
Obeticholic Acid Inhibits Anxiety via Alleviating Gut Microbiota-Mediated Microglia Accumulation in the Brain of High-Fat High-Sugar Diet Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
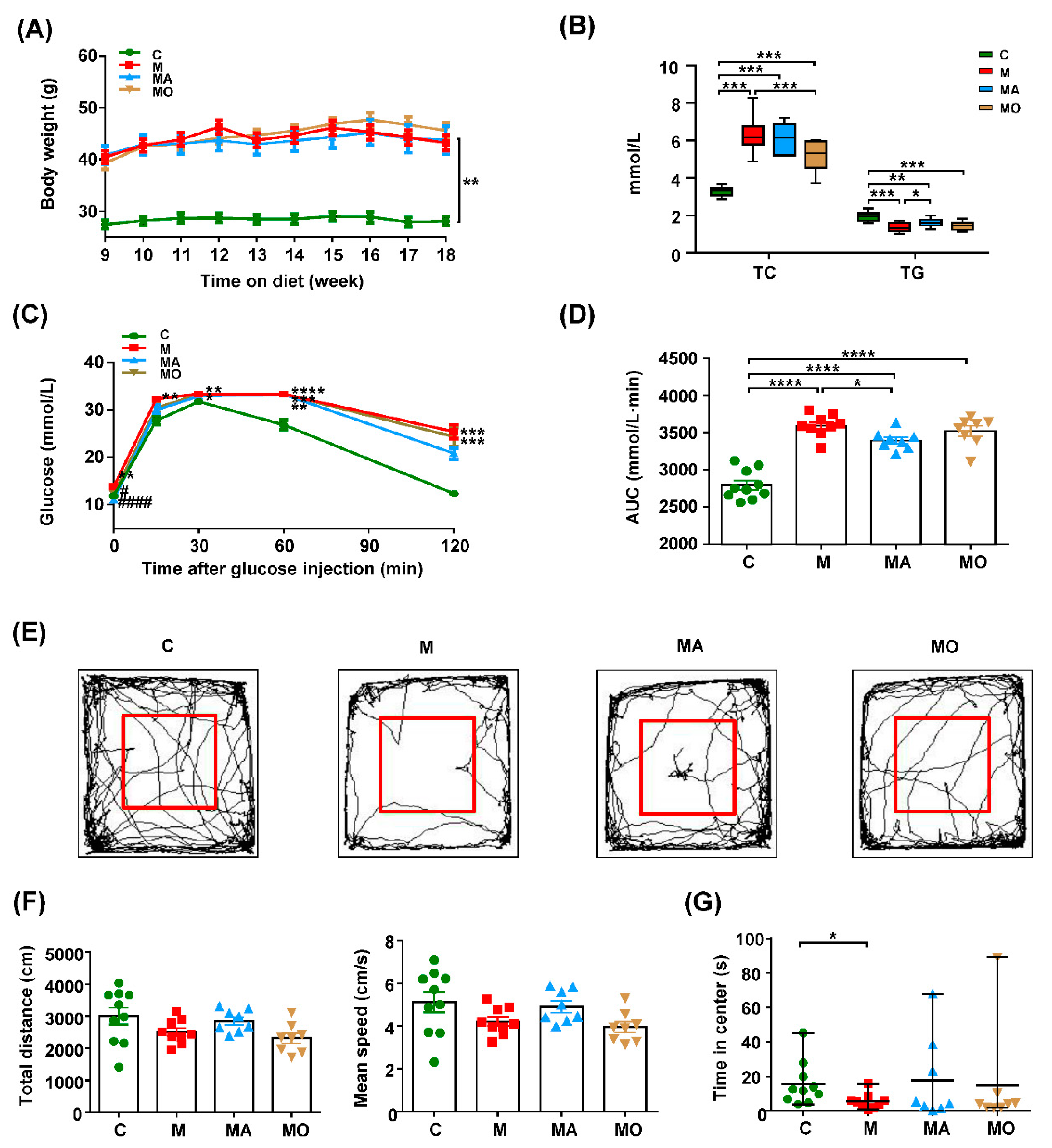
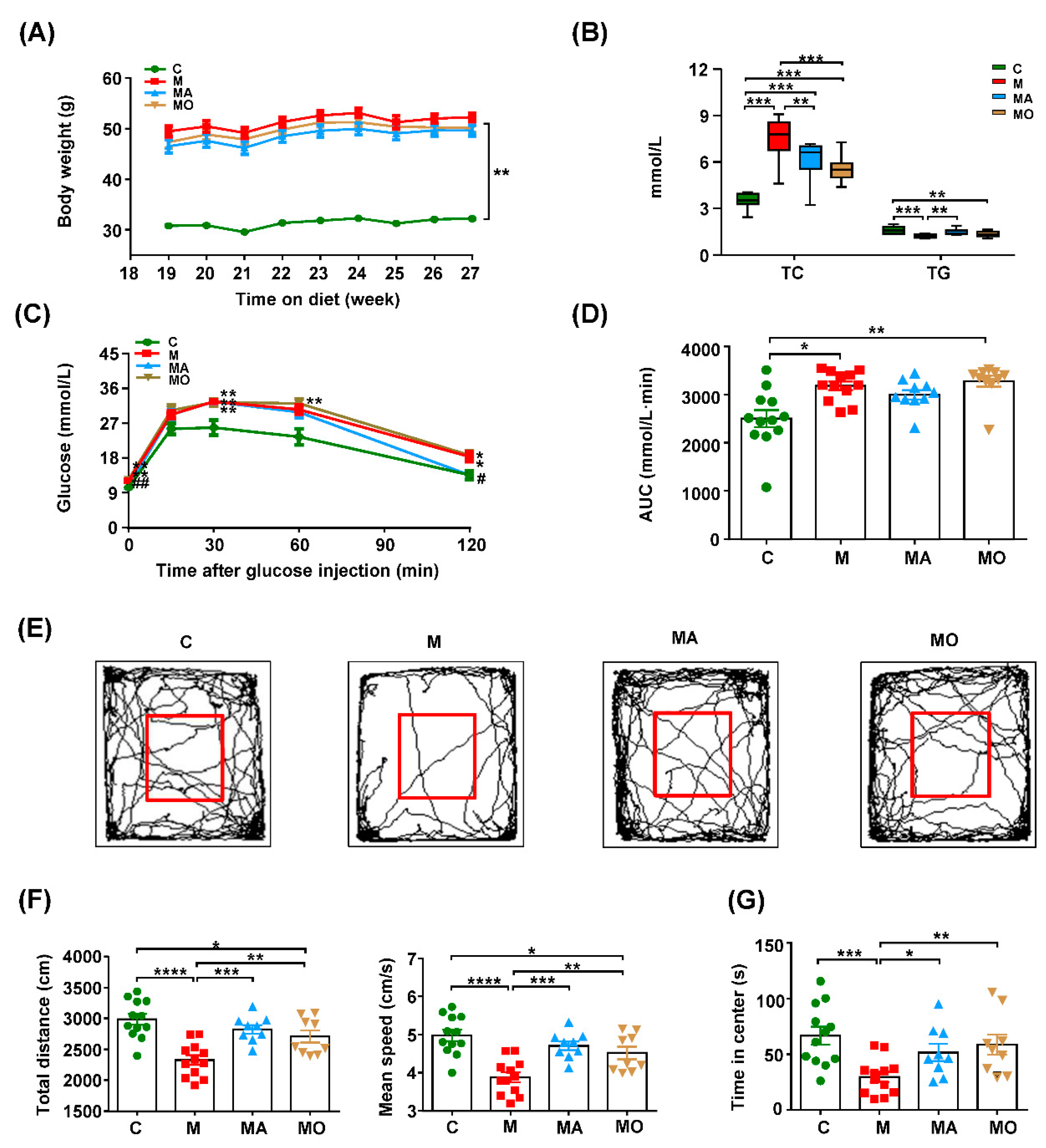
2.2. Measurement of Serum Biochemical Parameters
2.3. Glucose Tolerance Test
2.4. Histological Examination of the Liver
2.5. Morris Water Maze Test
2.6. Open Field Test
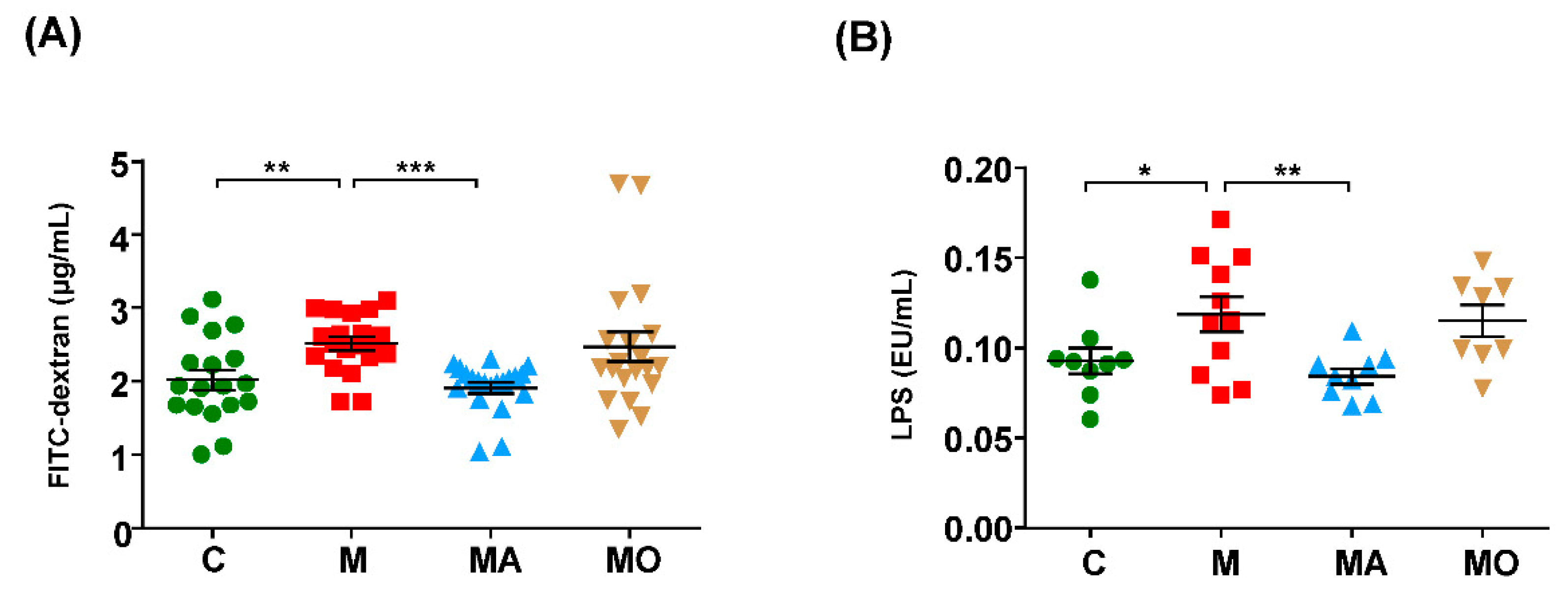
2.7. Intestinal Permeability In Vivo
2.8. Measurement of Serum Lipopolysaccharide Level
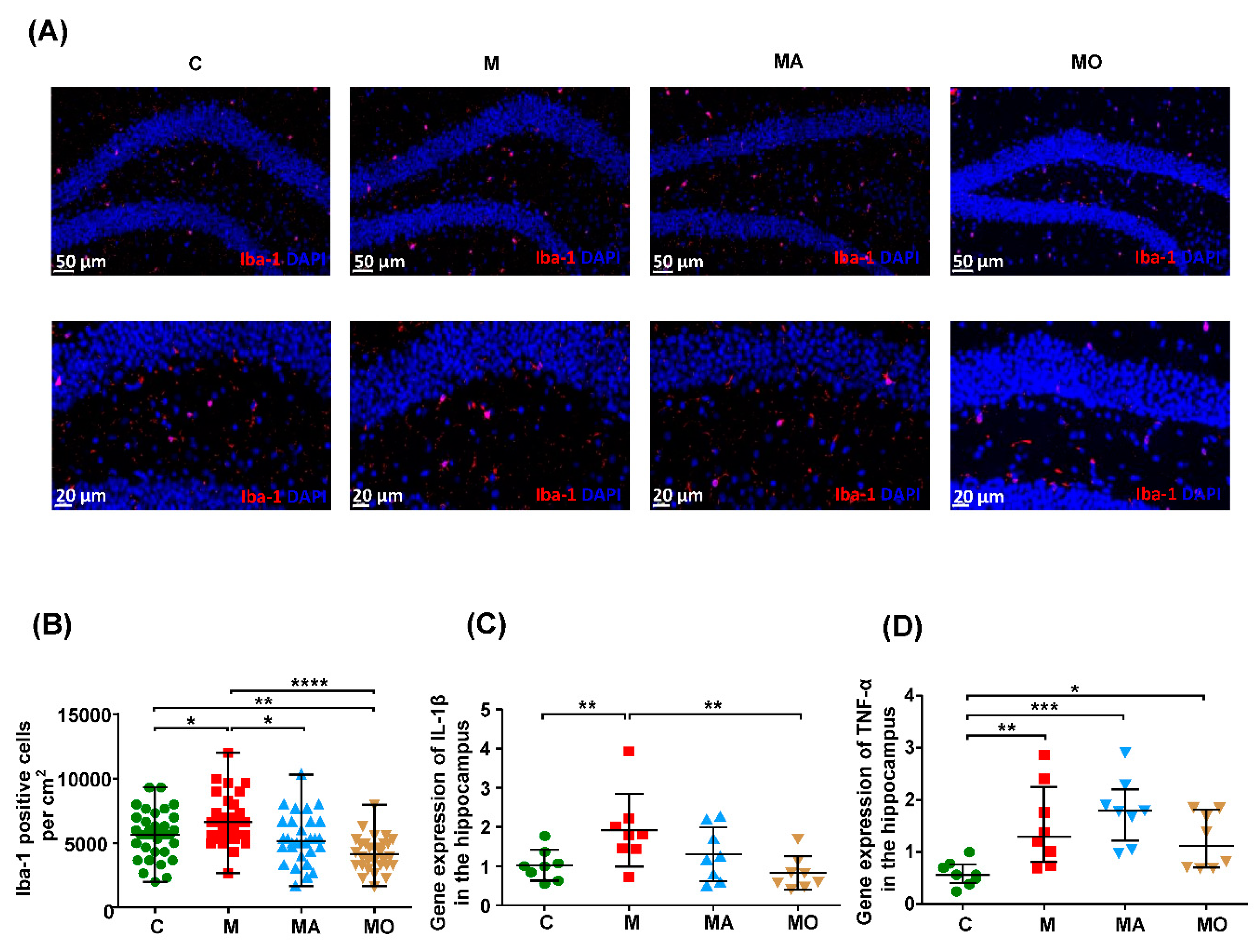
2.9. Immunofluorescence Staining of Microglial Cells in the Hippocampus
2.10. qPCR Analysis of Proinflammatory Cytokines in the Hippocampus
2.11. The Bacterial 16S rRNA Gene Sequencing
2.12. High-Throughput Untargeted Metabolomics Profiling
2.12.1. Chemical Reagents
2.12.2. Sample Preparation
2.12.3. Untargeted Metabolomics Profiling
2.12.4. Data Analysis
2.13. Targeted Profiling of Bile Acids
2.14. Statistical Analysis
3. Results
3.1. OCA Supplementation Inhibited Gut Microbiota-Mediated Anxiety in HFHS Diet-MDs Mice
3.2. OCA Supplementation Ameliorated Gut Microbiota-Mediated Microgliosis in HFHS Diet Mice
3.3. OCA Supplementation Improved Gut Microbiota-Mediated Leaky Gut and Endotoxemia in HFHS Diet Mice
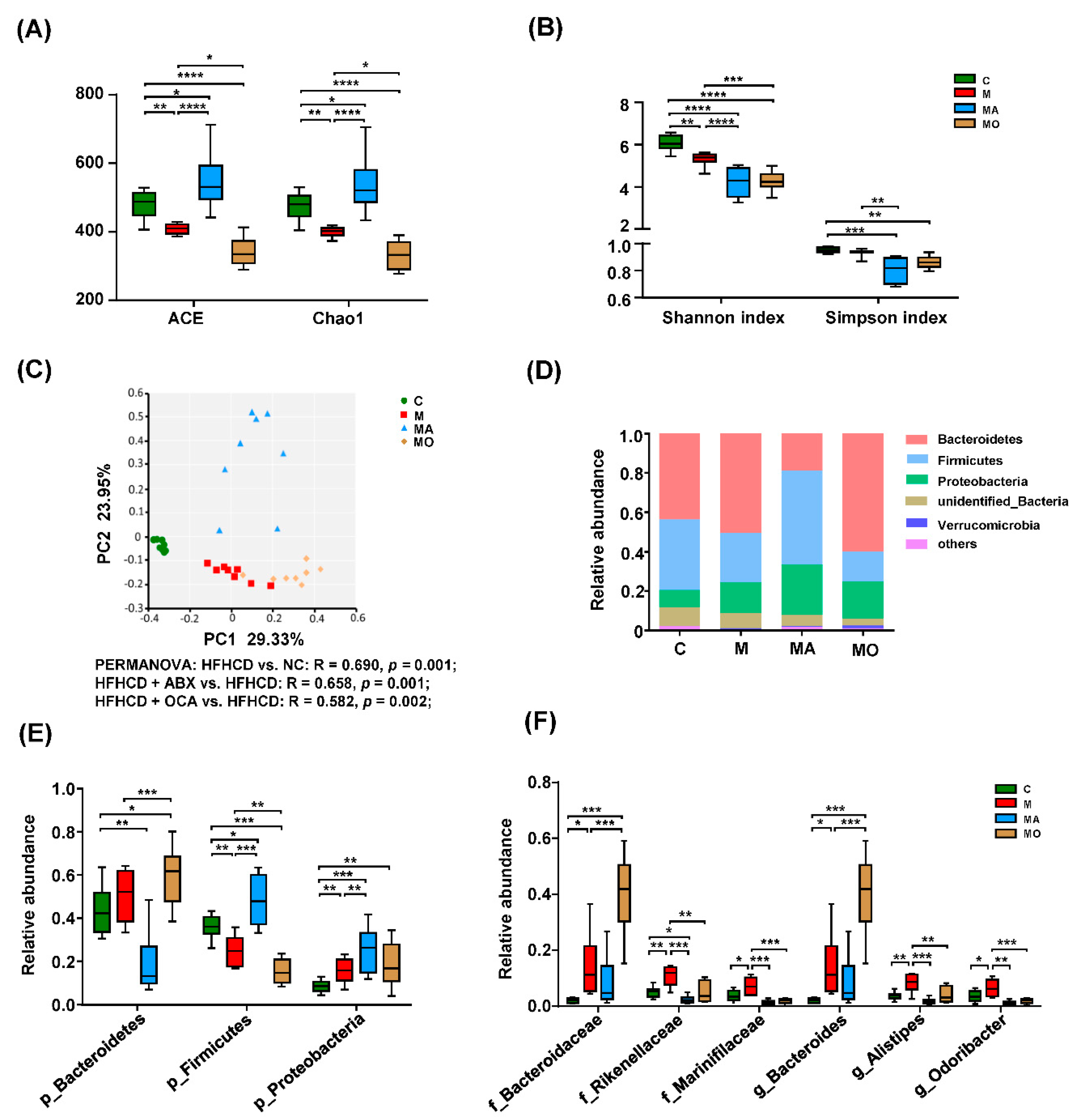
3.4. OCA Supplementation Modified Microbial Composition in HFHS Diet Mice
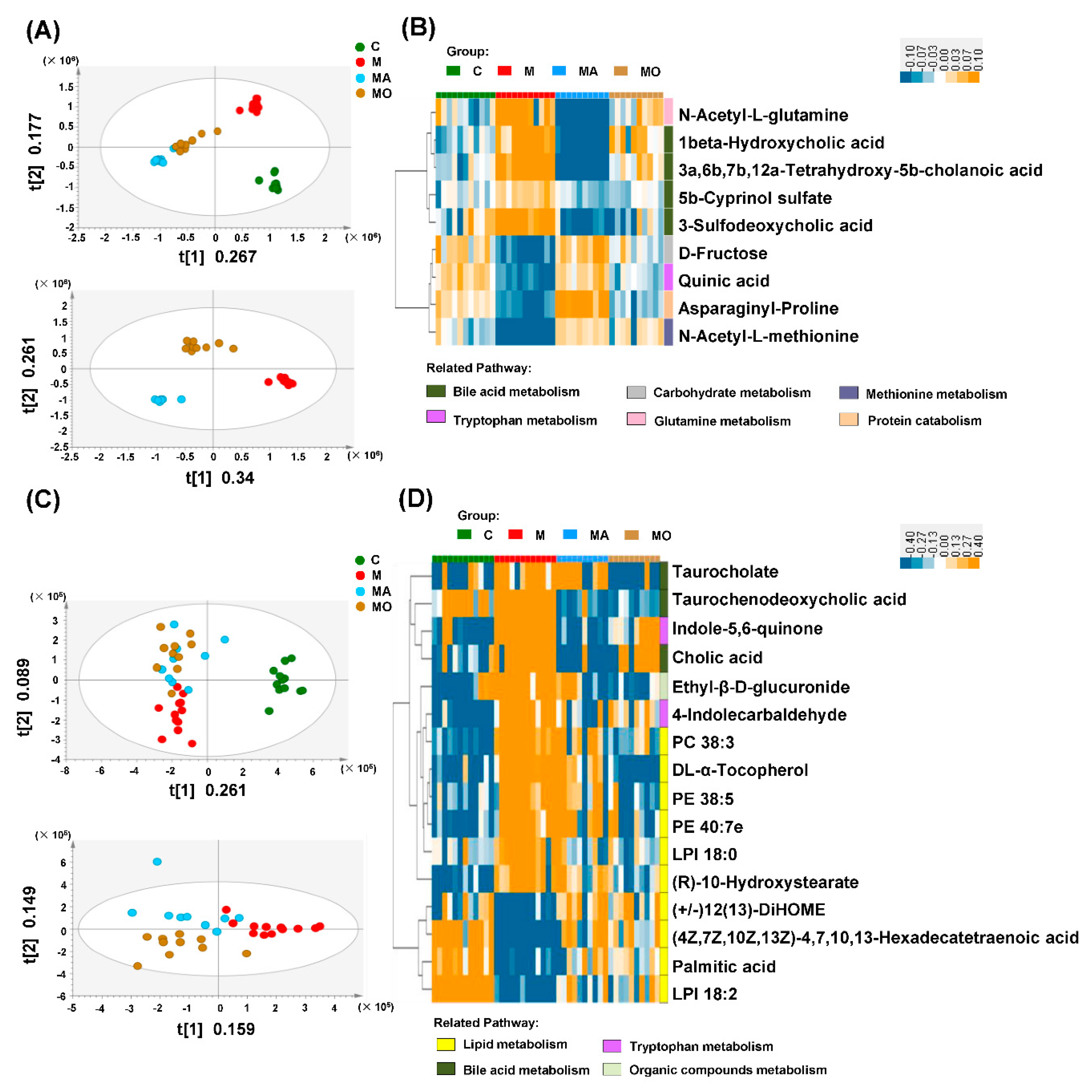
3.5. OCA Supplementation Improved Gut Microbiota-Mediated Metabolic Disturbance in HFHS Diet Mice
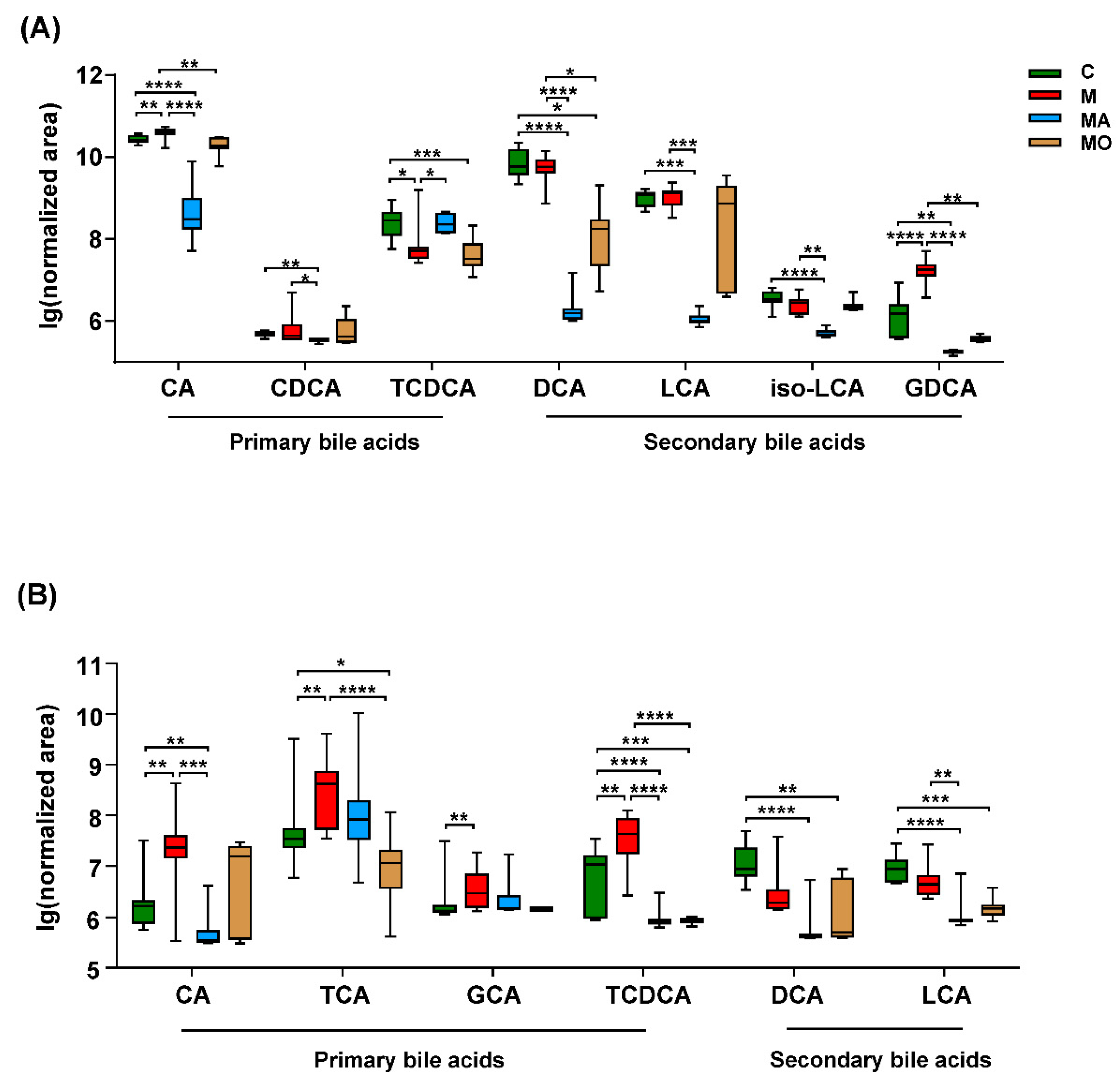
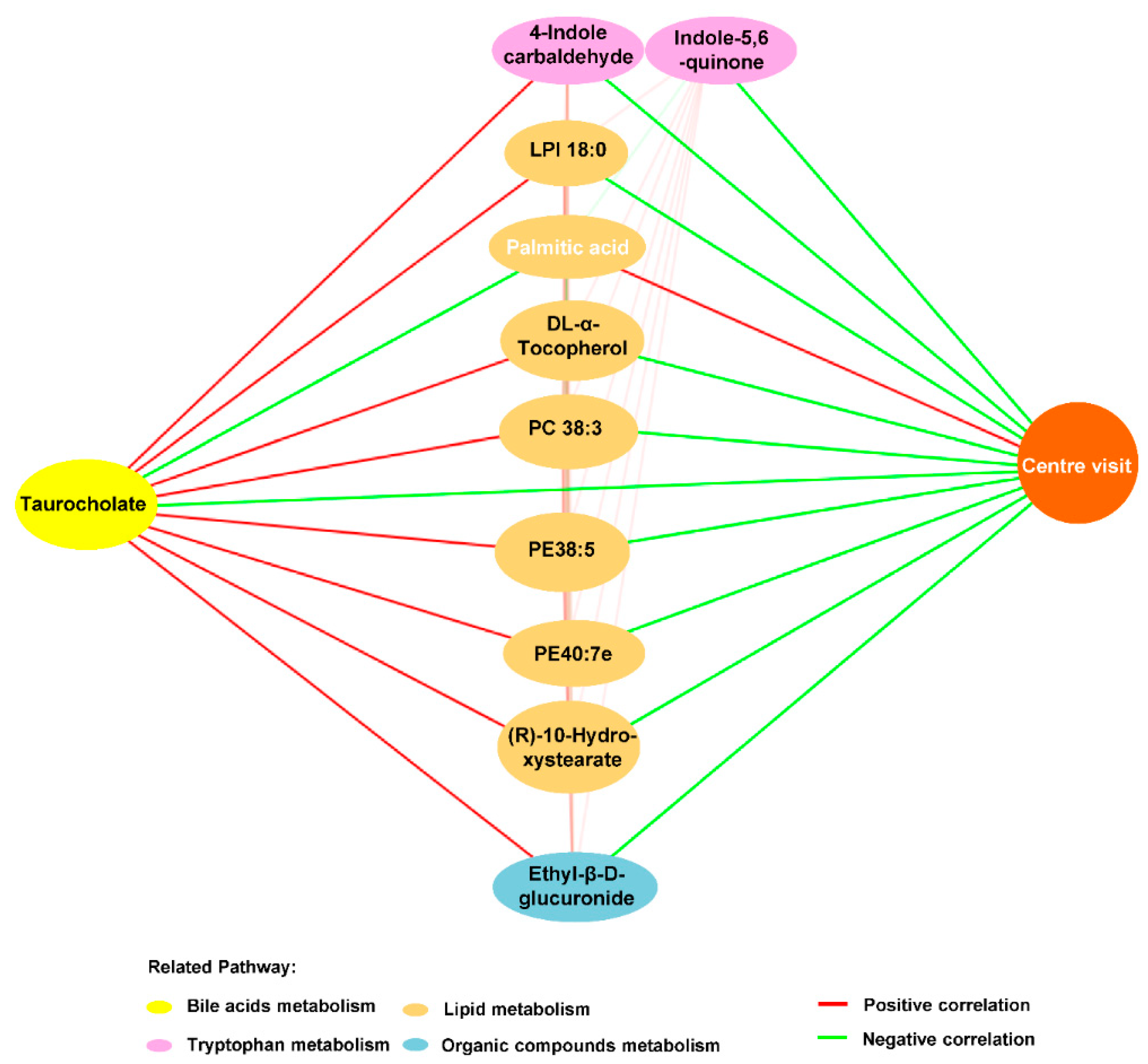
3.6. OCA-Reversed Bile Acid Linked Metabolic Disturbance to Anxiety in HFHS Diet Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Craske, M.G.; Stein, M.B. Anxiety. Lancet 2016, 388, 3048–3059. [Google Scholar] [CrossRef]
- Baxter, A.J.; Scott, K.M.; Vos, T.; Whiteford, H.A. Global Prevalence of Anxiety Disorders: A Systematic Review and Meta-regression. Psychol. Med. 2013, 43, 897–910. [Google Scholar] [CrossRef]
- Beaudreau, S.A.; O’Hara, R. Late-life Anxiety and Cognitive Impairment: A Review. Am. J. Geriatr. Psychiatry. 2008, 16, 790–803. [Google Scholar] [CrossRef]
- Fan, K.Q.; Li, Y.Y.; Wang, H.L.; Mao, X.T.; Guo, J.X.; Wang, F.; Huang, L.J.; Li, Y.N.; Ma, X.Y.; Gao, Z.J.; et al. Stress-Induced Metabolic Disorder in Peripheral CD4(+) T Cells Leads to Anxiety-like Behavior. Cell 2019, 179, 864–879.e819. [Google Scholar] [CrossRef]
