
Iron Deficiency without Anemia Decreases Physical Endurance and Mitochondrial Complex I Activity of Oxidative Skeletal Muscle in the Mouse


 , and
, and 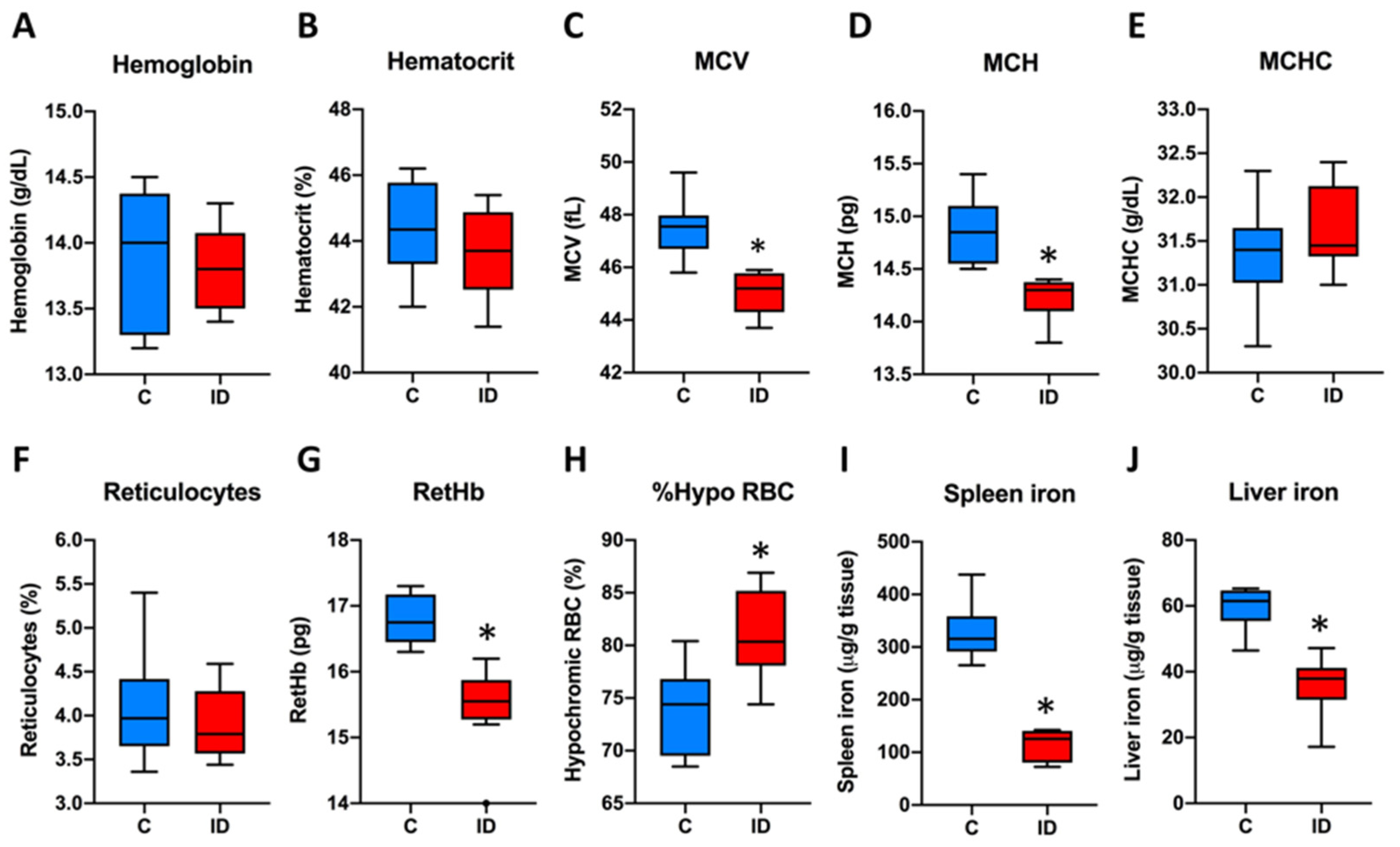{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Experimental Design
2.3. Physical Tests
2.4. Hematological Parameters
2.5. Tissue Iron Content
2.6. Mitochondrial Enzymatic Activities
2.7. Statistical Analysis
3. Results
3.1. Description of the Mouse Model of ID
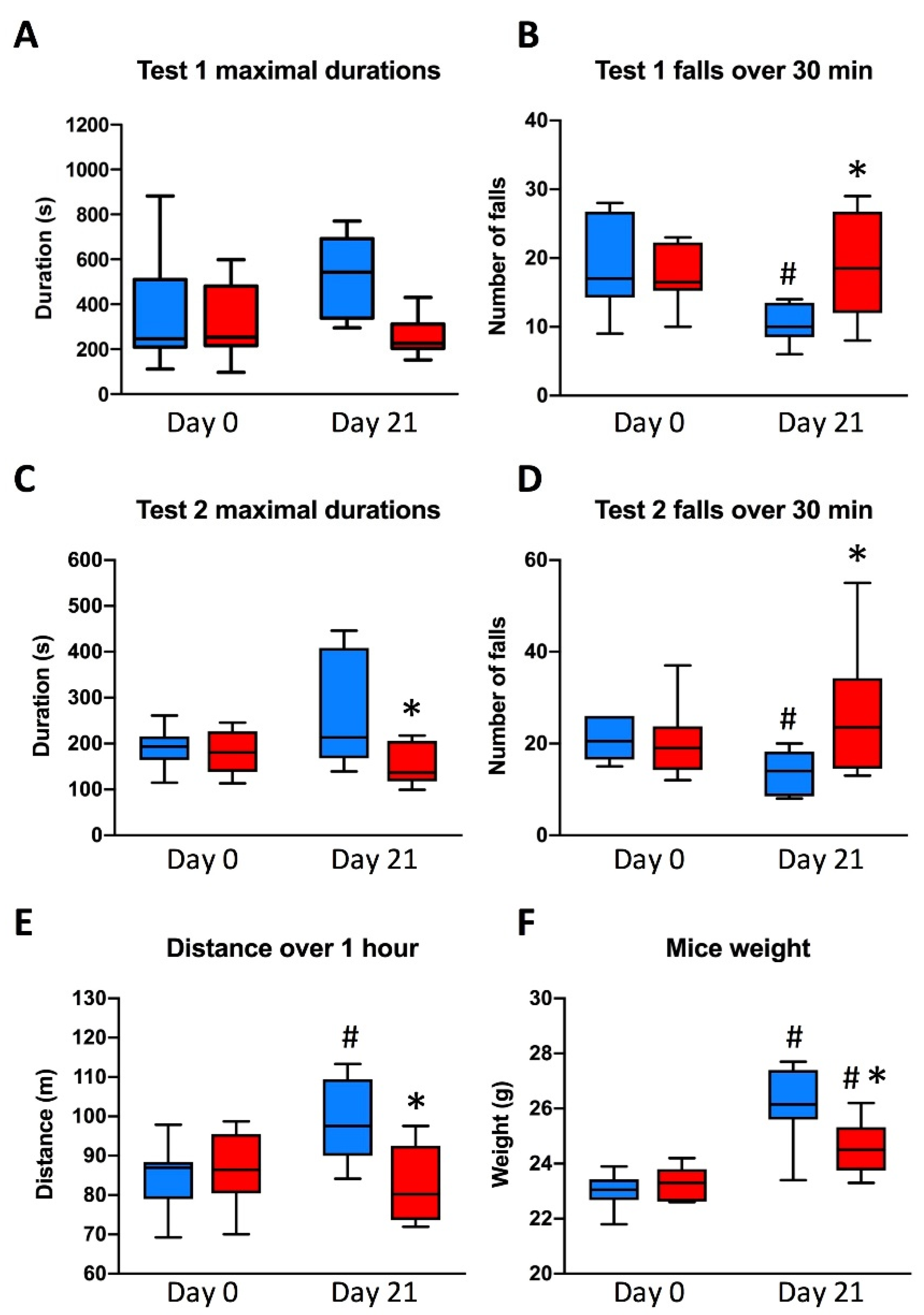
3.2. Animal Physical Endurance
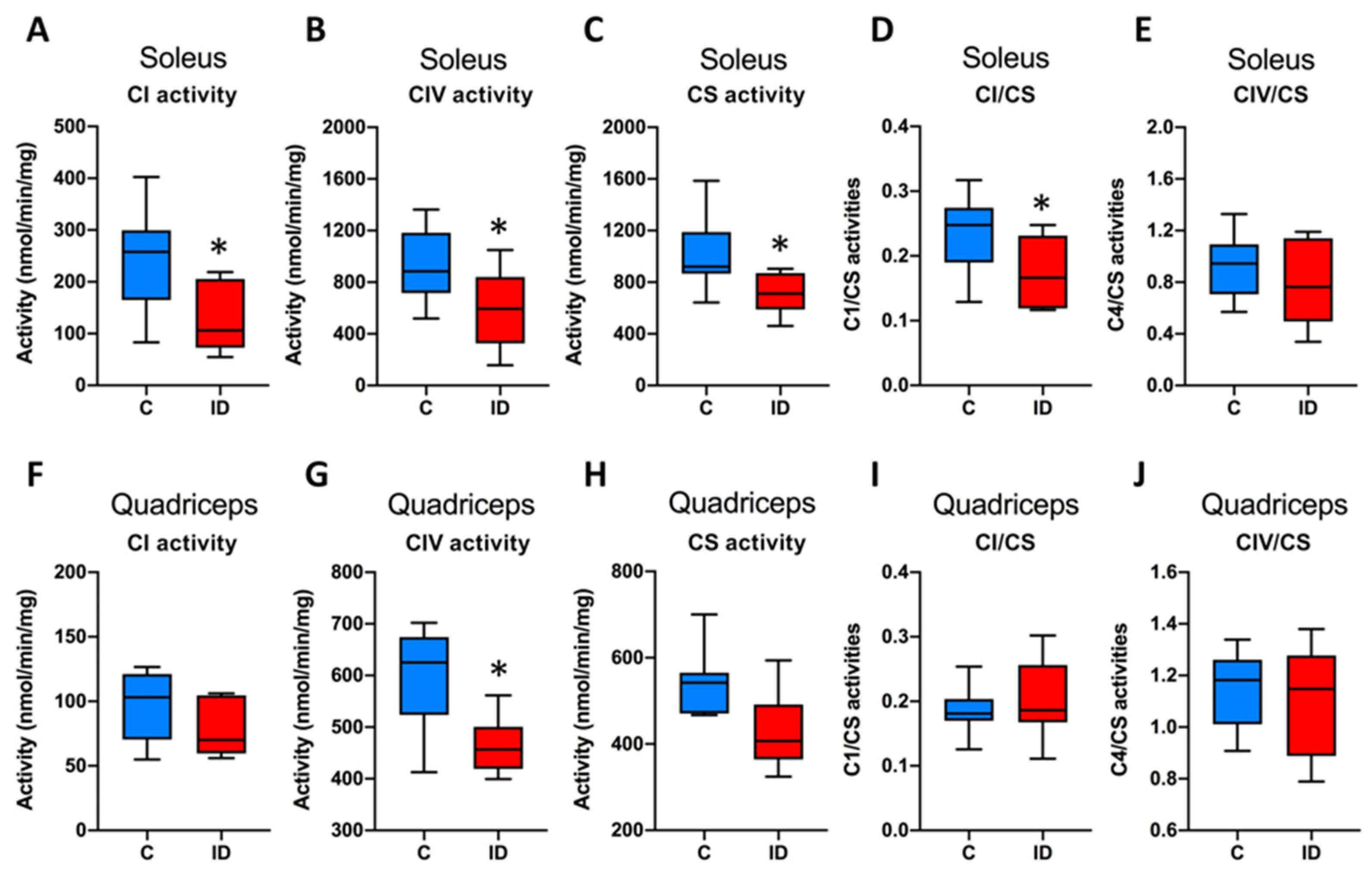
3.3. Mitochondrial Enzymatic Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krayenbuehl, P.-A.; Battegay, E.; Breymann, C.; Furrer, J.; Schulthess, G. Intravenous Iron for the Treatment of Fatigue in Nonanemic, Premenopausal Women with Low Serum Ferritin Concentration. Blood 2011, 118, 3222–3227. [Google Scholar] [CrossRef]
- Favrat, B.; Balck, K.; Breymann, C.; Hedenus, M.; Keller, T.; Mezzacasa, A.; Gasche, C. Evaluation of a Single Dose of Ferric Carboxymaltose in Fatigued, Iron-Deficient Women--PREFER a Randomized, Placebo-Controlled Study. PLoS ONE 2014, 9, e94217. [Google Scholar] [CrossRef]
- Piednoir, P.; Allou, N.; Driss, F.; Longrois, D.; Philip, I.; Beaumont, C.; Montravers, P.; Lasocki, S. Preoperative Iron Deficiency Increases Transfusion Requirements and Fatigue in Cardiac Surgery Patients: A Prospective Observational Study. Eur. J. Anaesthesiol. 2011, 28, 796–801. [Google Scholar] [CrossRef]
- Lasocki, S.; Chudeau, N.; Papet, T.; Tartiere, D.; Roquilly, A.; Carlier, L.; Mimoz, O.; Seguin, P.; Malledant, Y.; Asehnoune, K.; et al. Prevalence of Iron Deficiency on ICU Discharge and Its Relation with Fatigue: A Multicenter Prospective Study. Crit. Care Lond. Engl. 2014, 18, 542. [Google Scholar] [CrossRef]
- Brownlie, T.; Utermohlen, V.; Hinton, P.S.; Giordano, C.; Haas, J.D. Marginal Iron Deficiency without Anemia Impairs Aerobic Adaptation among Previously Untrained Women. Am. J. Clin. Nutr. 2002, 75, 734–742. [Google Scholar] [CrossRef]
