
Short-Term Calorie Restriction Maintains Plasma Insulin Concentrations along with a Reduction in Hepatic Insulin-Degrading Enzyme Levels in db/db Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment
2.2. Oral Glucose Tolerance Test
2.3. Tissue Collection
2.4. Plasma Glucose and Insulin Concentrations
2.5. Tissue Homogenization
2.6. Western Blotting
2.7. Glucose-Stimulated Insulin Secretion of Isolated Islets
2.8. Statistical Analysis
3. Results
3.1. Body Weight, Epididymal Fat Weight, and Total Food Intake
3.2. Fasting Plasma Glucose and Insulin Concentrations
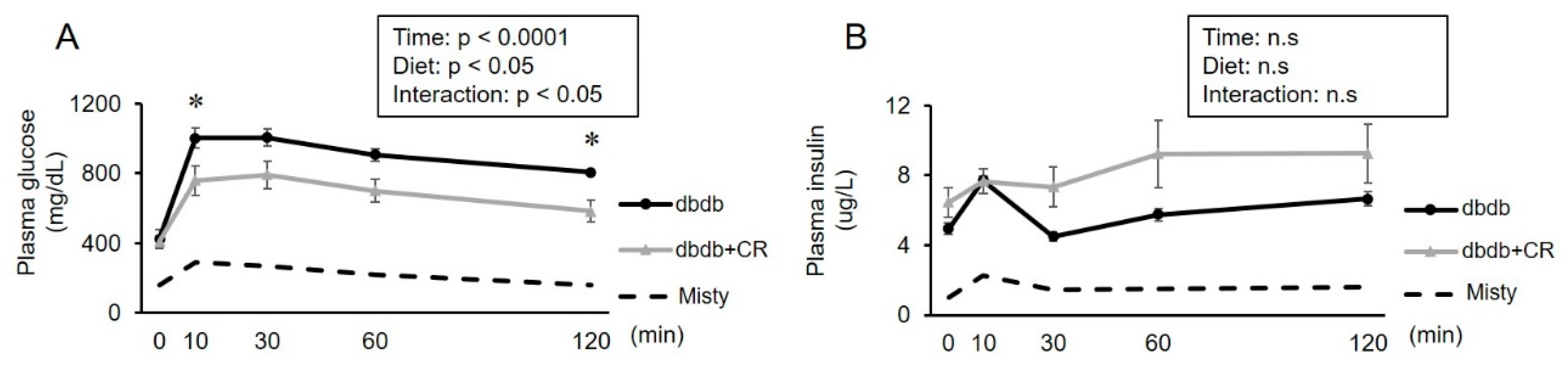
3.3. OGTT
3.4. GLUT-4 Protein Levels in TA Muscle and Epididymal Adipose Tissue
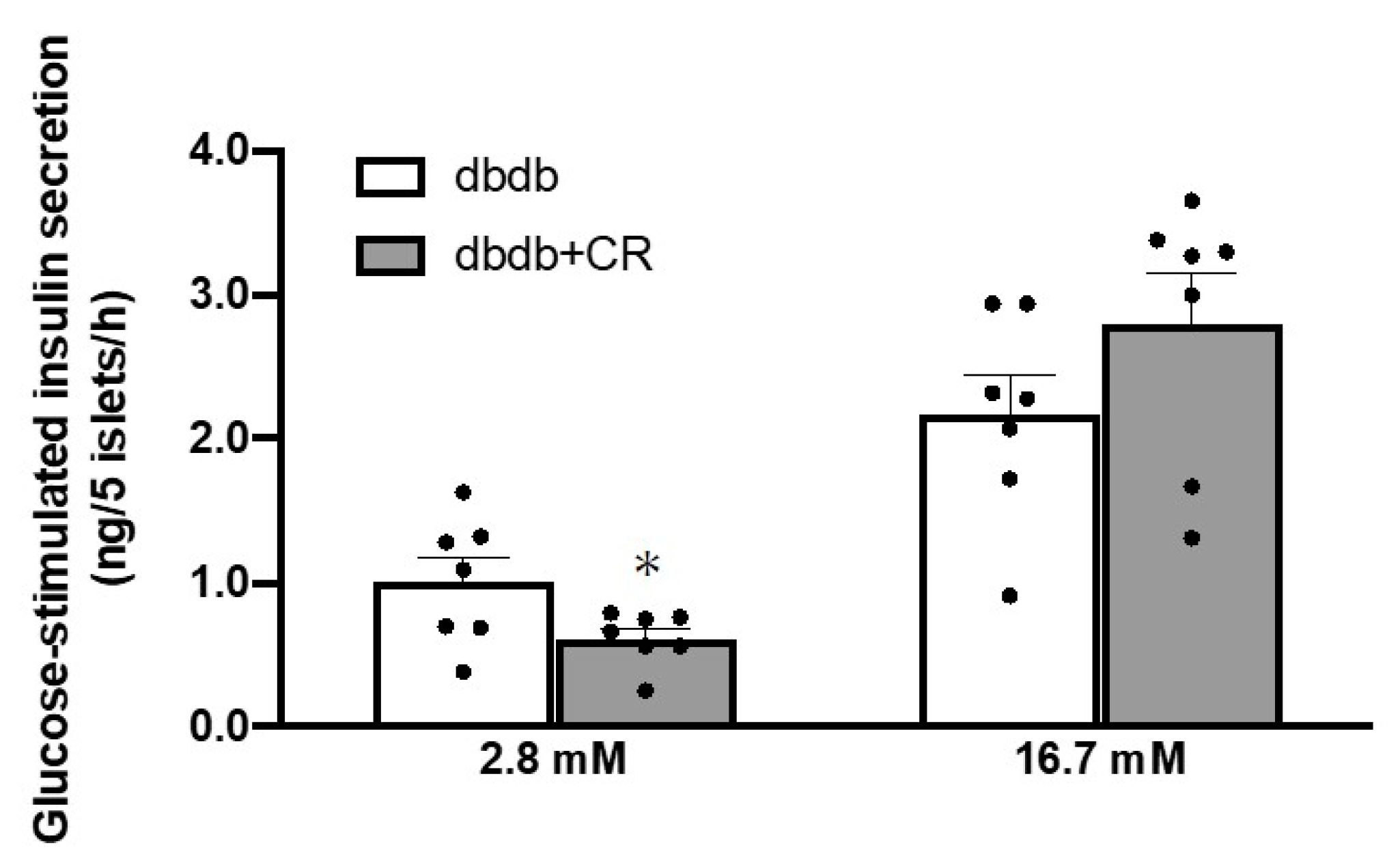
3.5. Glucose-Stimulated Insulin Secretion in Isolated Islets
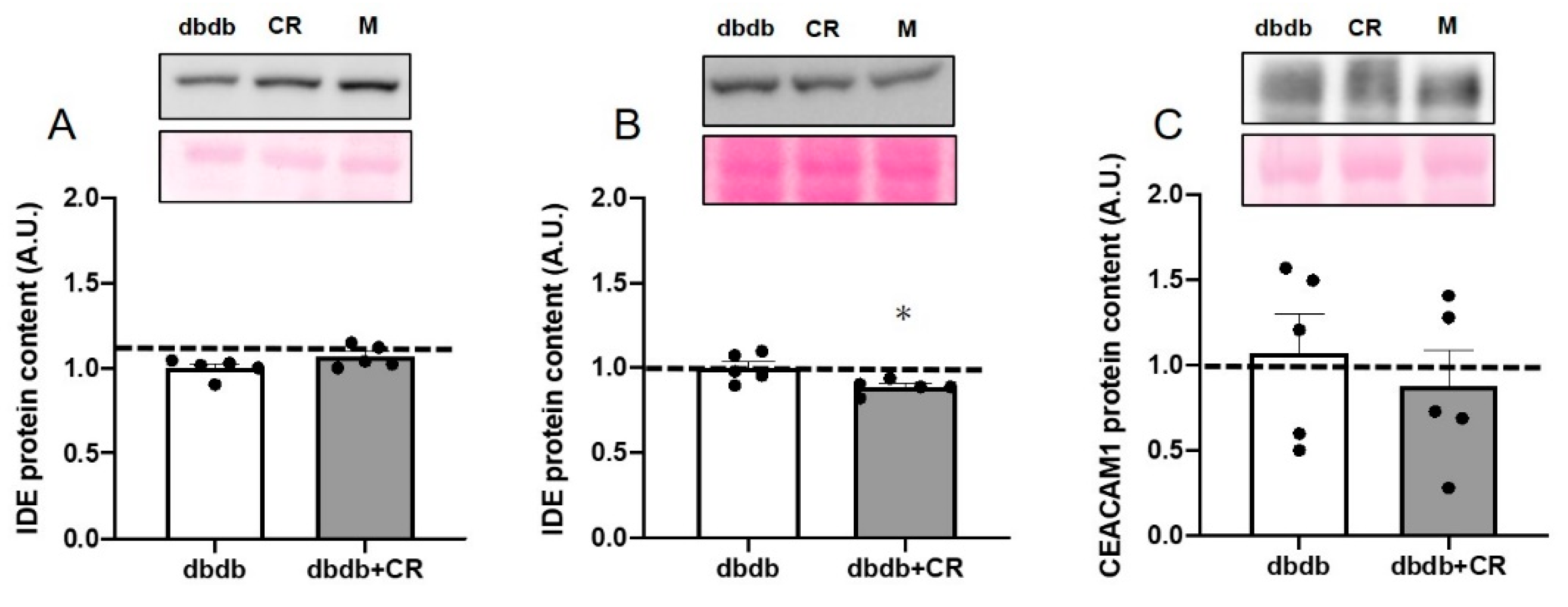
3.6. IDE Protein Levels in TA Muscle and Liver and CEACAM1 Protein Levels in Liver
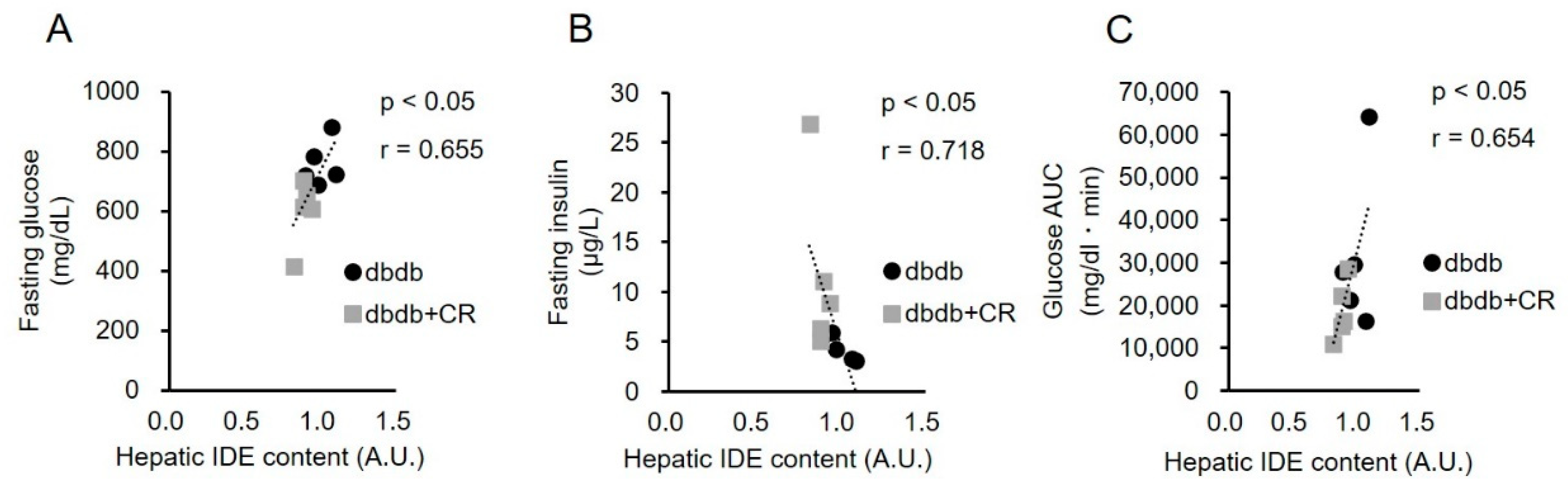
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Siddiqui, S. Obesity and diabetes: Interrelationship. Adv. Obes. Weight Manag. Control 2018, 8, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Nolte, L.A.; Hansen, P.A.; Han, D.-H.; Ferguson, K.; Thompson, P.A.; Holloszy, J.O. High-fat diet-induced muscle insulin resistance: Relationship to visceral fat mass. Am. J. Physiol. Integr. Comp. Physiol. 2000, 279, R2057–R2065. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, A.; Sumida, Y.; Urakawa, H.; Gabazza, E.C.; Murashima, S.; Maruyama, N.; Morioka, K.; Nakatani, K.; Yano, Y.; Adachi, Y. Increased visceral fat and serum levels of triglyceride are associated with insulin resistance in Japanese metabolically obese, normal weight subjects with normal glucose tolerance. Diabetes Care 2003, 26, 2341–2344. [Google Scholar] [CrossRef] [Green Version]