- Labenz, C.; Huber, Y.; Michel, M.; Nagel, M.; Galle, P.R.; Kostev, K.; Schattenberg, J.M. Nonalcoholic Fatty Liver Disease Increases the Risk of Anxiety and Depression. Hepatol. Commun. 2020, 4, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Snoek, F.J.; Bremmer, M.A.; Hermanns, N. Constructs of Depression and Distress in Diabetes: Time for an Appraisal. Lancet Diabetes Endocrinol. 2015, 3, 450–460. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The Gut Microbiota at the Intersection of Diet and Human Health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, P.; Daita, K.; Joyce, A.; Mirshahi, F.; Santhekadur, P.K. The Presence and Severity of Nonalcoholic Steatohepatitis is Associated with Specific Changes in Circulating Bile Acids. Hepatology 2018, 67, 534–548. [Google Scholar] [CrossRef]
- Mahmoudian Dehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider-Paisley, A.; Moseley, M.A.; Thompson, J.W.; et al. Altered Bile Acid Profile Associates with Cognitive Impairment in Alzheimer’s Disease-An Emerging Role for Gut Microbiome. Alzheimers Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K.; et al. Altered Microbiomes Distinguish Alzheimer’s Disease from Amnestic Mild Cognitive Impairment and Health in a Chinese Cohort. Brain Behav. Immun. 2019, 80, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Han, W.; Zhan, G.; Li, S.; Xiang, S.; Zhu, B.; Jiang, X.; Yang, L.; Luo, A.; Hua, F.; et al. Abnormal Gut Microbiota Composition Contributes to Cognitive Dysfunction in Streptozotocin-induced Diabetic Mice. Aging 2019, 11, 3262–3279. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhong, Z.; Wang, B.; Xia, X.; Yao, W.; Huang, L.; Wang, Y.; Ding, W. Early-life High-fat Diet-induced Obesity Programs Hippocampal Development and Cognitive Functions via Regulation of Gut Commensal Akkermansia muciniphila. Neuropsychopharmacology 2019, 44, 2054–2064. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acids as Metabolic Regulators and Nutrient Sensors. Annu. Rev. Nutr. 2019, 39, 175–200. [Google Scholar] [CrossRef]
- Bhargava, P.; Smith, M.D.; Mische, L.; Harrington, E.; Fitzgerald, K.C.; Martin, K.; Kim, S.; Reyes, A.A.; Gonzalez-Cardona, J.; Volsko, C.; et al. Bile Acid Metabolism is Altered in Multiple Sclerosis and Supplementation Ameliorates Neuroinflammation. J. Clin. Invest. 2020, 130, 3467–3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlström, A.; Sayin, S.I.; Marschall, H.U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Eslimi, D.; Oryan, S.; Nasehi, M.; Zarrindast, M.R. Effects of Opioidergic Systems upon Anxiolytic-like Behaviors Induced in Cholestatic Rats. Eur. J. Pharmacol. 2011, 670, 180–185. [Google Scholar] [CrossRef]
