
Natural Products in the Prevention of Metabolic Diseases: Lessons Learned from the 20th KAST Frontier Scientists Workshop

 ,
,  , , ,
, , ,  , ,
, ,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Inflammation, Oxidative Stress, and Natural Products
2.1. Antioxidant Effect of Triterpenoids and Tocopherols
2.2. Role of Tart Cherry (TC) in the Prevention of Obesity-Related Inflammation and Life Span Extension
2.3. Food-Derived Antioxidants and Lifespan
2.4. Role of Phytochemicals in the Regulation of Mitochondrial Functions under Oxidative Stress
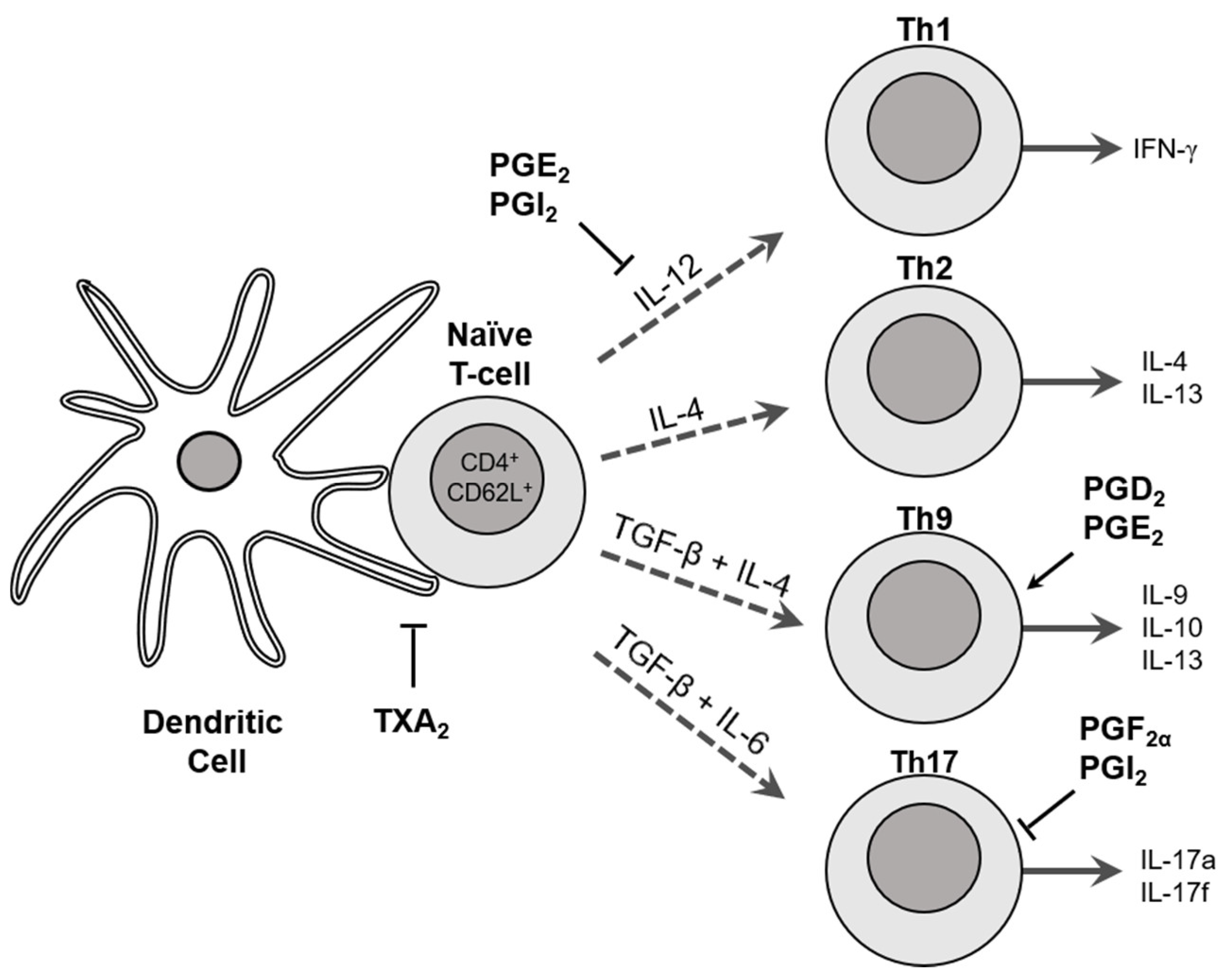
2.5. Role of Cyclooxygenases in T Cell Differentiation and Function
3. Neuroinflammation and Natural Products
3.1. Protective Effects of Phytochemicals on Blood–Brain Barrier Integrity after Ischemic Damage to the Brain
3.2. Fatty Acid Metabolites for the Treatment of Inflammation and Neuropathic Pain
3.3. Role of Passiflora incarnata L. (PI) in Neuroinflammation
4. Antioxidant and Anti-Inflammatory Compounds in Cancer Prevention
4.1. Roles of NAG-1 Activated by Phytochemicals in the Prevention of Inflammation-Mediated Tumorigenesis
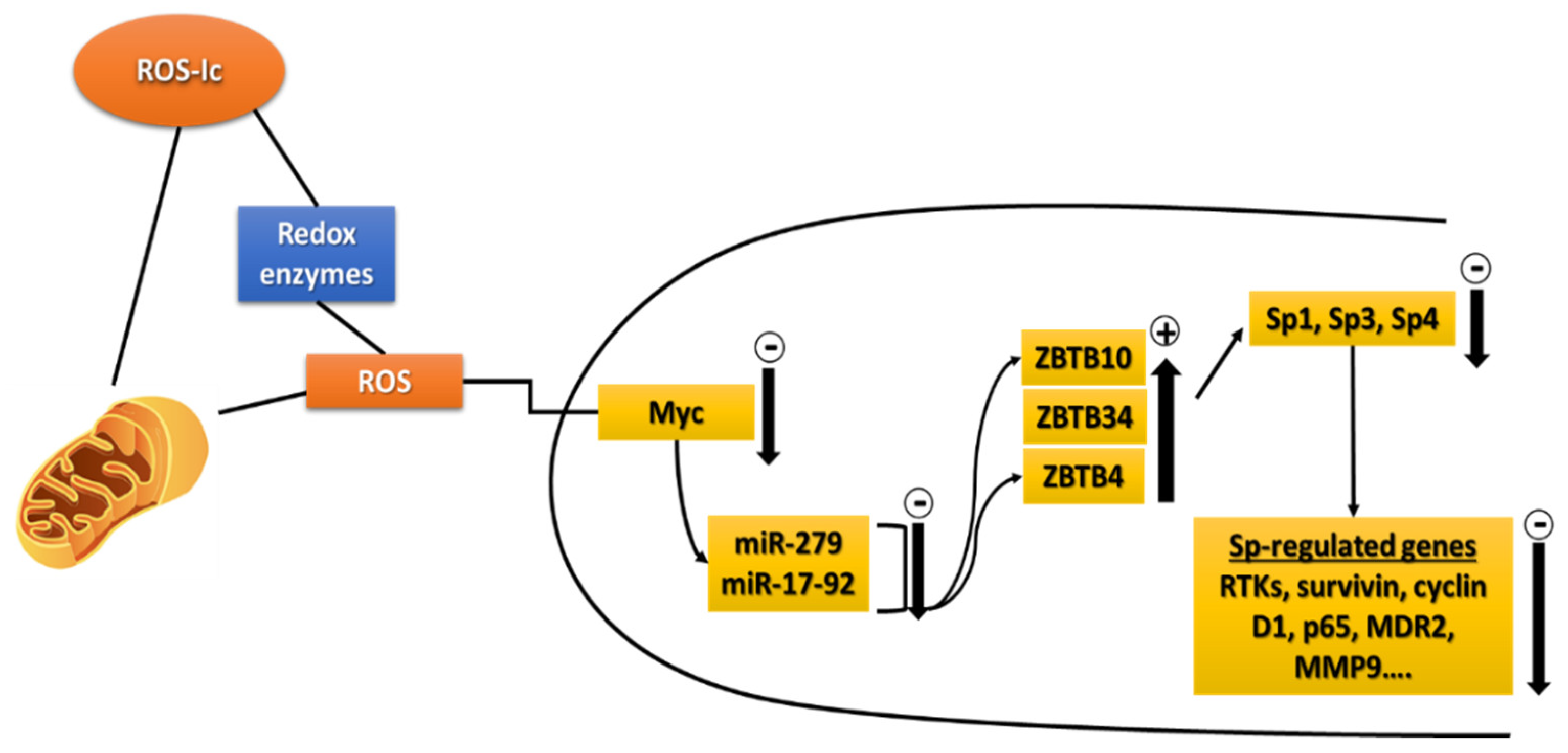
4.2. ROS-Inducing Natural Products as Anticancer Agents

{kind=link}
{kind=link}
| Phytochemical | Cell Line | Dose (μM) | Mechanism of Action | Reference |
|---|---|---|---|---|
| 2′-Hydroxyflavanone | HCT-116 | 5–40 | EGR-1 | [200] |
| 6-Gingerol | HCT-116 | 25–200 | PKCε, GSK-3β | [217] |
| 18α-Glycyrrhetinic acid | DU-145 | 100 | - | [208] |
| Apigenin | HCT-116, LoVo, SW480, HT-29 | 0.1–10 | PKCδ | [201] |
| Berberine | HCT-116, Caco-2, HepG2 | 1–100 | PKCε, GSK-3β, ERK1/2, EGR-1 | [218,219] |
| CAPE | HCT-116 | 1–25 | ATF3 | [203] |
| Capsaicin | HCT-116 | 1–100 | GSK3β, C/EBPβ, ATF3, PKCδ | [204] |
| Damnacanthal | HCT-116, LoVo | 1–100 | ERK, C/EBPβ | [220] |
| Diallyl disulfide | HCT-116 | 4.6–23 | p53 | [221] |
| DIM | HCT-116 | 12.5–50 | ATF3 | [222] |
| Green tea (EGCG/ECG) | HCT-116 | 1–100 | ATF3, EGR-1 | [202,223,224] |
| Genistein | HCT-116, A549 | 25–100 | p53 | [225,226] |
| Formononetin | HCT-116 | 6.25–400 | EGR-1 | [205] |
| Hispidulin | AGS | 6.25–100 | ERK1/2 | [212] |
| Indole-3-carbinol | HCT-116 | 25–100 | - | [222] |
| Isochaihulactone | A549, LNCaP, GBM8401 | 1.25–80 | EGR-1, ERK1/2, JNK, DDIT3 | [209,210,227,228] |
| Isoliquiritigenin | HCT-116 | 2.5–160 | EGR-1 | [205] |
| Platycodon D | U937 | 7.5–15 | EGR-1 | [229] |
| Pinosylvin | HCT-116 | 60 | p53 | [206] |
| Pseudolaric acid B | HT-29 | 1–25 | EGR-1 | [230] |
| Quercetin | HCT-116, Huh7 | 5–40 | EGR-1, p53 | [231] |
| Resveratrol | HCT-116, A549, U2OS, S2-013, CD18 | 10–100 | p53, RNA stability | [232,233] |
| Silibinin | HT-29 | 50–100 | EGR-1, p38 MAPK | [207] |
| Taiwanin A | A549, H460 | 1.25–80 | JNK | [211] |
| Xanthorrhizol | HCT-116 | 25–100 | - | [234] |
5. Identification of Biochemical Targets of Bioactive Compounds
6. Nano-Delivery of Bioactive Compounds in Foods for Disease Prevention
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mesika, R.; Reichmann, D. When safeguarding goes wrong: Impact of oxidative stress on protein homeostasis in health and neurodegenerative disorders. Adv. Protein Chem. Struct. Biol. 2019, 114, 221–264. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative stress: A key modulator in neurodegenerative diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [Green Version]
- Karam, B.S.; Chavez-Moreno, A.; Koh, W.; Akar, J.G.; Akar, F.G. Oxidative stress and inflammation as central mediators of atrial fibrillation in obesity and diabetes. Cardiovasc. Diabetol. 2017, 16, 120. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Karin, M. The fire within: Cell-Autonomous mechanisms in inflammation-driven cancer. Cancer Cell 2019, 35, 714–720. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Kunnumakkara, A.B.; Aggarwal, S.; Aggarwal, B.B. Inflammation, a double-edge sword for cancer and other age-related diseases. Front. Immunol. 2018, 9, 2160. [Google Scholar] [CrossRef]
- Albini, A.; DeCensi, A.; Cavalli, F.; Costa, A. Cancer prevention and interception: A new era for chemopreventive approaches. Clin. Cancer Res. 2016, 22, 4322–4327. [Google Scholar] [CrossRef] [Green Version]
- Ballon-Landa, E.; Parsons, J.K. Nutrition, physical activity, and lifestyle factors in prostate cancer prevention. Curr. Opin. Urol. 2018, 28, 55–61. [Google Scholar] [CrossRef]
- Albini, A.; Bassani, B.; Baci, D.; Dallaglio, K.; Gallazzi, M.; Corradino, P.; Bruno, A.; Noonan, D.M. Nutraceuticals and “repurposed” drugs of phytochemical origin in prevention and interception of chronic degenerative diseases and cancer. Curr. Med. Chem. 2019, 26, 973–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sporn, M.B.; Liby, K.T.; Yore, M.M.; Fu, L.; Lopchuk, J.M.; Gribble, G.W. New synthetic triterpenoids: Potent agents for prevention and treatment of tissue injury caused by inflammatory and oxidative stress. J. Nat. Prod. 2011, 74, 537–545. [Google Scholar] [CrossRef]
- Yang, C.S.; Luo, P.; Zeng, Z.; Wang, H.; Malafa, M.; Suh, N. Vitamin E and cancer prevention: Studies with different forms of tocopherols and tocotrienols. Mol. Carcinog. 2020, 59, 365–389. [Google Scholar] [CrossRef]
- Suh, N.; Wang, Y.; Honda, T.; Gribble, G.W.; Dmitrovsky, E.; Hickey, W.F.; Maue, R.A.; Place, A.E.; Porter, D.M.; Spinella, M.J.; et al. A novel synthetic oleanane triterpenoid, 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid, with potent differentiating, antiproliferative, and anti-inflammatory activity. Cancer Res. 1999, 59, 336–341. [Google Scholar] [PubMed]
- Dinkova-Kostova, A.T.; Liby, K.T.; Stephenson, K.K.; Holtzclaw, W.D.; Gao, X.; Suh, N.; Williams, C.; Risingsong, R.; Honda, T.; Gribble, G.W.; et al. Extremely potent triterpenoid inducers of the phase 2 response: Correlations of protection against oxidant and inflammatory stress. Proc. Natl. Acad. Sci. USA 2005, 102, 4584–4589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, T.; Gribble, G.W.; Suh, N.; Finlay, H.J.; Rounds, B.V.; Bore, L.; Favaloro, F.G., Jr.; Wang, Y.; Sporn, M.B. Novel synthetic oleanane and ursane triterpenoids with various enone functionalities in ring A as inhibitors of nitric oxide production in mouse macrophages. J. Med. Chem. 2000, 43, 1866–1877. [Google Scholar] [CrossRef] [PubMed]
- Liby, K.T.; Sporn, M.B. Synthetic oleanane triterpenoids: Multifunctional drugs with a broad range of applications for prevention and treatment of chronic disease. Pharm. Rev. 2012, 64, 972–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, J.; Picinich, S.C.; Yang, Z.; Zhao, Y.; Suh, N.; Kong, A.N.; Yang, C.S. Cancer-preventive activities of tocopherols and tocotrienols. Carcinogenesis 2010, 31, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Quinn, P.J. Vitamin E and its function in membranes. Prog. Lipid Res. 1999, 38, 309–336. [Google Scholar] [CrossRef]
- Mustacich, D.J.; Bruno, R.S.; Traber, M.G. Vitamin E. Vitam. Horm. 2007, 76, 1–21. [Google Scholar] [CrossRef]
- Niki, E.; Traber, M.G. A history of vitamin E. Ann. Nutr. Metab. 2012, 61, 207–212. [Google Scholar] [CrossRef]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traber, M.G. Vitamin E regulatory mechanisms. Annu. Rev. Nutr. 2007, 27, 347–362. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, J.X.; Wang, H.; Lim, J. Lessons learned from cancer prevention studies with nutrients and non-nutritive dietary constituents. Mol. Nutr. Food Res. 2016, 60, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohe, R.; Traber, M.G. Vitamin E: Function and metabolism. FASEB J. 1999, 13, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Benaksas, E.J.; Bolli, R.; Comp, P.; Grammas, P.; Hamdheydari, L.; Mou, S.; Pye, Q.N.; Stoddard, M.F.; Wallis, G.; et al. New perspectives on vitamin E: Gamma-tocopherol and carboxyelthylhydroxychroman metabolites in biology and medicine. Free Radic. Biol. Med. 2004, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A.; Appelqvist, L.A. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.A.; Thompson, I.M., Jr.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the risk of prostate cancer: The selenium and Vitamin E cancer prevention trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Liu, A.; Lee, M.J.; Wang, H.; Yu, S.; Chi, E.; Reuhl, K.; Suh, N.; Yang, C.S. Delta- and gamma-tocopherols inhibit phIP/DSS-induced colon carcinogenesis by protection against early cellular and DNA damages. Mol. Carcinog. 2017, 56, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Smolarek, A.K.; So, J.Y.; Thomas, P.E.; Lee, H.J.; Paul, S.; Dombrowski, A.; Wang, C.X.; Saw, C.L.; Khor, T.O.; Kong, A.N.; et al. Dietary tocopherols inhibit cell proliferation, regulate expression of ERalpha, PPARgamma, and Nrf2, and decrease serum inflammatory markers during the development of mammary hyperplasia. Mol. Carcinog. 2013, 52, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Smolarek, A.K.; So, J.Y.; Burgess, B.; Kong, A.N.; Reuhl, K.; Lin, Y.; Shih, W.J.; Li, G.; Lee, M.J.; Chen, Y.K.; et al. Dietary administration of delta- and gamma-tocopherol inhibits tumorigenesis in the animal model of estrogen receptor-positive, but not HER-2 breast cancer. Cancer Prev. Res. Phila 2012, 5, 1310–1320. [Google Scholar] [CrossRef] [Green Version]
