
Breastfeeding Practices Influence the Breast Milk Microbiota Depending on Pre-Gestational Maternal BMI and Weight Gain over Pregnancy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Mother-Infant Pairs
2.2. Breast Milk Samples and DNA Extraction
2.3. Total Bacterial Load by qPCR
2.4. S rRNA Amplicon Sequencing and Bioinformatics
2.5. Statistical Analysis
3. Results
3.1. Subjects and Clinical Data
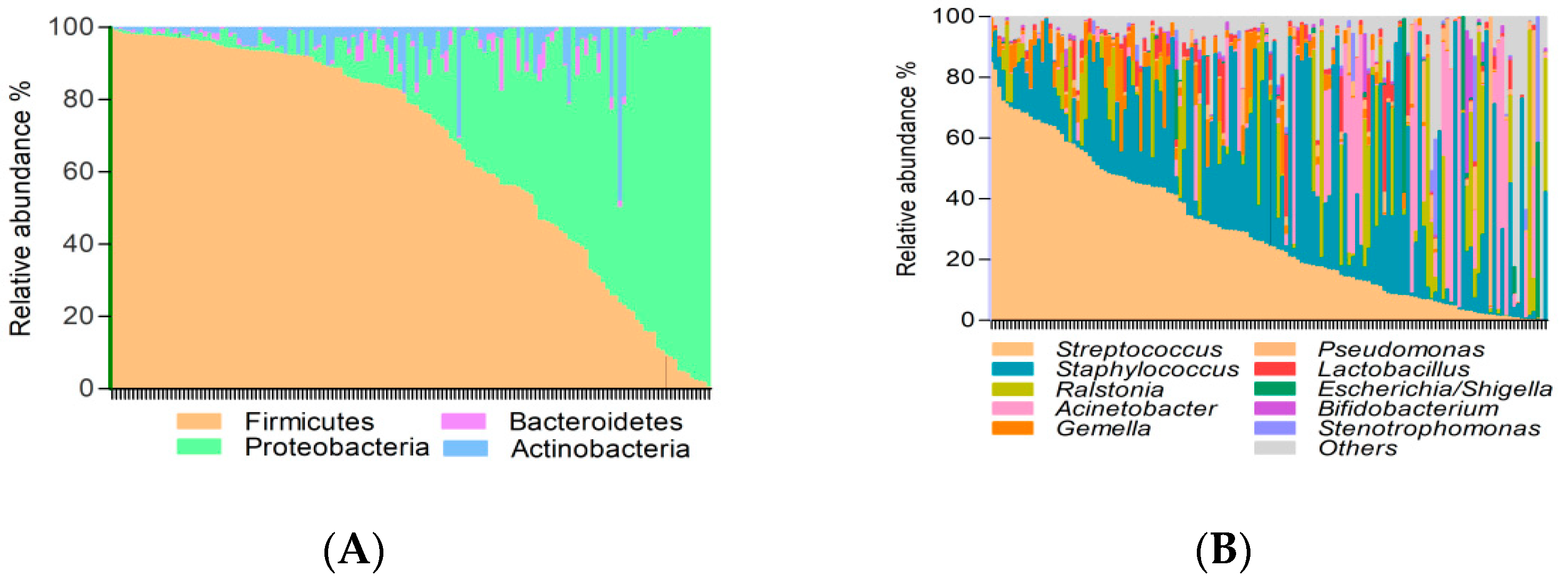
3.2. Factors Affecting Breast Milk Microbiota
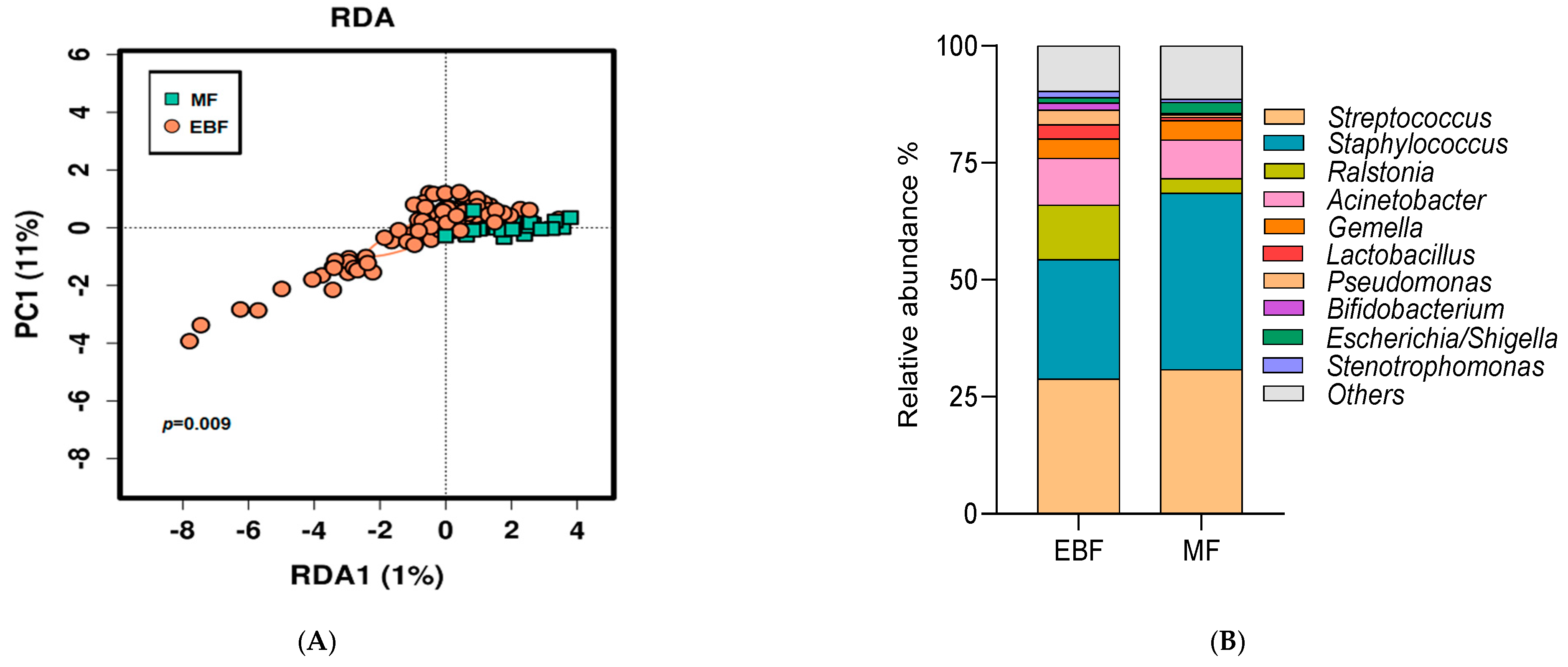
3.3. Breast Milk Microbiota Was Shaped by Breastfeeding Practices
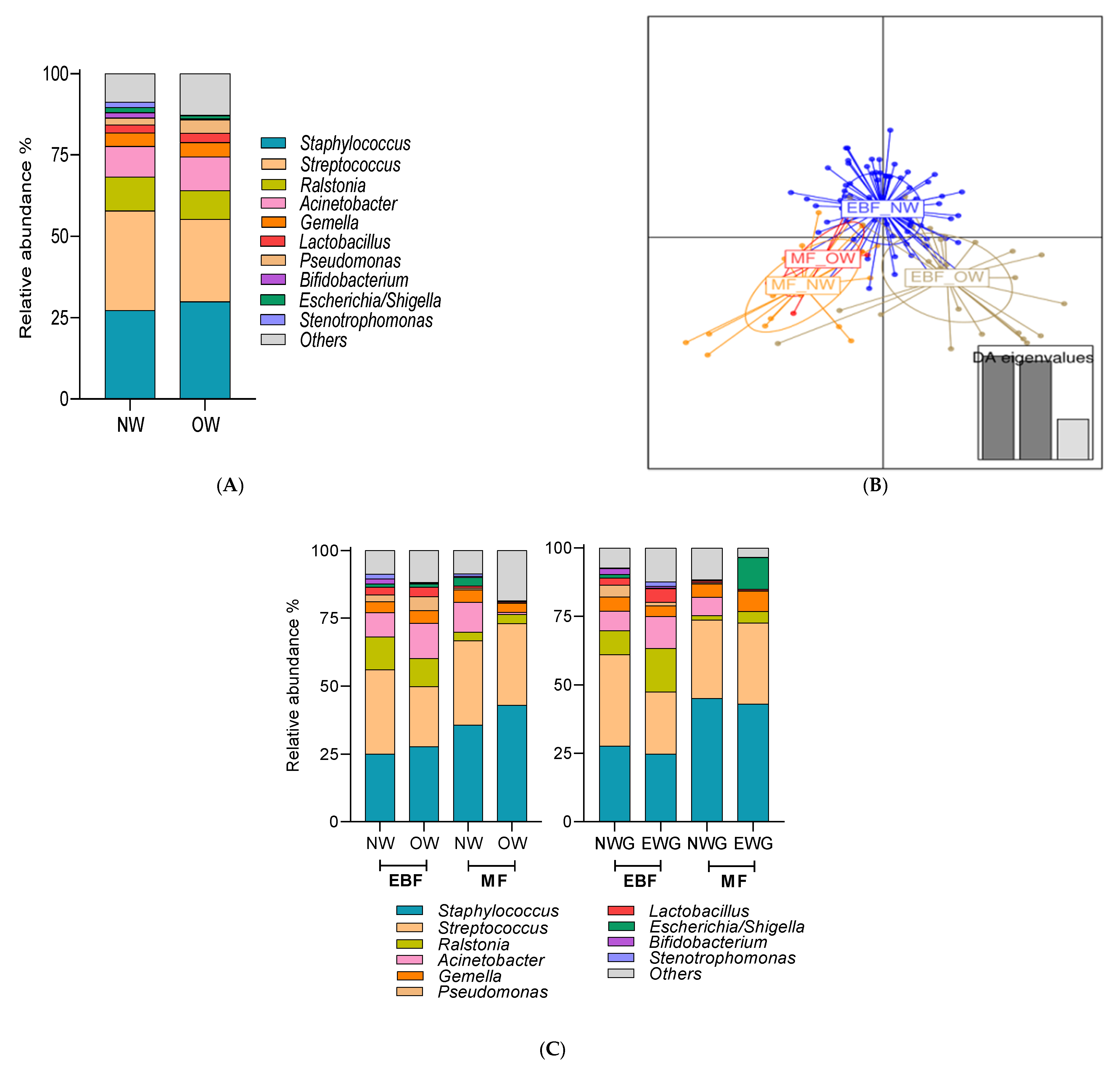
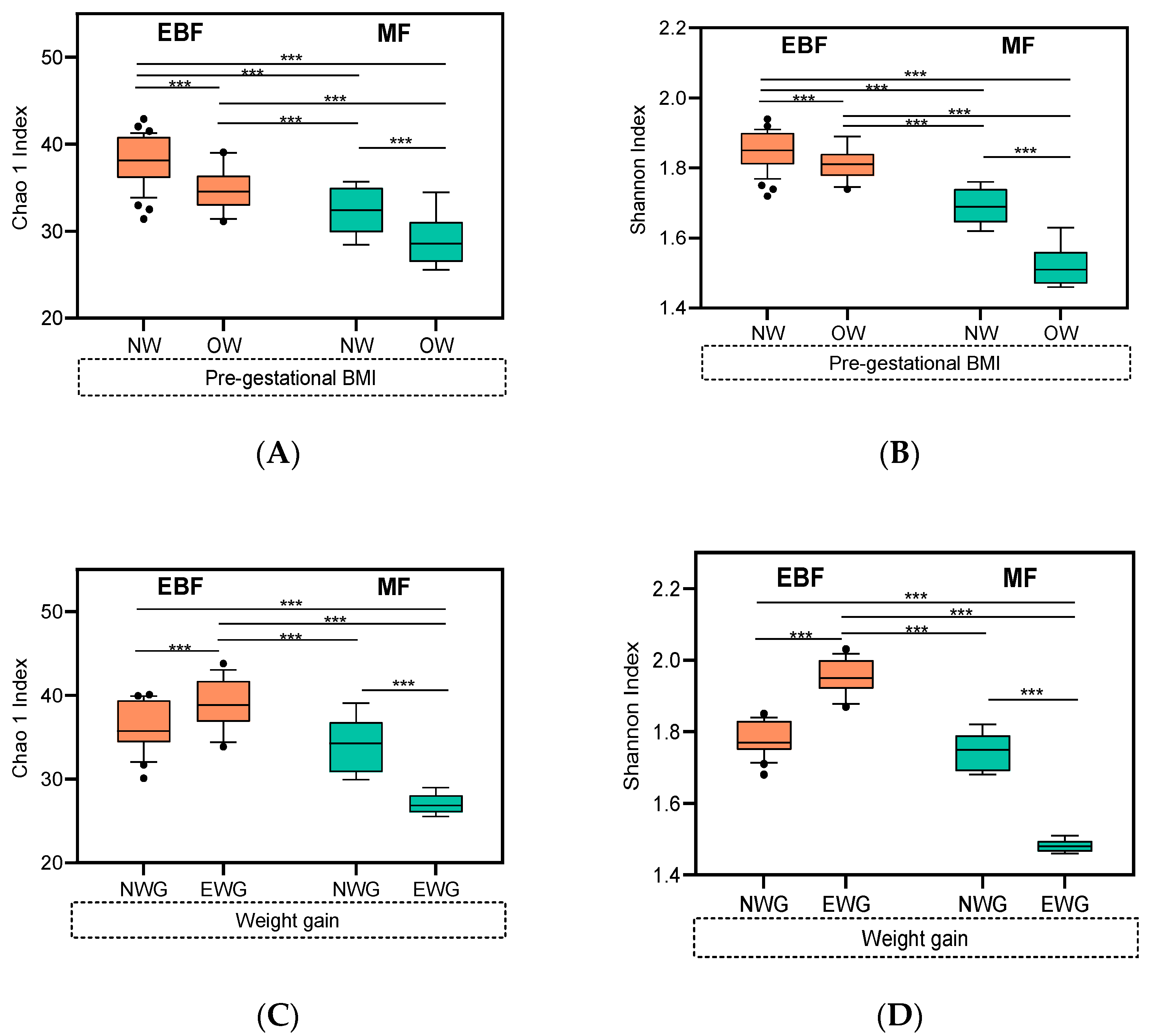
3.4. Impact of Breastfeeding Practices on Breast Milk Microbiota Composition Was Modulated by Pre-Gestational BMI and Weight Gain over Pregnancy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A purposeful contribution to the development of the infant microbiota and immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Exclusive Breastfeeding for Six Months Best for Babies Everywhere; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Hoppu, U.; Kalliomäki, M.; Laiho, K.; Isolauri, E. Breast milk—Immunomodulatory signals against allergic diseases. Allergy Eur. J. Allergy Clin. Immunol. 2001, 56, 23–26. [Google Scholar] [CrossRef]
- Pastor-Villaescusa, B.; Hurtado, J.A.; Gil-Campos, M.; Uberos, J.; Maldonado-Lobón, J.A.; Díaz-Ropero, M.P.; Bañuelos, O.; Fonollá, J.; Olivares, M.; the PROLAC Group. Effects of Lactobacillus fermentum CECT5716 Lc40 on infant growth and health: A randomised clinical trial in nursing women. Benef. Microbes 2020, 11, 235–244. [Google Scholar] [CrossRef]
- Jakaitis, B.M.; Denning, P.W. Human breast milk and the gastrointestinal innate immune system. Clin. Perinatol. 2014, 41, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Hermansson, H.; Kumar, H.; Collado, M.C.; Salminen, S.; Isolauri, E.; Rautava, S. Breast milk microbiota is shaped by mode of delivery and intrapartum antibiotic exposure. Front. Nutr. 2019, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, P.D.; Aparicio, M.; Alba, C.; Hechler, C.; Beijers, R.; Rodríguez, J.M.; Fernández, L.; de Weerth, C. Human milk microbiome and maternal postnatal psychosocial distress. Front. Microbiol. 2019, 10, 2333. [Google Scholar] [CrossRef]