- Koska, J.; Stefan, N.; Permana, P.A.; Weyer, C.; Sonoda, M.; Bogardus, C.; Smith, S.R.; Joanisse, D.R.; Funahashi, T.; Krakoff, J.; et al. Increased fat accumulation in liver may link insulin resistance with subcutaneous abdominal adipocyte enlargement, visceral adiposity, and hypoadiponectinemia in obese individuals. Am. J. Clin. Nutr. 2008, 87, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, J.E.; Dohm, G.L.; Leggett-Frazier, N.; Elton, C.W.; Tapscott, E.B.; Pories, W.P.; Caro, J.F. Restoration of insulin responsiveness in skeletal muscle of morbidly obese patients after weight loss: Effect on muscle glucose transport and glucose transporter GLUT4. J. Clin. Invest. 1992, 89, 701–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Diabetes Association. Nutrition recommendations and principles for people with diabetes mellitus. Diabetes Care 1994, 17, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Petersen, K.F.; Dufour, S.; Morino, K.; Yoo, P.S.; Cline, G.W.; Shulman, G.I. Reversal of muscle insulin resistance by weight reduction in young, lean, insulin-resistant offspring of parents with type 2 diabetes. Proc. Natl. Acad. Sci. USA 2012, 109, 8236–8240. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.P.; Jordan, R.C.; Frese, E.M.; Albert, S.G.; Villareal, D.T.; Weiss, A.; Jordan, R.C.; Frese, E.M.; Albert, S.G.; Villareal, D.T. Effects of Weight Loss on Lean Mass, Strength, Bone, and Aerobic Capacity. Med. Sci. Sport Exerc. 2017, 49, 206–217. [Google Scholar] [CrossRef]
- Marchal, J.; Blanc, S.; Epelbaum, J.; Aujard, F.; Pifferi, F. Effects of chronic calorie restriction or dietary resveratrol supplementation on insulin sensitivity markers in a primate, Microcebus murinus. PLoS ONE 2012, 7, e34289. [Google Scholar] [CrossRef]
- Matyi, S.; Jackson, J.; Garrett, K.; Deepa, S.S.; Unnikrishnan, A. The effect of different levels of dietary restriction on glucose homeostasis and metabolic memory. GeroScience 2018, 40, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Zhao, J.; Bai, M.; Li, C.; Zhang, L.; Chen, Y. Comparison of glycemic improvement between intermittent calorie restriction and continuous calorie restriction in diabetic mice. Nutr. Metab. 2019, 16, 60. [Google Scholar] [CrossRef]
- Dean, D.J.; Brozinick, J.T.; Cushman, S.W.; Cartee, G.D. Calorie restriction increases cell surface GLUT-4 in insulin-stimulated skeletal muscle. Am. J. Physiol. 1998, 275, E957–E964. [Google Scholar] [CrossRef] [PubMed]