- Guan, F.; Li, G.; Liu, A.B.; Lee, M.J.; Yang, Z.; Chen, Y.K.; Lin, Y.; Shih, W.; Yang, C.S. Delta- and gamma-tocopherols, but not alpha-tocopherol, inhibit colon carcinogenesis in azoxymethane-treated F344 rats. Cancer Prev. Res. Phila 2012, 5, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Bak, M.J.; Das Gupta, S.; Wahler, J.; Lee, H.J.; Li, X.; Lee, M.J.; Yang, C.S.; Suh, N. Inhibitory effects of gamma- and delta-Tocopherols on Estrogen-Stimulated breast cancer In Vitro and In Vivo. Cancer Prev. Res. Phila 2017, 10, 188–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Gupta, S.; Patel, M.; Wahler, J.; Bak, M.J.; Wall, B.; Lee, M.J.; Lin, Y.; Shih, W.J.; Cai, L.; Yang, C.S.; et al. Differential gene regulation and tumor-inhibitory activities of alpha-, delta-, and gamma-tocopherols in estrogen-mediated mammary carcinogenesis. Cancer Prev. Res. Phila 2017, 10, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Das Gupta, S.; Sae-tan, S.; Wahler, J.; So, J.Y.; Bak, M.J.; Cheng, L.C.; Lee, M.J.; Lin, Y.; Shih, W.J.; Shull, J.D.; et al. Dietary gamma-tocopherol-rich mixture inhibits estrogen-induced mammary tumorigenesis by modulating estrogen metabolism, antioxidant response, and PPARgamma. Cancer Prev. Res. Phila 2015, 8, 807–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Gupta, S.; So, J.Y.; Wall, B.; Wahler, J.; Smolarek, A.K.; Sae-Tan, S.; Soewono, K.Y.; Yu, H.; Lee, M.J.; Thomas, P.E.; et al. Tocopherols inhibit oxidative and nitrosative stress in estrogen-induced early mammary hyperplasia in ACI rats. Mol. Carcinog. 2015, 54, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Christen, S.; Woodall, A.A.; Shigenaga, M.K.; Southwell-Keely, P.T.; Duncan, M.W.; Ames, B.N. Gamma-tocopherol traps mutagenic electrophiles such as NO(X) and complements alpha-tocopherol: Physiological implications. Proc. Natl. Acad. Sci. USA 1997, 94, 3217–3222. [Google Scholar] [CrossRef] [Green Version]
- Hoglen, N.C.; Waller, S.C.; Sipes, I.G.; Liebler, D.C. Reactions of peroxynitrite with gamma-tocopherol. Chem. Res. Toxicol. 1997, 10, 401–407. [Google Scholar] [CrossRef]
- Jiang, Q.; Lykkesfeldt, J.; Shigenaga, M.K.; Shigeno, E.T.; Christen, S.; Ames, B.N. Gamma-tocopherol supplementation inhibits protein nitration and ascorbate oxidation in rats with inflammation. Free Radic. Biol. Med. 2002, 33, 1534–1542. [Google Scholar] [CrossRef]
- Ogawa, Y.; Saito, Y.; Nishio, K.; Yoshida, Y.; Ashida, H.; Niki, E. Gamma-tocopheryl quinone, not alpha-tocopheryl quinone, induces adaptive response through up-regulation of cellular glutathione and cysteine availability via activation of ATF4. Free Radic. Res. 2008, 42, 674–687. [Google Scholar] [CrossRef]
- Jiang, Q.; Ames, B.N. Gamma-tocopherol, but not alpha-tocopherol, decreases proinflammatory eicosanoids and inflammation damage in rats. FASEB J. 2003, 17, 816–822. [Google Scholar] [CrossRef]
- Jiang, Q.; Elson-Schwab, I.; Courtemanche, C.; Ames, B.N. Gamma-tocopherol and its major metabolite, in contrast to alpha-tocopherol, inhibit cyclooxygenase activity in macrophages and epithelial cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11494–11499. [Google Scholar] [CrossRef] [Green Version]
- Reiter, E.; Jiang, Q.; Christen, S. Anti-inflammatory properties of alpha- and gamma-tocopherol. Mol. Asp. Med. 2007, 28, 668–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, K.A.; de Pascual-Tereasa, S.; Needs, P.W.; Bao, Y.P.; O’Brien, N.M.; Williamson, G. Effect of flavonoids and vitamin E on cyclooxygenase-2 (COX-2) transcription. Mutat. Res. 2004, 551, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Kalupahana, N.S.; Moustaid-Moussa, N.; Claycombe, K.J. Immunity as a link between obesity and insulin resistance. Mol. Asp. Med. 2012, 33, 26–34. [Google Scholar] [CrossRef]
- Salvestrini, V.; Sell, C.; Lorenzini, A. Obesity may accelerate the aging process. Front. Endocrinol. Lausanne 2019, 10, 266. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Kalupahana, N.S.; Cekanova, M.; LeMieux, M.; Greer, B.; Moustaid-Moussa, N. Modulation of adipose tissue inflammation by bioactive food compounds. J. Nutr. Biochem. 2013, 24, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Jayarathne, S.; Stull, A.J.; Park, O.H.; Kim, J.H.; Thompson, L.; Moustaid-Moussa, N. Protective effects of anthocyanins in obesity-associated inflammation and changes in gut microbiome. Mol. Nutr. Food Res. 2019, 63, e1900149. [Google Scholar] [CrossRef]
- Thangthaeng, N.; Poulose, S.M.; Gomes, S.M.; Miller, M.G.; Bielinski, D.F.; Shukitt-Hale, B. Tart cherry supplementation improves working memory, hippocampal inflammation, and autophagy in aged rats. Age Dordr. 2016, 38, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Kalupahana, N.S.; Claycombe, K.J.; Moustaid-Moussa, N. (n-3) Fatty acids alleviate adipose tissue inflammation and insulin resistance: Mechanistic insights. Adv. Nutr. 2011, 2, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Jayarathne, S.; Stull, A.J.; Miranda, A.; Scoggin, S.; Claycombe-Larson, K.; Kim, J.H.; Moustaid-Moussa, N. Tart cherry reduces inflammation in adipose tissue of zucker fatty rats and cultured 3T3-L1 adipocytes. Nutrients 2018, 10, 1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirakosyan, A.; Seymour, E.M.; Llanes, D.E.U.; Kaufman, P.B.; Bolling, S.F. Chemical profile and antioxidant capacities of tart cherry products. Food Chem. 2009, 115, 20–25. [Google Scholar] [CrossRef]
- Seymour, E.M.; Lewis, S.K.; Urcuyo-Llanes, D.E.; Tanone, I.I.; Kirakosyan, A.; Kaufman, P.B.; Bolling, S.F. Regular tart cherry intake alters abdominal adiposity, adipose gene transcription, and inflammation in obesity-prone rats fed a high fat diet. J. Med. Food 2009, 12, 935–942. [Google Scholar] [CrossRef]
- Kim, B.; Lee, S.G.; Park, Y.K.; Ku, C.S.; Pham, T.X.; Wegner, C.J.; Yang, Y.; Koo, S.I.; Chun, O.K.; Lee, J.Y. Blueberry, blackberry, and blackcurrant differentially affect plasma lipids and pro-inflammatory markers in diet-induced obesity mice. Nutr. Res. Pract. 2016, 10, 494–500. [Google Scholar] [CrossRef]
- Kolehmainen, M.; Mykkanen, O.; Kirjavainen, P.V.; Leppanen, T.; Moilanen, E.; Adriaens, M.; Laaksonen, D.E.; Hallikainen, M.; Puupponen-Pimia, R.; Pulkkinen, L.; et al. Bilberries reduce low-grade inflammation in individuals with features of metabolic syndrome. Mol. Nutr. Food Res. 2012, 56, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- McCall, D.O.; McGartland, C.P.; McKinley, M.C.; Sharpe, P.; McCance, D.R.; Young, I.S.; Woodside, J.V. The effect of increased dietary fruit and vegetable consumption on endothelial activation, inflammation and oxidative stress in hypertensive volunteers. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 658–664. [Google Scholar] [CrossRef]
- Meydani, M. Nutrition interventions in aging and age-associated disease. Ann. N. Y. Acad. Sci. 2001, 928, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Jayarathne, S.; Ramalingam, L.; Edwards, H.; Vanapalli, S.A.; Moustaid-Moussa, N. Tart cherry increases lifespan in caenorhabditis elegans by altering metabolic signaling pathways. Nutrients 2020, 12, 1482. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.; Yue, Y.; Zheng, J.; Park, Y. Caenorhabditis elegans: A convenient in vivo model for assessing the impact of food bioactive compounds on obesity, aging, and Alzheimer’s disease. Annu. Rev. Food Sci. Technol. 2018, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.; Vantipalli, M.C.; Lithgow, G.J. Using Caenorhabditis elegans as a model for aging and age-related diseases. Ann. N. Y. Acad. Sci. 2006, 1067, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Sueishi, Y.; Nii, R.; Kakizaki, N. Resveratrol analogues like piceatannol are potent antioxidants as quantitatively demonstrated through the high scavenging ability against reactive oxygen species and methyl radical. Bioorg. Med. Chem. Lett. 2017, 27, 5203–5206. [Google Scholar] [CrossRef] [PubMed]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Thygesen, L.; Thulin, J.; Mortensen, A.; Skibsted, L.H.; Molgaard, P. Antioxidant activity of cichoric acid and alkamides from Echinacea purpurea, alone and in combination. Food Chem. 2007, 101, 74–81. [Google Scholar] [CrossRef]
- Shen, P.; Yue, Y.; Sun, Q.; Kasireddy, N.; Kim, K.H.; Park, Y. Piceatannol extends the lifespan of Caenorhabditis elegans via DAF-16. Biofactors 2017, 43, 379–387. [Google Scholar] [CrossRef]