- Zhu, Y.I.; Haas, J.D. Iron Depletion without Anemia and Physical Performance in Young Women. Am. J. Clin. Nutr. 1997, 66, 334–341. [Google Scholar] [CrossRef]
- DellaValle, D.M.; Haas, J.D. Impact of Iron Depletion without Anemia on Performance in Trained Endurance Athletes at the Beginning of a Training Season: A Study of Female Collegiate Rowers. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Hinton, P.S.; Giordano, C.; Brownlie, T.; Haas, J.D. Iron Supplementation Improves Endurance after Training in Iron-Depleted, Nonanemic Women. J. Appl. Physiol. (1985) 2000, 88, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.A.; Rozentryt, P.; Witkowska, A.; Nowak, J.; Hartmann, O.; Ponikowska, B.; Borodulin-Nadzieja, L.; Banasiak, W.; Polonski, L.; Filippatos, G.; et al. Iron Deficiency: An Ominous Sign in Patients with Systolic Chronic Heart Failure. Eur. Heart J. 2010, 31, 1872–1880. [Google Scholar] [CrossRef] [PubMed]
- von Haehling, S.; Gremmler, U.; Krumm, M.; Mibach, F.; Schön, N.; Taggeselle, J.; Dahm, J.B.; Angermann, C.E. Prevalence and Clinical Impact of Iron Deficiency and Anaemia among Outpatients with Chronic Heart Failure: The PrEP Registry. Clin. Res. Cardiol. Off. J. Ger. Card. Soc. 2017, 106, 436–443. [Google Scholar] [CrossRef]
- Klip, I.T.; Comin-Colet, J.; Voors, A.A.; Ponikowski, P.; Enjuanes, C.; Banasiak, W.; Lok, D.J.; Rosentryt, P.; Torrens, A.; Polonski, L.; et al. Iron Deficiency in Chronic Heart Failure: An International Pooled Analysis. Am. Heart J. 2013, 165, 575–582.e3. [Google Scholar] [CrossRef] [PubMed]
- Bekfani, T.; Pellicori, P.; Morris, D.; Ebner, N.; Valentova, M.; Sandek, A.; Doehner, W.; Cleland, J.G.; Lainscak, M.; Schulze, P.C.; et al. Iron Deficiency in Patients with Heart Failure with Preserved Ejection Fraction and Its Association with Reduced Exercise Capacity, Muscle Strength and Quality of Life. Clin. Res. Cardiol. Off. J. Ger. Card. Soc. 2019, 108, 203–211. [Google Scholar] [CrossRef]