- Xie, G.; Wang, X.; Jiang, R.; Zhao, A.; Yan, J.; Zheng, X.; Huang, F.; Liu, X.; Panee, J.; Rajani, C.; et al. Dysregulated Bile Acid Signaling Contributes to the Neurological Impairment in Murine Models of Acute and Chronic Liver Failure. EBioMedicine 2018, 37, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Mudaliar, S.; Henry, R.R.; Sanyal, A.J.; Morrow, L.; Marschall, H.U.; Kipnes, M.; Adorini, L.; Sciacca, C.I.; Clopton, P.; Castelloe, E.; et al. Efficacy and Safety of the Farnesoid X Receptor Agonist Obeticholic Acid in Patients with Type 2 Diabetes and Nonalcoholic Fatty Liver Disease. Gastroenterology 2013, 145, 574–582.e571. [Google Scholar] [CrossRef]
- Li, F.; Hao, X.; Chen, Y.; Bai, L.; Gao, X.; Lian, Z.; Wei, H.; Sun, R.; Tian, Z. The Microbiota Maintain Homeostasis of Liver-resident γδT-17 Cells in a Lipid Antigen/CD1d-dependent Manner. Nat. Commun. 2017, 7, 13839. [Google Scholar] [CrossRef]
- Lv, B.; Ma, L.; Tang, W.; Huang, P.; Yang, B.; Wang, L.; Chen, S.; Gao, Q.; Zhang, S.; Xia, J. FXR Acts as a Metastasis Suppressor in Intrahepatic Cholangiocarcinoma by Inhibiting IL-6-Induced Epithelial-Mesenchymal Transition. Cell. Physiol. Biochem. 2018, 48, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.R.; Françozo, M.C.; de Oliveira, G.G.; Ignacio, A.; Castoldi, A.; Zamboni, D.S.; Ramos, S.G.; Câmara, N.O.; de Zoete, M.R.; Palm, N.W.; et al. Gut Microbiota Translocation to the Pancreatic Lymph Nodes Triggers NOD2 Activation and Contributes to T1D Onset. J. Exp. Med. 2016, 213, 1223–1239. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Jiang, X.; Ling, Q.; Wu, L.; Wu, P.; Tang, R.; Xu, X.; Yang, M.; Zhang, L.; Zhu, W.; et al. Antibiotics-mediated Intestinal Microbiome Perturbation Aggravates Tacrolimus-induced Glucose Disorders in Mice. Front. Med. 2019, 13, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Lv, L.; Fang, D.; Wu, W.; Hu, C.; Xu, L.; Chen, Y.; Guo, J.; Hu, X.; Li, A.; et al. Administration of Lactobacillus Salivarius LI01 or Pediococcus Pentosaceus LI05 Prevents CCl(4)-induced Liver Cirrhosis by Protecting the Intestinal Barrier in Rats. Sci. Rep. 2017, 7, 6927. [Google Scholar] [CrossRef] [Green Version]
- Boehme, M.; van de Wouw, M. Mid-life Microbiota Crises: Middle Age is Associated with Pervasive Neuroimmune Alterations That are Reversed by Targeting the Gut Microbiome. Mol. Psychiatry 2020, 25, 2567–2583. [Google Scholar] [CrossRef]
