
Maternal Quercetin Consumption during Pregnancy May Help Regulate Total Cholesterol/HDL-Cholesterol Ratio without Effect on Cholesterol Levels in Male Progeny Consuming High-Fat Diet


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal Experiments
2.2.1. Institutional Approval of the Study Protocols
2.2.2. Animals
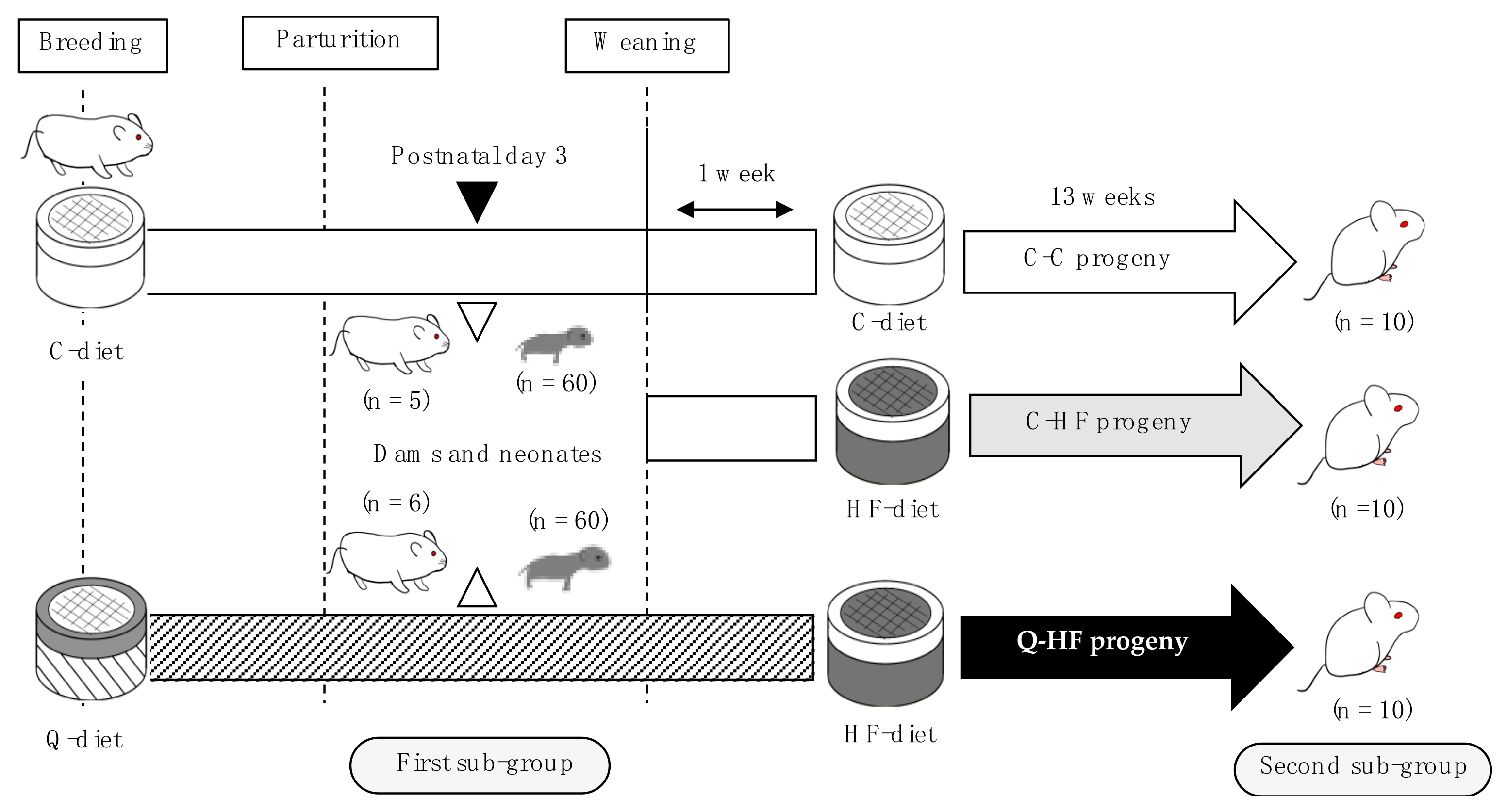
2.2.3. Breeding
2.2.4. Experimental Design
2.2.5. Sample Collections
2.3. Biochemical Parameters
2.3.1. Blood Biochemistry and Adipokines
2.3.2. Hepatic Lipid Analysis
2.4. Statistical Analysis
3. Results
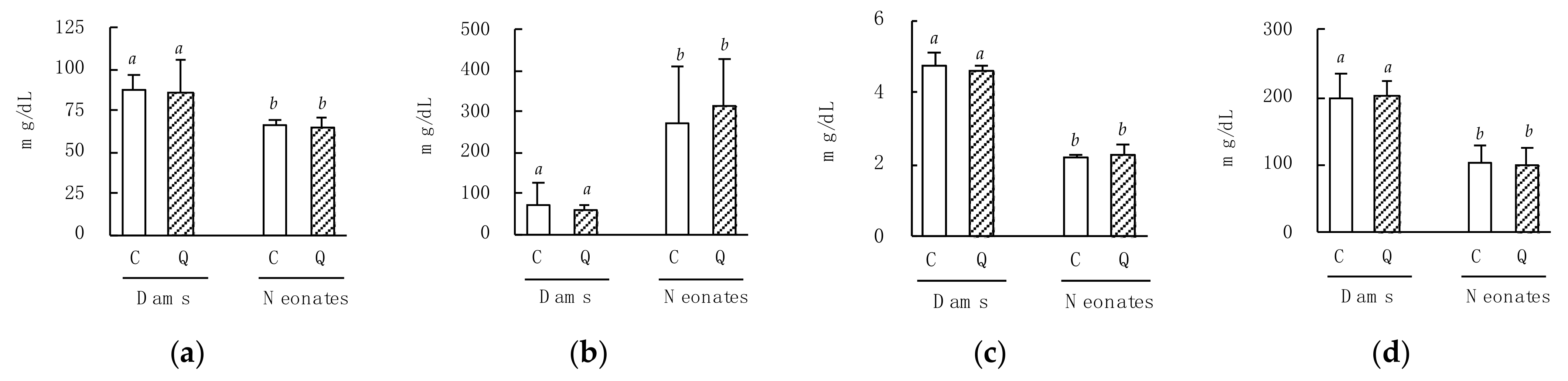
3.1. Effects of Maternal Quercetin Consumption on Neonatal Biological and Blood Parameters
3.2. Effects of Maternal Quercetin Consumption on Biological Parameters in Progeny Fed a HF Diet
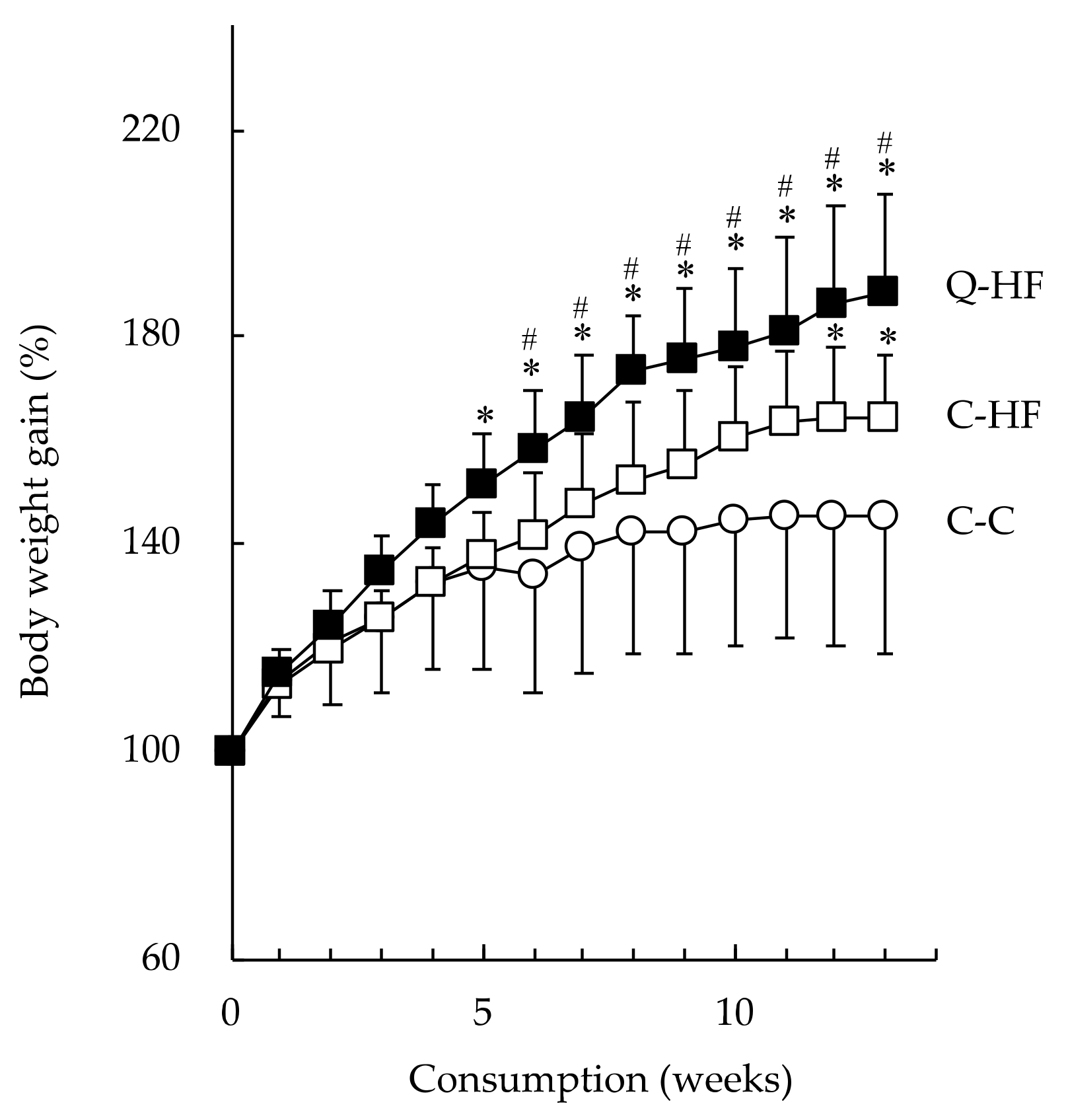
3.2.1. Effects on Food Intake, Body Weight, and Organ Weight
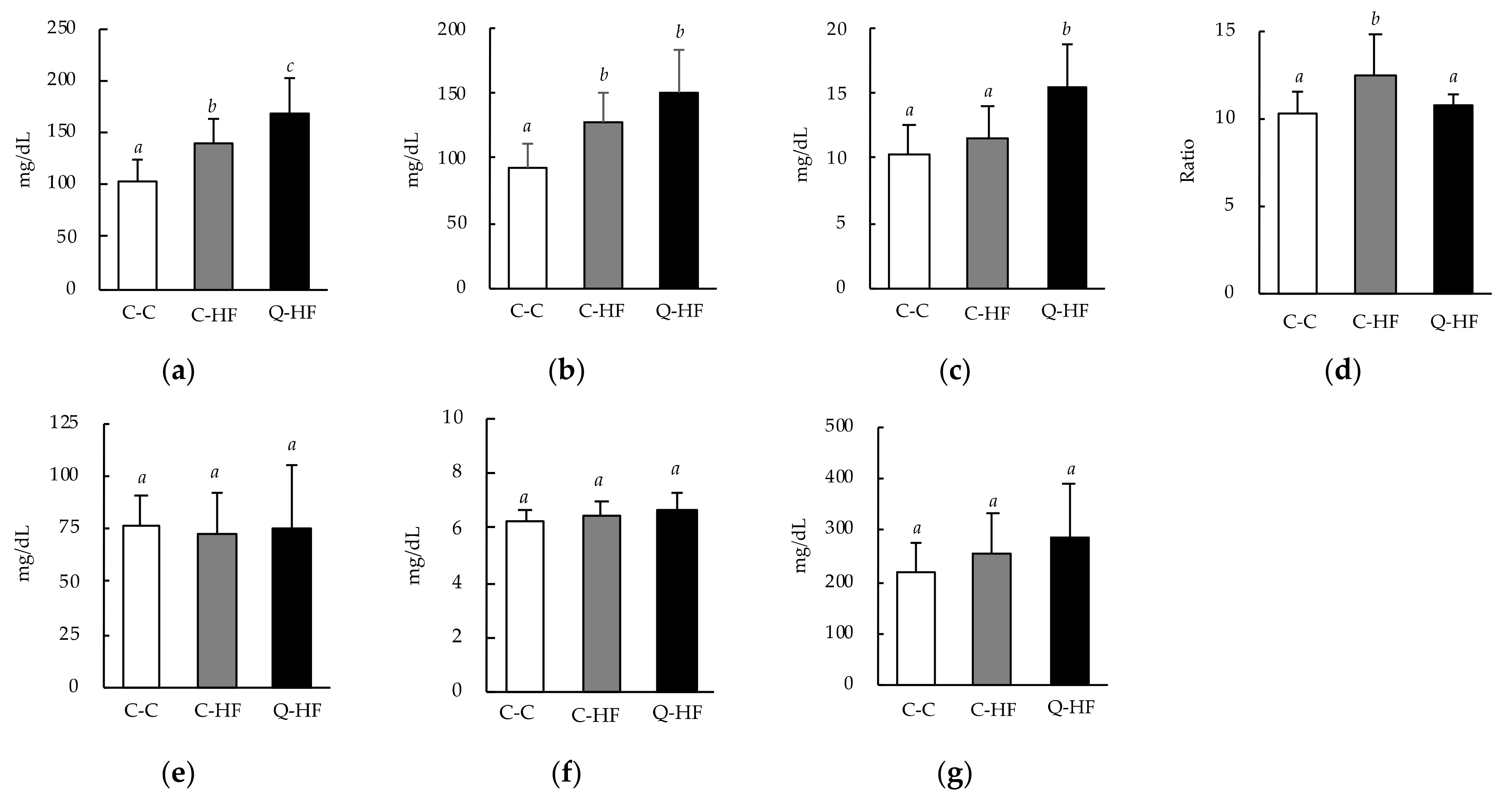
3.2.2. Effects on Blood Biochemical Parameters
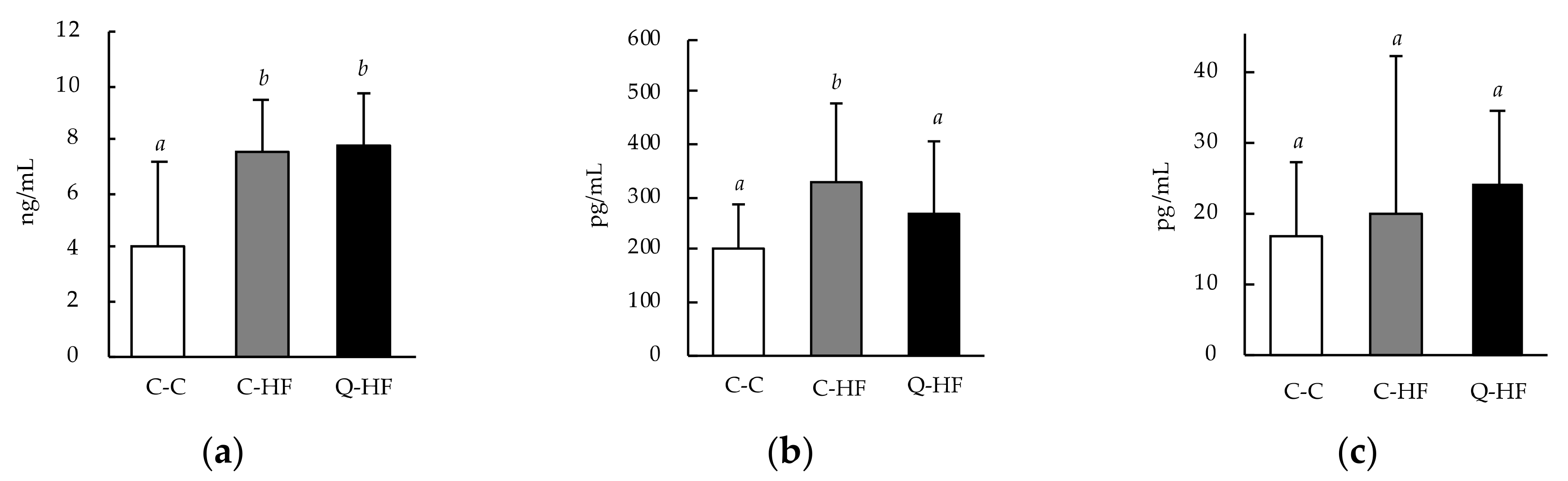
3.2.3. Effects on Blood Metabolic Hormones and Myokines
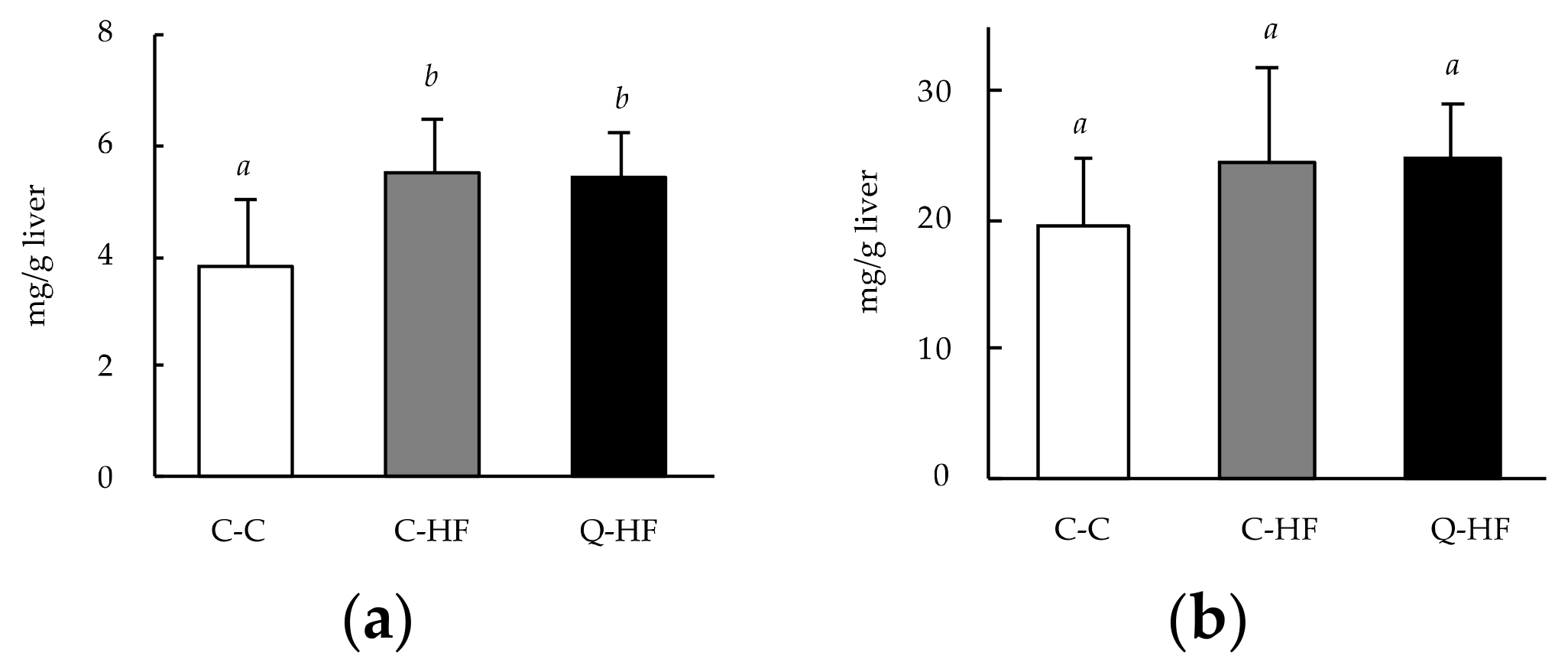
3.2.4. Effects on Hepatic Lipid Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caballero, B. Humans against obesity: Who Will Win? Adv. Nutr. 2019, 10, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Obesity: Preventing and Managing the Global Epidemic; World Health Organization: Geneva, Switzerland, 2000; Volume 894.
- Craig, P.; Halavatau, V.; Comino, E.; Caterson, I. Differences in body composition between Tongans and Australians: Time to rethink the healthy weight ranges? Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1806–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elagizi, A.; Kachur, S.; Lavie, C.J.; Carbone, S.; Pandey, A.; Ortega, F.B.; Milani, R.V. An overview and update on obesity and the obesity paradox in cardiovascular diseases. Prog. Cardiovasc. Dis. 2018, 61, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Vecchie, A.; Dallegri, F.; Carbone, F.; Bonaventura, A.; Liberale, L.; Portincasa, P.; Fruhbeck, G.; Montecucco, F. Obesity phenotypes and their paradoxical association with cardiovascular diseases. Eur. J. Intern. Med. 2018, 48, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. The importance of energy balance. Eur. Endocrinol. 2013, 9, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Carrera-Bastos, P.; Fontes-Villalba, M.; O’Keefe, J.H.; Lindeberg, S.; Cordain, L. The western diet and lifestyle and diseases of civilization. Res. Rep. Clin. Cardiol. 2011, 2, 15–35. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lee, J.; Kim, Y. Effect of deglycosylated rutin by acid hydrolysis on oobesity and hyperlipidemia in high-fat diet-induced obese mice. Nutrients 2020, 12, 1539. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, U.; Rubab, M.; Daliri, E.B.; Chelliah, R.; Javed, A.; Oh, D.H. Curcumin, quercetin, catechins and metabolic diseases: The role of gut microbiota. Nutrients 2021, 13, 206. [Google Scholar] [CrossRef]
- Jiang, H.; Horiuchi, Y.; Hironao, K.Y.; Kitakaze, T.; Yamashita, Y.; Ashida, H. Prevention effect of quercetin and its glycosides on obesity and hyperglycemia through activating AMPKalpha in high-fat diet-fed ICR mice. J. Clin. Biochem. Nutr. 2020, 67, 74–83. [Google Scholar] [CrossRef]
- Yokoyama, D.; Tanaka, W.; Hashizume, Y.; Tandia, M.; Sakono, M.; Shimoi, K.; Sakakibara, H. Daily consumption of monoglucosyl-rutin prevents high-fat diet-induced obesity by suppressing gastric inhibitory polypeptide secretion in mice. Funct. Foods Health Dis. 2018, 8, 353–371. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Muro, T.; Kobori, M.; Nishihira, J. Effect of daily ingestion of quercetin-rich onion powder for 12 weeks on visceral fat: A randomised, double-blind, placebo-controlled, parallel-group study. Nutrients 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, P.D.; Hanson, M.A.; Buklijas, T. A conceptual framework for the developmental origins of health and disease. J. Dev. Orig. Health Dis. 2010, 1, 6–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Mandy, M.; Nyirenda, M. Developmental Origins of Health and Disease: The relevance to developing nations. Int. Health 2018, 10, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.A.; Javurek, A.B.; Painter, M.S.; Murphy, C.R.; Conard, C.M.; Gant, K.L.; Howald, E.C.; Ellersieck, M.R.; Wiedmeyer, C.E.; Vieira-Potter, V.J.; et al. Effects of a maternal high-fat diet on offspring behavioral and metabolic parameters in a rodent model. J. Dev. Orig. Health Dis. 2017, 8, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Lee, C.T.; Chan, J.Y.H.; Tain, Y.L. The interplay between maternal and post-weaning high-fat diet and gut microbiota in the developmental programming of hypertension. Nutrients 2019, 11, 1982. [Google Scholar] [CrossRef] [Green Version]
- Akyol, A.; Cetin, A.K.; Gulec, A.; Dasgin, H.; Ayaz, A.; Onbasilar, I. Maternal low-quality protein diet exerts sex-specific effects on plasma amino acid profile and alters hepatic expression of methyltransferases in adult rat offspring. J. Dev. Orig. Health Dis. 2018, 9, 409–416. [Google Scholar] [CrossRef]
- Kawakubo-Yasukochi, T.; Morioka, M.; Ohe, K.; Yasukochi, A.; Ozaki, Y.; Hazekawa, M.; Nishinakagawa, T.; Ono, K.; Nakamura, S.; Nakashima, M. Maternal folic acid depletion during early pregnancy increases sensitivity to squamous tumor formation in the offspring in mice. J. Dev. Orig. Health Dis. 2019, 10, 683–691. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, W.C.; Wu, K.L.H.; Leu, S.; Chan, J.Y.H. Maternal high fructose intake increases the vulnerability to post-weaning high-fat diet-induced programmed hypertension in male offspring. Nutrients 2018, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhao, J.; Xu, H.; Lyv, Y.; Feng, X.; Fang, Y.; Xu, Y. Maternal quercetin administration during gestation and lactation decrease endoplasmic reticulum stress and related inflammation in the adult offspring of obese female rats. Eur. J. Nutr. 2014, 53, 1669–1683. [Google Scholar] [CrossRef]
- Braden, G.C.; Rasmussen, S.; Monette, S.; Tolwani, R.J. Effects of breeding configuration on maternal and weanling behavior in laboratory mice. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 369–376. [Google Scholar]
- Zhang, M.; Liu, W.; Zhou, Y.; Li, Y.; Qin, Y.; Xu, Y. Neurodevelopmental toxicity induced by maternal PM2.5 exposure and protective effects of quercetin and Vitamin C. Chemosphere 2018, 213, 182–196. [Google Scholar] [CrossRef]
- Yamaki, K.; Takahashi, Y. Additive beneficial effect of epigallocatechin gallate and quercetin on the arteriosclerosis index in mice. Food Sci. Technol. Res. 2017, 23, 355–361. [Google Scholar] [CrossRef]
- McMullin, T.S.; Lowe, E.R.; Bartels, M.J.; Marty, M.S. Dynamic changes in lipids and proteins of maternal, fetal, and pup blood and milk during perinatal development in CD and Wistar rats. Toxicol. Sci. 2008, 105, 260–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetulani, J. Early maternal separation: A rodent model of depression and a prevailing human condition. Pharmacol. Rep. 2013, 65, 1451–1461. [Google Scholar] [CrossRef]
- Md, A.H.; Deb, K.P.; Hossain, M.A.; Mannan, K.A.; Mostafa, G.; Hossain, M.M. Correlation between serum cholesterol and serum albumin level in childhood nephrotic syndrome. Uroll Nephrol. Open Access J. 2016, 3, 115–118. [Google Scholar]