- Toblli, J.E.; Lombraña, A.; Duarte, P.; Di Gennaro, F. Intravenous Iron Reduces NT-pro-Brain Natriuretic Peptide in Anemic Patients with Chronic Heart Failure and Renal Insufficiency. J. Am. Coll. Cardiol. 2007, 50, 1657–1665. [Google Scholar] [CrossRef]
- Okonko, D.O.; Grzeslo, A.; Witkowski, T.; Mandal, A.K.J.; Slater, R.M.; Roughton, M.; Foldes, G.; Thum, T.; Majda, J.; Banasiak, W.; et al. Effect of Intravenous Iron Sucrose on Exercise Tolerance in Anemic and Nonanemic Patients with Symptomatic Chronic Heart Failure and Iron Deficiency FERRIC-HF: A Randomized, Controlled, Observer-Blinded Trial. J. Am. Coll. Cardiol. 2008, 51, 103–112. [Google Scholar] [CrossRef]
- Anker, S.D.; Comin Colet, J.; Filippatos, G.; Willenheimer, R.; Dickstein, K.; Drexler, H.; Lüscher, T.F.; Bart, B.; Banasiak, W.; Niegowska, J.; et al. Ferric Carboxymaltose in Patients with Heart Failure and Iron Deficiency. N. Engl. J. Med. 2009, 361, 2436–2448. [Google Scholar] [CrossRef] [PubMed]
- Filippatos, G.; Farmakis, D.; Colet, J.C.; Dickstein, K.; Lüscher, T.F.; Willenheimer, R.; Parissis, J.; Gaudesius, G.; Mori, C.; von Eisenhart Rothe, B.; et al. Intravenous Ferric Carboxymaltose in Iron-Deficient Chronic Heart Failure Patients with and without Anaemia: A Subanalysis of the FAIR-HF Trial. Eur. J. Heart Fail. 2013, 15, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- van Veldhuisen, D.J.; Ponikowski, P.; van der Meer, P.; Metra, M.; Böhm, M.; Doletsky, A.; Voors, A.A.; Macdougall, I.C.; Anker, S.D.; Roubert, B.; et al. Effect of Ferric Carboxymaltose on Exercise Capacity in Patients With Chronic Heart Failure and Iron Deficiency. Circulation 2017, 136, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Finch, C.A.; Miller, L.R.; Inamdar, A.R.; Person, R.; Seiler, K.; Mackler, B. Iron Deficiency in the Rat. Physiological and Biochemical Studies of Muscle Dysfunction. J. Clin. Investig. 1976, 58, 447–453. [Google Scholar] [CrossRef]
