
Intrauterine Transfer of Polyunsaturated Fatty Acids in Mother–Infant Dyads as Analyzed at Time of Delivery


 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Collection
2.3. Fatty Acid Analysis
2.4. Clinical and Demographic Data
2.5. Statistical Analysis
3. Results
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Demmelmair, H.; Koletzko, B. Importance of fatty acids in the perinatal period. World Rev. Nutr. Diet. 2015, 112, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, J.A.; Mori, T.A.; Barden, A.; Beilin, L.J.; Holt, P.G.; Calder, P.C.; Taylor, A.L.; Prescott, S.L. Effects of n-3 polyunsaturated fatty acid supplementation in pregnancy on maternal and fetal erythrocyte fatty acid composition. Eur. J. Clin. Nutr. 2004, 58, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Fowden, A.L. The placenta: A multifaceted, transient organ. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larqué, E.; Demmelmair, H.; Gil-Sánchez, A.; Prieto-Sánchez, M.T.; Blanco, J.E.; Pagán, A.; Faber, F.L.; Zamora, S.; Parrilla, J.J.; Koletzko, B. Placental transfer of fatty acids and fetal implications. Am. J. Clin. Nutr. 2011, 94, 1908s–1913s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.M.; Childs, C.E.; Calder, P.C. New perspectives on placental fatty acid transfer. Prostaglandins Leukot. Essent. Fat. Acids 2018, 138, 24–29. [Google Scholar] [CrossRef]
- Herrera, E.; Ortega-Senovilla, H. Lipid metabolism during pregnancy and its implications for fetal growth. Curr. Pharm. Biotechnol. 2014, 15, 24–31. [Google Scholar] [CrossRef]
- Marceau, G.; Gallot, D.; Lemery, D.; Sapin, V. Metabolism of retinol during mammalian placental and embryonic development. Vitam. Horm. 2007, 75, 97–115. [Google Scholar] [CrossRef]