- Hong, J.; Stubbins, R.E.; Smith, R.R.; Harvey, A.E.; Nunez, N.P. Differential susceptibility to obesity between male, female and ovariectomized female mice. Nutr. J. 2009, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Lim, S.M.; Jung, J.I.; Kim, S.M.; Lee, J.K.; Kim, Y.H.; Cha, K.M.; Oh, T.K.; Moon, J.M.; Kim, T.Y.; et al. Gynostemma pentaphyllum extract ameliorates high-fat diet-Induced obesity in C57BL/6N mice by upregulating SIRT1. Nutrients 2019, 11, 2475. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Kim, J.; Park, H.J.; Kim, H. Anti-obesity effects of a prunus persica and nelumbo nucifera mixture in mice fed a high-fat diet. Nutrients 2020, 12, 3392. [Google Scholar] [CrossRef] [PubMed]
- Keller, U. Nutritional laboratory markers in malnutrition. J. Clin. Med. 2019, 8, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laclaustra, M.; Lopez-Garcia, E.; Civeira, F.; Garcia-Esquinas, E.; Graciani, A.; Guallar-Castillon, P.; Banegas, J.R.; Rodriguez-Artalejo, F. LDL cholesterol rises with BMI only in lean individuals: Cross-sectional U.S. and Spanish representative data. Diabetes Care 2018, 41, 2195–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Wu, J.; Tang, J.; Wang, J.J.; Lu, C.H.; Wang, P.X. Beneficial effect of higher dietary fiber intake on plasma HDL-C and TC/HDL-C ratio among Chinese rural-to-urban migrant workers. Int. J. Environ. Res. Public Health 2015, 12, 4726–4738. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wu, H.; Liu, Y.; Yang, L. High fat diet induced obesity model using four strainsof mice: Kunming, C57BL/6, BALB/c and ICR. Exp. Anim. 2020, 69, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.R.; Li, J.Y.; Dong, X.W.; Tan, Z.J.; Wu, W.Z.; Xie, Q.M.; Yang, Y.M. Apple polyphenols decrease atherosclerosis and hepatic steatosis in ApoE-/- mice through the ROS/MAPK/NF-kappaB pathway. Nutrients 2015, 7, 7085–7105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Urbina, A.; Rull, A.; Aldana-Ramos, J.; Santos, D.; Puig, N.; Farre-Cabrerizo, N.; Benitez, S.; Perez, A.; de