- Xiang, L.; Cao, X.L.; Xing, T.Y.; Mori, D.; Tang, R.Q.; Li, J.; Gao, L.J.; Qi, J.H. Mixture of Peanut Skin Extract and Fish Oil Improves Memory in Mice via Modulation of Anti-Oxidative Stress and Regulation of BDNF/ERK/CREB Signaling Pathways. Nutrients 2016, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-induced Inflammation in High-fat Diet-induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.Y.; Gillilland, M., 3rd; Wu, X.; Leelasinjaroen, P.; Zhang, G.; Zhou, H.; Ye, B.; Lu, Y.; Owyang, C. FODMAP Diet Modulates Visceral Nociception by Lipopolysaccharide-mediated Intestinal Inflammation and Barrier Dysfunction. J. Clin. Invest 2018, 128, 267–280. [Google Scholar] [CrossRef]
- Gomes, A.L.; Teijeiro, A.; Burén, S.; Tummala, K.S.; Yilmaz, M.; Waisman, A.; Theurillat, J.P.; Perna, C.; Djouder, N. Metabolic Inflammation-Associated IL-17A Causes Non-alcoholic Steatohepatitis and Hepatocellular Carcinoma. Cancer Cell 2016, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Hewing, B.; Parathath, S.; Mai, C.K.; Fiel, M.I.; Guo, L.; Fisher, E.A. Rapid Regression of Atherosclerosis with MTP Inhibitor Treatment. Atherosclerosis 2013, 227, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Syn, W.K.; Agboola, K.M.; Swiderska, M.; Michelotti, G.A.; Liaskou, E.; Pang, H.; Xie, G.; Philips, G.; Chan, I.S.; Karaca, G.F.; et al. NKT-associated Hedgehog and Osteopontin Drive Fibrogenesis in Non-alcoholic Fatty Liver Disease. Gut 2012, 61, 1323–1329. [Google Scholar] [CrossRef]
- Shvachiy, L.; Geraldes, V.; Amaro-Leal, Â.; Rocha, I. Intermittent Low-level Lead Exposure Provokes Anxiety, Hypertension, Autonomic Dysfunction and Neuroinflammation. Neurotoxicology 2018, 69, 307–319. [Google Scholar] [CrossRef]
- Ye, J.; Lv, L.; Wu, W.; Li, Y.; Shi, D.; Fang, D.; Guo, F.; Jiang, H.; Yan, R.; Ye, W.; et al. Butyrate Protects Mice Against Methionine-Choline-Deficient Diet-Induced Non-alcoholic Steatohepatitis by Improving Gut Barrier Function, Attenuating Inflammation and Reducing Endotoxin Levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S rRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lv, L.; Shi, D.; Ye, J.; Fang, D.; Guo, F.; Li, Y.; He, X.; Li, L. Protective Effect of Akkermansia Muciniphila against Immune-Mediated Liver Injury in a Mouse Model. Front. Microbiol. 2017, 8, 1804. [Google Scholar] [CrossRef]