- Fonseca, F.; Mucci, D.B.; Assumpção, R.P.; Marcondes, H.; Sardinha, F.L.C.; Silva, S.V.; Citelli, M.; Tavares do Carmo, M.D.G. Differential Long-Chain Polyunsaturated Fatty Acids Status and Placental Transport in Adolescent Pregnancies. Nutrients 2018, 10, 220. [Google Scholar] [CrossRef] [Green Version]
- Desoye, G.; Gauster, M.; Wadsack, C. Placental transport in pregnancy pathologies. Am. J. Clin. Nutr. 2011, 94, 1896s–1902s. [Google Scholar] [CrossRef] [Green Version]
- Herrera, E.; Ortega-Senovilla, H. Implications of Lipids in Neonatal Body Weight and Fat Mass in Gestational Diabetic Mothers and Non-Diabetic Controls. Curr. Diabetes Rep. 2018, 18, 7. [Google Scholar] [CrossRef]
- Jones, M.L.; Mark, P.J.; Waddell, B.J. Maternal dietary omega-3 fatty acids and placental function. Reproduction 2014, 147, R143–R152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larqué, E.; Demmelmair, H.; Berger, B.; Hasbargen, U.; Koletzko, B. In vivo investigation of the placental transfer of (13)C-labeled fatty acids in humans. J. Lipid Res. 2003, 44, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Meher, A.; Randhir, K.; Mehendale, S.; Wagh, G.; Joshi, S. Maternal Fatty Acids and Their Association with Birth Outcome: A Prospective Study. PLoS ONE 2016, 11, e0147359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Kawabata, T.; Kagawa, Y.; Kimura, F.; Miyazawa, T.; Tatsuta, N.; Saito, S.; Arima, T.; Yaegashi, N.; Nakai, K. Relationships between docosahexaenoic acid compositions of maternal and umbilical cord erythrocytes in pregnant Japanese women. Prostaglandins Leukot. Essent. Fat. Acids 2019, 147, 1–5. [Google Scholar] [CrossRef]