- Cortes-Macías, E.; Selma-Royo, M.; García-Mantrana, I.; Calatayud, M.; González, S.; Martínez-Costa, C.; Collado, M.C. Maternal diet shapes the breast milk microbiota composition and diversity: Impact of mode of delivery and antibiotic exposure. J. Nutr. 2021, 151, 330–340. [Google Scholar] [CrossRef]
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and variation of the human milk microbiota are influenced by maternal and early-life factors. Cell Host Microbe 2019, 25, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.E.; Carrothers, J.M.; Lackey, K.A.; Beatty, N.F.; York, M.A.; Brooker, S.L.; Shafii, B.; Price, W.J.; Settles, M.L.; McGuire, M.A.; et al. Human milk microbial community structure is relatively stable and related to variations in macronutrient and micronutrient intakes in healthy lactating women. J. Nutr. 2017, 147, 1739–1748. [Google Scholar]
- Drago, L.; Toscano, M.; De Grandi, R.; Grossi, E.; Padovani, E.M.; Peroni, D.G. Microbiota network and mathematic microbe mutualism in colostrum and mature milk collected in two different geographic areas: Italy versus Burundi. ISME J. 2017, 11, 875–884. [Google Scholar] [CrossRef]
- Urbaniak, C.; Angelini, M.; Gloor, G.B.; Reid, G. Human milk microbiota profiles in relation to birthing method, gestation and infant gender. Microbiome 2016, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, S.N.; Madan, J.C.; Karagas, M.R.; Morrison, H.G.; Hoen, A.G.; Christensen, B.C. Microbial communities in human milk relate to measures of maternal weight. Front. Microbiol. 2019, 10, 2886. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 2016, 21, 400–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butts, C.A.; Paturi, G.; Blatchford, P.; Bentley-Hewitt, K.L.; Hedderley, D.I.; Martell, S.; Dinnan, H.; Eady, S.L.; Wallace, A.J.; Glyn-Jones, S.; et al. Microbiota composition of breast milk from women of different ethnicity from the Manawatu—Wanganui region of New Zealand. Nutrients 2020, 12, 1756. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Jiang, J.; Lu, M.; Tong, W.; Zhou, R.; Li, J.; Yuan, J.; Wang, F.; Li, D. Human milk microbiota development during lactation and its relation to maternal geographic location and gestational hypertensive status. Gut Microbes 2020, 11, 1438–1449. [Google Scholar] [CrossRef] [PubMed]
- Shenker, N.S.; Perdones-Montero, A.; Burke, A.; Stickland, S.; McDonald, J.A.K.; Alexander-Hardiman, K.; Flanagan, J.; Takats, Z.; Cameron, S.J.S. Metabolomic and metataxonomic fingerprinting of human milk suggests compositional stability over a natural term of breastfeeding to 24 months. Nutrients 2020, 12, 3450. [Google Scholar] [CrossRef] [PubMed]
- García-Mantrana, I.; Alcántara, C.; Selma-Royo, M.; Boix-Amorós, A.; Dzidic, M.; Gimeno-Alcañiz, J.; Úbeda-Sansano, I.; Sorribes-Monrabal, I.; Escuriet, R.; Gil-Raga, F.; et al. MAMI: A birth cohort focused on maternal-infant microbiota during early life. BMC Pediatrics 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic; Report of a WHO; Consultation No. 894; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- National Researh Council. Weight Gain during Pregnancy: Reexamining the Guidelines; The National Academies Press: Washington, DC, USA, 2009. [Google Scholar] [CrossRef]
- Boix-Amorós, A.; Collado, M.C.; Mira, A. Relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 2016, 7, 492. [Google Scholar] [CrossRef] [Green Version]
- Cruaud, P.; Vigneron, A.; Lucchetti-Miganeh, C.; Ciron, P.E.; Godfroy, A.; Cambon-Bonavita, M.A. Influence of DNA extraction method, 16S rRNA targeted hypervariable regions, and sample origin on microbial diversity detected by 454 pyrosequencing in marine chemosynthetic ecosystems. Appl. Environ. Microbiol. 2014, 80, 4626–4639. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- García-Mantrana, I.; Selma-Royo, M.; González, S.; Parra-Llorca, A.; Martínez-Costa, C.; Collado, M.C. Distinct maternal microbiota clusters are associated with diet during pregnancy: Impact on neonatal microbiota and infant growth during the first 18 months of life. Gut Microbes 2020, 11, 962–978. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome—Environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IBM Corp. Released 2020. IBM SPSS Statistics for Windows, Version 27.0; IBM Corp: Armonk, NY, USA, 2020. [Google Scholar]
- Allaire, J.J. RStudio: Integrated Development for R; R Studio Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Grote, V.; Verduci, E.; Scaglioni, S.; Vecchi, F.; Contarini, G.; Giovannini, M.; Koletzko, B.; Agostoni, C. Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur. J. Clin. Nutr. 2016, 70, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moossavi, S.; Miliku, K.; Sepehri, S.; Khafipour, E.; Azad, M.B. The prebiotic and probiotic properties of human milk: Implications for infant immune development and pediatric asthma. Front. Pediatr. 2018, 6, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toscano, M.; De Grandi, R.; Grossi, E.; Drago, L. Role of the human breast milk-associated microbiota on the newborns’ immune system: A mini review. Front. Microbiol. 2017, 8, 2100. [Google Scholar] [CrossRef] [PubMed]
- Ladomenou, F.; Moschandreas, J.; Kafatos, A.; Tselentis, Y.; Galanakis, E. Protective effect of exclusive breastfeeding against infections during infancy: A prospective study. Arch. Dis. Child. 2010, 95, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, S.; Song, Y.; Feng, Y.; Lv, N.; Xue, Y.; Liu, F.; Gosalbes, M.J. The perturbation of infant gut microbiota caused by cesarean delivery is partially restored by exclusive breastfeeding. Front. Microbiol. 2019, 10, 598. [Google Scholar] [CrossRef] [Green Version]
- Ho, N.T.; Li, F.; Lee-sarwar, K.A.; Tun, H.M.; Brown, B.P.; Pannaraj, P.S.; Bender, J.M.; Azad, M.B.; Thompson, A.L.; Weiss, S.T.; et al. Meta-analysis of effects of exclusive breastfeeding on infant gut microbiota across populations. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Azad, M.B.; Vehling, L.; Tun, H.M.; Konya, T.B.; Guttman, D.S.; Field, C.J.; Lefebvre, D.; Sears, M.R.; Becker, A.B.; et al. Association of exposure to formula in the hospital and subsequent infant feeding practices with gut microbiota and risk of overweight in the first year of life. JAMA 2018, 172, e181161. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.; Peura, S.; Hammar, U.; Vicenzi, S.; Hedman, A.; Almqvist, C.; Andolf, E.; Pershagen, G.; Dicksved, J. Oral microbiota development in early childhood. Sci. Rep. 2019, 9, 19025. [Google Scholar] [CrossRef] [PubMed]
- Holgerson, P.L.; Vestman, N.R.; Claesson, R.; Öhman, C.; Domellöf, M.; Tanner, A.C.R.; Hernell, O.; Johansson, I. Oral microbial profile discriminates breastfed fron formula-fed infants. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Holgerson, P.L.; Esberg, A.; Sjödin, A.; West, C.E.; Johansson, I. A longitudinal study of the development of the saliva microbiome in infants 2 days to 5 years compared to the microbiome in adolescents. Sci. Rep. 2020, 10, 9629. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, M.; Collado, M.C.; Abrahamsson, T.; Artacho, A.; Stensson, M.; Jenmalm, M.C.; Mira, A. Oral microbiome development during childhood: An ecological succession influenced by postnatal factors and associated with tooth decay. ISME J. 2018, 12, 2292–2306. [Google Scholar] [CrossRef]
- Timby, N.; Domellöf, M.; Holgerson, P.L.; West, C.E.; Lönnerdal, B.; Hernell, O.; Johansson, I. Oral microbiota in infants fed a formula supplemented with bovine milk fat globule membranes—A randomized controlled trial. PLoS ONE 2017, 12, e0169831. [Google Scholar] [CrossRef] [PubMed]
- Al-Shehri, S.S.; Sweeney, E.L.; Cowley, D.M.; Liley, H.G.; Ranasinghe, P.D.; Charles, B.G.; Shaw, P.N.; Vagenas, D.; Duley, J.A.; Knox, C.L. Deep sequencing of the 16S ribosomal RNA of the neonatal oral microbiome: A comparison of breast-fed and formula-fed infants. Sci. Rep. 2016, 6, 38309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeMay-Nedjelski, L.; Asbury, M.R.; Butcher, J.; Ley, S.H.; Hanley, A.J.; Kiss, A.; Unger, S.; Copeland, J.K.; Wang, P.W.; Stintzi, A.; et al. Maternal Diet and Infant Feeding Practices Are Associated with