- Peng, Y.; Sun, Q.; Gao, R.; Park, Y. AAK-2 and SKN-1 are involved in chicoric-acid-induced lifespan extension in Caenorhabditis elegans. J. Agric. Food Chem. 2019, 67, 9178–9186. [Google Scholar] [CrossRef]
- Yue, Y.; Shen, P.; Xu, Y.; Park, Y. p-Coumaric acid improves oxidative and osmosis stress responses in Caenorhabditis elegans. J. Sci. Food Agric. 2019, 99, 1190–1197. [Google Scholar] [CrossRef]
- Farina, F.; Lambert, E.; Commeau, L.; Lejeune, F.-X.; Roudier, N.; Fonte, C.; Parker, J.A.; Boddaert, J.; Verny, M.; Baulieu, E.-E. The stress response factor daf-16/FOXO is required for multiple compound families to prolong the function of neurons with Huntington’s disease. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kwon, G.; Park, J.; Kim, J.-K.; Lim, Y.-H. Brief communication: SIR-2.1-dependent lifespan extension of Caenorhabditis elegans by oxyresveratrol and resveratrol. Exp. Biol. Med. 2016, 241, 1757–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.-G.; Huang, J.-A.; Li, J.; Yu, P.-H.; Xiong, Z.; Zhang, J.-W.; Gong, Y.-S.; Liu, Z.-H.; Chen, J.-H. Black tea increased survival of Caenorhabditis elegans under stress. J. Agric. Food Chem. 2014, 62, 11163–11169. [Google Scholar] [CrossRef]
- Aparecida Paiva, F.; de Freitas Bonomo, L.; Ferreira Boasquivis, P.; Borges Raposo de Paula, I.T.; Guerra, J.F.d.C.; Mendes Leal, W.; Silva, M.E.; Pedrosa, M.L.; Oliveira, R.d.P. Carqueja (Baccharis trimera) protects against oxidative stress and β-amyloid-induced toxicity in Caenorhabditis elegans. Oxidative Med. Cell. Longev. 2015, 2015, 740162. [Google Scholar] [CrossRef] [Green Version]
- Grünz, G.; Haas, K.; Soukup, S.; Klingenspor, M.; Kulling, S.E.; Daniel, H.; Spanier, B. Structural features and bioavailability of four flavonoids and their implications for lifespan-extending and antioxidant actions in C. elegans. Mech. Ageing Dev. 2012, 133, 1–10. [Google Scholar] [CrossRef]
- Shen, P.; Yue, Y.; Park, Y. A Living model for obesity and aging research: Caenorhabditis elegans. Crit. Rev. Food Sci. Nutr. 2017, 58, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Zhu, L.J.; Yen, K.; Tissenbaum, H.A. Uncoupling lifespan and healthspan in Caenorhabditis elegans longevity mutants. Proc. Natl. Acad. Sci. USA 2015, 112, E277–E286. [Google Scholar] [CrossRef] [Green Version]
- Harrell, C.R.; Jankovic, M.G.; Fellabaum, C.; Volarevic, A.; Djonov, V.; Arsenijevic, A.; Volarevic, V. Molecular mechanisms responsible for anti-inflammatory and immunosuppressive effects of mesenchymal stem cell-derived factors. Adv. Exp. Med. Biol. 2019, 1084, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, Q.; Wang, Y.; Li, L.; Bu, H.; Bao, J. Senescence of mesenchymal stem cells (Review). Int. J. Mol. Med. 2017, 39, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.R.; Kang, K.S. Aging-related genes in mesenchymal stem cells: A mini-review. Gerontology 2013, 59, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yoon, Y.M.; Han, Y.S.; Lee, J.H.; Hur, J.; Lee, S.H. Administration of Cripto in GRP78 overexpressed human MSCs enhances stem cell viability and angiogenesis during human MSC transplantation therapy. Cell Prolif. 2018, 51, e12463. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Han, Y.S.; Lee, S.H. Potentiation of biological effects of mesenchymal stem cells in ischemic conditions by melatonin via upregulation of cellular prion protein expression. J. Pineal Res. 2017, 62, e12385. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Kim, J.S.; Chae, C.W.; Lim, J.R.; Kim, S.Y.; Lee, J.E.; Park, M.C.; Yoon, J.H.; et al. O-cyclic phytosphingosine-1-phosphate stimulates HIF1alpha-dependent glycolytic reprogramming to enhance the therapeutic potential of mesenchymal stem cells. Cell Death Dis. 2019, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial role of phytochemicals on oxidative stress and age-related diseases. Biomed. Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saud, B.; Malla, R.; Shrestha, K. A review on the effect of plant extract on mesenchymal stem cell proliferation and differentiation. Stem Cells Int. 2019, 2019, 7513404. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jung, Y.H.; Oh, J.Y.; Choi, G.E.; Chae, C.W.; Kim, J.S.; Lim, J.R.; Kim, S.Y.; Lee, S.J.; Seong, J.K.; et al. BICD1 mediates HIF1α nuclear translocation in mesenchymal stem cells during hypoxia adaptation. Cell Death Differ. 2019, 26, 1716–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Bliek, A.M.; Sedensky, M.M.; Morgan, P.G. Cell biology of the mitochondrion. Genetics 2017, 207, 843–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, S.H.; Choi, G.E.; Oh, J.Y.; Lee, H.J.; Kim, J.S.; Chae, C.W.; Choi, D.; Han, H.J. Author correction: Succinate promotes stem cell migration through the GPR91-dependent regulation of DRP1-mediated mitochondrial fission. Sci. Rep. 2018, 8, 13326. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Ko, S.H.; Lee, S.J.; Lee, S.H.; Han, H.J. BNIP3 induction by hypoxia stimulates FASN-dependent free fatty acid production enhancing therapeutic potential of umbilical cord blood-derived human mesenchymal stem cells. Redox Biol. 2017, 13, 426–443. [Google Scholar] [CrossRef]
- Liu, S.P.; Shibu, M.A.; Tsai, F.J.; Hsu, Y.M.; Tsai, C.H.; Chung, J.G.; Yang, J.S.; Tang, C.H.; Wang, S.; Li, Q.; et al. Tetramethylpyrazine reverses high-glucose induced hypoxic effects by negatively regulating HIF-1alpha induced BNIP3 expression to ameliorate H9c2 cardiomyoblast apoptosis. Nutr. Metab. 2020, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabacka, M.M.; Gawin, M.; Pierzchalska, M. Phytochemical modulators of mitochondria: The search for chemopreventive agents and supportive therapeutics. Pharmaceuticals 2014, 7, 913–942. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, S.; Zununi Vahed, S.; Ahmadian, E.; Maleki Dizaj, S.; Abedi, A.; Hosseiniyan Khatibi, S.M.; Samiei, M. Stem cell therapy: Curcumin does the trick. Phytother. Res. 2019, 33, 2927–2937. [Google Scholar] [CrossRef] [Green Version]
- Mohar, D.S.; Malik, S. The Sirtuin system: The holy grail of resveratrol? J. Clin. Exp. Cardiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.H.; Lee, H.J.; Kim, J.S.; Lee, S.J.; Han, H.J. EphB2 signaling-mediated Sirt3 expression reduces MSC senescence by maintaining mitochondrial ROS homeostasis. Free Radic. Biol. Med. 2017, 110, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Choi, G.E.; Lee, H.J.; Jung, Y.H.; Chae, C.W.; Kim, J.S.; Lee, C.K.; Han, H.J. 17beta-Estradiol protects mesenchymal stem cells against high glucose-induced mitochondrial oxidants production via Nrf2/Sirt3/MnSOD signaling. Free Radic. Biol. Med. 2019, 130, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yun, C.W.; Hur, J.; Lee, S.H. Fucoidan rescues p-cresol-induced cellular senescence in mesenchymal stem cells via FAK-Akt-TWIST axis. Mar. Drugs 2018, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Ryu, J.M.; Han, Y.S.; Zia, M.F.; Kwon, H.Y.; Noh, H.; Han, H.J.; Lee, S.H. Fucoidan improves bioactivity and vasculogenic potential of mesenchymal stem cells in murine hind limb ischemia associated with chronic kidney disease. J. Mol. Cell. Cardiol. 2016, 97, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Lee, J.H.; Jung, J.S.; Noh, H.; Baek, M.J.; Ryu, J.M.; Yoon, Y.M.; Han, H.J.; Lee, S.H. Fucoidan protects mesenchymal stem cells against oxidative stress and enhances vascular regeneration in a murine hindlimb ischemia model. Int. J. Cardiol. 2015, 198, 187–195. [Google Scholar] [CrossRef]
- Carey, M.A.; Germolec, D.R.; Langenbach, R.; Zeldin, D.C. Cyclooxygenase enzymes in allergic inflammation and asthma. Prostaglandins Leukot. Essent. Fat. Acids 2003, 69, 157–162. [Google Scholar] [CrossRef]
- Peebles, R.S., Jr.; Hashimoto, K.; Morrow, J.D.; Dworski, R.; Collins, R.D.; Hashimoto, Y.; Christman, J.W.; Kang, K.H.; Jarzecka, K.; Furlong, J.; et al. Selective cyclooxygenase-1 and -2 inhibitors each increase allergic inflammation and airway hyperresponsiveness in mice. Am. J. Respir. Crit. Care Med. 2002, 165, 1154–1160. [Google Scholar] [CrossRef]
- Halushka, M.K.; Walker, L.P.; Halushka, P.V. Genetic variation in cyclooxygenase 1: Effects on response to aspirin. Clin. Pharm. 2003, 73, 122–130. [Google Scholar] [CrossRef]
- Szczeklik, W.; Sanak, M.; Szczeklik, A. Functional effects and gender association of COX-2 gene polymorphism G-765C in bronchial asthma. J. Allergy Clin. Immunol. 2004, 114, 248–253. [Google Scholar] [CrossRef]
- Lee, C.R.; Bottone, F.G., Jr.; Krahn, J.M.; Li, L.; Mohrenweiser, H.W.; Cook, M.E.; Petrovich, R.M.; Bell, D.A.; Eling, T.E.; Zeldin, D.C. Identification and functional characterization of polymorphisms in human cyclooxygenase-1 (PTGS1). Pharm. Genom. 2007, 17, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Murata, T.; Tanaka, H.; Matsuoka, T.; Sakata, D.; Yoshida, N.; Katagiri, K.; Kinashi, T.; Tanaka, T.; Miyasaka, M.; et al. Thromboxane A2 modulates interaction of dendritic cells and T cells and regulates acquired immunity. Nat. Immunol. 2003, 4, 694–701. [Google Scholar] [CrossRef]