- Cao, H.; Huang, H.; Xu, W.; Chen, D.; Yu, J.; Li, J.; Li, L. Fecal Metabolome Profiling of Liver Cirrhosis and Hepatocellular Carcinoma Patients by Ultra Performance Liquid Chromatography-mass Spectrometry. Anal. Chim. Acta 2011, 691, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Tan, Y.; Yin, P.; Ye, G.; Gao, P.; Lu, X.; Wang, H.; Xu, G. Metabolic Characterization of Hepatocellular Carcinoma Using Nontargeted Tissue Metabolomics. Cancer Res. 2013, 73, 4992–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonato, M.; Fochi, I.; Vedovelli, L.; Giambelluca, S.; Carollo, C.; Padalino, M.; Carnielli, V.P.; Cogo, P. Urinary Metabolomics Reveals Kynurenine Pathway Perturbation in Newborns with Transposition of Great Arteries after Surgical Repair. Metabolomics 2019, 15, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Chen, D.; Chen, Y.; Hu, Z.; Cao, M.; Xie, Q.; Chen, Y.; Xu, J.; Zheng, S.; Li, L. Metabonomic Profiles Discriminate Hepatocellular Carcinoma from Liver Cirrhosis by Ultraperformance Liquid Chromatography-mass Spectrometry. J. Proteome. Res. 2012, 11, 1217–1227. [Google Scholar] [CrossRef]
- Hoon, M.J.L.d.; Imoto, S.; Nolan, J.; Miyano, S. Open Source Clustering Software. Bioinformatics 2004, 20, 1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Uña, M.; Varela-Rey, M.; Mestre, D.; Fernández-Ares, L.; Fresnedo, O.; Fernandez-Ramos, D.; Gutiérrez-de Juan, V.; Martin-Guerrero, I.; García-Orad, A.; Luka, Z.; et al. S-Adenosylmethionine Increases Circulating Very-low Density Lipoprotein Clearance in Non-alcoholic Fatty Liver Disease. J. Hepatol. 2015, 62, 673–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Nozaki, Y.; Wada, K.; Yoneda, M.; Fujimoto, Y.; Fujitake, M.; Endo, H.; Takahashi, H.; Inamori, M.; Kobayashi, N.; et al. Dysfunctional Very-low-density Lipoprotein Synthesis and Release is a Key Factor in Nonalcoholic Steatohepatitis Pathogenesis. Hepatology 2009, 50, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Studer, V.; Motta, C.; Polidoro, S.; Perugini, J.; Macchiarulo, G.; Giovannetti, A.M.; Pareja-Gutierrez, L.; Calò, A.; Colonna, I.; et al. Neuroinflammation Drives Anxiety and Depression in Relapsing-remitting Multiple Sclerosis. Neurology 2017, 89, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Alsiö, J.; Hulting, A.L.; Schiöth, H.B. Withdrawal from Free-choice High-fat High-sugar Diet Induces Craving Only in Obesity-prone Animals. Psychopharmacology 2009, 204, 431–443. [Google Scholar] [CrossRef]
- Baker, K.D.; Reichelt, A.C. Impaired Fear Extinction Retention and Increased Anxiety-like Behaviours Induced by Limited Daily Access to a High-fat/High-sugar Diet in Male Rats: Implications for Diet-induced Prefrontal Cortex Dysregulation. Neurobiol. Learn. Mem. 2016, 136, 127–138. [Google Scholar] [CrossRef]
