
Plant-Based, Antioxidant-Rich Snacks Elevate Plasma Antioxidant Ability and Alter Gut Bacterial Composition in Older Adults

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
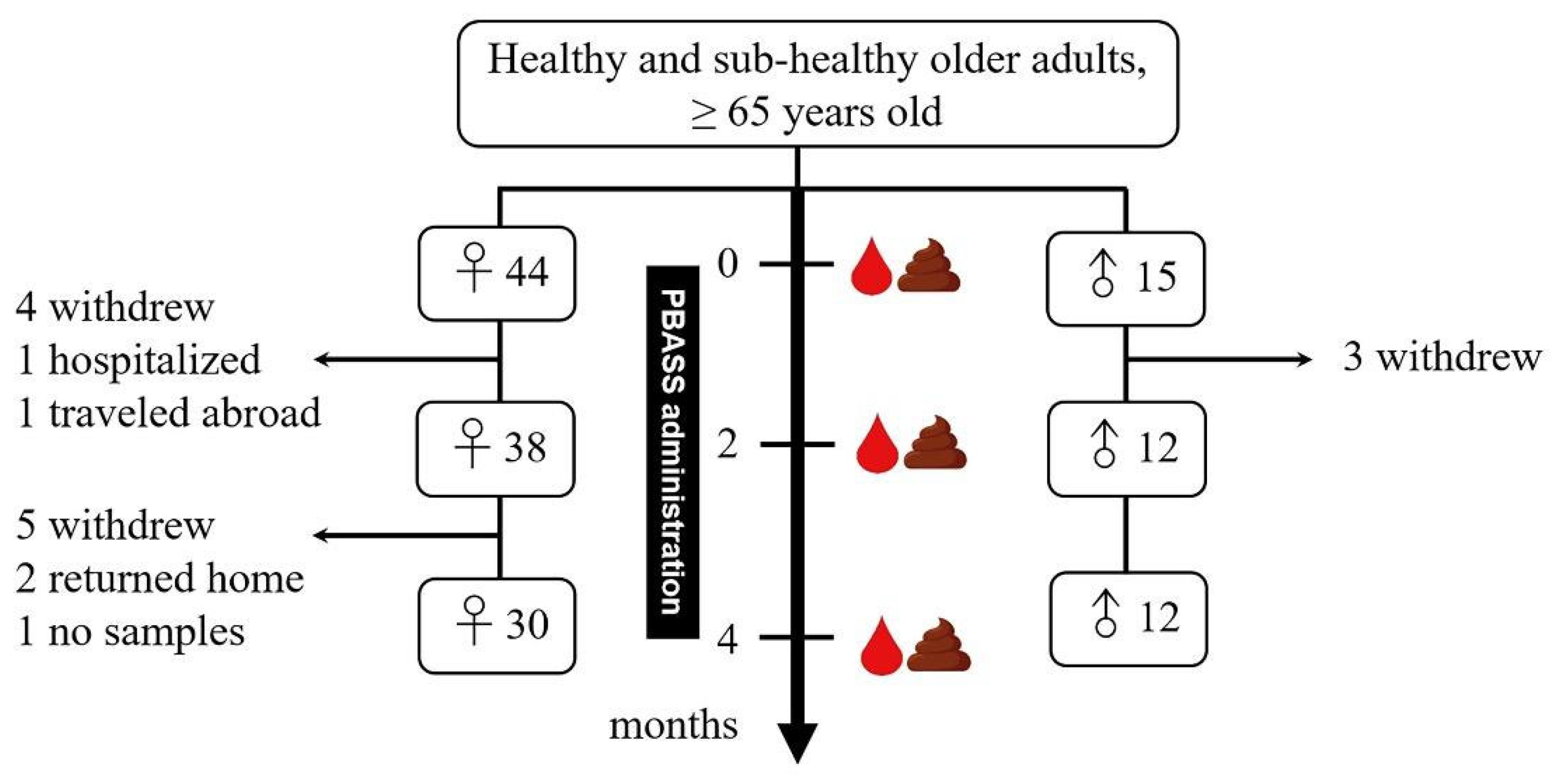
2.1. Study Participants and Ethics
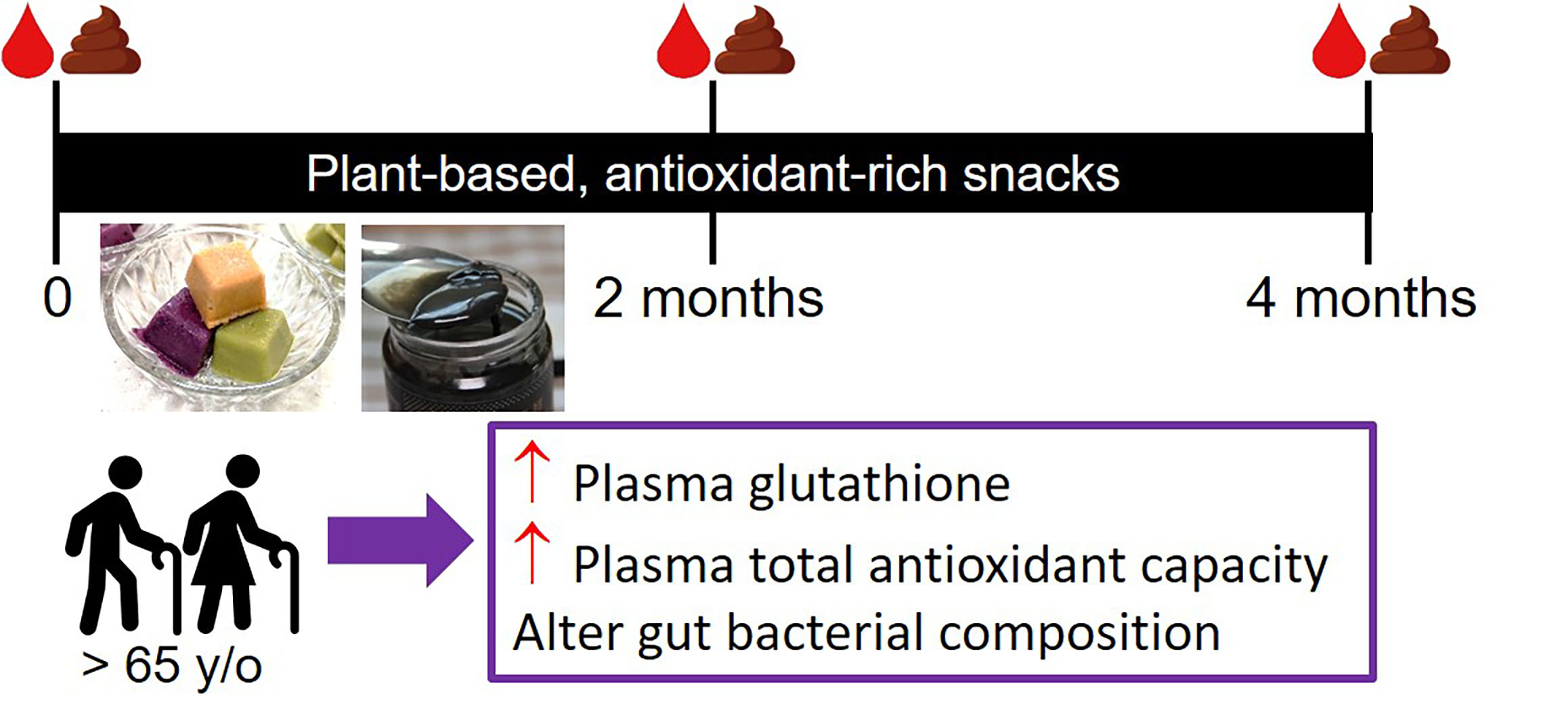
2.2. Study Design
2.3. Composition of Plant-Based Antioxidant Smoothies and Sesame Seed Snacks
2.4. Serological and Biochemical Analyses
2.5. Indicators of Oxidative Status in the Plasma and Erythrocytes
2.6. Short Chain Fatty Acids in the Feces
2.7. Gut Microbiota Composition
2.8. Statistical Analyses
3. Results
3.1. Anthropometric Measurements
3.2. Complete Blood Counts and Serum Biochemical Parameters
3.3. Oxidative Status in the Plasma
3.4. Oxidative Status in the Erythrocytes
3.5. Short-Chain Fatty Acid Levels in the Feces
3.6. Fecal Bacterial Composition
3.7. Power and Effect Size Calculation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Almeida, A.J.P.O.; Rezende, M.S.d.A.; Dantas, S.H.; Silva, S.d.L.; de Oliveira, J.C.P.L.; de Azevedo, F.d.L.A.A.; Alves, R.M.F.R.; de Menezes, G.M.S.; dos Santos, P.F.; Goncalves, T.A.F.; et al. Unveiling the Role of Inflammation and Oxidative Stress on Age-Related Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 1954398. [Google Scholar] [CrossRef]
- Population Projections for the R.O.C. (Taiwan): 2020~2070. Available online: https://www.ndc.gov.tw/en/cp.aspx?n=2E5DCB04C64512CC&s=002ABF0E676F4DB5 (accessed on 28 July 2021).
