
Placental Antioxidant Defenses and Autophagy-Related Genes in Maternal Obesity and Gestational Diabetes Mellitus

 , , , , ,
, , , , ,  , , , and
, , , and
Abstract
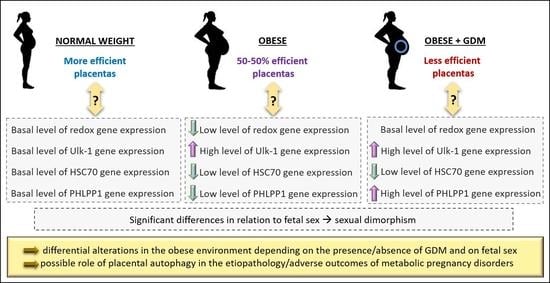
:
1. Introduction
2. Materials and Methods
2.1. Population
- Normal Weight (NW) women: 18 ≤ BMI < 25, n = 16;
- Obese (OB) women: BMI ≥ 30, n = 31.
- NW women: 11.5 ≤ GWG ≤ 16;
- OB women: 5 ≤ GWG ≤ 9.
2.2. Placental Tissue Sample Collection
2.3. Gene Expression Analysis
2.4. Statistical Analysis
3. Results
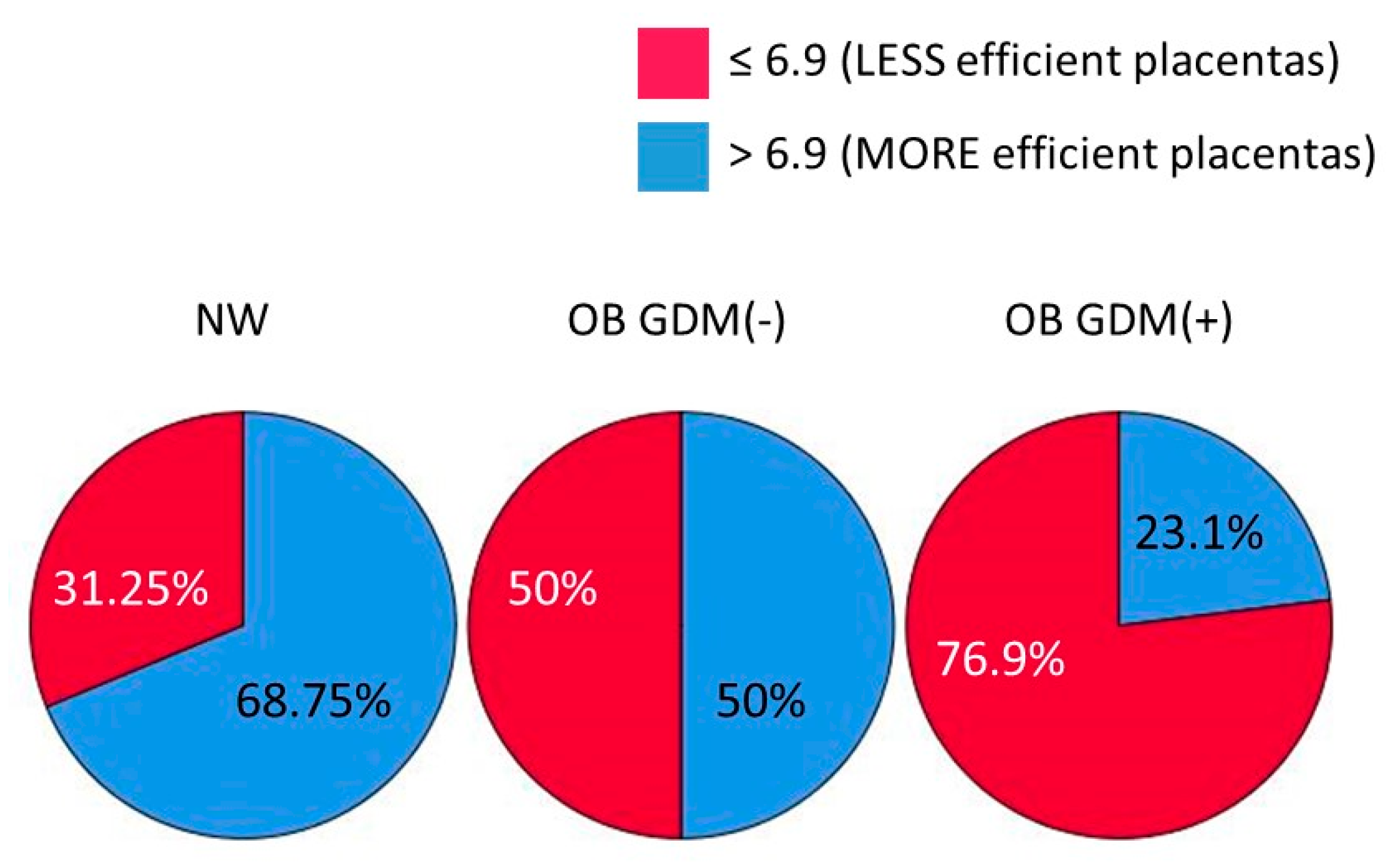
3.1. Clinical Data
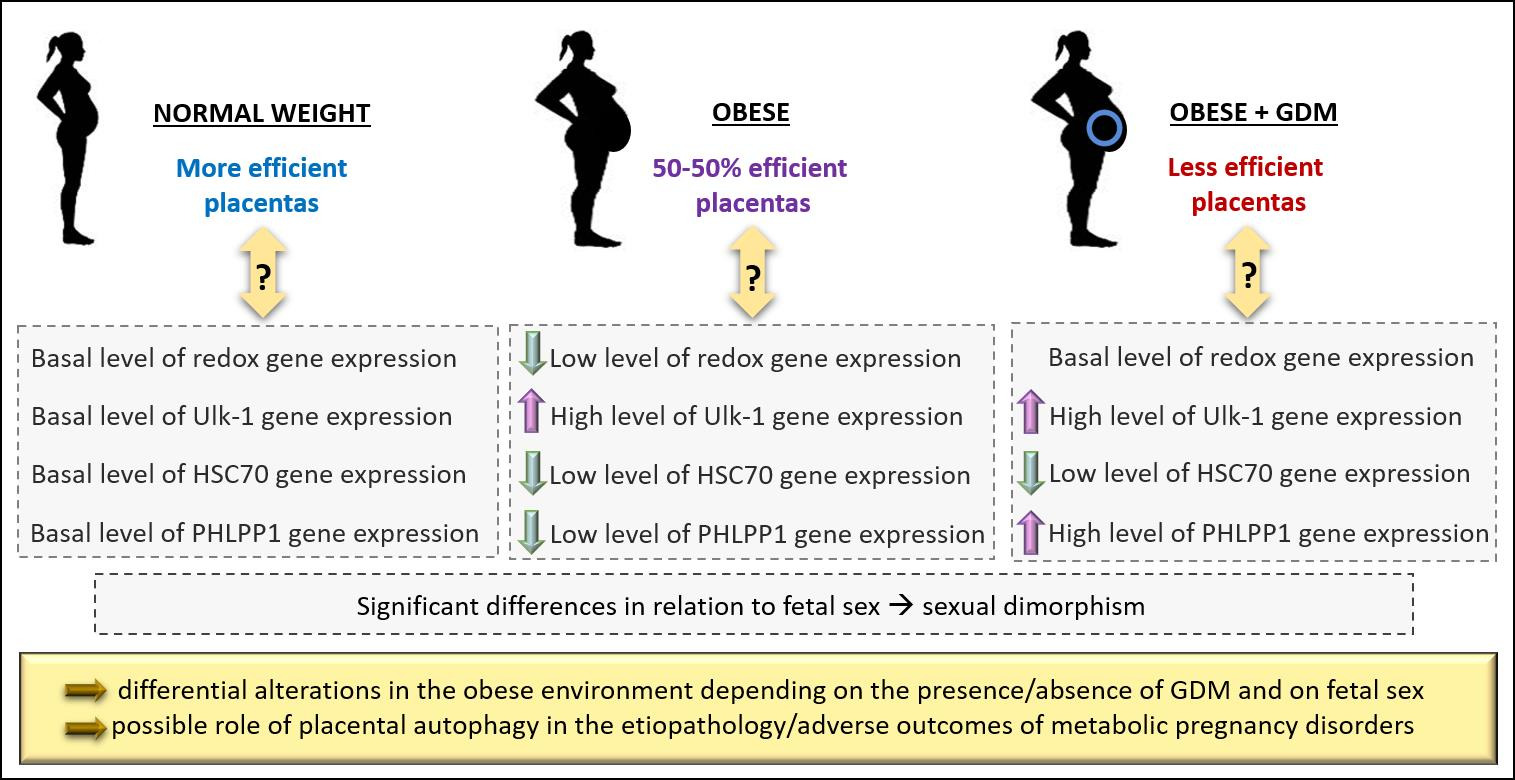
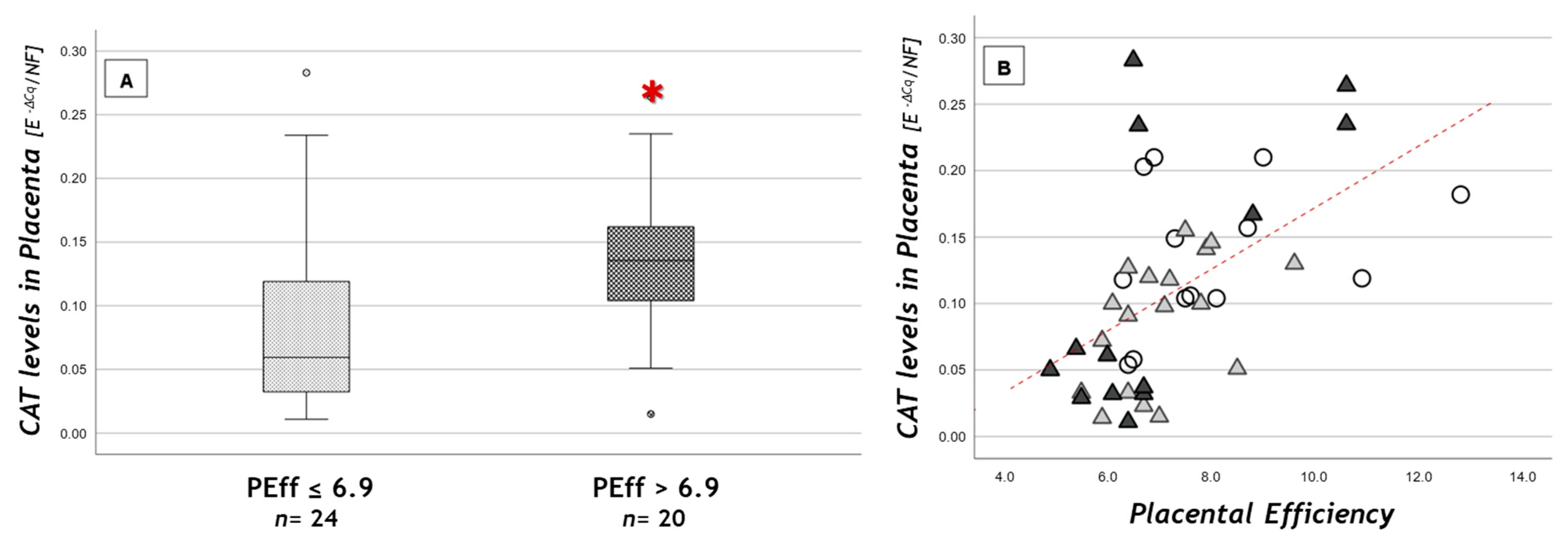
3.2. Expression of Antioxidant Defense Genes in Placenta
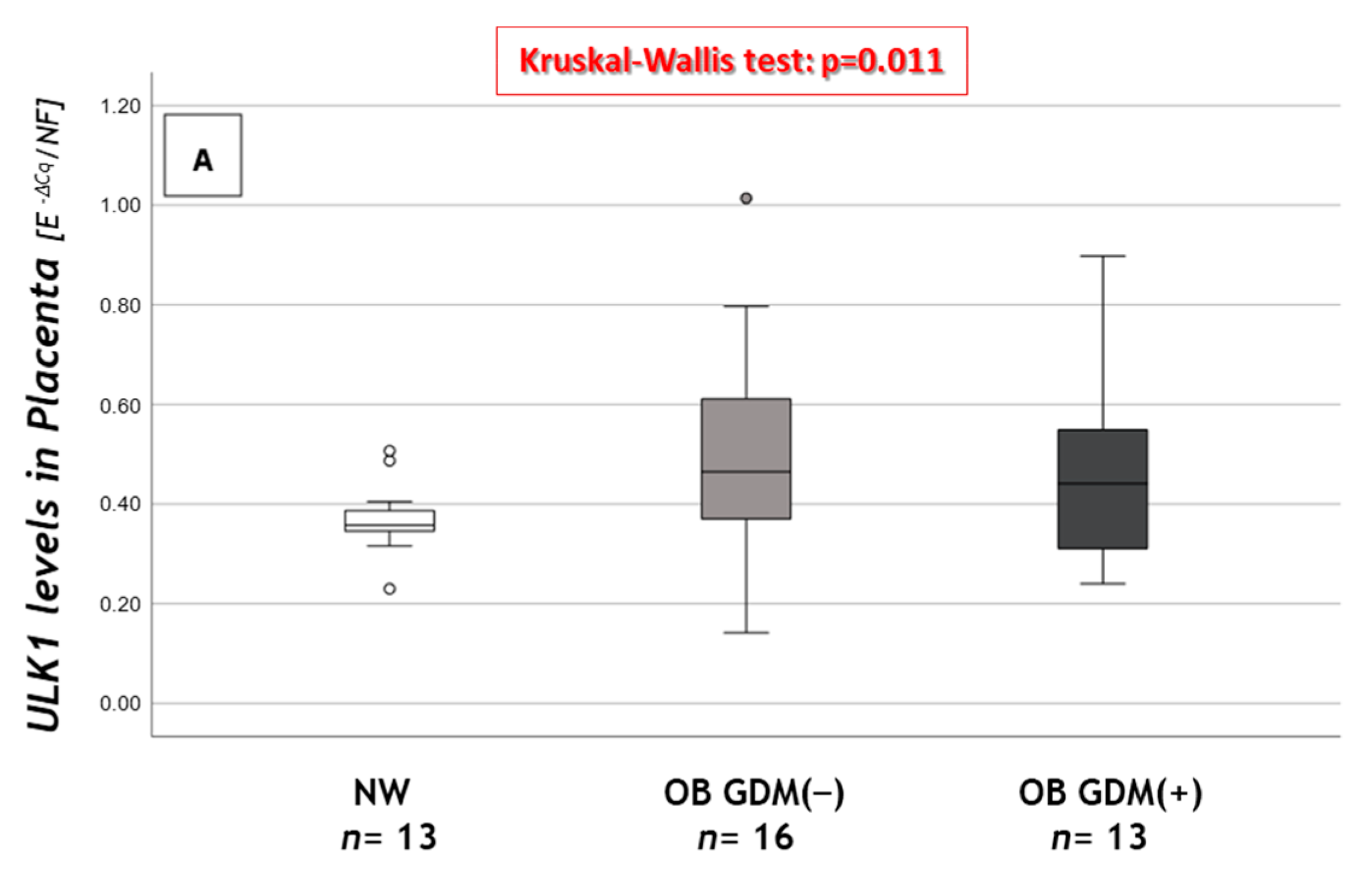
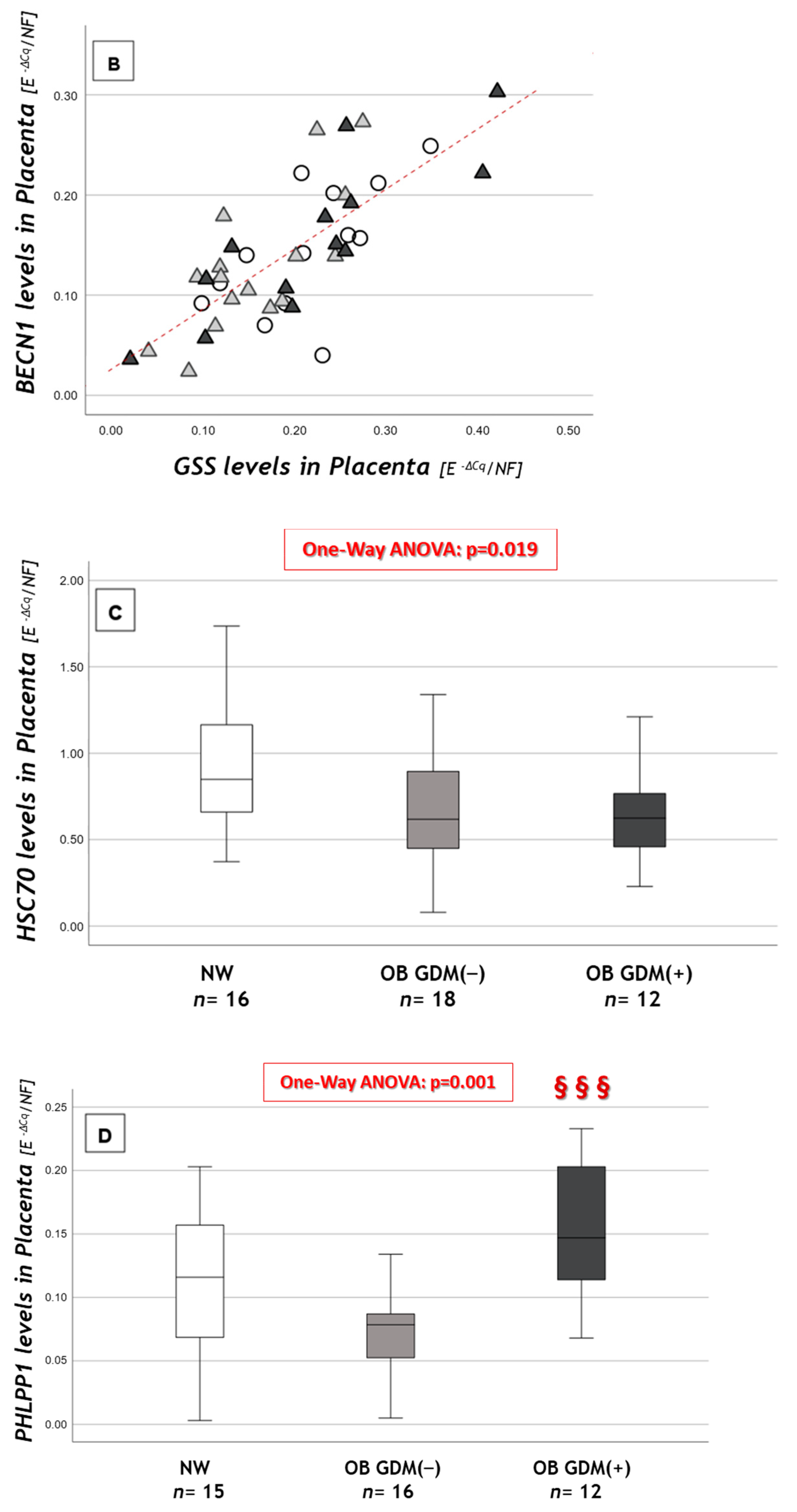
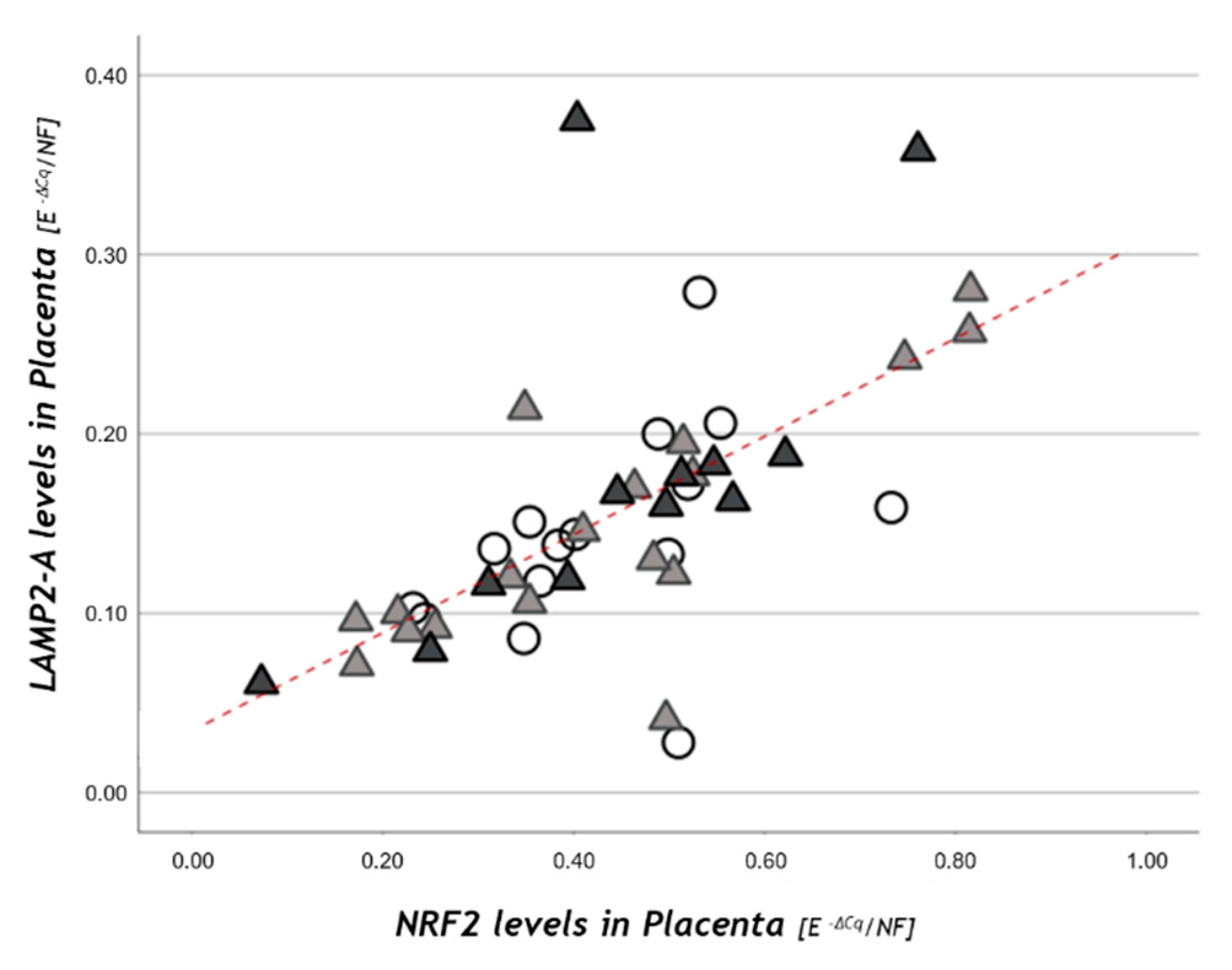
3.3. Autophagy-Related Gene Expression
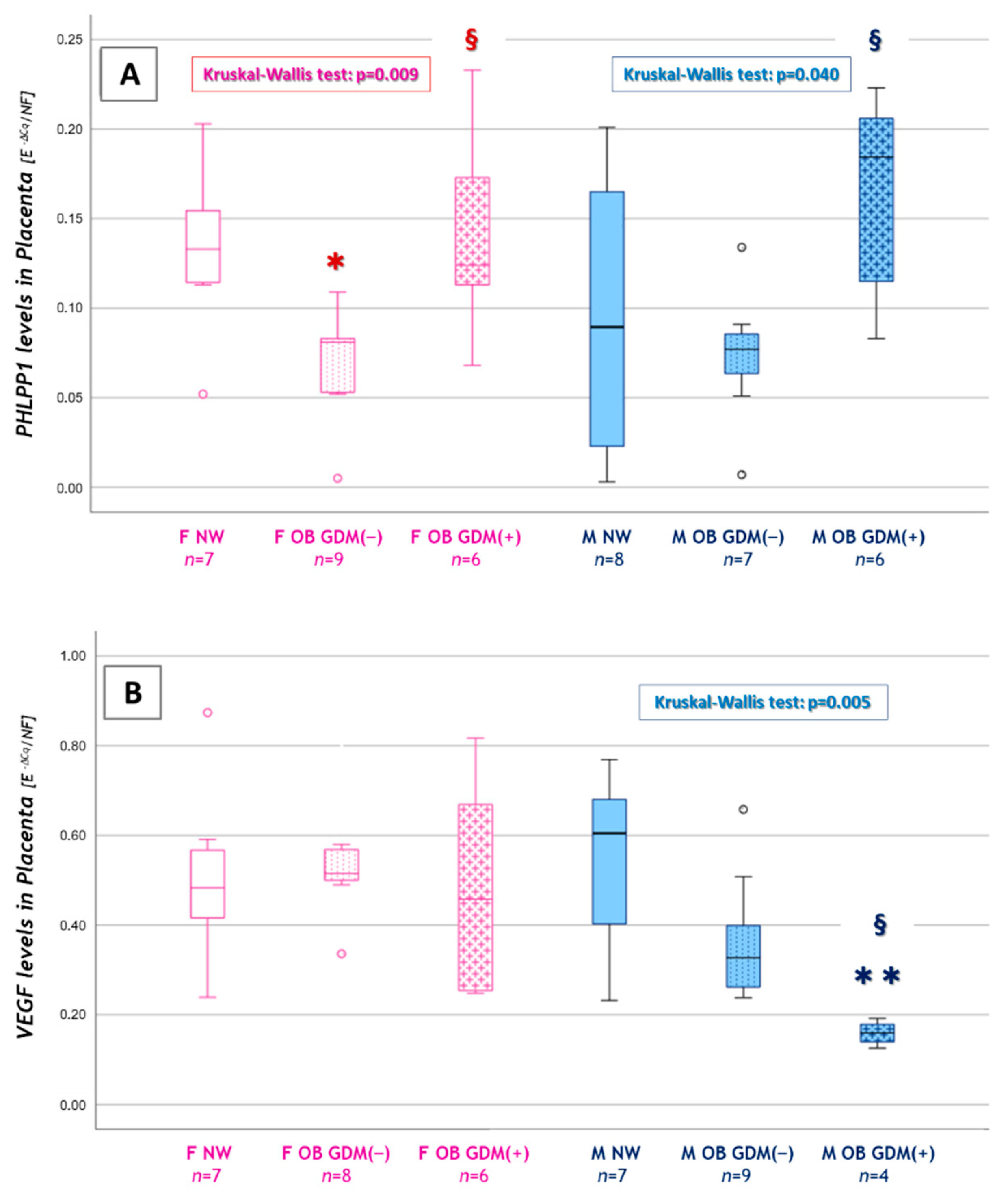
3.4. Sexually Dimorphic Placental Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaefer-Graf, U.; Napoli, A.; Nolan, C.J.; the Diabetic Pregnancy Study Group. Diabetes in pregnancy: A new decade of challenges ahead. Diabetologia 2018, 61, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef] [PubMed]
- Bordin, P.; Dotto, L.; Battistella, L.; Rosso, E.; Pecci, L.; Valent, F.; Collarile, P.; Vanin, M. Gestational diabetes mellitus yesterday, today and tomorrow: A 13 year italian cohort study. Diabetes Res. Clin. Pract. 2020, 167, 108360. [Google Scholar] [CrossRef]
- Martino, J.; Sebert, S.; Segura, M.T.; García-Valdés, L.; Florido, J.; Padilla, M.C.; Marcos, A.; Rueda, R.; McArdle, H.J.; Budge, H.; et al. Maternal Body Weight and Gestational Diabetes Differentially Influence Placental and Pregnancy Outcomes. J. Clin. Endocrinol. Metab. 2016, 101, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.P.; Thornburg, K.L.; Osmond, C.; Kajantie, E.; Eriksson, J.G. Beyond birthweight: The maternal and placental origins of chronic disease. J. Dev. Orig. Health Dis. 2010, 1, 360–364. [Google Scholar] [CrossRef]
- Saben, J.; Lindsey, F.; Zhong, Y.; Thakali, K.; Badger, T.M.; Andres, A.; Gomez-Acevedo, H.; Shankar, K. Maternal obesity is associated with a lipotoxic placental environment. Placenta 2014, 35, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandò, C.; Anelli, G.M.; Novielli, C.; Panina-Bordignon, P.; Massari, M.; Mazzocco, M.I.; Cetin, I. Impact of Obesity and Hyperglycemia on Placental Mitochondria. Oxid. Med. Cell. Longev. 2018, 14, 2378189. [Google Scholar] [CrossRef]