- Smuts, C.M.; Huang, M.; Mundy, D.; Plasse, T.; Major, S.; Carlson, S.E. A randomized trial of docosahexaenoic acid supplementation during the third trimester of pregnancy. Obs. Gynecol. 2003, 101, 469–479. [Google Scholar] [CrossRef]
- Al, M.D.M.; Houwelingen, A.C.V.; Hornstra, G. Relation between birth order and the maternal and neonatal docosahexaenoic acid status. Eur. J. Clin. Nutr. 1997, 51, 548–553. [Google Scholar] [CrossRef] [Green Version]
- American College of Obstetricians and Gynecologists Committee. Opinion No 579: Definition of term pregnancy. Obs. Gynecol. 2013, 122, 1139–1140. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.C.; Sampson, L.; Stampfer, M.J.; Rosner, B.; Bain, C.; Witschi, J.; Hennekens, C.H.; Speizer, F.E. Reproducibility and validity of a semiquantitative food frequency questionnaire. Am. J. Epidemiol. 1985, 122, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.; Hein, N.; Hanson, C.; Smith, L.M.; Anderson-Berry, A.; Richter, C.K.; Stessy Bisselou, K.; Kusi Appiah, A.; Kris-Etherton, P.; Skulas-Ray, A.C.; et al. Omega-3 Fatty Acid Intake by Age, Gender, and Pregnancy Status in the United States: National Health and Nutrition Examination Survey 2003–2014. Nutrients 2019, 11, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordgren, T.M.; Lyden, E.; Anderson-Berry, A.; Hanson, C. Omega-3 Fatty Acid Intake of Pregnant Women and Women of Childbearing Age in the United States: Potential for Deficiency? Nutrients 2017, 9, 197. [Google Scholar] [CrossRef] [Green Version]
- Jasani, B.; Simmer, K.; Patole, S.K.; Rao, S.C. Long chain polyunsaturated fatty acid supplementation in infants born at term. Cochrane Database Syst Rev. 2017, 3, CD000376. [Google Scholar] [CrossRef]