- Ageing: Healthy Ageing and Functional Ability. Available online: https://www.who.int/westernpacific/news/q-a-detail/ageing-healthy-ageing-and-functional-ability (accessed on 28 July 2021).
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. BioMed Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narmaki, E.; Siassi, F.; Fariba, K.; Qorbani, M.; Shiraseb, F.; Ataie-Jafari, A.; Sotoudeh, G. Dietary diversity as a proxy measure of blood antioxidant status in women. Nutrition 2015, 31, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Tian, G.; Xue, H.; Zhang, X.; Zhao, Y.; Cheng, G. Higher adherence to the ‘vegetable-rich’ dietary pattern is related to longer telomere length in women. Clin. Nutr. 2018, 37, 1232–1237. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Cichero, J.A.Y. Age-Related Changes to Eating and Swallowing Impact Frailty: Aspiration, Choking Risk, Modified Food Texture and Autonomy of Choice. Geriatrics 2018, 3, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Meij, B.S.; Wijnhoven, H.A.H.; Lee, J.S.; Houston, D.K.; Hue, T.; Harris, T.B.; Kritchevsky, S.B.; Newman, A.B.; Visser, M. Poor Appetite and Dietary Intake in Community-Dwelling Older Adults. J. Am. Geriatr. Soc. 2017, 65, 2190–2197. [Google Scholar] [CrossRef] [Green Version]
- Shikany, J.M.; Demmer, R.T.; Johnson, A.J.; Fino, N.F.; Meyer, K.; Ensrud, K.E.; Lane, N.E.; Orwoll, E.S.; Kado, D.M.; Zmuda, J.M.; et al. Association of dietary patterns with the gut microbiota in older, community-dwelling men. Am. J. Clin. Nutr. 2019, 110, 1003–1014. [Google Scholar] [CrossRef]
- Ott, A.; Senger, M.; Lotzbeyer, T.; Gefeller, O.; Sieber, C.C.; Volkert, D. Effects of a Texture-Modified, Enriched, and Reshaped Diet on Dietary Intake and Body Weight of Nursing Home Residents with Chewing and/or Swallowing Problems: An Enable Study. J. Nutr. Gerontol. Geriatr. 2019, 38, 361–376. [Google Scholar] [CrossRef]
- Xu, T.; Zhu, G.; Han, S. Prevalence of Suboptimal Health Status and the Relationships between Suboptimal Health Status and Lifestyle Factors among Chinese Adults Using a Multi-Level Generalized Estimating Equation Model. Int. J. Environ. Res. Public Health 2020, 17, 763. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Xie, F.; Yan, S.; Hu, X.; Jin, B.; Wang, J.; Wu, J.; Yin, D.; Xie, Q. Subhealth: Definition, criteria for diagnosis and potential prevalence in the central region of China. BMC Public Health 2013, 13, 446. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Y. The Effect of Anti-Oxidation Dietary Nutrition Education on the Dietary Nutrition Knowledge, Attitudes, Behaviors and Compliance of Elderly in Institution. MSD Thesis, Fu Jen Catholic University, New Taipei City, Taiwan, 2019. [Google Scholar]