- Li, M.; Wu, X.; An, P.; Dang, H.; Liu, Y.; Liu, R. Effects of resveratrol on autophagy and the expression of inflammasomes in a placental trophoblast oxidative stress model. Life Sci. 2020, 256, 117890. [Google Scholar] [CrossRef]
- De Andrade Ramos, B.R.; Witkin, S.S. The influence of oxidative stress and autophagy cross regulation on pregnancy outcome. Cell Stress Chaperones 2016, 5, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkoong, S.; Cho, C.S.; Semple, I.; Lee, J.H. Autophagy Dysregulation and Obesity-Associated Pathologies. Mol. Cells 2018, 41, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.A.; Salmani, J.M.M.; Jiang, Z.; Feng, L.; Song, J.; Jia, X.; Chen, B. Autophagy: An overview and its roles in cancer and obesity. Clin. Chim. Acta 2017, 468, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Arias, E.; Koga, H.; Diaz, A.; Mocholi, E.; Patel, B.; Cuervo, A.M. Lysosomal mTORC2/PHLPP1/Akt Regulate Chaperone-Mediated Autophagy. Mol. Cell 2015, 59, 270–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiffin, R.; Christian, C.; Knecht, E.; Cuervo, A.M. Activation of chaperone-mediated autophagy during oxidative stress. Mol. Biol. Cell 2004, 15, 4829–4840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Suh, Y.; Cuervo, A.M. Regulated degradation of Chk1 by chaperone-mediated autophagy in response to DNA damage. Nat. Commun. 2015, 6, 6823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuervo, A.M.; Knecht, E.; Terlecky, S.R.; Dice, J.F. Activation of a selective pathway of lysosomal proteolysis in rat liver by prolonged starvation. Am. J. Physiol. 1995, 269, C1200–C1208. [Google Scholar] [CrossRef] [PubMed]
- Eisermann, D.J.; Wenzel, U.; Fitzenberger, E. Inhibition of chaperone-mediated autophagy prevents glucotoxicity in the Caenorhabditis elegans mev-1 mutant by activation of the proteasome. Biochem. Biophys. Res. Commun. 2017, 484, 171–175. [Google Scholar] [CrossRef]
- Zhao, X.; Jiang, Y.; Jiang, T.; Han, X.; Wang, Y.; Chen, L.; Feng, X. Physiological and pathological regulation of autophagy in pregnancy. Arch. Gynecol. Obstet. 2020, 302, 293–303. [Google Scholar] [CrossRef]