- Mennitti, L.V.; Oliveira, J.L.; Morais, C.A.; Estadella, D.; Oyama, L.M.; Oller do Nascimento, C.M.; Pisani, L.P. Type of fatty acids in maternal diets during pregnancy and/or lactation and metabolic consequences of the offspring. J. Nutr. Biochem. 2015, 26, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Tallima, H.; El Ridi, R. Arachidonic acid: Physiological roles and potential health benefits—A review. J. Adv. Res. 2018, 11, 33–41. [Google Scholar] [CrossRef]
- Wang, W.; Yang, J.; Edin, M.L.; Wang, Y.; Luo, Y.; Wan, D.; Yang, H.; Song, C.-Q.; Xue, W.; Sanidad, K.Z.; et al. Targeted Metabolomics Identifies the Cytochrome P450 Monooxygenase Eicosanoid Pathway as a Novel Therapeutic Target of Colon Tumorigenesis. Cancer Res. 2019, 79, 1822–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsani, E.; Della Vedova, A.M.; Rezzani, R.; Rodella, L.F.; Cristini, C. Correlation between human nervous system development and acquisition of fetal skills: An overview. Brain Dev. 2019, 41, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Soothill, P.W.; Nicolaides, K.H.; Rodeck, C.H.; Gamsu, H. Blood gases and acid-base status of the human second-trimester fetus. Obs. Gynecol. 1986, 68, 173–176. [Google Scholar] [CrossRef]
- Haggarty, P. Effect of placental function on fatty acid requirements during pregnancy. Eur. J. Clin. Nutr. 2004, 58, 1559–1570. [Google Scholar] [CrossRef] [Green Version]
- Cetin, I.; Alvino, G.; Cardellicchio, M. Long chain fatty acids and dietary fats in fetal nutrition. J. Physiol. 2009, 587, 3441–3451. [Google Scholar] [CrossRef]
- Elliott, E.; Hanson, C.K.; Anderson-Berry, A.L.; Nordgren, T.M. The role of specialized pro-resolving mediators in maternal-fetal health. Prostaglandins Leukot. Essent. Fat. Acids 2017, 126, 98–104. [Google Scholar] [CrossRef]