- Lo, H.C.; Hsu, T.H.; Lee, C.H. Extracellular Polysaccharopeptides from Fermented Turkey Tail Medicinal Mushroom, Trametes versicolor (Agaricomycetes), Mitigate Oxidative Stress, Hyperglycemia, and Hyperlipidemia in Rats with Type 2 Diabetes Mellitus. Int. J. Med. Mushrooms 2020, 22, 417–429. [Google Scholar] [CrossRef]
- De Baere, S.; Eeckhaut, V.; Steppe, M.; De Maesschalck, C.; De Backer, P.; Van Immerseel, F.; Croubels, S. Development of a HPLC-UV method for the quantitative determination of four short-chain fatty acids and lactic acid produced by intestinal bacteria during in vitro fermentation. J. Pharm. Biomed. Anal. 2013, 80, 107–115. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Salzberg, S.L. Ultrafast and accurate 16S rRNA microbial community analysis using Kraken 2. Microbiome 2020, 8, 124. [Google Scholar] [CrossRef]
- Foster, Z.S.; Sharpton, T.J.; Grunwald, N.J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Myhrstad, M.C.W.; Tunsjo, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation-Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Lin, X.; Abbasi, A.M.; Zheng, B. Phytochemical Contents and Antioxidant and Antiproliferative Activities of Selected Black and White Sesame Seeds. BioMed Res. Int. 2016, 2016, 8495630. [Google Scholar] [CrossRef] [Green Version]
- Tonin, F.S.; Steimbach, L.M.; Wiens, A.; Perlin, C.M.; Pontarolo, R. Impact of Natural Juice Consumption on Plasma Antioxidant Status: A Systematic Review and Meta-Analysis. Molecules 2015, 20, 22146–22156. [Google Scholar] [CrossRef] [Green Version]
- Rink, S.M.; Mendola, P.; Mumford, S.L.; Poudrier, J.K.; Browne, R.W.; Wactawski-Wende, J.; Perkins, N.J.; Schisterman, E.F. Self-report of fruit and vegetable intake that meets the 5 a day recommendation is associated with reduced levels of oxidative stress biomarkers and increased levels of antioxidant defense in premenopausal women. J. Acad. Nutr. Diet. 2013, 113, 776–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davinelli, S.; Maes, M.; Corbi, G.; Zarrelli, A.; Willcox, D.C.; Scapagnini, G. Dietary phytochemicals and neuro-inflammaging: From mechanistic insights to translational challenges. Immun. Ageing 2016, 13, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partula, V.; Mondot, S.; Torres, M.J.; Kesse-Guyot, E.; Deschasaux, M.; Assmann, K.; Latino-Martel, P.; Buscail, C.; Julia, C.; Galan, P.; et al. Associations between usual diet and gut microbiota composition: Results from the Milieu Interieur cross-sectional study. Am. J. Clin. Nutr. 2019, 109, 1472–1483. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wu, X.; Qiu, L.; Wang, D.; Gan, M.; Chen, X.; Wei, H.; Xu, F. Analysis of the intestinal microbial community structure of healthy and long-living elderly residents in Gaotian Village of Liuyang City. Appl. Microbiol. Biotechnol. 2015, 99, 9085–9095. [Google Scholar] [CrossRef]