- Sequea, D.A.; Sharma, N.; Arias, E.B.; Cartee, G.D. Calorie restriction enhances insulin-stimulated glucose uptake and akt phosphorylation in both fast-twitch and slow-twitch skeletal muscle of 24-month-old rats. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2012, 67, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [CrossRef]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef]
- Smith, U. Impaired (’diabetic’) insulin signaling and action occur in fat cells long before glucose intolerance—Is insulin resistance initiated in the adipose tissue? Int. J. Obes. 2002, 26, 897–904. [Google Scholar] [CrossRef] [Green Version]
- King, B.C.; Blom, A.M. Non-traditional roles of complement in type 2 diabetes: Metabolism, insulin secretion and homeostasis. Mol. Immunol. 2017, 84, 34–42. [Google Scholar] [CrossRef]
- Cnop, M.; Welsh, N.; Jonas, J.C.; Jörns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic β-cell death in type 1 and type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54, S97–S107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dor, Y.; Glaser, B. Beta-Cell Dedifferentiation and Type 2 Diabetes. N. Engl. J. Med. 2013, 368, 572–573. [Google Scholar] [CrossRef]
- Guillausseau, P.J.; Meas, T.; Virally, M.; Laloi-Michelin, M.; Médeau, V.; Kevorkian, J.P. Abnormalities in insulin secretion in type 2 diabetes mellitus. Diabetes Metab. 2008, 34, S43–S48. [Google Scholar] [CrossRef]
- Gao, X.; Yan, D.; Zhao, Y.; Tao, H.; Zhou, Y. Moderate calorie restriction to achieve normal weight reverses β-cell dysfunction in diet-induced obese mice: Involvement of autophagy. Nutr. Metab. 2015, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Najjar, S.M.; Perdomo, G. Hepatic Insulin Clearance: Mechanism and Physiology. Physiology (Bethesda) 2019, 34, 198–215. [Google Scholar] [CrossRef] [PubMed]