- Zemdegs, J.; Quesseveur, G.; Jarriault, D.; Pénicaud, L.; Fioramonti, X.; Guiard, B.P. High-fat Diet-induced Metabolic Disorders Impairs 5-HT Function and Anxiety-like Behavior in Mice. Br. J. Pharmacol. 2016, 173, 2095–2110. [Google Scholar] [CrossRef]
- Rooney, S.; Sah, A.; Unger, M.S.; Kharitonova, M.; Sartori, S.B.; Schwarzer, C.; Aigner, L.; Kettenmann, H.; Wolf, S.A.; Singewald, N. Neuroinflammatory Alterations in Trait Anxiety: Modulatory Effects of Minocycline. Transl. Psychiatry 2020, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Haroon, E.; Daguanno, A.W.; Woolwine, B.J.; Goldsmith, D.R.; Baer, W.M.; Wommack, E.C.; Felger, J.C.; Miller, A.H. Antidepressant Treatment Resistance is Associated with Increased Inflammatory Markers in Patients with Major Depressive Disorder. Psychoneuroendocrinology 2018, 95, 43–49. [Google Scholar] [CrossRef]
- Ho, P.P.; Steinman, L. Obeticholic Acid, a Synthetic Bile Acid Agonist of the Farnesoid X Receptor, Attenuates Experimental Autoimmune Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2016, 113, 1600–1605. [Google Scholar] [CrossRef] [Green Version]
- McMillin, M.; Frampton, G.; Grant, S.; Khan, S.; Diocares, J.; Petrescu, A.; Wyatt, A.; Kain, J.; Jefferson, B.; DeMorrow, S. Bile Acid-Mediated Sphingosine-1-Phosphate Receptor 2 Signaling Promotes Neuroinflammation during Hepatic Encephalopathy in Mice. Front. Cell. Neurosci. 2017, 11, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, M.; McMillin, M.; Galindo, C.; Frampton, G.; Pae, H.Y.; DeMorrow, S. Bile Acids Permeabilize the Blood Brain Barrier after Bile Duct Ligation in Rats via Rac1-dependent Mechanisms. Dig. Liver Dis. 2014, 46, 527–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro-Cardoso, V.F.; Corlianò, M.; Singaraja, R.R. Bile Acids: A Communication Channel in the Gut-Brain Axis. NeuroMolecular Med. 2021, 23, 99–117. [Google Scholar] [CrossRef]
- Friedman, E.S.; Li, Y.; Shen, T.D.; Jiang, J.; Chau, L.; Adorini, L.; Babakhani, F.; Edwards, J.; Shapiro, D.; Zhao, C.; et al. FXR-Dependent Modulation of the Human Small Intestinal Microbiome by the Bile Acid Derivative Obeticholic Acid. Gastroenterology 2018, 155, 1741–1752.e1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.H.; Tu, J.L.; Li, X.H.; Hua, Q.; Liu, W.Z.; Liu, Y.; Pan, B.X.; Hu, P.; Zhang, W.H. Neuroinflammation Induces Anxiety- and Depressive-like Behavior by Modulating Neuronal Plasticity in the Basolateral Amygdala. Brain Behav. Immun. 2020, 91, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Izvolskaia, M.; Sharova, V. Prenatal Programming of Neuroendocrine System Development by Lipopolysaccharide: Long-Term Effects. Int. J. Mol. Sci. 2018, 19, 3695. [Google Scholar] [CrossRef] [Green Version]