Gonzalo-Calvo, D.; Escola-Gil, J.C.; et al. Subcutaneous administration of apolipoprotein J-derived mimetic peptide d-[113–122]apoJ improves LDL and HDL function and prevents atherosclerosis in LDLR-KO mice. Biomolecules 2020, 10, 829. [Google Scholar] [CrossRef]
- Yu, Y.; Luo, T.; Liu, S.; Song, G.; Han, J.; Wang, Y.; Yao, S.; Feng, L.; Qin, S. Chitosan oligosaccharides attenuate atherosclerosis and decrease non-HDL in ApoE-/- mice. J. Atheroscler. Thromb. 2015, 22, 926–941. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.G.; Myers, G.L.; Sakurabayashi, I.; Bachmann, L.M.; Caudill, S.P.; Dziekonski, A.; Edwards, S.; Kimberly, M.M.; Korzun, W.J.; Leary, E.T.; et al. Seven direct methods for measuring HDL and LDL cholesterol compared with ultracentrifugation reference measurement procedures. Clin. Chem. 2010, 56, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Champy, M.F.; Selloum, M.; Piard, L.; Zeitler, V.; Caradec, C.; Chambon, P.; Auwerx, J. Mouse functional genomics requires standardization of mouse handling and housing conditions. Mamm. Genome 2004, 15, 768–783. [Google Scholar] [CrossRef]
- Lin, X.; Chavez, M.R.; Bruch, R.C.; Kilroy, G.E.; Simmons, L.A.; Lin, L.; Braymer, H.D.; Bray, G.A.; York, D.A. The effects of a high fat diet on leptin mRNA, serum leptin and the response to leptin are not altered in a rat strain susceptible to high fat diet-induced obesity. J. Nutr. 1998, 128, 1606–1613. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, R.; Ogata, H.; Murase, M.; Harada, N.; Suzuki, K.; Joo, E.; Sankoda, A.; Iida, A.; Izumoto, T.; Tsunekawa, S.; et al. Glucose-dependent insulinotropic polypeptide is required for moderate high-fat diet- but not high-carbohydrate diet-induced weight gain. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E572–E583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabe, D.; Seino, Y. Two incretin hormones GLP-1 and GIP: Comparison of their actions in insulin secretion and beta cell preservation. Prog. Biophys. Mol. Biol. 2011, 107, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Nasteska, D.; Harada, N.; Suzuki, K.; Yamane, S.; Hamasaki, A.; Joo, E.; Iwasaki, K.; Shibue, K.; Harada, T.; Inagaki, N. Chronic reduction of GIP secretion alleviates obesity and insulin resistance under high-fat diet conditions. Diabetes 2014, 63, 2332–2343. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal models of metabolic syndrome: A review. Nutr. Metab. 2016, 13, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, H.; Honda, Y.; Nakagawa, S.; Ashida, H.; Kanazawa, K. Simultaneous determination of all polyphenols in vegetables, fruits, and teas. J. Agric. Food Chem. 2003, 51, 571–581. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal-Fat Diet | High-Fat Diet (HF) | ||
|---|---|---|---|
| – (C) | Quercetin (Q) | ||
| in 100 g diets | |||
| β-Cornstarch (g) | 39.75 | 38.75 | 16.75 |
| α-Cornstarch (g) | 13.2 | 13.2 | 13.2 |
| Casein (g) | 20.0 | 20.0 | 20.0 |
| Soybean oil (g) | 7.0 | 7.0 | 7.0 |
| Lard (g) | – | – | 23.0 |
| Sucrose (g) | 10.0 | 10.0 | 10.0 |
| Cellulose (g) | 5.0 | 5.0 | 5.0 |
| Vitamin mixture (g) | 1.0 | 1.0 | 1.0 |
| Mineral mixture (g) | 3.5 | 3.5 | 3.5 |
| L-Cysteine (g) | 0.3 | 0.3 | 0.3 |
| Choline bitartrate (g) | 0.25 | 0.25 | 0.25 |
| t-Butylhydroquinone (mg) | 1.4 | 1.4 | 1.4 |
| Quercetin (g) | – | 1.0 | – |
| Energy (kcal/g) | 3.948 | 3.908 | 5.098 |
| Dams | Neonates | |||
|---|---|---|---|---|
| C-Diet | Q-Diet | C-Diet | Q-Diet | |
| Body weight (g) | 42.1 ± 4.1 | 41.4 ± 4.1 | 3.1 ± 1.0 | 3.3 ± 0.3 |
| Number of animals per dam | 5 | 6 | 12 ± 3 | 10 ± 3 |
| Food intake (g/mice/day) | 7.2 ± 1.1 | 7.7 ± 2.4 | – | – |
| Quercetin intake (mg/mice/day) | – | 77 ± 24 | – | – |
| C–C | C–HF | Q–HF | ANOVA p Values | ||