Variation in the Human Milk Microbiota at 3 Months Postpartum in a Cohort of Women with High Rates of Gestational Glucose Intolerance. J. Nutr. 2021, 151, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Solís, G.; de los Reyes-Gavilan, C.G.; Fernández, N.; Margolles, A.; Gueimonde, M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe 2010, 16, 307–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef]
- Lemay-Nedjelski, L.; Butcher, J.; Ley, S.H.; Asbury, M.R.; Hanley, A.J.; Kiss, A.; Unger, S.; Copeland, J.K.; Wang, P.W.; Zinman, B.; et al. Examining the relationship between maternal body size, gestational glucose tolerance status, mode of delivery and ethnicity on human milk microbiota at three months post-partum. BMC Microbiol. 2020, 20, 219. [Google Scholar] [CrossRef]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Marítn-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Heikkilä, M.P.; Saris, P.E.J. Inhibition of Staphylococcus aureus by the commensal bacteria of human milk. J. Appl. Microbiol. 2003, 95, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatr. Res. 2012, 72, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.W.; Watanabe, K.; Hsu, C.C.; Chao, S.H.; Yang, Z.H.; Lin, Y.J.; Chen, C.C.; Cao, Y.M.; Huang, H.C.; Chang, C.H.; et al. Bacterial composition and diversity in breast milk samples from mothers living in Taiwan and Mainland China. Front. Microbiol. 2017, 8, 965. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selma-Royo, M.; Calatayud, M.; García-Mantrana, I.; Parra-Llorca, A.; Escuriet, R.; Martínez-Costa, C.; Collado, M.C. Perinatal environment shapes microbiota colonization and infant growth: Impact on host response and intestinal function. Microbiome 2020, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Haugen, M.; Brantsæter, A.L.; Winkvist, A.; Lissner, L.; Alexander, J.; Oftedal, B.; Magnus, P.; Meltzer, H.M. Associations of pre-pregnancy body mass index and gestational weight gain with pregnancy outcome and postpartum weight retention: A prospective observational cohort study. BMC Pregnancy Childbirth 2014, 14, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.E.