- Katamura, K.; Shintaku, N.; Yamauchi, Y.; Fukui, T.; Ohshima, Y.; Mayumi, M.; Furusho, K. Prostaglandin E2 at priming of naive CD4+ T cells inhibits acquisition of ability to produce IFN-gamma and IL-2, but not IL-4 and IL-5. J. Immunol. 1995, 155, 4604–4612. [Google Scholar]
- Zhou, W.; Blackwell, T.S.; Goleniewska, K.; O’Neal, J.F.; Fitzgerald, G.A.; Lucitt, M.; Breyer, R.M.; Peebles, R.S., Jr. Prostaglandin I2 analogs inhibit Th1 and Th2 effector cytokine production by CD4 T cells. J. Leukoc. Biol. 2007, 81, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Carey, M.A.; Germolec, D.R.; Bradbury, J.A.; Gooch, R.A.; Moorman, M.P.; Flake, G.P.; Langenbach, R.; Zeldin, D.C. Accentuated T helper type 2 airway response after allergen challenge in cyclooxygenase-1-/- but not cyclooxygenase-2-/- mice. Am. J. Respir. Crit. Care Med. 2003, 167, 1509–1515. [Google Scholar] [CrossRef]
- Li, H.; Bradbury, J.A.; Dackor, R.T.; Edin, M.L.; Graves, J.P.; DeGraff, L.M.; Wang, P.M.; Bortner, C.D.; Maruoka, S.; Lih, F.B.; et al. Cyclooxygenase-2 regulates Th17 cell differentiation during allergic lung inflammation. Am. J. Respir. Crit. Care Med. 2011, 184, 37–49. [Google Scholar] [CrossRef]
- Durrant, D.M.; Metzger, D.W. Emerging roles of T helper subsets in the pathogenesis of asthma. Immunol. Investig. 2010, 39, 526–549. [Google Scholar] [CrossRef]
- Li, H.; Edin, M.L.; Bradbury, J.A.; Graves, J.P.; DeGraff, L.M.; Gruzdev, A.; Cheng, J.; Dackor, R.T.; Wang, P.M.; Bortner, C.D.; et al. Cyclooxygenase-2 inhibits T helper cell type 9 differentiation during allergic lung inflammation via down-regulation of IL-17RB. Am. J. Respir. Crit. Care Med. 2013, 187, 812–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdi, J.G.; Mahdi, A.J.; Mahdi, A.J.; Bowen, I.D. The historical analysis of aspirin discovery, its relation to the willow tree and antiproliferative and anticancer potential. Cell Prolif. 2006, 39, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Botting, R.M. The mechanism of action of aspirin. Thromb. Res. 2003, 110, 255–258. [Google Scholar] [CrossRef]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the war against inflammation with natural products. Front. Pharmacol. 2018, 9, 976. [Google Scholar] [CrossRef]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from omega-6 and omega-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Edin, M.L.; Gruzdev, A.; Cheng, J.; Bradbury, J.A.; Graves, J.P.; DeGraff, L.M.; Zeldin, D.C. Regulation of T helper cell subsets by cyclooxygenases and their metabolites. Prostaglandins Other Lipid Mediat. 2013, 104–105, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Askew, R.L.; Capo-Lugo, C.E.; Naidech, A.; Prabhakaran, S. Differential effects of time to initiation of therapy on disability and quality of life in patients with mild and moderate to severe ischemic stroke. Arch. Phys. Med. Rehabil. 2020, 101, 1515–1522. [Google Scholar] [CrossRef]
- Khoshnam, S.E.; Winlow, W.; Farzaneh, M.; Farbood, Y.; Moghaddam, H.F. Pathogenic mechanisms following ischemic stroke. Neurol. Sci. 2017, 38, 1167–1186. [Google Scholar] [CrossRef] [PubMed]
- Siesjo, B.K. Pathophysiology and treatment of focal cerebral ischemia. Part I: Pathophysiology. J. Neurosurg. 1992, 77, 169–184. [Google Scholar] [CrossRef]
- Song, D.; Xu, J.; Du, T.; Yan, E.; Hertz, L.; Walz, W.; Peng, L. Inhibition of brain swelling after ischemia-reperfusion by beta-adrenergic antagonists: Correlation with increased K+ and decreased Ca2+ concentrations in extracellular fluid. Biomed. Res. Int 2014, 2014, 873590. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Okami, N.; Sakata, H.; Maier, C.M.; Narasimhan, P.; Goeders, C.E.; Chan, P.H. Oxidative stress in ischemic brain damage: Mechanisms of cell death and potential molecular targets for neuroprotection. Antioxid. Redox Signal. 2011, 14, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.I.; Geuss, E.; Kleikers, P.W.M.; Mencl, S.; Herrmann, A.M.; Buendia, I.; Egea, J.; Meuth, S.G.; Lopez, M.G.; Kleinschnitz, C.; et al. NOX4-dependent neuronal autotoxicity and BBB breakdown explain the superior sensitivity of the brain to ischemic damage. Proc. Natl. Acad. Sci. USA 2017, 114, 12315–12320. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chen, S.; Luo, Y.; Han, Z. Crosstalk between Inflammation and the BBB in Stroke. Curr. Neuropharmacol. 2020, 18, 1227–1236. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, Q.; Anrather, J.; Shi, F.D. Immune interventions in stroke. Nat. Rev. Neurol. 2015, 11, 524–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keaney, J.; Campbell, M. The dynamic blood-brain barrier. FEBS J. 2015, 282, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Yamaguchi, T.; Kirino, T.; Orzi, F.; Klatzo, I. The effects of 5-minute ischemia in Mongolian gerbils: I. Blood-brain barrier, cerebral blood flow, and local cerebral glucose utilization changes. Acta Neuropathol. 1983, 60, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Pillai, D.R.; Dittmar, M.S.; Baldaranov, D.; Heidemann, R.M.; Henning, E.C.; Schuierer, G.; Bogdahn, U.; Schlachetzki, F. Cerebral ischemia-reperfusion injury in rats—a 3 T MRI study on biphasic blood-brain barrier opening and the dynamics of edema formation. J. Cereb. Blood Flow Metab. 2009, 29, 1846–1855. [Google Scholar] [CrossRef]
- Knowland, D.; Arac, A.; Sekiguchi, K.J.; Hsu, M.; Lutz, S.E.; Perrino, J.; Steinberg, G.K.; Barres, B.A.; Nimmerjahn, A.; Agalliu, D. Stepwise recruitment of transcellular and paracellular pathways underlies blood-brain barrier breakdown in stroke. Neuron 2014, 82, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Zhang, L.; Pu, H.; Mao, L.; Hu, X.; Jiang, X.; Xu, N.; Stetler, R.A.; Zhang, F.; Liu, X.; et al. Rapid endothelial cytoskeletal reorganization enables early blood-brain barrier disruption and long-term ischaemic reperfusion brain injury. Nat. Commun. 2016, 7, 10523. [Google Scholar] [CrossRef]
- Song, J.; Wu, C.; Korpos, E.; Zhang, X.; Agrawal, S.M.; Wang, Y.; Faber, C.; Schafers, M.; Korner, H.; Opdenakker, G.; et al. Focal MMP-2 and MMP-9 activity at the blood-brain barrier promotes chemokine-induced leukocyte migration. Cell Rep. 2015, 10, 1040–1054. [Google Scholar] [CrossRef] [Green Version]
- Romanic, A.M.; White, R.F.; Arleth, A.J.; Ohlstein, E.H.; Barone, F.C. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: Inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke 1998, 29, 1020–1030. [Google Scholar] [CrossRef] [Green Version]
- Candelario-Jalil, E.; Yang, Y.; Rosenberg, G.A. Diverse roles of matrix metalloproteinases and tissue inhibitors of metalloproteinases in neuroinflammation and cerebral ischemia. Neuroscience 2009, 158, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Malemud, C.J. Matrix metalloproteinases (MMPs) in health and disease: An overview. Front. Biosci. 2006, 11, 1696–1701. [Google Scholar] [CrossRef]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Maier, C.M.; Hsieh, L.; Crandall, T.; Narasimhan, P.; Chan, P.H. Evaluating therapeutic targets for reperfusion-related brain hemorrhage. Ann. Neurol. 2006, 59, 929–938. [Google Scholar] [CrossRef]
- Simard, M.; Arcuino, G.; Takano, T.; Liu, Q.S.; Nedergaard, M. Signaling at the gliovascular interface. J. Neurosci. 2003, 23, 9254–9262. [Google Scholar] [CrossRef] [PubMed]
- Solenov, E.; Watanabe, H.; Manley, G.T.; Verkman, A.S. Sevenfold-reduced osmotic water permeability in primary astrocyte cultures from AQP-4-deficient mice, measured by a fluorescence quenching method. Am. J. Physiol. Cell Physiol. 2004, 286, C426–C432. [Google Scholar] [CrossRef]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat. Med. 2000, 6, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.W.; Jung, Y.S. Hypoxia induces FoxO3a-mediated dysfunction of blood-brain barrier. Biochem. Biophys. Res. Commun. 2014, 450, 1638–1642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Investig. 2000, 106, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Ye, Z.; Pan, Y.; Li, X.; Fu, X.; Zhang, B.; Li, Y.; Lin, W.; Li, X.; Gao, Q. Vascular endothelial growth factor aggravates cerebral ischemia and reperfusion-induced blood-brain-barrier disruption through regulating LOC102640519/HOXC13/ZO-1 signaling. Exp. Cell Res. 2018, 369, 275–283. [Google Scholar] [CrossRef]