- Tian, R.; Ning, D.; He, Z.; Zhang, P.; Spencer, S.J.; Gao, S.; Shi, W.; Wu, L.; Zhang, Y.; Yang, Y.; et al. Small and mighty: Adaptation of superphylum Patescibacteria to groundwater environment drives their genome simplicity. Microbiome 2020, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Zhu, J.; Yang, T.; Guo, M.; Li, Q.; Chen, J.; Li, T. The Gut Microbiota and Associated Metabolites Are Altered in Sleep Disorder of Children With Autism Spectrum Disorders. Front. Psychiatry 2020, 11, 855. [Google Scholar] [CrossRef]
- Cherbuy, C.; Bellet, D.; Robert, V.; Mayeur, C.; Schwiertz, A.; Langella, P. Modulation of the Caecal Gut Microbiota of Mice by Dietary Supplement Containing Resistant Starch: Impact Is Donor-Dependent. Front. Microbiol. 2019, 10, 1234. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| Body height (cm) | 157.7 ± 7.8 | 158.4 ± 7.5 | 157.6 ± 7.2 | 0.815 | 0.22 ± 1.63 | −0.10 ± 0.78 | 0.153 |
| Body weight (kg) | 59.9 ± 8.8 b | 61.1 ± 9.5 a | 60.5 ± 9.7 ab | 0.028 | 0.33 ± 2.84 | −0.05 ± 1.44 | 0.219 |
| BMI (kg/m2) | 24.0 ± 2.8 | 24.3 ± 3.1 | 24.3 ± 3.2 | 0.250 | 0.00 ± 0.78 | −0.16 ± 0.55 | 0.440 |
| Waist circumference (cm) | 85.4 ± 9.0 | 86.3 ± 10.0 | 85.8 ± 11.0 | 0.741 | −1.75 ± 2.90 | −4.31 ± 4.41 | 0.830 |
| Hip circumference (cm) | 96.7 ± 5.1 | 97.9 ± 5.5 | 97.7 ± 11.6 | 0.132 | −0.04 ± 2.15 | −6.07 ± 4.44 | 0.480 |
| Waist-to-hip ratio | 0.88 ± 0.07 | 0.88 ± 0.08 | 0.88 ± 0.07 | 0.939 | −0.03 ± 0.02 | −0.01 ± 0.04 | 0.264 |
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| RBC (106/mL) | 4.26 ± 0.45 a | 4.26 ± 0.50 a | 4.17 ± 0.47 b | 0.004 | 0.00 ± 0.18 | −0.09 ± 0.16 | 0.025 |
| Hemoglobin (g/dL) | 12.97 ± 1.46 ab | 12.88 ± 1.54 b | 13.07 ± 1.51 a | 0.033 | −0.09 ± 0.58 | 0.10 ± 0.59 | 0.140 |
| Hematocrit (%) | 40.47 ± 4.26 a | 41.05 ± 4.80 a | 38.78 ± 4.27 b | <0.001 | 0.58 ± 1.88 | −1.69 ± 1.76 | <0.001 |
| Albumin (g/dL) | 5.03 ± 1.14 a | 4.44 ± 0.38 b | 4.17 ± 0.33 c | <0.001 | −0.59 ± 1.12 | −0.86 ± 1.15 | 0.285 |
| Cholesterol (mg/dL) | 185.1 ± 42.0 | 191.9 ± 46.4 | 191.8 ± 37.5 | 0.334 | 6.8 ± 33.4 | 6.7 ± 30.3 | 0.995 |
| LDL-C (mg/dL) | 102.0 ± 34.7 | 110.3 ± 38.3 | 103.2 ± 30.0 | 0.076 | 8.4 ± 23.6 | 1.3 ± 20.3 | 0.143 |
| HDL-C (mg/dL) | 61.2 ± 12.1 b | 68.4 ± 12.4 a | 61.8 ± 14.8 b | <0.001 | 7.2 ± 7.4 | 0.6 ± 7.1 | <0.001 |