- Davies, K.J.; Maguire, J.J.; Brooks, G.A.; Dallman, P.R.; Packer, L. Muscle Mitochondrial Bioenergetics, Oxygen Supply, and Work Capacity during Dietary Iron Deficiency and Repletion. Am. J. Physiol. 1982, 242, E418–E427. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J.; Donovan, C.M.; Refino, C.J.; Brooks, G.A.; Packer, L.; Dallman, P.R. Distinguishing Effects of Anemia and Muscle Iron Deficiency on Exercise Bioenergetics in the Rat. Am. J. Physiol. 1984, 246, E535–E543. [Google Scholar] [CrossRef]
- Willis, W.T.; Brooks, G.A.; Henderson, S.A.; Dallman, P.R. Effects of Iron Deficiency and Training on Mitochondrial Enzymes in Skeletal Muscle. J. Appl. Physiol. (1985) 1987, 62, 2442–2446. [Google Scholar] [CrossRef]
- Blayney, L.; Bailey-Wood, R.; Jacobs, A.; Henderson, A.; Muir, J. The Effects of Iron Deficiency on the Respiratory Function and Cytochrome Content of Rat Heart Mitochondria. Circ. Res. 1976, 39, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Rineau, E.; Gaillard, T.; Gueguen, N.; Procaccio, V.; Henrion, D.; Prunier, F.; Lasocki, S. Iron Deficiency without Anemia Is Responsible for Decreased Left Ventricular Function and Reduced Mitochondrial Complex I Activity in a Mouse Model. Int. J. Cardiol. 2018, 266, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Melenovsky, V.; Hlavata, K.; Sedivy, P.; Dezortova, M.; Borlaug, B.A.; Petrak, J.; Kautzner, J.; Hajek, M. Skeletal Muscle Abnormalities and Iron Deficiency in Chronic Heart FailureAn Exercise 31P Magnetic Resonance Spectroscopy Study of Calf Muscle. Circ. Heart Fail. 2018, 11, e004800. [Google Scholar] [CrossRef]
- Charles-Edwards, G.; Amaral, N.; Sleigh, A.; Ayis, S.; Catibog, N.; McDonagh, T.; Monaghan, M.; Amin-Youssef, G.; Kemp, G.J.; Shah, A.M.; et al. Effect of Iron Isomaltoside on Skeletal Muscle Energetics in Patients With Chronic Heart Failure and Iron Deficiency. Circulation 2019, 139, 2386–2398. [Google Scholar] [CrossRef] [PubMed]