- Bannenberg, G.; Serhan, C.N. Specialized pro-resolving lipid mediators in the inflammatory response: An update. Biochim. Biophys. Acta 2010, 1801, 1260–1273. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhivaki, D.; Lo-Man, R. Unique aspects of the perinatal immune system. Nat. Rev. Immunol. 2017, 17, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, W.F. Essential fatty acid requirements in infancy. Am. J. Clin. Nutr. 1976, 29, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in US women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Median | IQR | Min | Max | |
|---|---|---|---|---|
| Infant CGA (weeks) | 39.45 | 39.0–40.2 | 37.4 | 41.0 |
| Infant birthweight (g) | 3467 | 3172.5–3758.0 | 2955 | 4893 |
| Infant birth length (cm) | 50.50 | 48.3–50.8 | 45.7 | 53.3 |
| Infant birth head circumference (cm) | 34.60 | 33.0–35.6 | 32.4 | 36.8 |
| Maternal pre-pregnancy BMI (kg/m2) | 24.7 | 21.2–28.9 | 18.2 | 43.9 |
| Count (n) | Percentage (%) | |||
| Infant sex: | ||||
| Male | 11 | 55.0 | ||
| Female | 9 | 45.0 | ||
| Maternal race: | ||||
| White | 10 | 50.0 | ||
| Black/African American | 4 | 20.0 | ||
| Hispanic/Latino | 3 | 15.0 | ||
| Asian/Pacific Islander | 1 | 5.0 | ||
| Native American/Alaskan | 0 | 0.0 | ||
| Other/Unknown | 2 | 10.0 | ||
| Delivery mode: | ||||
| Vaginal | 18 | 90.0 | ||
| Caesarean | 2 | 10.0 |
| Median | IQR | 33rd %ile | 67th %ile | Min | Max | |
|---|---|---|---|---|---|---|
| Linoleic Acid (µg/mL) | 1482.6 | 1322.0–1618.9 | 1342.4 | 1584.5 | 973.9 | 2122.3 |
| α-Linolenic Acid (µg/mL) | 35.6 | 28.5–39.4 | 33.8 | 39.4 | 24.2 | 52.5 |
| Arachidonic Acid (µg/mL) | 272.7 | 202.8–312.6 | 224.21 | 289.2 | 183.5 | 417.6 |