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Jewell, S.; Grummer-Strawn, L. Maternal obesity and breast-feeding practices 1,2. Am. J. Clin. Nutr. 2003, 77, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojo-Okunola, A.; Claassen-Weitz, S.; Mwaikono, K.S.; Gardner-Lubbe, S.; Stein, D.J.; Zar, H.J.; Nicol, M.P.; Du Toit, E. Influence of socio-economic and psychosocial profiles on the human breast milk bacteriome of south african women. Nutrients 2019, 11, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilha, M.; Brejnrod, A.; Danneskiold-Samsøe, N.B.; Hoffmann, C.; Iaucci, J.d.M.; Cabral, V.P.; Xavier-Santos, D.; Taddei, C.R.; Kristiansen, K.; Saad, S.M.I. Response of the human milk microbiota to a maternal prebiotic intervention is individual and influenced by maternal age. Nutrients 2020, 12, 1081. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
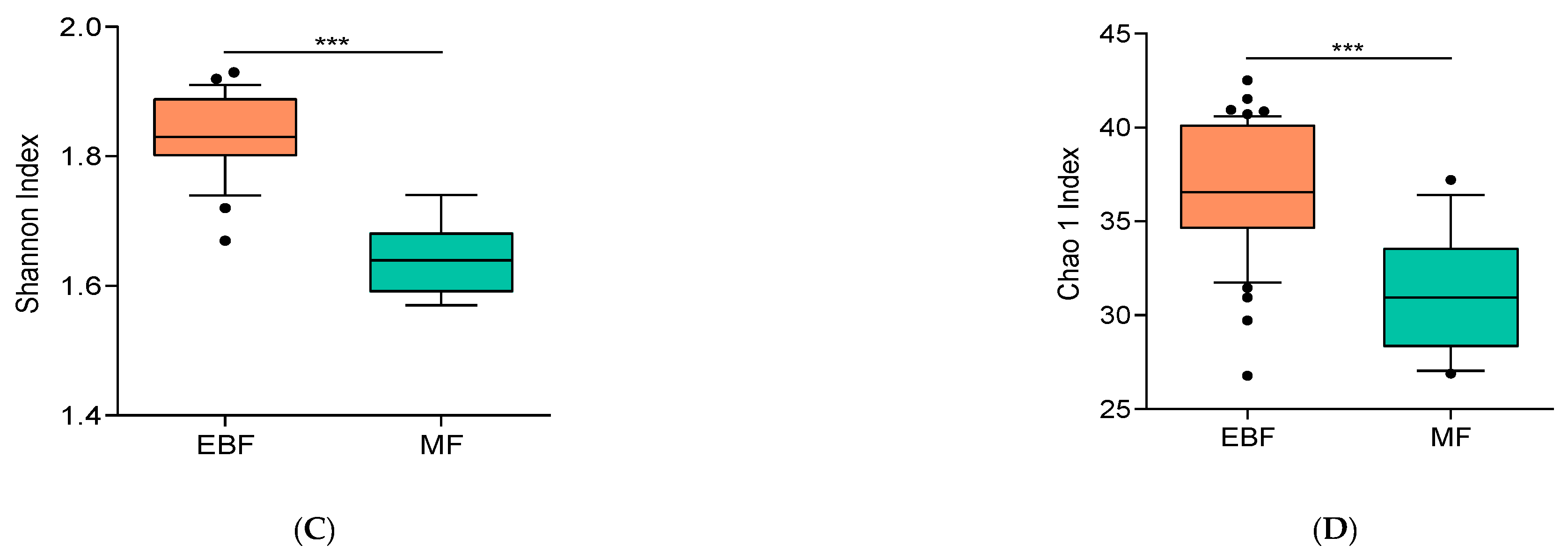{kind=link}
{kind=link}
{kind=link}
| All | Exclusive Breastfeeding (n = 111) | Mixed Feeding (n = 25) | p | |
|---|---|---|---|---|
| Maternal characteristics | ||||
| Maternal age (years) | 34.44 ± 3.79 | 34.39 ± 3.84 | 34.68 ± 3.61 | 0.728 |
| Gestational age (weeks) | 40 (39,40) | 40 (39–41) | 39 (39,40) | 0.005 |
| Pre-gestational BMI (Kg/m2) * | 22.84 (21.01–25.39) | 22.62 (20.96–25.42) | 23.01 (21.29–25.60) | 0.386 |
| Normal weight (NW) | 97 (71.3) | 79 (71.2) | 18 (72.0) | 0.985 |
| Over weight (OW) | 38 (27.9) | 31 (27.9) | 7 (28.0) | |
| Weight gain (kg) over pregnancy ǂ | 12 (9.5–15) | 12 (10–15) | 10 (7.8–14) | 0.874 |
| Low weight gain (LWG) | 43 (31.6) | 35 (31.5) | 8 (32.0) | 0.876 |
| Normal weight gain (NWG) | 60 (44.1) | 48 (43.2) | 12 (48.0) | |
| Excessive weight gain (EWG) | 32 (23.5) | 27 (24.3) | 5 (20.0) | |
| Intrapartum antibiotic exposure (%) | 54 (39.7) | 46 (41.4) | 8 (32.0) | 0.383 |
| Antibiotic during pregnancy (%) | 42 (30.9) | 34 (30.6) | 8 (32.0) | 0.915 |
| Antibiotic treatment during 1 month (%) | 11 (8.1) | 8 (7.2) | 3 (12.0) | 0.435 |
| Infant characteristics | ||||