- Burnes, L.A.; Kolker, S.J.; Danielson, J.F.; Walder, R.Y.; Sluka, K.A. Enhanced Muscle Fatigue Occurs in Male but Not Female ASIC3-/- Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1347–R1355. [Google Scholar] [CrossRef]
- Lasocki, S.; Millot, S.; Andrieu, V.; Lettéron, P.; Pilard, N.; Muzeau, F.; Thibaudeau, O.; Montravers, P.; Beaumont, C. Phlebotomies or Erythropoietin Injections Allow Mobilization of Iron Stores in a Mouse Model Mimicking Intensive Care Anemia. Crit. Care Med. 2008, 36, 2388–2394. [Google Scholar] [CrossRef]
- Medja, F.; Allouche, S.; Frachon, P.; Jardel, C.; Malgat, M.; Mousson de Camaret, B.; Slama, A.; Lunardi, J.; Mazat, J.P.; Lombès, A. Development and Implementation of Standardized Respiratory Chain Spectrophotometric Assays for Clinical Diagnosis. Mitochondrion 2009, 9, 331–339. [Google Scholar] [CrossRef]
- McLane, J.A.; Fell, R.D.; McKay, R.H.; Winder, W.W.; Brown, E.B.; Holloszy, J.O. Physiological and Biochemical Effects of Iron Deficiency on Rat Skeletal Muscle. Am. J. Physiol. 1981, 241, C47–C54. [Google Scholar] [CrossRef] [PubMed]
- Brutsaert, T.D.; Hernandez-Cordero, S.; Rivera, J.; Viola, T.; Hughes, G.; Haas, J.D. Iron Supplementation Improves Progressive Fatigue Resistance during Dynamic Knee Extensor Exercise in Iron-Depleted, Nonanemic Women. Am. J. Clin. Nutr. 2003, 77, 441–448. [Google Scholar] [CrossRef]
- Ponikowski, P.; van Veldhuisen, D.J.; Comin-Colet, J.; Ertl, G.; Komajda, M.; Mareev, V.; McDonagh, T.; Parkhomenko, A.; Tavazzi, L.; Levesque, V.; et al. Beneficial Effects of Long-Term Intravenous Iron Therapy with Ferric Carboxymaltose in Patients with Symptomatic Heart Failure and Iron Deficiency†. Eur. Heart J. 2015, 36, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Augusto, V.; Padovani, C.; Eduardo, G.; Campos, R. Skeletal Muscle Fiber Types in C57BL6J Mice. J. Morphol. Sci. 2004, 21, 89–94. [Google Scholar]