| Eicosapentaenoic Acid (µg/mL) | 6.0 | 4.0–9.3 | 5.0 | 7.2 | 2.0 | 18.5 |
| Docosapentaenoic-n3 Acid (µg/mL) | 10.1 | 9.0–12.0 | 9.2 | 12.0 | 7.0 | 20.8 |
| Docosahexaenoic Acid (µg/mL) | 72.7 | 59.4–81.7 | 62.4 | 80.4 | 37.6 | 161.3 |
| Total Fatty Acids (µg/mL) | 4794.0 | 4062.2–5208.2 | 4601.1 | 5093.7 | 3423.8 | 7138.1 |
| Median | IQR | Min | Max | |
|---|---|---|---|---|
| Linoleic Acid (µg/mL) | 136.8 | 101.0–184.7 | 89.6 | 1342.6 |
| α-Linolenic Acid (µg/mL) | 2.5 | 1.2–3.1 | 0.7 | 40.0 |
| Arachidonic Acid (µg/mL) | 162.6 | 125.8–181.7 | 89.4 | 251.6 |
| Eicosapentaenoic Acid (µg/mL) | 1.2 | 0.9–2.5 | 0.7 | 8.4 |
| Docosapentaenoic-n3 Acid (µg/mL) | 2.4 | 1.7–3.5 | 0.7 | 10.0 |
| Docosahexaenoic Acid (µg/mL) | 36.7 | 30.1–46.5 | 19.2 | 83.3 |
| Total Fatty Acids (µg/mL) | 1155.5 | 964.5–1341.8 | 721.2 | 4621.2 |
| Median | IQR | Min | Max | |
|---|---|---|---|---|
| Linoleic Acid (µg/mg) | 1.11 | 1.0–1.3 | 0.72 | 1.56 |
| α-Linolenic Acid (µg/mg) | 0.01 | 0.009–0.012 | 0.01 | 0.02 |
| Arachidonic Acid (µg/mg) | 2.09 | 1.8–2.3 | 1.28 | 2.51 |
| Eicosapentaenoic Acid (µg/mg) | 0.01 | 0.005–0.010 | 0.00 | 0.02 |
| Docosapentaenoic-n3 Acid (µg/mg) | 0.07 | 0.05–0.08 | 0.03 | 0.12 |
| Docosahexaenoic Acid (µg/mg) | 0.32 | 0.26–0.44 | 0.18 | 0.64 |
| Total Fatty Acids (µg/mg) | 9.97 | 8.63–10.72 | 6.82 | 12.46 |
| Median | IQR | Min | Max | |
|---|---|---|---|---|
| Linoleic Acid (g) | 9.42 | 8.2–14.2 | 1.8 | 18.7 |
| α-Linolenic Acid (g) | 1.19 | 0.9–1.4 | 0.2 | 1.9 |
| Arachidonic Acid (g) | 0.12 | 0.1–0.2 | 0.0 | 0.3 |
| Eicosapentaenoic Acid (g) | 0.01 | 0–0.05 | 0.0 | 0.4 |
| Docosapentaenoic-n3 Acid (g) | 0.02 | 0.01–0.02 | 0.0 | 0.1 |
| Docosahexaenoic Acid (g) | 0.06 | 0.03–0.12 | 0.0 | 0.4 |