- Ji, B.; Zhou, F.; Han, L.; Yang, J.; Fan, H.; Li, S.; Li, J.; Zhang, X.; Wang, X.; Chen, X.; et al. Sodium tanshinone IIA sulfonate enhances effectiveness Rt-PA treatment in acute ischemic stroke patients associated with ameliorating blood-brain barrier damage. Transl. Stroke Res. 2017, 8, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Lee, J.S.; Jang, H.J.; Kim, S.M.; Chang, M.S.; Park, S.H.; Kim, K.S.; Bae, J.; Park, J.W.; Lee, B.; et al. Chlorogenic acid ameliorates brain damage and edema by inhibiting matrix metalloproteinase-2 and 9 in a rat model of focal cerebral ischemia. Eur. J. Pharm. 2012, 689, 89–95. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, Z.; Jin, T. Crocin protects against cerebral- ischemia-induced damage in aged rats through maintaining the integrity of blood-brain barrier. Restor. Neurol. Neurosci. 2017, 35, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, L.; Kong, L.; Zhu, Z.; Zhang, W.; Song, J.; Chang, J.; Du, G. Pinocembrin protects blood-brain barrier function and expands the therapeutic time window for tissue-type plasminogen activator treatment in a rat thromboembolic stroke model. Biomed. Res. Int. 2018, 2018, 8943210. [Google Scholar] [CrossRef]
- Zhang, W.; Song, J.K.; Zhang, X.; Zhou, Q.M.; He, G.R.; Xu, X.N.; Rong, Y.; Zhou, W.X.; Du, G.H. Salvianolic acid A attenuates ischemia reperfusion induced rat brain damage by protecting the blood brain barrier through MMP-9 inhibition and anti-inflammation. Chin. J. Nat. Med. 2018, 16, 184–193. [Google Scholar] [CrossRef]
- Li, S.; Bian, L.; Fu, X.; Ai, Q.; Sui, Y.; Zhang, A.; Gao, H.; Zhong, L.; Lu, D. Gastrodin pretreatment alleviates rat brain injury caused by cerebral ischemic-reperfusion. Brain Res. 2019, 1712, 207–216. [Google Scholar] [CrossRef]
- Chang, C.Y.; Chen, J.Y.; Wu, M.H.; Hu, M.L. Therapeutic treatment with vitamin C reduces focal cerebral ischemia-induced brain infarction in rats by attenuating disruptions of blood brain barrier and cerebral neuronal apoptosis. Free Radic. Biol. Med. 2020, 155, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.D.; Liu, N.N.; Zhang, S.; Ma, G.D.; Yang, H.G.; Kong, L.L.; Du, G.H. Salvianolic acid A prevented cerebrovascular endothelial injury caused by acute ischemic stroke through inhibiting the Src signaling pathway. Acta Pharm. Sin. 2020, 42, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, R.N.; Li, L.H.; Qu, Y.Z.; Gao, G.D. Astragaloside IV reduces cerebral edema post-ischemia/reperfusion correlating the suppression of MMP-9 and AQP4. Eur. J. Pharm. 2013, 715, 189–195. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Liang, C.; Tan, R.; Tan, L.; Tan, R. Pharmacodynamic effect of ellagic acid on ameliorating cerebral ischemia/reperfusion injury. Pharmacology 2019, 104, 320–331. [Google Scholar] [CrossRef]
- Nag, S.; Venugopalan, R.; Stewart, D.J. Increased caveolin-1 expression precedes decreased expression of occludin and claudin-5 during blood-brain barrier breakdown. Acta Neuropathol. 2007, 114, 459–469. [Google Scholar] [CrossRef]
- Lee, B.K.; Hyun, S.W.; Jung, Y.S. Yuzu and hesperidin ameliorate blood-brain barrier disruption during hypoxia via antioxidant activity. Antioxid. Basel 2020, 9, 843. [Google Scholar] [CrossRef]
- Liu, J.; Chen, L.; Zhang, X.; Pan, L.; Jiang, L. The protective effects of juglanin in cerebral ischemia reduce blood-brain barrier permeability via inhibition of VEGF/VEGFR2 signaling. Drug Des. Dev. Ther. 2020, 14, 3165–3175. [Google Scholar] [CrossRef]
- Chen, H.; Guan, B.; Chen, X.; Chen, X.; Li, C.; Qiu, J.; Yang, D.; Liu, K.J.; Qi, S.; Shen, J. Baicalin attenuates blood-brain barrier disruption and hemorrhagic transformation and improves neurological outcome in ischemic stroke rats with delayed t-PA treatment: Involvement of ONOO(-)-MMP-9 pathway. Transl. Stroke Res. 2018, 9, 515–529. [Google Scholar] [CrossRef]
- Li, W.; Suwanwela, N.C.; Patumraj, S. Curcumin prevents reperfusion injury following ischemic stroke in rats via inhibition of NFkappaB, ICAM-1, MMP-9 and caspase-3 expression. Mol. Med. Rep. 2017, 16, 4710–4720. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, Y.; Zhang, X.; Chen, R.; Zhang, L.; Xue, J.; Gao, X. Dl-3-N-Butylphthalide alleviates the blood-brain barrier permeability of focal cerebral ischemia reperfusion in mice. Neuroscience 2019, 413, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Guo, Y.; Yang, W.; Zheng, P.; Zeng, J.; Tong, W. Protective effect of ginsenoside Rb1 on integrity of blood-brain barrier following cerebral ischemia. Exp. Brain Res. 2015, 233, 2823–2831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.; Hu, G.; Zhang, G.; Zhao, G.; Shi, M. Ginsenoside Rd attenuates blood-brain barrier damage by suppressing proteasome-mediated signaling after transient forebrain ischemia. Neuroreport 2020, 31, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.B.; Wang, W.; Gao, J.M.; Li, F.; Shi, J.S.; Gong, Q.H. Icariside II attenuates cerebral ischemia/reperfusion-induced blood-brain barrier dysfunction in rats via regulating the balance of MMP9/TIMP1. Acta Pharm. Sin. 2020, 41, 1547–1556. [Google Scholar] [CrossRef]
- Jang, J.W.; Lee, J.K.; Lee, M.C.; Piao, M.S.; Kim, S.H.; Kim, H.S. Melatonin reduced the elevated matrix metalloproteinase-9 level in a rat photothrombotic stroke model. J. Neurol. Sci. 2012, 323, 221–227. [Google Scholar] [CrossRef]
- Jin, Z.; Ke, J.; Guo, P.; Wang, Y.; Wu, H. Quercetin improves blood-brain barrier dysfunction in rats with cerebral ischemia reperfusion via Wnt signaling pathway. Am. J. Transl. Res. 2019, 11, 4683–4695. [Google Scholar]
- Wei, H.; Wang, S.; Zhen, L.; Yang, Q.; Wu, Z.; Lei, X.; Lv, J.; Xiong, L.; Xue, R. Resveratrol attenuates the blood-brain barrier dysfunction by regulation of the MMP-9/TIMP-1 balance after cerebral ischemia reperfusion in rats. J. Mol. Neurosci. 2015, 55, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.W.; Lee, J.K.; Hur, H.; Kim, T.W.; Joo, S.P.; Piao, M.S. Rutin improves functional outcome via reducing the elevated matrix metalloproteinase-9 level in a photothrombotic focal ischemic model of rats. J. Neurol. Sci. 2014, 339, 75–80. [Google Scholar] [CrossRef]
- Mondal, N.K.; Behera, J.; Kelly, K.E.; George, A.K.; Tyagi, P.K.; Tyagi, N. Tetrahydrocurcumin epigenetically mitigates mitochondrial dysfunction in brain vasculature during ischemic stroke. Neurochem. Int. 2019, 122, 120–138. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Zhang, Z.; Zou, Y.; Tian, Q.; Han, S.; Xu, Z.; Liao, J.; Gao, L.; Chen, Q.; Li, M. Tetramethylpyrazine attenuates blood-brain barrier disruption in ischemia/reperfusion injury through the JAK/STAT signaling pathway. Eur. J. Pharm. 2019, 854, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.D.; Hammock, B.D. The 2014 Bernard, B. Brodie award lecture-epoxide hydrolases: Drug metabolism to therapeutics for chronic pain. Drug Metab. Dispos. 2015, 43, 788–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, K.M.; McReynolds, C.B.; Schmidt, W.K.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for pain, inflammatory and neurodegenerative diseases. Pharmacol. Ther. 2017, 180, 62–76. [Google Scholar] [CrossRef]
- Inceoglu, B.; Bettaieb, A.; Trindade da Silva, C.A.; Lee, K.S.; Haj, F.G.; Hammock, B.D. Endoplasmic reticulum stress in the peripheral nervous system is a significant driver of neuropathic pain. Proc. Natl. Acad. Sci. USA. 2015, 112, 9082–9087. [Google Scholar] [CrossRef] [Green Version]
- Cervenka, L.; Huskova, Z.; Kopkan, L.; Kikerlova, S.; Sedlakova, L.; Vanourkova, Z.; Alanova, P.; Kolar, F.; Hammock, B.D.; Hwang, S.H.; et al. Two pharmacological epoxyeicosatrienoic acid-enhancing therapies are effectively antihypertensive and reduce the severity of ischemic arrhythmias in rats with angiotensin II-dependent hypertension. J. Hypertens. 2018, 36, 1326–1341. [Google Scholar] [CrossRef]
- Chábová, Č.V.; Kujal, P.; Škaroupková, P.; Varňourková, Z.; Vacková, Š.; Husková, Z.; Kikerlová, S.; Sadowski, J.; Kompanowska-Jezierska, E.; Baranowska, I.; et al. Combined Inhibition of soluble epoxide hydrolase and renin-angiotensin system exhibits superior renoprotection to renin-angiotensin system blockade in 5/6 nephrectomized ren-2 transgenic hypertensive rats with established chronic kidney disease. Kidney Blood Press. Res. 2018, 43, 329–349. [Google Scholar] [CrossRef]
- Zarriello, S.; Tuazon, J.P.; Corey, S.; Schimmel, S.; Rajani, M.; Gorsky, A.; Incontri, D.; Hammock, B.D.; Borlongan, C.V. Humble beginnings with big goals: Small molecule soluble epoxide hydrolase inhibitors for treating CNS disorders. Prog. Neurobiol. 2019, 172, 23–39. [Google Scholar] [CrossRef]
- Swardfager, W.; Hennebelle, M.; Yu, D.; Hammock, B.D.; Levitt, A.J.; Hashimoto, K.; Taha, A.Y. Metabolic/inflammatory/vascular comorbidity in psychiatric disorders; soluble epoxide hydrolase (sEH) as a possible new target. Neurosci. Biobehav. Rev. 2018, 87, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Tu, R.; Armstrong, J.; Lee, K.S.S.; Hammock, B.D.; Sapirstein, A.; Koehler, R.C. Soluble epoxide hydrolase inhibition decreases reperfusion injury after focal cerebral ischemia. Sci. Rep. 2018, 8, 5279. [Google Scholar] [CrossRef]