- Hinchliffe, P.; Sazanov, L.A. Organization of Iron-Sulfur Clusters in Respiratory Complex I. Science 2005, 309, 771–774. [Google Scholar] [CrossRef]
- Klempa, K.L.; Willis, W.T.; Chengson, R.; Dallman, P.R.; Brooks, G.A. Iron Deficiency Decreases Gluconeogenesis in Isolated Rat Hepatocytes. J. Appl. Physiol. (1985) 1989, 67, 1868–1872. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.A.; Dallman, P.R.; Brooks, G.A. Glucose Turnover and Oxidation Are Increased in the Iron-Deficient Anemic Rat. Am. J. Physiol. 1986, 250, E414–E421. [Google Scholar] [CrossRef]
- Eisenstein, R.S. Iron Regulatory Proteins and the Molecular Control of Mammalian Iron Metabolism. Annu. Rev. Nutr. 2000, 20, 627–662. [Google Scholar] [CrossRef]
- Cartier, L.J.; Ohira, Y.; Chen, M.; Cuddihee, R.W.; Holloszy, J.O. Perturbation of Mitochondrial Composition in Muscle by Iron Deficiency. Implications Regarding Regulation of Mitochondrial Assembly. J. Biol. Chem. 1986, 261, 13827–13832. [Google Scholar] [CrossRef]
- Dong, F.; Zhang, X.; Culver, B.; Chew, H.G.; Kelley, R.O.; Ren, J. Dietary Iron Deficiency Induces Ventricular Dilation, Mitochondrial Ultrastructural Aberrations and Cytochrome c Release: Involvement of Nitric Oxide Synthase and Protein Tyrosine Nitration. Clin. Sci. 2005, 109, 277–286. [Google Scholar] [CrossRef]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Schroder, H.D.; Boushel, R.; Helge, J.W.; Dela, F.; et al. Biomarkers of Mitochondrial Content in Skeletal Muscle of Healthy Young Human Subjects. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; González-Juanatey, J.R.; Harjola, V.-P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the Diagnosis and Treatment of Acute and Chronic Heart Failure: The Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure of the European Society of Cardiology (ESC). Developed with the Special Contribution of the Heart Failure Association (HFA) of the ESC. Eur. J. Heart Fail. 2016, 18, 891–975. [Google Scholar] [CrossRef]