- Lasselin, J.; Schedlowski, M.; Karshikoff, B.; Engler, H.; Lekander, M.; Konsman, J.P. Comparison of Bacterial Lipopolysaccharide-induced Sickness Behavior in Rodents and Humans: Relevance for Symptoms of Anxiety and Depression. Neurosci. Biobehav. Rev. 2020, 115, 15–24. [Google Scholar] [CrossRef]
- Shishkina, G.T.; Bannova, A.V.; Komysheva, N.P.; Dygalo, N.N. Anxiogenic-like Effect of Chronic Lipopolysaccharide is Associated with Increased Expression of Matrix Metalloproteinase 9 in the Rat Amygdala. Stress 2020, 23, 708–714. [Google Scholar] [CrossRef]
- Järbrink-Sehgal, E.; Andreasson, A. The Gut Microbiota and Mental Health in Adults. Curr. Opin. Neurobiol. 2020, 62, 102–114. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhao, J.; Li, C.; Zhang, M.; Wei, L.; Zhang, X.; Kurskaya, O.; Bi, H.; Gao, T. Effect of Combined Chronic Predictable and Unpredictable Stress on Depression-like Symptoms in Mice. Ann. Transl. Med. 2020, 8, 942. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, G. Alistipes: The Influence of a Commensal on Anxiety and Depression. Catalyst 2019, 3, 2–10. [Google Scholar]
- Haleem, D.J.; Mahmood, K. Brain Serotonin in High-fat Diet-induced Weight Gain, Anxiety and Spatial Memory in Rats. Nutr. Neuroence 2019, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; He, T.; Johnston, L.J.; Ma, X. Host-microbiome Interactions: The Aryl Hydrocarbon Receptor as a Critical Node in Tryptophan Metabolites to Brain Signaling. Gut Microbes 2020, 11, 1203–1219. [Google Scholar] [CrossRef]
- Lee, Y.H.; Lin, C.H.; Hsu, P.C.; Sun, Y.Y.; Huang, Y.J.; Zhuo, J.H.; Wang, C.Y.; Gan, Y.L.; Hung, C.C.; Kuan, C.Y.; et al. Aryl Hydrocarbon Receptor Mediates Both Proinflammatory and Anti-inflammatory Effects in Lipopolysaccharide-activated Microglia. Glia 2015, 63, 1138–1154. [Google Scholar] [CrossRef] [PubMed]
- Ramos-García, N.A.; Orozco-Ibarra, M. Aryl Hydrocarbon Receptor in Post-Mortem Hippocampus and in Serum from Young, Elder, and Alzheimer’s Patients. Int. J. Mol. Sci. 2020, 21, 1983. [Google Scholar] [CrossRef] [Green Version]
- Zang, X.; Zheng, X.; Hou, Y.; Hu, M.; Wang, H.; Bao, X.; Zhou, F.; Wang, G.; Hao, H. Regulation of Proinflammatory Monocyte Activation by the Kynurenine-AhR axis Underlies Immunometabolic Control of Depressive Behavior in Mice. FASEB J. 2018, 32, 1944–1956. [Google Scholar] [CrossRef] [Green Version]