- Ren, Q.; Ma, M.; Yang, J.; Nonaka, R.; Yamaguchi, A.; Ishikawa, K.I.; Kobayashi, K.; Murayama, S.; Hwang, S.H.; Saiki, S.; et al. Soluble epoxide hydrolase plays a key role in the pathogenesis of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, E5815–E5823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Ren, Q.; Yang, J.; Zhang, K.; Xiong, Z.; Ishima, T.; Pu, Y.; Hwang, S.H.; Toyoshima, M.; Iwayama, Y.; et al. Key role of soluble epoxide hydrolase in the neurodevelopmental disorders of offspring after maternal immune activation. Proc. Natl. Acad. Sci. USA 2019, 116, 7083–7088. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Zhang, B.; Xu, M.; Morisseau, C.; Hwang, S.H.; Hammock, B.D.; Li, Q.X. 1-Trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl) urea, a selective and potent dual inhibitor of soluble epoxide hydrolase and p38 kinase intervenes in alzheimer’s signaling in human nerve cells. ACS Chem. Neurosci. 2019, 10, 4018–4030. [Google Scholar] [CrossRef]
- Wang, W.; Yang, J.; Edin, M.L.; Wang, Y.; Luo, Y.; Wan, D.; Yang, H.; Song, C.Q.; Xue, W.; Sanidad, K.Z.; et al. Targeted metabolomics identifies the cytochrome P450 monooxygenase eicosanoid pathway as a novel therapeutic target of colon tumorigenesis. Cancer Res. 2019, 79, 1822–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panigrahy, D.; Gartung, A.; Yang, J.; Yang, H.; Gilligan, M.M.; Sulciner, M.L.; Bhasin, S.S.; Bielenberg, D.R.; Chang, J.; Schmidt, B.A.; et al. Preoperative stimulation of resolution and inflammation blockade eradicates micrometastases. J. Clin. Investig. 2019, 129, 2964–2979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, A.; Galuppo, L.; Hood, D.; Hwang, S.H.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase activity and pharmacologic inhibition in horses with chronic severe laminitis. Equine Vet. J. 2017, 49, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McReynolds, C.B.; Hwang, S.H.; Yang, J.; Wan, D.; Wagner, K.; Morisseau, C.; Li, D.; Schmidt, W.K.; Hammock, B.D. Pharmaceutical effects of inhibiting the soluble epoxide hydrolase in canine osteoarthritis. Front. Pharmacol. 2019, 10, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammock, B.D.; McReynolds, C.B.; Wagner, K.; Buckpitt, A.; Cortes-Puch, I.; Croston, G.; Lee, K.S.S.; Yang, J.; Schmidt, W.K.; Hwang, S.H. Movement to the clinic of soluble epoxide hydrolase inhibitor EC5026 as an analgesic for neuropathic pain and for use as a nonaddictive opioid alternative. J. Med. Chem. 2021, 64, 1856–1872. [Google Scholar] [CrossRef]
- McReynolds, C.B.; Cortes-Puch, I.; Ravindran, R.; Khan, I.H.; Hammock, B.G.; Shih, P.-a.B.; Hammock, B.D.; Yang, J. Plasma Linoleate Diols Are Potential Biomarkers for Severe COVID-19 Infections. Front. Physiol. 2021, 12, 663869. [Google Scholar] [CrossRef]
- Ferrara, M.; De Gennaro, L. How much sleep do we need? Sleep Med. Rev. 2001, 5, 155–179. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Kim, Y.; Yoon, S.; Kim, S.J.; Yi, S.S. Sleep-inducing effect of Passiflora incarnata L. extract by single and repeated oral administration in rodent animals. Food Sci. Nutr. 2020, 8, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Roth, T. Insomnia: Definition, prevalence, etiology, and consequences. J. Clin. Sleep Med. 2007, 3, S7–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, L.L.; Guo, H.; Song, N.N.; Jia, Z.P.; Hu, X.T.; Huang, J.F.; Ding, Y.Q.; Richter-Levin, G.; Zhou, Q.X.; Xu, L. Light exposure before learning improves memory consolidation at night. Sci. Rep. 2015, 5, 15578. [Google Scholar] [CrossRef]
- Lucassen, E.A.; Zhao, X.; Rother, K.I.; Mattingly, M.S.; Courville, A.B.; de Jonge, L.; Csako, G.; Cizza, G. Sleep Extension Study Group. Evening chronotype is associated with changes in eating behavior, more sleep apnea, and increased stress hormones in short sleeping obese individuals. PLoS ONE 2013, 8, e56519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Duan, P.; Yao, L.; Hou, H. Shiftwork-Mediated disruptions of circadian rhythms and sleep homeostasis cause serious health problems. Int. J. Genom. 2018, 2018, 8576890. [Google Scholar] [CrossRef] [PubMed]
- Potter, G.D.; Skene, D.J.; Arendt, J.; Cade, J.E.; Grant, P.J.; Hardie, L.J. Circadian rhythm and sleep disruption: Causes, metabolic consequences, and countermeasures. Endocr. Rev. 2016, 37, 584–608. [Google Scholar] [CrossRef] [Green Version]
- Brites, D.; Fernandes, A. Neuroinflammation and depression: Microglia activation, extracellular microvesicles and microRNA dysregulation. Front. Cell Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Leproult, R.; Van Cauter, E. Role of sleep and sleep loss in hormonal release and metabolism. Endocr. Dev. 2010, 17, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.H.; Lim, K.; Yang, H.S.; Lee, J.K.; Kim, Y.; Park, S.K.; Kim, S.H.; Park, S.; Kim, T.H.; Moon, J.S.; et al. Improvement in neurogenesis and memory function by administration of Passiflora incarnata L. extract applied to sleep disorder in rodent models. J. Chem. Neuroanat. 2019, 98, 27–40. [Google Scholar] [CrossRef]
- Miroddi, M.; Calapai, G.; Navarra, M.; Minciullo, P.L.; Gangemi, S. Passiflora incarnata L.: Ethnopharmacology, clinical application, safety and evaluation of clinical trials. J. Ethnopharmacol. 2013, 150, 791–804. [Google Scholar] [CrossRef]
- He, M.; Min, J.-W.; Kong, W.-L.; He, X.-H.; Li, J.-X.; Peng, B.-W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Malar, D.S.; Suryanarayanan, V.; Prasanth, M.I.; Singh, S.K.; Balamurugan, K.; Devi, K.P. Vitexin inhibits Abeta25-35 induced toxicity in Neuro-2a cells by augmenting Nrf-2/HO-1 dependent antioxidant pathway and regulating lipid homeostasis by the activation of LXR-alpha. Toxicol. Vitr. 2018, 50, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Soulimani, R.; Younos, C.; Jarmouni, S.; Bousta, D.; Misslin, R.; Mortier, F. Behavioural effects of Passiflora incarnata L. and its indole alkaloid and flavonoid derivatives and maltol in the mouse. J. Ethnopharmacol. 1997, 57, 11–20. [Google Scholar] [CrossRef]
- Choi, J.S.; Islam, M.N.; Ali, M.Y.; Kim, E.J.; Kim, Y.M.; Jung, H.A. Effects of C-glycosylation on anti-diabetic, anti-Alzheimer’s disease and anti-inflammatory potential of apigenin. Food Chem. Toxicol. 2014, 64, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, B.; Kimiagar, M.; Sadeghniiat, K.; Shirazi, M.M.; Hedayati, M.; Rashidkhani, B. The effect of magnesium supplementation on primary insomnia in elderly: A double-blind placebo-controlled clinical trial. J. Res. Med. Sci. 2012, 17, 1161–1169. [Google Scholar]
- Akhondzadeh, S.; Naghavi, H.R.; Vazirian, M.; Shayeganpour, A.; Rashidi, H.; Khani, M. Passionflower in the treatment of generalized anxiety: A pilot double-blind randomized controlled trial with oxazepam. J. Clin. Pharm. Ther. 2001, 26, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Shal, B.; Ding, W.; Ali, H.; Kim, Y.S.; Khan, S. Anti-neuroinflammatory potential of natural products in attenuation of alzheimer’s disease. Front. Pharm. 2018, 9, 548. [Google Scholar] [CrossRef]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory networks underlying colorectal cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kim, J.H.; Lee, J.H.; Lim, Y.; Lee, Y.H. 2’-Hydroxyflavanone induces apoptosis through Egr-1 involving expression of Bax, p21, and NAG-1 in colon cancer cells. Mol. Nutr. Food Res. 2012, 56, 761–774. [Google Scholar] [CrossRef]
- Zhong, Y.; Krisanapun, C.; Lee, S.-H.; Nualsanit, T.; Sams, C.; Peungvicha, P.; Baek, S.J. Molecular targets of apigenin in colorectal cancer cells: Involvement of p21, NAG-1 and p53. Eur. J. Cancer 2010, 46, 3365–3374. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.U.; Lee, B.-S.; Lee, S.-H.; Baek, S.J.; Shin, Y.S.; Kim, C.-H. Expression of NSAID-activated gene-1 by EGCG in head and neck cancer: Involvement of ATM-dependent p53 expression. J. Nutr. Biochem. 2013, 24, 986–999. [Google Scholar] [CrossRef]
- Park, M.-H.; Chung, C.; Lee, S.H.; Baek, S.J.; Kim, J.S. Caffeic acid phenethyl ester induces the expression of NAG-1 via activating transcription factor 3. J. Life Sci. 2018, 28, 37–42. [Google Scholar] [CrossRef]
- Lee, S.H.; Krisanapun, C.; Baek, S.J. NSAID-activated gene-1 as a molecular target for capsaicin-induced apoptosis through a novel molecular mechanism involving GSK3beta, C/EBPbeta and ATF3. Carcinogenesis 2010, 31, 719–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auyeung, K.K.; Ko, J.K. Novel herbal flavonoids promote apoptosis but differentially induce cell cycle arrest in human colon cancer cell. Investig. New Drugs 2010, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Chung, H.-J.; Park, H.J.; Kim, G.D.; Ahn, Y.-H.; Lee, S.K. Suppression of Src/ERK and GSK-3/β-catenin signaling by pinosylvin inhibits the growth of human colorectal cancer cells. Food Chem. Toxicol. 2013, 55, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.M.; Min, K.J.; Kim, S.; Park, J.W.; Kim, D.E.; Chun, K.S.; Kim, Y.H.; Lee, T.J.; Kim, S.H.; Choi, Y.H.; et al. Silibinin induces apoptosis of HT29 colon carcinoma cells through early growth response-1 (EGR-1)-mediated non-steroidal anti-inflammatory drug-activated gene-1 (NAG-1) up-regulation. Chem. Biol. Interact. 2014, 211, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Shetty, A.V.; Thirugnanam, S.; Dakshinamoorthy, G.; Samykutty, A.; Zheng, G.; Chen, A.; Bosland, M.C.; Kajdacsy-Balla, A.; Gnanasekar, M. 18α-glycyrrhetinic acid targets prostate cancer cells by down-regulating inflammation-related genes. Int. J. Oncol. 2011, 39, 635–640. [Google Scholar] [CrossRef] [Green Version]