- Rezende, L.F.; Camargo, R.L.; Branco, R.C.S.; Cappelli, A.P.G.; Boschero, A.C.; Carneiro, E.M. Reduced insulin clearance and lower insulin-degrading enzyme expression in the liver might contribute to the thrifty phenotype of protein-restricted mice. Br. J. Nutr. 2014, 112, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Kurauti, M.A.; Freitas-Dias, R.; Ferreira, S.M.; Vettorazzi, J.F.; Nardelli, T.R.; Araujo, H.N.; Santos, G.J.; Carneiro, E.M.; Boschero, A.C.; Rezende, L.F.; et al. Acute Exercise Improves Insulin Clearance and Increases the Expression of Insulin-Degrading Enzyme in the Liver and Skeletal Muscle of Swiss Mice. PLoS ONE 2016, 11, e0160239. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-W.; Villani, V.; Buono, R.; Wei, M.; Kumar, S.; Yilmaz, O.H.; Cohen, P.; Sneddon, J.B.; Perin, L.; Longo, V.D. Fasting-Mimicking Diet Promotes Ngn3-Driven β-Cell Regeneration to Reverse Diabetes. Cell 2017, 168, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, Y.; Takeda, R.; Kano, Y.; Hoshino, D. Effects of acute 3-h swimming exercise on insulin secretion capacity of pancreatic islets. J. Phys. Fit. Sport. Med. 2020, 9, 173–179. [Google Scholar] [CrossRef]
- Kawanaka, K.; Tabata, I.; Katsuta, S.; Higuchi, M. Changes in insulin-stimulated glucose transport and GLUT-4 protein in rat skeletal muscle after training. J. Appl. Physiol. 1997, 83, 2043–2047. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Choi, G.H.; Choi, H.I.; Ryu, J.; Jung, C.Y.; Lee, W. Calorie restriction improves whole-body glucose disposal and insulin resistance in association with the increased adipocyte-specific GLUT4 expression in Otsuka Long-Evans Tokushima Fatty rats. Arch. Biochem. Biophys. 2005, 436, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Tozzo, E.; Gnudi, L.; Kahn, B.B. Amelioration of Insulin Resistance in Streptozotocin Diabetic Mice by Transgenic Overexpression of GLUT4 Driven by an Adipose-Specific Promoter 1. Endocrinology 1997, 138, 1604–1611. [Google Scholar] [CrossRef] [PubMed]