- Galante, M.; Jani, H.; Vanes, L.; Daniel, H.; Fisher, E.M.C.; Tybulewicz, V.L.J.; Bliss, T.V.P.; Morice, E. Impairments in Motor Coordination without Major Changes in Cerebellar Plasticity in the Tc1 Mouse Model of Down Syndrome. Hum. Mol. Genet. 2009, 18, 1449–1463. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Chen, X.; Yan, H.; Deurenberg, P.; Garby, L.; Hautvast, J.G. Functional Consequences of Iron Supplementation in Iron-Deficient Female Cotton Mill Workers in Beijing, China. Am. J. Clin. Nutr. 1994, 59, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Crouter, S.E.; DellaValle, D.M.; Haas, J.D. Relationship between Physical Activity, Physical Performance, and Iron Status in Adult Women. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2012, 37, 697–705. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rineau, E.; Gueguen, N.; Procaccio, V.; Geneviève, F.; Reynier, P.; Henrion, D.; Lasocki, S. Iron Deficiency without Anemia Decreases Physical Endurance and Mitochondrial Complex I Activity of Oxidative Skeletal Muscle in the Mouse. Nutrients 2021, 13, 1056. https://doi.org/10.3390/nu13041056
Rineau E, Gueguen N, Procaccio V, Geneviève F, Reynier P, Henrion D, Lasocki S. Iron Deficiency without Anemia Decreases Physical Endurance and Mitochondrial Complex I Activity of Oxidative Skeletal Muscle in the Mouse. Nutrients. 2021; 13(4):1056. https://doi.org/10.3390/nu13041056
Chicago/Turabian StyleRineau, Emmanuel, Naïg Gueguen, Vincent Procaccio, Franck Geneviève, Pascal Reynier, Daniel Henrion, and Sigismond Lasocki. 2021. "Iron Deficiency without Anemia Decreases Physical Endurance and Mitochondrial Complex I Activity of Oxidative Skeletal Muscle in the Mouse" Nutrients 13, no. 4: 1056. https://doi.org/10.3390/nu13041056