| Total Omega-3 Acid (g) | 1.33 | 0.9–1.6 | 0.22 | 2.01 |
| n | Tertile 1 (Lowest) | Tertile 2 | Tertile 3 (Highest) | p-Value | |
|---|---|---|---|---|---|
| Linoleic Acid | 20 | 10.3% | 9.2% | 9.8% | 0.86 |
| α-linolenic Acid | 20 | 4.8% | 7.0% | 6.8% | 0.68 |
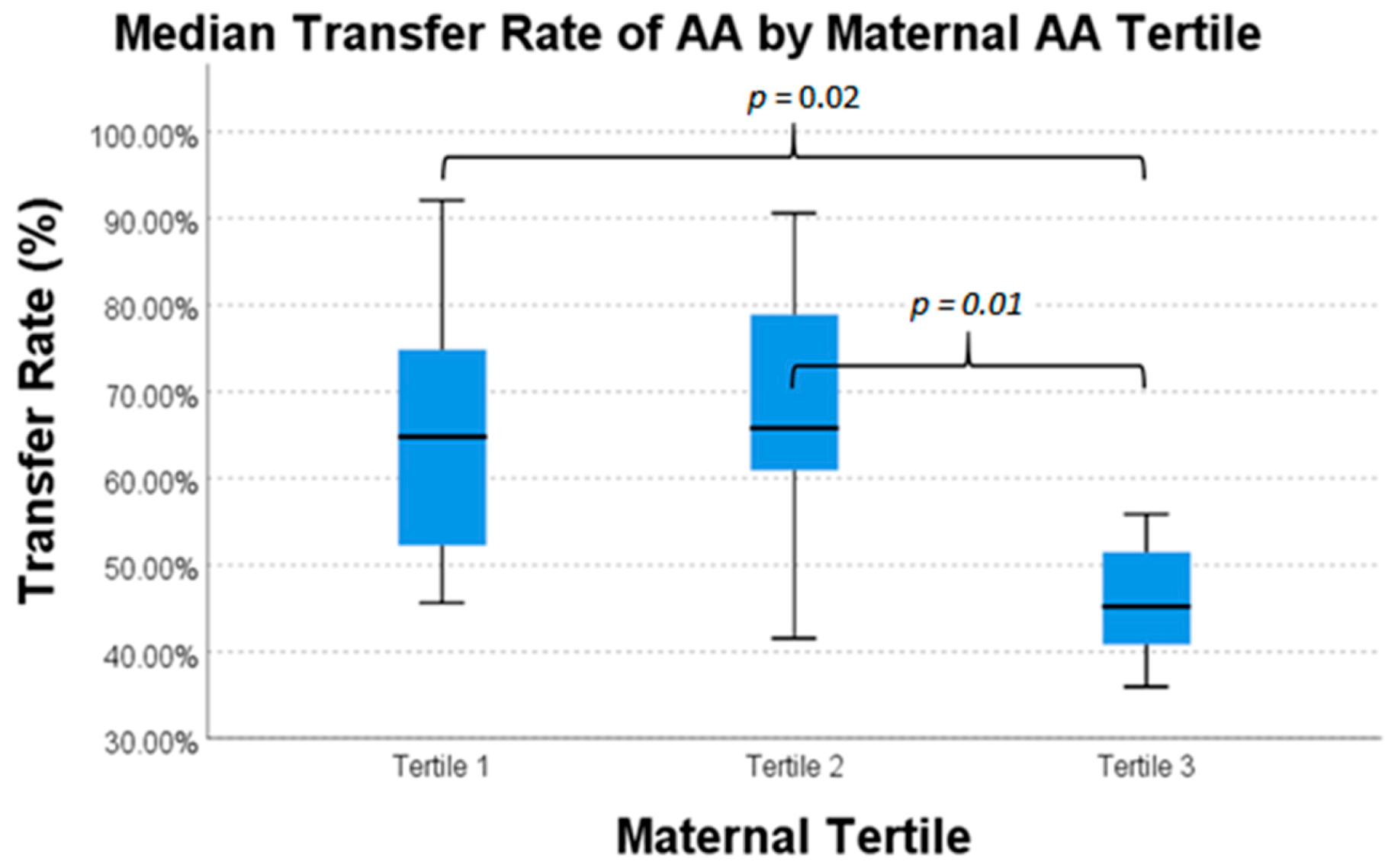
| Arachidonic Acid | 20 | 64.8% | 65.8% | 45.2% | 0.02 * |
| Docosapentaenoic Acid | 20 | 31.0% | 20.3% | 20.1% | 0.68 |
| Docosahexaenoic Acid | 20 | 57.8% | 52.4% | 45.9% | 0.60 |
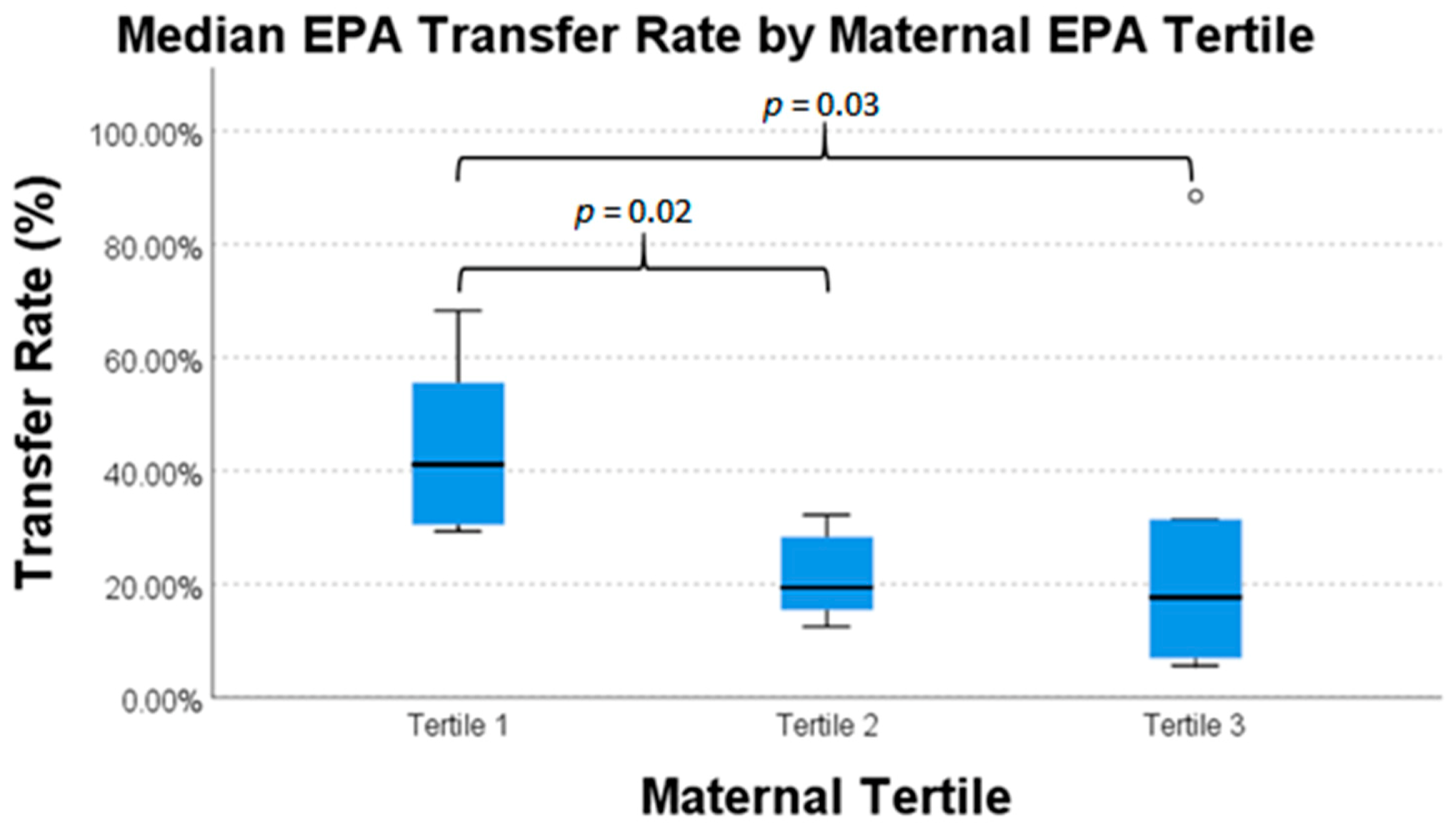
| Eicosapentaenoic Acid | 20 | 41.1% | 19.3% | 17.7% | 0.03 * |
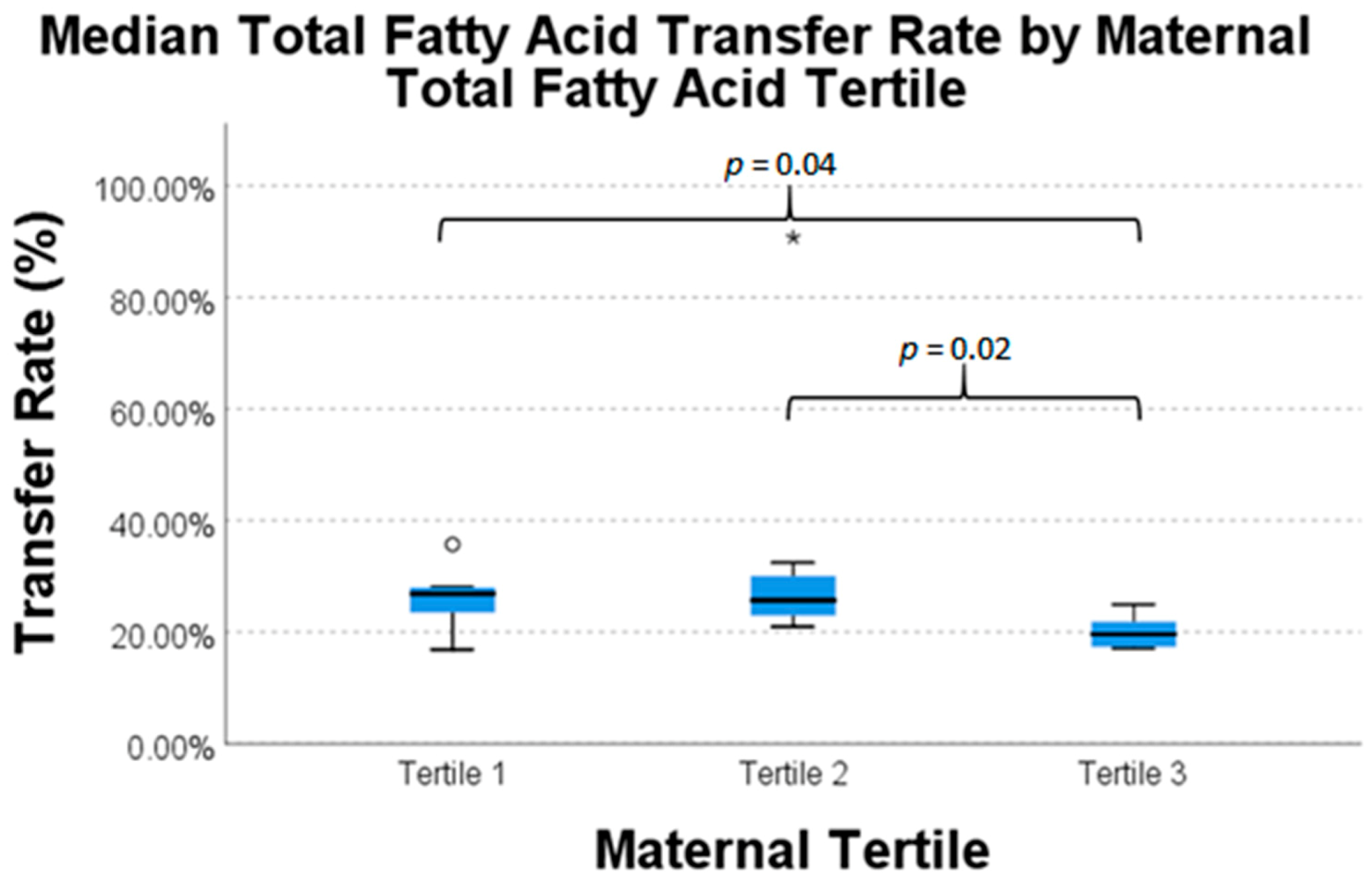
| Total Fatty Acids | 20 | 26.9% | 25.7% | 19.6% | 0.05 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woodard, V.; Thoene, M.; Van Ormer, M.; Thompson, M.; Hanson, C.; Natarajan, S.K.; Mukherjee, M.; Yuil-Valdes, A.; Nordgren, T.M.; Ulu, A.; et al. Intrauterine Transfer of Polyunsaturated Fatty Acids in Mother–Infant Dyads as Analyzed at Time of Delivery. Nutrients 2021, 13, 996. https://doi.org/10.3390/nu13030996
Woodard V, Thoene M, Van Ormer M, Thompson M, Hanson C, Natarajan SK, Mukherjee M, Yuil-Valdes A, Nordgren TM, Ulu A, et al. Intrauterine Transfer of Polyunsaturated Fatty Acids in Mother–Infant Dyads as Analyzed at Time of Delivery. Nutrients. 2021; 13(3):996. https://doi.org/10.3390/nu13030996
Chicago/Turabian StyleWoodard, Vanessa, Melissa Thoene, Matthew Van Ormer, Maranda Thompson, Corrine Hanson, Sathish Kumar Natarajan, Maheswari Mukherjee, Ana Yuil-Valdes, Tara M. Nordgren, Arzu Ulu, and et al. 2021. "Intrauterine Transfer of Polyunsaturated Fatty Acids in Mother–Infant Dyads as Analyzed at Time of Delivery" Nutrients 13, no. 3: 996. https://doi.org/10.3390/nu13030996