| BUN (mg/dL) | 18.12 ± 6.32 a | 19.00 ± 8.68 a | 16.64 ± 6.01 b | 0.032 | 0.88 ± 4.31 | −1.48 ± 4.10 | 0.012 |
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| GSH (mg/mL) | 212.1 ± 28.9 c | 334.4 ± 41.2 b | 364.0 ± 35.1 a | <0.001 | 122.3 ± 44.0 | 151.9 ± 38.2 | 0.002 |
| GSSG (mg/mL) | 36.4 ± 18.4 a | 23.0 ± 9.4 b | 34.7 ± 8.1 a | <0.001 | −13.5 ± 19.9 | −1.7 ± 19.9 | 0.008 |
| GSH/GSSG | 6.66 ± 2.01 c | 16.48 ± 5.64 a | 11.27 ± 4.00 b | <0.001 | 9.82 ± 5.69 | 4.61 ± 4.70 | <0.001 |
| TSH (nmol/mL) | 226.2 ± 46.5 ab | 222.4 ± 24.0 b | 247.2 ± 45.2 a | 0.015 | −3.9 ± 46.7 | 21.0 ± 62.5 | 0.042 |
| NPSH (nmol/mL) | 29.9 ± 3.9 | 28.1 ± 6.3 | 29.2 ± 1.7 | 0.439 | −1.8 ± 6.8 | −0.7 ± 4.3 | 0.390 |
| PBSH (nmol/mL) | 197.1 ± 45.9 ab | 194.3 ± 24.0 b | 218.0 ± 45.3 a | 0.019 | −2.8 ± 46.3 | 20.9 ± 61.9 | 0.050 |
| TAC (nmol/mL) | 8007 ± 555 c | 8443 ± 679 b | 9226 ± 691 a | <0.001 | 436 ± 554 | 1219 ± 587 | <0.001 |
| TBARS (nmol/mL) | 7.17 ± 1.40 | 6.92 ± 1.38 | 6.93 ± 1.48 | 0.325 | −0.25 ± 1.52 | −0.23 ± 1.54 | 0.962 |
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| GSH (mg/109 cell) | 464.1 ± 98.0 a | 197.8 ± 28.9 b | 181.6 ± 22.4 c | <0.001 | −266.3 ± 86.5 | −282.5 ± 98.2 | 0.425 |
| GSSG (mg/109 cell) | 140.3 ± 85.8 a | 65.2 ± 11.9 b | 69.3 ± 23.9 b | <0.001 | −75.1 ± 85.7 | −71.1 ± 85.8 | 0.830 |
| GSH/GSSG | 4.21 ± 0.26 a | 3.09 ± 0.07 b | 2.85 ± 0.13 b | <0.001 | −1.13 ± 0.29 | −1.37 ± 0.29 | 0.559 |
| TSH (nmol/109 cell) | 2.18 ± 0.57 b | 2.35 ± 0.54 b | 2.69 ± 0.55 a | 0.004 | 0.17 ± 0.77 | 0.50 ± 0.83 | 0.061 |
| NPSH (nmol/109 cell) | 0.14 ± 0.036 | 0.12 ± 0.03 | 0.13 ± 0.03 | 0.064 | −0.01 ± 0.03 | −0.01 ± 0.03 | 0.591 |
| PBSH (nmol/109 cell) | 2.04 ± 0.57 b | 2.23 ± 0.53 a | 2.56 ± 0.55 a | 0.003 | 0.18 ± 0.78 | 0.51 ± 0.83 | 0.065 |
| TAC (nmol/109 cell) | 2327 ± 355 | 2357 ± 361 | 2292 ± 457 | 0.526 | 30 ± 324 | −35 ± 502 | 0.487 |
| TBARS (nmol/109 cell) | 48.54 ± 13.73 | 47.48 ± 11.51 | 49.27 ± 10.43 | 0.494 | −1.06 ± 11.16 | 0.73 ± 10.76 | 0.459 |
| SOD | 45.56 ± 8.62 a | 36.20 ± 6.45 b | 35.24 ± 5.41 b | <0.001 | −9.35 ± 6.74 | −10.32 ± 7.31 | 0.530 |
| GPx | 172.4 ± 31.5 a | 168.3 ± 34.7 a | 149.2 ± 24.9 b | <0.001 | −4.0 ± 24.8 | −23.1 ± 20.7 | <0.001 |
| Catalase | 0.72 ± 0.62 | 0.55 ± 0.33 | 0.89 ± 1.02 | 0.052 | −0.16 ± 0.68 | 0.17 ± 1.22 | 0.128 |
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| Acetic acid (μmol/g) | 40.98 ± 16.83 | 38.59 ± 17.86 | 30.10 ± 17.26 | 0.179 | −2.39 ± 13.00 | −4.89 ± 12.92. | 0.968 |
| Propionic acid (μmol/g) | 40.88 ± 19.21 | 42.26 ± 20.48 | 38.89 ± 20.85 | 0.218 | 1.37 ± 17.72 | −2.00 ± 21.14 | 0.275 |
| Butyric acid (μmol/g) | 36.06 ± 18.20 | 31.61 ± 16.76 | 34.21 ± 19.06 | 0.525 | −4.46 ± 21.9 | −1.85 ± 20.71 | 0.711 |