- Hubbi, M.E.; Hu, H.; Kshitiz, A.I.; Levchenko, A.; Semenza, G.L. Chaperone-mediated autophagy targets hypoxia-inducible factor-1α (HIF-1α) for lysosomal degradation. J. Biol. Chem. 2013, 288, 10703–10714. [Google Scholar] [CrossRef] [Green Version]
- Hung, T.H.; Hsieh, T.T.; Chen, S.F.; Li, M.J.; Yeh, Y.L. Autophagy in the human placenta throughout gestation. PLoS ONE 2013, 8, e83475. [Google Scholar] [CrossRef]
- Hung, T.H.; Huang, S.Y.; Chen, S.F.; Wu, C.P.; Hsieh, T.T. Decreased placental apoptosis and autophagy in pregnancies complicated by gestational diabetes with large-for-gestational age fetuses. Placenta 2020, 90, 27–36. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Gao, X.; Weintraub, S.; Myatt, L.; Maloyan, A. Sexual dimorphism in activation of placental autophagy in obese women with evidence for fetal programming from a placenta-specific mouse model. Autophagy 2016, 12, 752–769. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, K.M.; Yaktine, A.L.; Institute of Medicine (US); National Research Council (US); Committee to Reexamine IOM Pregnancy Weight Guidelines (Eds.) Weight Gain during Pregnancy: Reexamining the Guidelines; National Academies Press: Washington, DC, USA, 2009; p. 20669500. [CrossRef]
- Hod, M.; Kapur, A.; Sacks, D.A.; Hadar, E.; Agarwal, M.; Di Renzo, G.C.; Cabero Roura, L.; McIntyre, H.D.; Morris, J.L.; Divakar, H. The International Federation of Gynecology and Obstetrics (FIGO) Initiative on gestational diabetes mellitus: A pragmatic guide for diagnosis, management, and care. Int. J. Gynaecol. Obstet. 2015, 13, S173–S211. [Google Scholar] [CrossRef] [Green Version]
- International Association of Diabetes and Pregnancy Study Groups Consensus Panel; Metzger, B.E.; Gabbe, S.G.; Persson, B.; Buchanan, T.A.; Catalano, P.A.; Damm, P.; Dyer, A.R.; Leiva, A.D.; Hod, M.; et al. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 3, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.; Taricco, E.; Cardellicchio, M.; Mandò, C.; Massari, M.; Savasi, V.; Cetin, I. The role of obesity and gestational diabetes on placental size and fetal oxygenation. Placenta 2021, 103, 59–63. [Google Scholar] [CrossRef]
