves Endurance and Recovery from Exhaustion in Male C57BL/6J Mice. Nutrients. 2022; 14(3):404. https://doi.org/10.3390/nu14030404
Chicago/Turabian StyleLiu, Bingyang, Yang Gou, Takamasa Tsuzuki, Takako Yamada, Tetsuo Iida, Sixian Wang, Ryoichi Banno, Yukiyasu Toyoda, and Teruhiko Koike. 2022. "d-Allulose Improves Endurance and Recovery from Exhaustion in Male C57BL/6J Mice" Nutrients 14, no. 3: 404. https://doi.org/10.3390/nu14030404