- Farris, W.; Mansourian, S.; Chang, Y.; Lindsley, L.; Eckman, E.A.; Frosch, M.P.; Eckman, C.B.; Tanzi, R.E.; Selkoe, D.J.; Guénette, S. Insulin-degrading enzyme regulates the levels of insulin, amyloid β-protein, and the β-amyloid precursor protein intracellular domain in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 4162–4167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Hay, S.O.; Kang, D.; McBride, M.; Li, L.; Zhao, J.; Leissring, M.A. Deletion of Insulin-Degrading Enzyme Elicits Antipodal, Age-Dependent Effects on Glucose and Insulin Tolerance. PLoS ONE 2011, 6, e20818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Ghani, M.A. Contributions of beta-Cell Dysfunction and Insulin Resistance to the Pathogenesis of Impaired Glucose Tolerance and Impaired Fasting Glucose. Diabetes Care 2006, 29, 1130–1139. [Google Scholar] [CrossRef]
- Wei, X.; Ke, B.; Zhao, Z.; Ye, X.; Gao, Z.; Ye, J. Regulation of insulin degrading enzyme activity by obesity-associated factors and pioglitazone in liver of diet-induced obese mice. PLoS ONE 2014, 9, e95399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, B.; Fernández-Díaz, C.M.; Parrado-Fernández, C.; González-Casimiro, C.M.; Postigo-Casado, T.; Lobatón, C.D.; Leissring, M.A.; Cózar-Castellano, I.; Perdomo, G. Hepatic insulin-degrading enzyme regulates glucose and insulin homeostasis in diet-induced obese mice. Metabolism 2020, 113, 154352. [Google Scholar] [CrossRef]
- Steneberg, P.; Bernardo, L.; Edfalk, S.; Lundberg, L.; Backlund, F.; Östenson, C.G.; Edlund, H. The type 2 diabetes-associated gene Ide is required for insulin secretion and suppression of α-synuclein levels in β-cells. Diabetes 2013, 62, 2004–2014. [Google Scholar] [CrossRef] [Green Version]
- Villa-Pérez, P.; Merino, B.; Fernández-Díaz, C.M.; Cidad, P.; Lobatón, C.D.; Moreno, A.; Muturi, H.T.; Ghadieh, H.E.; Najjar, S.M.; Leissring, M.A.; et al. Liver-specific ablation of insulin-degrading enzyme causes hepatic insulin resistance and glucose intolerance, without affecting insulin clearance in mice. Metabolism 2018, 88, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nagakura, T.; Yasuda, N.; Yamazaki, K.; Ikuta, H.; Tanaka, I. Enteroinsular axis of db/db mice and efficacy of dipeptidyl peptidase IV inhibition. Metabolism 2003, 52, 81–86. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Madrigal-Matute, J.; Scheibye-Knudsen, M.; Fang, E.; Aon, M.; González-Reyes, J.A.; Cortassa, S.; Kaushik, S.; Gonzalez-Freire, M.; Patel, B.; et al. Effects of Sex, Strain, and Energy Intake on Hallmarks of Aging in Mice. Cell Metab. 2016, 23, 1093–1112. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| dbdb | dbdb+CR | Misty | |
|---|---|---|---|
| Initial body weight (g) | 39.1 ± 0.7 | 39.2 ± 0.7 | 20.0 ± 0.3 |
| Final body weight (g) | 39.0 ± 0.7 | 33.4 ± 0.9 ** | 21.2 ± 0.2 |
| Total food intake (g) | 107 ± 4 | 64 ± 1*** | 72 ± 1 |
| Liver weight (g) | 2.1 ± 0.1 | 1.9 ± 0.1 ** | 1.3 ± 0.1 |
| Epidydimal fat weight (g) | 1.7 ± 0.1 | 1.5 ± 0.1 * | 0.3 ± 0.1 |
| Plasma glucose concentration (10 weeks) (mg/dL) | 672 ± 18 | 644 ± 67 | 167 ± 7 |
| Plasma glucose concentration (13 weeks) (mg/dL) | 759 ± 34 | 599 ± 24 * | 199 ± 6 |
| ΔPlasma glucose concentration (13 vs.10 weeks) (mg/dL) | 87 ± 28 | −45 ± 36 * | 32 ± 7 |
| Plasma insulin concentration (10 weeks) (µg/L) | 8.2 ± 1.7 | 9.5 ± 2.5 | 0.5 ± 0.1 |
| Plasma insulin concentration (13 weeks) (µg/L) | 4.4 ± 0.6 | 11.6 ± 4.0 | 0.3 ± 0.1 |
| ΔPlasma insulin concentration (13 vs. 10 weeks) (µg/L) | −3.7 ± 1.8 | 2.1 ± 1.7 * | −0.2 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonaka, Y.; Takeda, R.; Kano, Y.; Hoshino, D. Short-Term Calorie Restriction Maintains Plasma Insulin Concentrations along with a Reduction in Hepatic Insulin-Degrading Enzyme Levels in db/db Mice. Nutrients 2021, 13, 1190. https://doi.org/10.3390/nu13041190
Nonaka Y, Takeda R, Kano Y, Hoshino D. Short-Term Calorie Restriction Maintains Plasma Insulin Concentrations along with a Reduction in Hepatic Insulin-Degrading Enzyme Levels in db/db Mice. Nutrients. 2021; 13(4):1190. https://doi.org/10.3390/nu13041190
Chicago/Turabian StyleNonaka, Yudai, Reo Takeda, Yutaka Kano, and Daisuke Hoshino. 2021. "Short-Term Calorie Restriction Maintains Plasma Insulin Concentrations along with a Reduction in Hepatic Insulin-Degrading Enzyme Levels in db/db Mice" Nutrients 13, no. 4: 1190. https://doi.org/10.3390/nu13041190