- Anelli, G.M.; Mandò, C.; Letizia, T.; Mazzocco, M.I.; Novielli, C.; Lisso, F.; Personeni, C.; Vago, T.; Cetin, I. Placental ESRRG-CYP19A1 Expressions and Circulating 17-Beta Estradiol in IUGR Pregnancies. Front. Pediatr. 2019, 7, 154. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetin, I.; Taricco, E.; Mandò, C.; Radaelli, T.; Boito, S.; Nuzzo, A.M.; Giussani, D.A. Fetal Oxygen and Glucose Consumption in Human Pregnancy Complicated by Fetal Growth Restriction. Hypertension 2020, 3, 748–754. [Google Scholar] [CrossRef]
- Zambon, M.; Mandò, C.; Lissoni, A.; Anelli, G.M.; Novielli, C.; Cardellicchio, M.; Leone, R.; Monari, M.N.; Massari, M.; Cetin, I.; et al. Inflammatory and Oxidative Responses in Pregnancies with Obesity and Periodontal Disease. Reprod. Sci. 2018, 10, 1474–1484. [Google Scholar] [CrossRef]
- Pantham, P.; Aye, I.L.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 7, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Mandò, C.; De Palma, C.; Stampalija, T.; Anelli, G.M.; Figus, M.; Novielli, C.; Parisi, F.; Clementi, E.; Ferrazzi, E.; Cetin, I. Placental mitochondrial content and function in intrauterine growth restriction and preeclampsia. Am. J. Physiol. Endocrinol. Metab. 2014, 4, E404–E413. [Google Scholar] [CrossRef]
- Kenney, M.C.; Chwa, M.; Atilano, S.R.; Falatoonzadeh, P.; Ramirez, C.; Malik, D.; Tarek, M.; Del Carpio, J.C.; Nesburn, A.B.; Boyer, D.S.; et al. Molecular and bioenergetic differences between cells with African versus European inherited mitochondrial DNA haplogroups: Implications for population susceptibility to diseases. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 2, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Durán, A.; Pacheu-Grau, D.; López-Gallardo, E.; Díez-Sánchez, C.; Montoya, J.; López-Pérez, M.J.; Ruiz-Pesini, E. Unmasking the causes of multifactorial disorders: OXPHOS differences between mitochondrial haplogroups. Hum. Mol. Genet. 2010, 17, 3343–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalano, P.; Demouzon, S.H. Maternal obesity and metabolic risk to the offspring: Why lifestyle interventions may have not achieved the desired outcomes. Int. J. Obes. 2015, 4, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.M.; Horgan, G.W.; Bhattacharya, S. Placental weight and efficiency in relation to maternal body mass index and the risk of pregnancy complications in women delivering singleton babies. Placenta 2012, 8, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Fattuoni, C.; Mandò, C.; Palmas, F.; Anelli, G.M.; Novielli, C.; Laudicina, E.P.; Savasi, V.M.; Barberini, L.; Dessì, A.; Pintus, R.; et al. Preliminary metabolomics analysis of placenta in maternal obesity. Placenta 2018, 61, 89–95. [Google Scholar] [CrossRef]