- Chiu, S.C.; Wang, M.J.; Yang, H.H.; Chen, S.P.; Huang, S.Y.; Chen, Y.L.; Lin, S.Z.; Harn, H.J.; Pang, C.Y. Activation of NAG-1 via JNK signaling revealed an isochaihulactone-triggered cell death in human LNCaP prostate cancer cells. BMC Cancer 2011, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.L.; Su, K.J.; Chen, C.J.; Wei, C.W.; Lin, C.J.; Yiang, G.T.; Lin, S.Z.; Harn, H.J.; Chen, Y.L. Synergistic anti-tumor activity of isochaihulactone and paclitaxel on human lung cancer cells. J. Cell Physiol. 2012, 227, 213–222. [Google Scholar] [CrossRef]
- Harn, H.-J.; Chuang, H.-M.; Chang, L.-F.; Huang, A.; Hsieh, S.-T.; Lin, S.-Z.; Chou, C.-W.; Kuo, Y.-H.; Chiou, T.-W. Taiwanin A targets non-steroidal anti-inflammatory drug-activated gene-1 in human lung carcinoma. Fitoterapia 2014, 99, 227–235. [Google Scholar] [CrossRef]
- Yu, C.Y.; Su, K.Y.; Lee, P.L.; Jhan, J.Y.; Tsao, P.H.; Chan, D.C.; Chen, Y.L. Potential Therapeutic role of hispidulin in gastric cancer through induction of apoptosis via NAG-1 signaling. Evid. Based Complement. Altern. Med. 2013, 2013, 518301. [Google Scholar] [CrossRef]
- Anticancer Agents from Natural Products; Cragg, G.; Kingston, D.G.I.; Newman, D.J. (Eds.) CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2012. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, B.B.; Tiwari, V.K. Natural products: An evolving role in future drug discovery. Eur. J. Med. Chem. 2011, 46, 4769–4807. [Google Scholar] [CrossRef]
- Safe, S.; Abbruzzese, J.; Abdelrahim, M.; Hedrick, E. Specificity protein transcription factors and cancer: Opportunities for drug development. Cancer Prev. Res. Phila 2018, 11, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Cekanova, M.; Baek, S.J. Multiple mechanisms are involved in 6-gingerol-induced cell growth arrest and apoptosis in human colorectal cancer cells. Mol. Carcinog. 2008, 47, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Piyanuch, R.; Sukhthankar, M.; Wandee, G.; Baek, S.J. Berberine, a natural isoquinoline alkaloid, induces NAG-1 and ATF3 expression in human colorectal cancer cells. Cancer Lett. 2007, 258, 230–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auyeung, K.K.; Ko, J.K. Coptis chinensis inhibits hepatocellular carcinoma cell growth through nonsteroidal anti-inflammatory drug-activated gene activation. Int. J. Mol. Med. 2009, 24, 571–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nualsanit, T.; Rojanapanthu, P.; Gritsanapan, W.; Lee, S.H.; Lawson, D.; Baek, S.J. Damnacanthal, a noni component, exhibits antitumorigenic activity in human colorectal cancer cells. J. Nutr. Biochem. 2012, 23, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Bottone, F.G., Jr.; Baek, S.J.; Nixon, J.B.; Eling, T.E. Diallyl disulfide (DADS) induces the antitumorigenic NSAID-activated gene (NAG-1) by a p53-dependent mechanism in human colorectal HCT 116 cells. J. Nutr. 2002, 132, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Kim, J.-S.; Yamaguchi, K.; Eling, T.E.; Baek, S.J. Indole-3-carbinol and 3,3′-diindolylmethane induce expression of NAG-1 in a p53-independent manner. Biochem. Biophys. Res. Commun. 2005, 328, 63–69. [Google Scholar] [CrossRef]
- Baek, S.J.; Kim, J.-S.; Jackson, F.R.; Eling, T.E.; McEntee, M.F.; Lee, S.-H. Epicatechin gallate-induced expression of NAG-1 is associated with growth inhibition and apoptosis in colon cancer cells. Carcinogenesis 2004, 25, 2425–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-N.; Sukhthankar, M.; Lee, S.-H.; Yoon, J.-H.; Baek, S.J. Green tea catechin (−)-epicatechin gallate induces tumour suppressor protein ATF3 via EGR-1 activation. Eur. J. Cancer 2007, 43, 2404–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, L.C.; Baek, S.J.; Call, A.; Eling, T.E. Nonsteroidal anti-inflammatory drug-activated gene (NAG-1) is induced by genistein through the expression of p53 in colorectal cancer cells. Int. J. Cancer 2003, 105, 747–753. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, M.Y.; Moon, J.Y.; Hwang, J.W.; Lee, S.Y.; Joo, Y.M.; Han, S.I.; Park, H.G.; Kang, H.S. Implication of NAG-1 in synergistic induction of apoptosis by combined treatment of sodium salicylate and PI3K/MEK1/2 inhibitors in A549 human lung adenocarcinoma cells. Biochem. Pharm. 2008, 75, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.F.; Tao, M.; Ho, L.I.; Chiou, T.W.; Lin, S.Z.; Su, H.L.; Harn, H.J. Isochaihulactone-induced DDIT3 causes ER stress-PERK independent apoptosis in glioblastoma multiforme cells. Oncotarget 2017, 8, 4051–4061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.L.; Lin, P.C.; Chen, S.P.; Lin, C.C.; Tsai, N.M.; Cheng, Y.L.; Chang, W.L.; Lin, S.Z.; Harn, H.J. Activation of nonsteroidal anti-inflammatory drug-activated gene-1 via extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase revealed a isochaihulactone-triggered apoptotic pathway in human lung cancer A549 cells. J. Pharm. Exp. Ther. 2007, 323, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.Y.; Kim, G.Y.; Li, W.; Choi, B.T.; Kim, N.D.; Kang, H.S.; Choi, Y.H. Implication of intracellular ROS formation, caspase-3 activation and Egr-1 induction in platycodon D-induced apoptosis of U937 human leukemia cells. Biomed. Pharm. 2009, 63, 86–94. [Google Scholar] [CrossRef]
- Ko, J.K.; Leung, W.C.; Ho, W.K.; Chiu, P. Herbal diterpenoids induce growth arrest and apoptosis in colon cancer cells with increased expression of the nonsteroidal anti-inflammatory drug-activated gene. Eur. J. Pharm. 2007, 559, 1–13. [Google Scholar] [CrossRef]
- Lim, J.H.; Park, J.W.; Min, D.S.; Chang, J.S.; Lee, Y.H.; Park, Y.B.; Choi, K.S.; Kwon, T.K. NAG-1 up-regulation mediated by EGR-1 and p53 is critical for quercetin-induced apoptosis in HCT116 colon carcinoma cells. Apoptosis 2007, 12, 411–421. [Google Scholar] [CrossRef]
- Baek, S.J.; Wilson, L.C.; Eling, T.E. Resveratrol enhances the expression of non-steroidal anti-inflammatory drug-activated gene (NAG-1) by increasing the expression of p53. Carcinogenesis 2002, 23, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Golkar, L.; Ding, X.Z.; Ujiki, M.B.; Salabat, M.R.; Kelly, D.L.; Scholtens, D.; Fought, A.J.; Bentrem, D.J.; Talamonti, M.S.; Bell, R.H.; et al. Resveratrol inhibits pancreatic cancer cell proliferation through transcriptional induction of macrophage inhibitory cytokine-1. J. Surg. Res. 2007, 138, 163–169. [Google Scholar] [CrossRef]
- Woo, M.N.; Jeon, S.M.; Shin, Y.C.; Lee, M.K.; Kang, M.A.; Choi, M.S. Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice. Mol. Nutr. Food Res. 2009, 53, 1603–1611. [Google Scholar] [CrossRef]
- Jutooru, I.; Chadalapaka, G.; Abdelrahim, M.; Basha, M.R.; Samudio, I.; Konopleva, M.; Andreeff, M.; Safe, S. Methyl 2-cyano-3,12-dioxooleana-1,9-dien-28-oate decreases specificity protein transcription factors and inhibits pancreatic tumor growth: Role of microRNA-27a. Mol. Pharmacol. 2010, 78, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Jutooru, I.; Chadalapaka, G.; Lei, P.; Safe, S. Inhibition of NFkappaB and pancreatic cancer cell and tumor growth by curcumin is dependent on specificity protein down-regulation. J. Biol. Chem. 2010, 285, 25332–25344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chintharlapalli, S.; Papineni, S.; Lei, P.; Pathi, S.; Safe, S. Betulinic acid inhibits colon cancer cell and tumor growth and induces proteasome-dependent and -independent downregulation of specificity proteins (Sp) transcription factors. BMC Cancer 2011, 11, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhy, S.U.; Kim, K.; Larsen, L.; Rosengren, R.J.; Safe, S. Curcumin and synthetic analogs induce reactive oxygen species and decreases specificity protein (Sp) transcription factors by targeting microRNAs. BMC Cancer 2012, 12, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jutooru, I.; Chadalapaka, G.; Sreevalsan, S.; Lei, P.; Barhoumi, R.; Burghardt, R.; Safe, S. Arsenic trioxide downregulates specificity protein (Sp) transcription factors and inhibits bladder cancer cell and tumor growth. Exp. Cell Res. 2010, 316, 2174–2188. [Google Scholar] [CrossRef] [Green Version]