| Observed OTUs | 319.5 ± 98.4 a | 243.4 ± 71.1 b | 252.4 ± 69.4 b | <0.001 | −76.0 ± 14.4 | −67.1 ± 17.4 | 0.254 |
| Chao1 | 391.1 ± 112.5 a | 301.2 ± 85.4 b | 310.2 ± 77.9 b | <0.001 | −89.9 ± 17.0 | −80.9 ± 19.4 | 0.412 |
| ACE | 387.9 ± 111.4 a | 294.6 ± 78.9 b | 308.8 ± 77.9b | <0.001 | −93.3 ± 16.3 | −79.1 ± 19.8 | 0.225 |
| Shannon | 5.09 ± 0.74 | 4.98 ± 0.68 | 5.00 ± 0.66 | 0.298 | −0.11 ± 0.13 | −0.08 ± 0.12 | 0.596 |
| Simpson | 0.92 ± 0.06 | 0.92 ± 0.05 | 0.93 ± 0.04 | 0.425 | 0.00 ± 0.01 | 0.01 ± 0.01 | 0.596 |
| Parameters | Baseline | 2nd Month | 4th Month | Adjusted p-Value | Δ2mo | Δ4mo | Δp-Value |
|---|---|---|---|---|---|---|---|
| p_Firmicutes (%) | 54.30 ± 20.15 | 55.95 ± 15.63 | 57.59 ± 13.77 | 0.333 | 1.65 ± 19.88 | 3.29 ± 22.36 | 0.730 |
| c_Bacilli | 7.18 ± 9.41 a | 4.20 ± 10.92 b | 4.78 ± 8.38 b | <0.001 | −2.98 ± 11.75 | −2.40 ± 10.79 | 0.818 |
| o_Lactobacillales | 6.68 ± 11.32 | 3.78 ± 8.45 | 5.55 ± 8.84 | 0.451 | −2.90 ± 14.35 | −1.13 ± 14.93 | 0.591 |
| f_Streptococcaceae | 4.12 ± 7.61 | 4.14 ± 6.42 | 1.63 ± 2.49 | 0.234 | 0.02 ± 8.51 | −2.49 ± 8.02 | 0.179 |
| g_Streptococcus | 0.057 ± 0.087 a | 0.018 ± 0.03 b | 0.024 ± 0.039 b | 0.003 | −0.040 ± 0.089 | −0.033 ± 0.095 | 0.765 |
| f_Lactobacillaceae | 1.088 ± 3.215 | 2.364 ± 8.976 | 1.733 ± 6.439 | 0.745 | 1.276 ± 9.511 | 0.644 ± 7.430 | 0.742 |
| g_Lactobacillus | 0.009 ± 0.026 b | 0.023 ± 0.097 a | 0.020 ± 0.056 a | 0.005 | 0.013 ± 0.098 | 0.010 ± 0.056 | 0.869 |
| s_Lactobacillus_salivarius | 0.003 ± 0.012 | 0.013 ± 0.079 | 0.012 ± 0.040 | 0.543 | 0.010 ± 0.078 | 0.009 ± 0.040 | 0.931 |
| s_Lactobacillus_gasseri | 0.002 ± 0.008 | 0.006 ± 0.030 | 0.003 ± 0.011 | 0.231 | 0.004 ± 0.031 | 0.001 ± 0.012 | 0.539 |
| g_Ruminiclostridium_5 | 0.008 ± 0.011 a | 0.004 ± 0.006 b | 0.004 ± 0.012 b | 0.003 | −0.004 ± 0.013 | −0.003 ± 0.017 | 0.905 |
| g_Ruminococcaceae_UCG_014 | 0.016 ± 0.029 | 0.009 ± 0.019 | 0.009 ± 0.019 | 0.165 | −0.007 ± 0.022 | −0.007 ± 0.032 | 0.972 |
| g_Agathobacter | 0.014 ± 0.030 b | 0.017 ± 0.024 b | 0.019 ± 0.028 a | 0.022 | 0.003 ± 0.036 | 0.005 ± 0.039 | 0.854 |
| g_Megasphaera | 0.012 ± 0.026 | 0.020 ± 0.049 | 0.010 ± 0.029 | 0.963 | 0.008 ± 0.056 | −0.002 ± 0.041 | 0.364 |
| p_Bacteroidetes (%) | 21.80 ± 17.26 b | 29.23 ± 15.19 a | 25.01 ± 13.12 ab | 0.031 | 7.43 ± 17.45 | 3.21 ± 18.39 | 0.296 |
| f_Muribaculaceae | 0.470 ± 1.099 | 0.559 ± 1.305 | 0.375 ± 0.857 | 0.698 | 0.089 ± 1.190 | −0.095 ± 1.467 | 0.541 |
| s_Bacteroides_plebeius_DSM_17135 | 0.008 ± 0.016 | 0.009 ± 0.029 | 0.005 ± 0.014 | 0.575 | 0.002 ± 0.029 | −0.002 ± 0.014 | 0.449 |