- Scifres, C.M.; Parks, W.T.; Feghali, M.; Caritis, S.N.; Catov, J.M. Placental maternal vascular malperfusion and adverse pregnancy outcomes in gestational diabetes mellitus. Placenta 2017, 49, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Hastie, R.; Lappas, M. The effect of pre-existing maternal obesity and diabetes on placental mitochondrial content and electron transport chain activity. Placenta 2014, 9, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell. Biol. 2010, 10, 1634–1650. [Google Scholar] [CrossRef]
- Pietrocola, F.; Bravo-San Pedro, J.M. Targeting Autophagy to Counteract Obesity-Associated Oxidative Stress. Antioxidants 2021, 1, 102. [Google Scholar] [CrossRef]
- Oh, S.Y.; Roh, C.R. Autophagy in the placenta. Obstet. Gynecol. Sci. 2017, 3, 241–259. [Google Scholar] [CrossRef]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaro, I.E.; Albornoz, A.; Molina, A.; Moreno, J.; Cordero, K.; Criollo, A.; Budini, M. Chaperone Mediated Autophagy in the Crosstalk of Neurodegenerative Diseases and Metabolic Disorders. Front. Endocrinol. 2019, 9, 778. [Google Scholar] [CrossRef] [PubMed]
- Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays Biochem. 2017, 6, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Menon, M.B.; Dhamija, S. Beclin 1 Phosphorylation—At the Center of Autophagy Regulation. Front. Cell Dev. Biol. 2018, 6, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huanhuan, L.V.; Zhen, C.; Liu, J.; Yang, P.; Hu, L.; Shang, P. Unraveling the Potential Role of Glutathione in Multiple Forms of Cell Death in Cancer Therapy. Oxid. Med. Cell. Longev. 2019, 3150145. [Google Scholar] [CrossRef] [Green Version]
- Lo Dico, A.; Martelli, C.; Diceglie, C.; Ottobrini, L. The Multifaceted Role of CMA in Glioma: Enemy or Ally? Review. Int. J. Mol. Sci. 2021, 22, 2217. [Google Scholar] [CrossRef]
- Mathur, A.; Pandey, V.K.; Kakkar, P. PHLPP: A putative cellular target during insulin resistance and type 2 diabetes. J. Endocrinol. 2017, 233, R185–R198. [Google Scholar] [CrossRef] [Green Version]