| Gender: Female (%) | 76 (55.9) | 59 (53.2) | 17 (68.0) | 0.177 |
| Birth mode: vaginal birth (%) | 85 (62.5) | 68 (61.3) | 17 (68.0) | 0.530 |
| Height at birth (cm) | 49.86 ± 2.10 | 50.12 ± 2.07 | 48.74 ± 1.89 | 0.003 |
| Weight at birth (g) | 3.32 ± 0.44 | 3.36 ± 0.42 | 3.12 ± 0.49 | 0.013 |
| Antibiotic treatment 1 month (%) | 8 (5.9) | 7 (6.3) | 1 (4.0) | 0.658 |
| Total breastfeeding duration (months) | 8.37 ± 3.92 | 9.41 ± 3.24 | 3.76± 3.32 | <0.001 |
| Breastfeeding Practices One Month | |||||
|---|---|---|---|---|---|
| Genus | Rel.abund (%) | IRR | 95% CI | p | |
| Streptococcus | 29.19 | EBF vs. MF | 0.92 | 0.85–1.00 | 0.033 |
| Staphylococcus | 27.78 | EBF vs. MF | 0.69 | 0.64–0.74 | <0.001 |
| Ralstonia | 10.11 | EBF vs. MF | 3.72 | 2.94–4.72 | <0.001 |
| Acinetobacter | 9.64 | EBF vs. MF | 1.26 | 1.08–1.47 | 0.003 |
| Gemella | 4.20 | EBF vs. MF | 0.95 | 0.76–1.20 | 0.678 |
| Pseudomonas | 2.69 | EBF vs. MF | 9.60 | 4.53–20.28 | <0.001 |
| Lactobacillus | 2.57 | EBF vs. MF | 6.43 | 3.42-12.10 | <0.001 |
| Escherichia/Shigella | 1.40 | EBF vs. MF | 0.42 | 0.31–0.58 | <0.001 |
| Bifidobacterium | 1.25 | EBF vs. MF | 9.52 | 3.03–29.92 | <0.001 |
| Stenotrophomonas | 1.23 | EBF vs. MF | 1.33 | 0.77–2.28 | 0.303 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés-Macías, E.; Selma-Royo, M.; Martínez-Costa, C.; Collado, M.C. Breastfeeding Practices Influence the Breast Milk Microbiota Depending on Pre-Gestational Maternal BMI and Weight Gain over Pregnancy. Nutrients 2021, 13, 1518. https://doi.org/10.3390/nu13051518
Cortés-Macías E, Selma-Royo M, Martínez-Costa C, Collado MC. Breastfeeding Practices Influence the Breast Milk Microbiota Depending on Pre-Gestational Maternal BMI and Weight Gain over Pregnancy. Nutrients. 2021; 13(5):1518. https://doi.org/10.3390/nu13051518
Chicago/Turabian StyleCortés-Macías, Erika, Marta Selma-Royo, Cecilia Martínez-Costa, and Maria Carmen Collado. 2021. "Breastfeeding Practices Influence the Breast Milk Microbiota Depending on Pre-Gestational Maternal BMI and Weight Gain over Pregnancy" Nutrients 13, no. 5: 1518. https://doi.org/10.3390/nu13051518