- de Araujo, L.S.; Pessler, K.; Sühs, K.W.; Novoselova, N.; Klawonn, F.; Kuhn, M.; Kaever, V.; Müller-Vahl, K.; Trebst, C.; Skripuletz, T.; et al. Phosphatidylcholine PC ae C44:6 in Cerebrospinal Fluid is a Sensitive Biomarker for Bacterial Meningitis. J. Transl. Med. 2020, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Berkecz, R.; Körmöczi, T.; Tömösi, F. Plasma Phospholipid Profiling of a Mouse Model of Anxiety Disorder by Hydrophilic Interaction Liquid Chromatography Coupled to High-resolution Mass Spectrometry. Biomed. Chromatogr. BMC 2018, 32, e4202. [Google Scholar] [CrossRef] [Green Version]
- Demirkan, A.; Isaacs, A.; Ugocsai, P.; Liebisch, G.; Struchalin, M.; Rudan, I.; Wilson, J.F.; Pramstaller, P.P.; Gyllensten, U.; Campbell, H.; et al. Plasma Phosphatidylcholine and Sphingomyelin Concentrations are Associated with Depression and Anxiety Symptoms in a Dutch Family-based Lipidomics Study. J. Psychiatr. Res. 2013, 47, 357–362. [Google Scholar] [CrossRef]
- Liśkiewicz, A.; Przybyła, M.; Wojakowska, A.; Marczak, Ł.; Bogus, K.; Nowacka-Chmielewska, M.; Liśkiewicz, D.; Małecki, A.; Barski, J.; Lewin-Kowalik, J.; et al. Physical Activity Reduces Anxiety and Regulates Brain Fatty Acid Synthesis. Mol. Brain 2020, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Blandina, B.M.; Jonathan, C.E.; Gabriel, G.-R.; Rodriguez-Landa, F.J.; Contreras, C.M. A Fatty Acids Mixture Reduces Anxiety-Like Behaviors in Infant Rats Mediated by GABAA Receptors. BioMed Res. Int. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Han, Y.; Zheng, Z.; Zhu, S.; Chen, J.; Yao, Y.; Yue, S.; Teufel, A.; Weng, H.; Li, L.; et al. Obeticholic Acid Inhibits Anxiety via Alleviating Gut Microbiota-Mediated Microglia Accumulation in the Brain of High-Fat High-Sugar Diet Mice. Nutrients 2021, 13, 940. https://doi.org/10.3390/nu13030940
Wu L, Han Y, Zheng Z, Zhu S, Chen J, Yao Y, Yue S, Teufel A, Weng H, Li L, et al. Obeticholic Acid Inhibits Anxiety via Alleviating Gut Microbiota-Mediated Microglia Accumulation in the Brain of High-Fat High-Sugar Diet Mice. Nutrients. 2021; 13(3):940. https://doi.org/10.3390/nu13030940
Chicago/Turabian StyleWu, Li, Yuqiu Han, Zhipeng Zheng, Shuai Zhu, Jun Chen, Yuanyuan Yao, Siqing Yue, Andreas Teufel, Honglei Weng, Lanjuan Li, and et al. 2021. "Obeticholic Acid Inhibits Anxiety via Alleviating Gut Microbiota-Mediated Microglia Accumulation in the Brain of High-Fat High-Sugar Diet Mice" Nutrients 13, no. 3: 940. https://doi.org/10.3390/nu13030940