- Andreozzi, F.; Procopio, C.; Greco, A.; Mannino, G.C.; Miele, C.; Raciti, G.A.; Iadicicco, C.; Beguinot, F.; Pontiroli, A.E.; Hribal, M.L.; et al. Increased levels of the Akt-specific phosphatase PH domain leucine-rich repeat protein phosphatase (PHLPP)-1 in obese participants are associated with insulin resistance. Diabetologia 2011, 54, 1879–1887. [Google Scholar] [CrossRef] [Green Version]
- Leng, S.; Zhang, W.; Zheng, Y.; Liberman, Z.; Rhodes, C.J.; Eldar-Finkelman, H.; Sun, X.J. Glycogen synthase kinase 3 beta mediates high glucose-induced ubiquitination and proteasome degradation of insulin receptor substrate 1. J. Endocrinol. 2010, 2, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Pajares, M.; Rojo, A.I.; Arias, E.; Díaz-Carretero, A.; Cuervo, A.M.; Cuadrado, A. Transcription factor NFE2L2/NRF2 modulates chaperone-mediated autophagy through the regulation of LAMP2A. Autophagy 2018, 8, 1310–1322. [Google Scholar] [CrossRef]
- Osei-Kumah, A.; Smith, R.; Jurisica, I.; Caniggia, I.; Clifton, V.L. Sex-specific differences in placental global gene expression in pregnancies complicated by asthma. Placenta 2011, 8, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Clifton, V.L. Sexually dimorphic effects of maternal asthma during pregnancy on placental glucocorticoid metabolism and fetal growth. Cell Tissue Res. 2005, 1, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Anelli, G.M.; Cardellicchio, M.; Novielli, C.; Antonazzo, P.; Mazzocco, M.I.; Cetin, I.; Mandò, C. Mitochondrial content and hepcidin are increased in obese pregnant mothers. J. Matern. Fetal Neonatal Med. 2018, 18, 2388–2395. [Google Scholar] [CrossRef] [PubMed]
- Mandò, C.; Calabrese, S.; Mazzocco, M.I.; Novielli, C.; Anelli, G.M.; Antonazzo, P.; Cetin, I. Sex specific adaptations in placental biometry of overweight and obese women. Placenta 2016, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
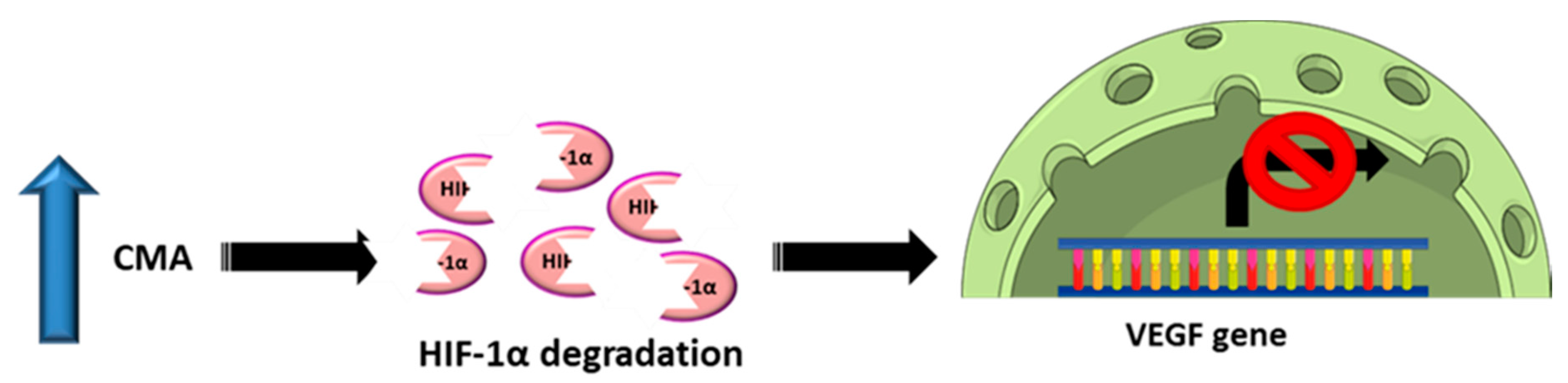{kind=link}
| Gene | Forward Sequence (5′–3′) | Reverse Sequence (3′–5′) | Fw Position * | Rev Position * | Amplicon Length |
|---|---|---|---|---|---|
| YWHAZ | ACTTTTGGTACATTGTGGCTTCAA | CCGCCAGGACAAACCAGTAT | 1110–1133 | 1203–1184 | 51 bp |
| β–ACTIN | ATCAAGATCATTGCTCCTCCTGA | CTGCTTGCTGATCCACATCTG | 1063–1085 | 1164–1144 | 59 bp |
| CAT | TAAGACTGACCAGGGCA | CAAACCTTGGTGAGATCGAA | 788–804 | 988–969 | 165 bp |
| GSS | ATGCTGTGCAGATGGACTTCAACC | TGGATGTCAAACAGACGAGCGGTA | 285–308 | 408–385 | 77 bp |
| GSR | AACATCCCAACTGTGGTCTTCAGC | TTGGTAACTGCGTGATACATCGGG | 1239–1262 | 1378–1355 | 93 bp |