- Pathi, S.S.; Lei, P.; Sreevalsan, S.; Chadalapaka, G.; Jutooru, I.; Safe, S. Pharmacologic doses of ascorbic acid repress specificity protein (Sp) transcription factors and Sp-regulated genes in colon cancer cells. Nutr. Cancer 2011, 63, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; O’Reilly, S.; Liang, H.; Maher, V.M.; Sleight, S.D.; McCormick, J.J. Down-regulation of overexpressed sp1 protein in human fibrosarcoma cell lines inhibits tumor formation. Cancer Res. 2005, 65, 1007–1017. [Google Scholar] [PubMed]
- Hedrick, E.; Cheng, Y.; Jin, U.H.; Kim, K.; Safe, S. Specificity protein (Sp) transcription factors Sp1, Sp3 and Sp4 are non-oncogene addiction genes in cancer cells. Oncotarget 2016, 7, 22245–22256. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.K.; Mattie, M.D.; Berger, C.E.; Benz, S.C.; Benz, C.C. Rapid alteration of microRNA levels by histone deacetylase inhibition. Cancer Res. 2006, 66, 1277–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillotson, L.G. RIN ZF, a novel zinc finger gene, encodes proteins that bind to the CACC element of the gastrin promoter. J. Biol. Chem. 1999, 274, 8123–8128. [Google Scholar] [CrossRef] [Green Version]
- Mertens-Talcott, S.U.; Chintharlapalli, S.; Li, X.; Safe, S. The oncogenic microRNA-27a targets genes that regulate specificity protein transcription factors and the G2-M checkpoint in MDA-MB-231 breast cancer cells. Cancer Res. 2007, 67, 11001–11011. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Chadalapaka, G.; Lee, S.O.; Yamada, D.; Sastre-Garau, X.; Defossez, P.A.; Park, Y.Y.; Lee, J.S.; Safe, S. Identification of oncogenic microRNA-17-92/ZBTB4/specificity protein axis in breast cancer. Oncogene 2012, 31, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Chadalapaka, G.; Jutooru, I.; Safe, S. Celastrol decreases specificity proteins (Sp) and fibroblast growth factor receptor-3 (FGFR3) in bladder cancer cells. Carcinogenesis 2012, 33, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Jutooru, I.; Guthrie, A.S.; Chadalapaka, G.; Pathi, S.; Kim, K.; Burghardt, R.; Jin, U.H.; Safe, S. Mechanism of action of phenethylisothiocyanate and other reactive oxygen species-inducing anticancer agents. Mol. Cell. Biol. 2014, 34, 2382–2395. [Google Scholar] [CrossRef] [Green Version]
- Kasiappan, R.; Jutooru, I.; Karki, K.; Hedrick, E.; Safe, S. Benzyl Isothiocyanate (BITC) induces reactive oxygen species-dependent repression of STAT3 Protein by down-regulation of specificity proteins in pancreatic cancer. J. Biol. Chem. 2016, 291, 27122–27133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karki, K.; Hedrick, E.; Kasiappan, R.; Jin, U.H.; Safe, S. Piperlongumine induces Reactive Oxygen Species (ROS)-Dependent Downregulation of specificity protein transcription factors. Cancer Prev. Res. Phila 2017, 10, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hagan, H.M.; Wang, W.; Sen, S.; Destefano Shields, C.; Lee, S.S.; Zhang, Y.W.; Clements, E.G.; Cai, Y.; Van Neste, L.; Easwaran, H.; et al. Oxidative damage targets complexes containing DNA methyltransferases, SIRT1, and polycomb members to promoter CpG Islands. Cancer Cell 2011, 20, 606–619. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Zhang, B.; Su, W.W.; Williams, P.G.; Li, Q.X. C-Glycosylflavones Alleviate Tau Phosphorylation and Amyloid Neurotoxicity through GSK3beta Inhibition. ACS Chem. Neurosci. 2016, 7, 912–923. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Li, Q.X. Discovery of selective, substrate-competitive, and passive membrane permeable glycogen synthase kinase-3beta inhibitors: Synthesis, biological evaluation, and molecular modeling of new C-Glycosylflavones. ACS Chem. Neurosci. 2018, 9, 1166–1183. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Liang, Z.; Li, Y.; Zhi, Y.; Yi, L.; Bai, S.; Forest, K.H.; Nichols, R.A.; Dong, Y.; Li, Q.X. Isoorientin, a GSK-3β inhibitor, rescues synaptic dysfunction, spatial memory deficits and attenuates pathological progression in APP/PS1 model mice. Behav. Brain Res. 2021, 398, 112968. [Google Scholar] [CrossRef] [PubMed]
- Doello, S.; Liang, Z.; Cho, I.K.; Kim, J.B.; Li, Q.X. Cytotoxic effects of 24-Methylenecyloartanyl ferulate on A549 nonsmall cell lung cancer cells through MYBBP1A up-regulation and AKT and aurora B kinase inhibition. J. Agric. Food Chem. 2018, 66, 3726–3733. [Google Scholar] [CrossRef]
- Kim, H.; Kim, D.; Jang, G.; Lee, S.; Jang, H.; Cho, I.; Li, Q.X.; Lee, S.; Kim, J. Purification of cycloartenyl ferulate, 24-methylenecycloartanyl ferulate, campesteryl ferulate and sitosteryl ferulate from rice bran and their effects on the NLRP3 inflammasome. Acad. J. Agric. Res. 2016, 4, 411–419. [Google Scholar] [CrossRef]
- Chang, C.L.; Cho, I.K.; Li, Q.X. Insecticidal activity of basil oil, trans-anethole, estragole, and linalool to adult fruit flies of Ceratitis capitata, Bactrocera dorsalis, and Bactrocera cucurbitae. J. Econ. Entomol. 2009, 102, 203–209. [Google Scholar] [CrossRef]
- Li, A.S.; Iijima, A.; Huang, J.; Li, Q.X.; Chen, Y. Putative mode of action of the monoterpenoids linalool, methyl eugenol, estragole, and citronellal on ligand-gated ion channels. Engineering 2020, 6, 541–545. [Google Scholar] [CrossRef]
- Sun, B.T.D.; Pan, D.; Baker, M.R.; Liang, Z.; Wang, Z.; Lei, J.; Liu, S.; Hu, C.Y.; Li, Q.X. Dihydromyricetin imbues anti-adipogenic effects on 3T3-L1 cells via direct interactions with 78-kDa glucose regulated protein. J. Nutr. 2021. [Google Scholar] [CrossRef]
- Boon, C.S.; Mcclements, D.J.; Weiss, J.; Decker, E.A. Factors influencing the chemical stability of carotenoids in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Pan, K.; Zhong, Q. Organic nanoparticles in foods: Fabrication, characterization, and utilization. Annu. Rev. Food Sci. Technol. 2016, 7, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Baskar, V.; Selvaraj, D.; Nile, A.; Xiao, J.; Kai, G. Nanotechnologies in food science: Applications, recent trends, and future perspectives. Nano-Micro Lett. 2020, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- McClements, D.J. Nanotechnology approaches for improving the healthiness and sustainability of the modern food supply. ACS Omega 2020, 5, 29623–29630. [Google Scholar] [CrossRef] [PubMed]


| Phytochemical | Animal | Mechanism of Action | Reference |
|---|---|---|---|
| Ascorbic acid | Rat | Downregulation of MMP-2 and MMP-9 | [145] |
| Astragaloside IV | Rat | Downregulation of MMP-9 and AQP4 | [147] |
| Baicalin | Rat | Downregulation of MMP-9 | [152] |
| Chlorogenic acid | Rat | Downregulation of MMP-2 and MMP-9 | [140] |
| Crocin | Rat | Downregulation of MMP-2 and MMP-9 | [141] |
| Curcumin | Rat | Downregulation of MMP-9 | [153] |
| Dl-3-n-butylphthalide | Mouse | Downregulation of Caveolin-1 | [154] |
| Ellagic acid | Rat | Downregulation of AQP4 and MMP-9 | [148] |
| Gastrodin | Rat | Downregulation of MMP-2 and MMP-9 | [144] |
| Ginsenoside Rb1 | Mouse | Downregulation of MMP-9 | [155] |
| Ginsenoside Rd | Rat | Downregulation of NF-κB and MMP-9 | [156] |
| Hesperidin | Mouse | Inhibition of FoxO3a nuclear translocation Downregulation of MMP-3/9 | [150] |
| Icariside II | Rat | Downregulation of MMP-9 Upregulation of TIMP-1 | [157] |
| Juglanin | Mouse | Downregulation of VEGF and VEGFR2 | [151] |
| Melatonin | Rat | Downregulation of MMP-9 | [158] |
| Pinocembrin | Rat | Downregulation of MMP-2 and MMP-9 | [142] |
| Quercetin | Rat | Downregulation of MMP-9 | [159] |
| Resveratrol | Rat | Downregulation of MMP-9 Upregulation of TIMP-1 | [160] |
| Rutin | Rat | Downregulation of MMP-9 | [161] |
| Salvianolic acid A | Rat | Downregulation ofMMP-9 Upregulation of TIMP-1 Src phosphorylation at Tyr416 | [143,146] |
| Sodium tanshinone IIA sulfonate (with rt-PA) | Human | Downregulation of MMP-9 and TIMP-1 | [139] |
| Tetrahydrocurcumin | Mouse | Downregulation of MMP-9 Upregulation of TIMP-2 | [162] |
| Tetramethylpyrazine | Rat | Downregulation of JAK/STAT phosphorylation | [163] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.J.; Hammock, B.D.; Hwang, I.-K.; Li, Q.X.; Moustaid-Moussa, N.; Park, Y.; Safe, S.; Suh, N.; Yi, S.-S.; Zeldin, D.C.; et al. Natural Products in the Prevention of Metabolic Diseases: Lessons Learned from the 20th KAST Frontier Scientists Workshop. Nutrients 2021, 13, 1881. https://doi.org/10.3390/nu13061881
Baek SJ, Hammock BD, Hwang I-K, Li QX, Moustaid-Moussa N, Park Y, Safe S, Suh N, Yi S-S, Zeldin DC, et al. Natural Products in the Prevention of Metabolic Diseases: Lessons Learned from the 20th KAST Frontier Scientists Workshop. Nutrients. 2021; 13(6):1881. https://doi.org/10.3390/nu13061881
Chicago/Turabian StyleBaek, Seung J., Bruce D. Hammock, In-Koo Hwang, Qing X. Li, Naima Moustaid-Moussa, Yeonhwa Park, Stephen Safe, Nanjoo Suh, Sun-Shin Yi, Darryl C. Zeldin, and et al. 2021. "Natural Products in the Prevention of Metabolic Diseases: Lessons Learned from the 20th KAST Frontier Scientists Workshop" Nutrients 13, no. 6: 1881. https://doi.org/10.3390/nu13061881