| s_Bacteroides_thetaiotaomicron | 0.006 ± 0.008 b | 0.015 ± 0.025 a | 0.009 ± 0.012 ab | 0.017 | 0.009 ± 0.024 | 0.004 ± 0.014 | 0.228 |
| s_Bacteroides_coprocola_DSM_17136 | 0.002 ± 0.010 | 0.003 ± 0.009 | 0.005 ± 0.016 | 0.879 | 0.000 ± 0.014 | 0.003 ± 0.019 | 0.560 |
| p_Proteobacteria (%) | 10.16 ± 9.33 | 8.32 ± 9.21 | 9.77 ± 10.38 | 0.160 | −1.84 ± 11.54 | −0.38 ± 11.32 | 0.571 |
| c_Deltaproteobacteria | 0.909 ± 1.460 a | 0.372 ± 0.414 b | 0.413 ± 0.549 b | 0.006 | −0.537 ± 1.520 | −0.497 ± 1.629 | 0.908 |
| o_Desulfovibrionales | 0.488 ± 0.647 | 0.473 ± 0.607 | 0.733 ± 1.399 | 0.756 | −0.015 ± 0.843 | 0.245 ± 1.188 | 0.263 |
| f_Desulfovibrionaceae | 0.733 ± 1.399 | 0.488 ± 0.647 | 0.473 ± 0.607 | 0.745 | −0.245 ± 1.454 | −0.260 ± 1.114 | 0.959 |
| p_Actinobacteria (%) | 3.04 ± 4.30 a | 1.92 ± 4.22 b | 1.69 ± 3.36 b | 0.001 | −1.13 ± 5.65 | −1.36 ± 4.76 | 0.845 |
| c_Actinobacteria | 1.947 ± 3.674 a | 1.255 ± 4.070 b | 0.759 ± 1.671 b | <0.001 | −0.692 ± 5.266 | −1.188 ± 3.776 | 0.629 |
| o_Bifidobacteriales | 1.06 ± 2.83 | 0.85 ± 1.77 | 1.71 ± 4.63 | 0.656 | −0.22 ± 3.25 | 0.65 ± 5.44 | 0.393 |
| f_Bifidobacteriaceae | 1.708 ± 4.630 | 1.063 ± 2.826 | 0.847 ± 1.774 | 0.749 | −0.645 ± 5.683 | −0.862 ± 4.883 | 0.856 |
| p_Fusobacteria (%) | 0.648 ± 2.908 | 0.726 ± 3.284 | 0.934 ± 3.071 | 0.210 | 0.077 ± 4.200 | 0.286 ± 3.060 | 0.619 |
| o_Fusobacteriales | 0.182 ± 0.519 | 0.574 ± 3.255 | 0.316 ± 1.118 | 0.364 | 0.392 ± 3.319 | 0.134 ± 3.154 | 0.620 |
| f_Fusobacteriaceae | 0.093 ± 0.371 | 0.018 ± 0.052 | 0.057 ± 0.326 | 0.104 | −0.075 ± 0.309 | −0.036 ± 0.459 | 0.656 |
| g_Fusobacterium | 0.006 ± 0.028 | 0.007 ± 0.033 | 0.003 ± 0.011 | 0.198 | 0.001 ± 0.041 | −0.003 ± 0.030 | 0.660 |
| p_Patescibacteria (%) | 0.135 ± 0.299 a | 0.025 ± 0.054 b | 0.016 ± 0.020 b | 0.011 | −0.110 ± 0.295 | −0.119 ± 0.303 | 0.899 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-Y.; Lo, H.-C.; Yang, F.L.; Liu, Y.-F.; Wu, W.-M.; Chou, C.-C. Plant-Based, Antioxidant-Rich Snacks Elevate Plasma Antioxidant Ability and Alter Gut Bacterial Composition in Older Adults. Nutrients 2021, 13, 3872. https://doi.org/10.3390/nu13113872
Zhang J-Y, Lo H-C, Yang FL, Liu Y-F, Wu W-M, Chou C-C. Plant-Based, Antioxidant-Rich Snacks Elevate Plasma Antioxidant Ability and Alter Gut Bacterial Composition in Older Adults. Nutrients. 2021; 13(11):3872. https://doi.org/10.3390/nu13113872
Chicago/Turabian StyleZhang, Jing-Yao, Hui-Chen Lo, Feili Lo Yang, Yi-Fang Liu, Wen-Mein Wu, and Chi-Chun Chou. 2021. "Plant-Based, Antioxidant-Rich Snacks Elevate Plasma Antioxidant Ability and Alter Gut Bacterial Composition in Older Adults" Nutrients 13, no. 11: 3872. https://doi.org/10.3390/nu13113872