| GPX1 | CGCAACGATGTTGCCTGGAACTTT | AGGCTCGATGTCAATGGTCTGGAA | 563–586 | 664–641 | 55 bp |
| ULK1 | TCATCTTCAGCCACGCTG | CACGGTGCTGGAACATCT | 2724–2741 | 2789–2772 | 31 bp |
| BECN1 | GGCTGAGAGACTGGATCAGG | CTGCGTCTGGGCATAACG | 806–825 | 932–915 | 90 bp |
| HSC70 | ATTGATCTTGGCACCACCTA | ACATAGCTTGAAGTGGTTCG | 103–122 | 203–184 | 62 bp |
| LAMP–2A | TGCTGGCTACCATGGGGC TG | GCAGCTGCCTGTGGAGTGAGT | 825–844 | 922–902 | 58 bp |
| PHLPP1 | CCTACCTTCTCCAGTGCACT | CCAGCAGTTCCAAGTTTCCT | 3796–3815 | 3916–3897 | 82 bp |
| NRF2 | CAGCGACGGAAAGAGTATGA | AAGAAACCTGGGAGTAG | 271–291 | 471–454 | 163 bp |
| HIF–1α | TGATTGCATCTCCATCTCCTACC | GACTCAAAGCGACAGATAACACG | 2205–2227 | 2381–2359 | 132 bp |
| VEGF | CGAGGGCCTGGAGTGTGT | CGCATAATCTGCATGGTGATG | 335–352 | 391–371 | 19 bp |
| One-Way ANOVA or Kruskal–Wallis Test | NW n = 16 | OB GDM(−) n = 18 | OB GDM(+) n = 13 | |
|---|---|---|---|---|
| Maternal Data | ||||
| Age (years) A | p = 0.002 | 35.7 ± 3.5 | 30.2 ± 6.0 ** | 35.8 ± 4.3 §§ |
| Pregestational BMI (kg/m2) B | p = 0.000 | 21.18 ± 2.11 | 34.80 ± 4.06 *** | 35.05 ± 3.41 *** |
| OGTT I Value (mg/dL) A | ns | 84.00 ± 5.12 | 79.92 ± 6.52 | 89.73 ± 15.08 |
| OGTT II Value (mg/dL) A | p = 0.000 | 117.73 ± 19.14 | 113.10 ± 22.57 | 164.67 ± 35.99 *** §§§ |
| OGTT III Value (mg/dL) B | p = 0.001 | 105.36 ± 36.28 | 92.20 ± 14.19 | 162.89 ± 32.28 ** §§§ |
| Gestational Weight Gain (kg) A | ns | 10.93 ± 2.86 | 8.44 ± 6.06 | 6.62 ± 5.91 |
| Hemoglobin (mg/dL), at 34–36 weeks B | ns | 12.13 ± 1.16 | 11.35 ± 0.76 | 11.37 ± 0.97 |
| Fetal (F) and Placental (P) Data at Delivery | ||||
| Gestational Age (weeks) B | ns | 39.26 ± 0.50 | 39.16 ± 0.30 | 39.09 ± 0.16 |
| Fetal Weight (g) A | ns | 3337.94 ± 247.71 | 3339.44 ± 417.17 | 3330.00 ± 336.54 |
| Placental Weight (g) A | ns | 434.25 ± 80.43 | 482.22 ± 81.50 | 497.46 ± 102.55 |
| Placental Surface (cm2) A | ns | 245.59 ± 45.25 | 232.34 ± 85.03 | 245.44 ± 46.41 |
| Placental Thickness (cm) A | ns | 1.81 ± 0.42 | 2.21 ± 0.58 | 2.04 ± 0.55 |
| Placental Efficiency (F/P weight) B | ns | 7.97 ± 1.75 | 7.04 ± 1.04 | 6.98 ± 1.85 |
| MALE FETUSES (n = 24) | FEMALE FETUSES (n = 23) | |||||||
|---|---|---|---|---|---|---|---|---|
| One-Way ANOVA or Kruskal–Wallis Test | NW n = 9 | OB GDM(−) n = 9 | OB GDM(+) n = 6 | One-Way ANOVA or Kruskal–Wallis Test | NW n = 7 | OB GDM(−) n = 9 | OB GDM(+) n = 7 | |