|---|---|---|---|---|---|
| Maternal Diet | Progeny Diet | ||||
| Body weight | |||||
| Initial (g) | 37.8 ± 2.9 a | 38.7 ± 2.1 a | 37.7 ± 2.9 a | 0.582 | 0.747 |
| Final (g) | 54.6 ± 6.2 a | 63.4 ± 4.5 b | 71.3 ± 12.2 c | 0.001 | <0.001 |
| Food consumption | |||||
| (g/mouse/day) | 5.1 | 4.0 | 4.1 | – | – |
| (kcal/mouse/day) | 20.1 | 20.4 | 20.9 | – | – |
| Absolute organ weight (g) | |||||
| Liver | 2.18 ± 0.29 a | 2.45 ± 0.36 a | 3.56 ± 1.53 b | 0.003 | 0.041 |
| Kidney | 0.85 ± 0.06 a | 0.79 ± 0.08 a | 0.77 ± 0.15 a | 0.186 | 0.111 |
| Spleen | 0.21 ± 0.05 a | 0.22 ± 0.07 a | 0.20 ± 0.05 a | 0.514 | 0.827 |
| Heart | 0.27 ± 0.03 a | 0.27 ± 0.02 a | 0.26 ± 0.03 a | 0.221 | 0.411 |
| Lung | 0.25 ± 0.03 a | 0.25 ± 0.02 a | 0.25 ± 0.04 a | 0.588 | 1.000 |
| Visceral fat * | 2.70 ± 1.21 a | 4.23 ± 0.79 b | 4.74 ± 1.37 b | 0.012 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takashima, M.; Tanaka, W.; Matsuyama, H.; Tajiri, H.; Sakakibara, H. Maternal Quercetin Consumption during Pregnancy May Help Regulate Total Cholesterol/HDL-Cholesterol Ratio without Effect on Cholesterol Levels in Male Progeny Consuming High-Fat Diet. Nutrients 2021, 13, 1242. https://doi.org/10.3390/nu13041242
Takashima M, Tanaka W, Matsuyama H, Tajiri H, Sakakibara H. Maternal Quercetin Consumption during Pregnancy May Help Regulate Total Cholesterol/HDL-Cholesterol Ratio without Effect on Cholesterol Levels in Male Progeny Consuming High-Fat Diet. Nutrients. 2021; 13(4):1242. https://doi.org/10.3390/nu13041242
Chicago/Turabian StyleTakashima, Masakatsu, Wataru Tanaka, Hiroki Matsuyama, Hayato Tajiri, and Hiroyuki Sakakibara. 2021. "Maternal Quercetin Consumption during Pregnancy May Help Regulate Total Cholesterol/HDL-Cholesterol Ratio without Effect on Cholesterol Levels in Male Progeny Consuming High-Fat Diet" Nutrients 13, no. 4: 1242. https://doi.org/10.3390/nu13041242