| Fetal (F) Weight (g) A | ns | 3371.67 ± 287.26 | 3368.89 ± 450.68 | 3374.17 ± 369.49 | ns | 3294.57 ± 198.68 | 3310.00 ± 405.90 | 3292.14 ± 283.58 |
| Placental (P) Weight (g) A | ns | 451.44 ± 79.26 | 495.56 ± 95.93 | 441.17 ± 105.75 | p = 0.011 | 412.14 ± 82.35 | 468.89 ± 67.17 | 545.71 ± 76.35 ** |
| Placental Efficiency (F/P weight) B | ns | 7.76 ± 2.02 | 6.92 ± 0.89 | 8.05 ± 2.27 | p = 0.012 | 8.24 ± 1.46 | 7.16 ± 1.22 | 6.07 ± 0.68 ** |
| Antioxidant Defenses | NW n = 16 | OB GDM(−) n = 18 | OB GDM(+) n = 13 |
|---|---|---|---|
| CATA | 0.146 ± 0.054 | 0.087 ± 0.048 | 0.115 ± 0.104 |
| GSSA | 0.215 ± 0.071 | 0.159 ± 0.068 | 0.218 ± 0.114 |
| GSRB | 0.265 ± 0.131 | 0.213 ± 0.116 | 0.294 ± 0.180 |
| GPX1A | 0.207 ± 0.067 | 0.152 ± 0.062 | 0.187 ± 0.061 |
| Macroautophagy and CMA | One-Way Anova | NW n = 16 | OB GDM(−) n = 18 | OB GDM(+) n = 13 |
|---|---|---|---|---|
| BECN1A | ns | 0.145 ± 0.063 | 0.142 ± 0.088 | 0.155 ± 0.078 |
| LAMP-2AA | ns | 0.143 ± 0.058 | 0.147 ± 0.068 | 0.179 ± 0.097 |
| Autophagy-Related Genes | NW n = 16 | OB GDM(−) n = 18 | OB GDM(+) n = 13 |
|---|---|---|---|
| NRF2A | 0.433 ± 0.128 | 0.437 ± 0.203 | 0.449 ± 0.181 |
| HIF-1αB | 0.235 ± 0.080 | 0.275 ± 0.168 | 0.218 ± 0.087 |
| VEGFA | 0.524± 0.198 | 0.447 ± 0.158 | 0.354 ± 0.243 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diceglie, C.; Anelli, G.M.; Martelli, C.; Serati, A.; Lo Dico, A.; Lisso, F.; Parisi, F.; Novielli, C.; Paleari, R.; Cetin, I.; et al. Placental Antioxidant Defenses and Autophagy-Related Genes in Maternal Obesity and Gestational Diabetes Mellitus. Nutrients 2021, 13, 1303. https://doi.org/10.3390/nu13041303
Diceglie C, Anelli GM, Martelli C, Serati A, Lo Dico A, Lisso F, Parisi F, Novielli C, Paleari R, Cetin I, et al. Placental Antioxidant Defenses and Autophagy-Related Genes in Maternal Obesity and Gestational Diabetes Mellitus. Nutrients. 2021; 13(4):1303. https://doi.org/10.3390/nu13041303
Chicago/Turabian StyleDiceglie, Cecilia, Gaia Maria Anelli, Cristina Martelli, Anais Serati, Alessia Lo Dico, Fabrizia Lisso, Francesca Parisi, Chiara Novielli, Renata Paleari, Irene Cetin, and et al. 2021. "Placental Antioxidant Defenses and Autophagy-Related Genes in Maternal Obesity and Gestational Diabetes Mellitus" Nutrients 13, no. 4: 1303. https://doi.org/10.3390/nu13041303