
The Nutraceutical N-Palmitoylethanolamide (PEA) Reveals Widespread Molecular Effects Unmasking New Therapeutic Targets in Murine Varicocele

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Animals and Surgical Procedures
2.3. Materials
2.4. Experimental Groups
2.5. Histological and Morphometric Evaluation
2.6. Immunohistochemical Analysis
2.7. Western Blot Analysis
2.8. Statistical Evaluation
3. Results
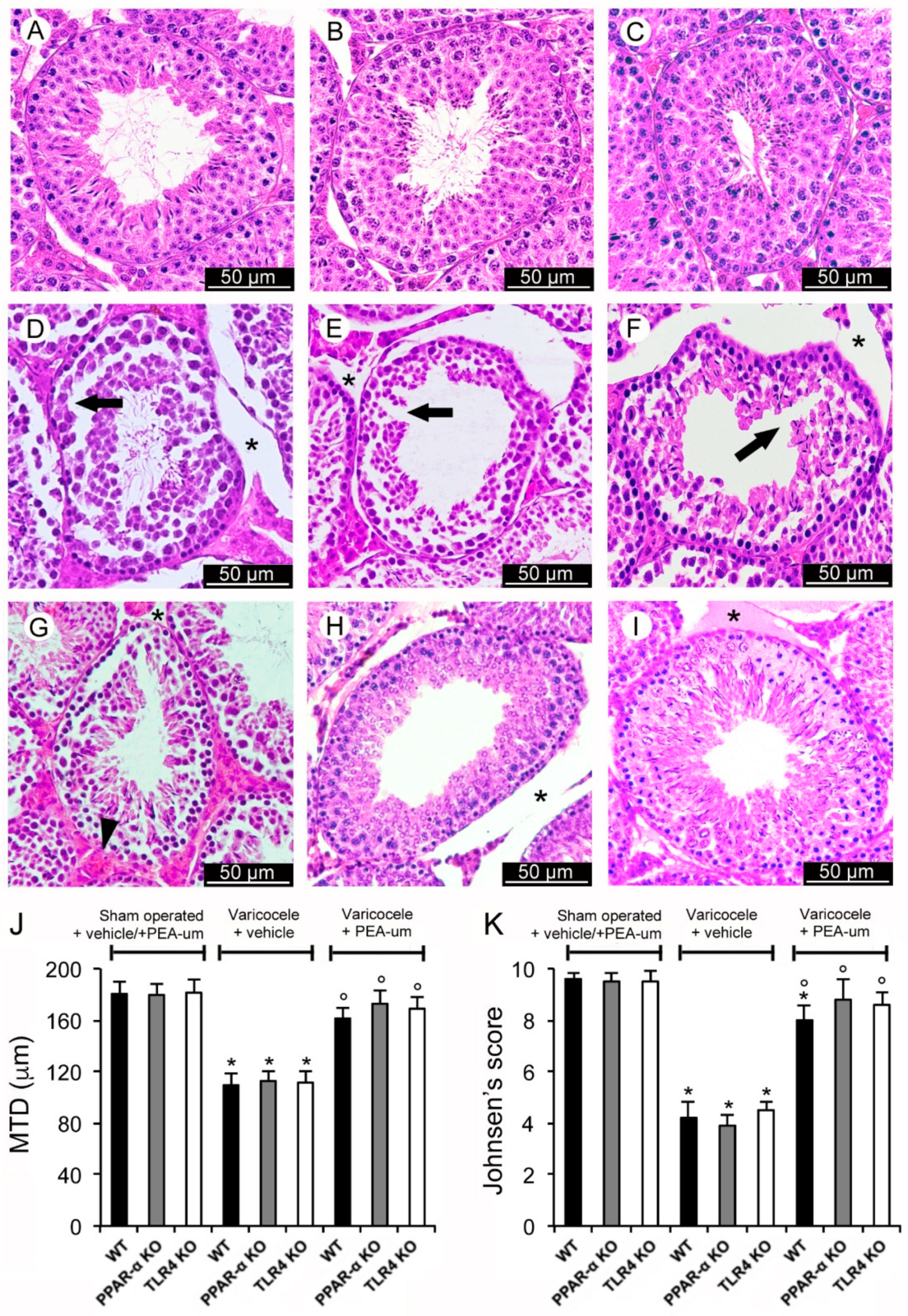
3.1. Histopathological Evaluation
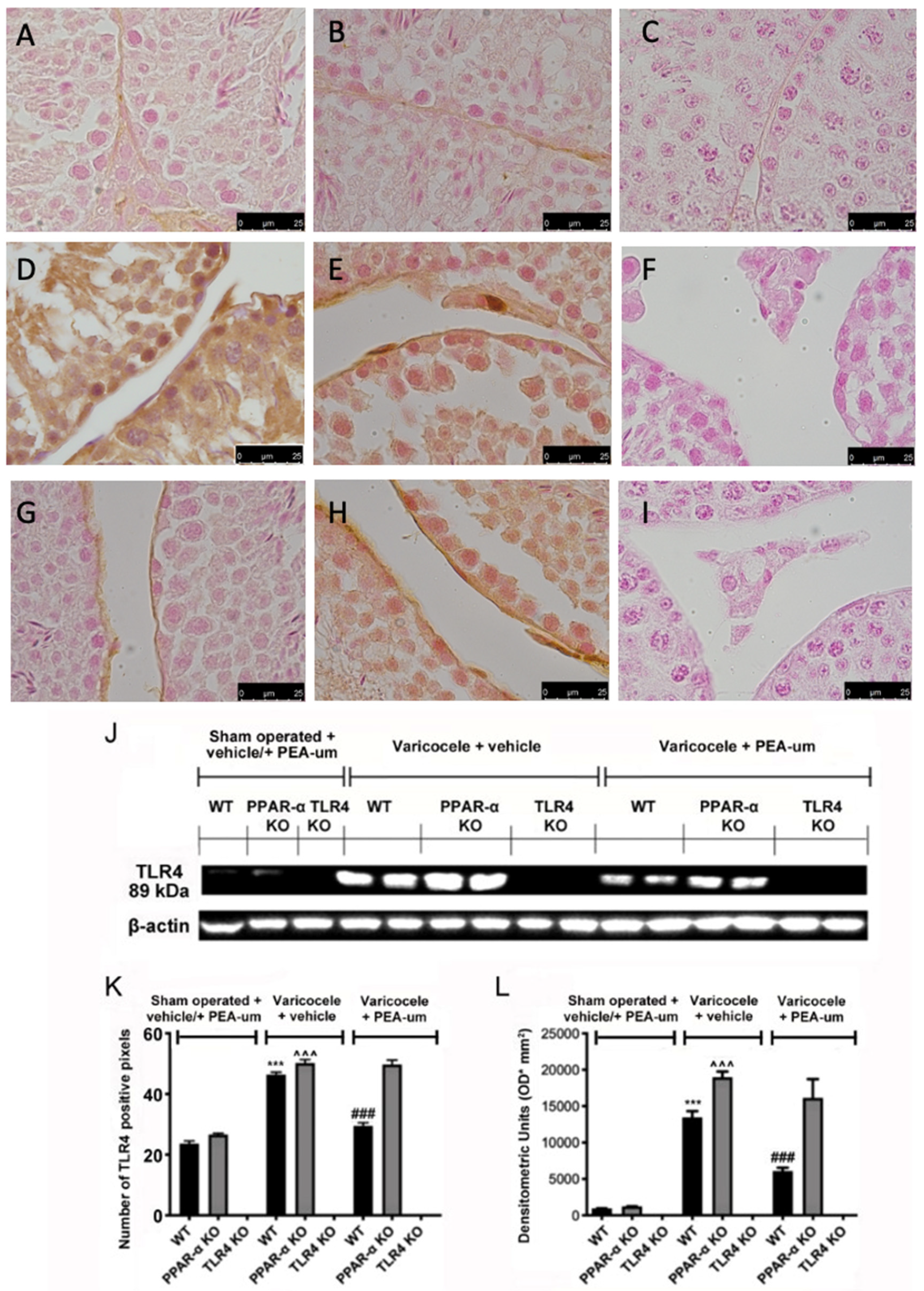
3.2. Genetic Deficiency of PPAR-α and TLR4 KO
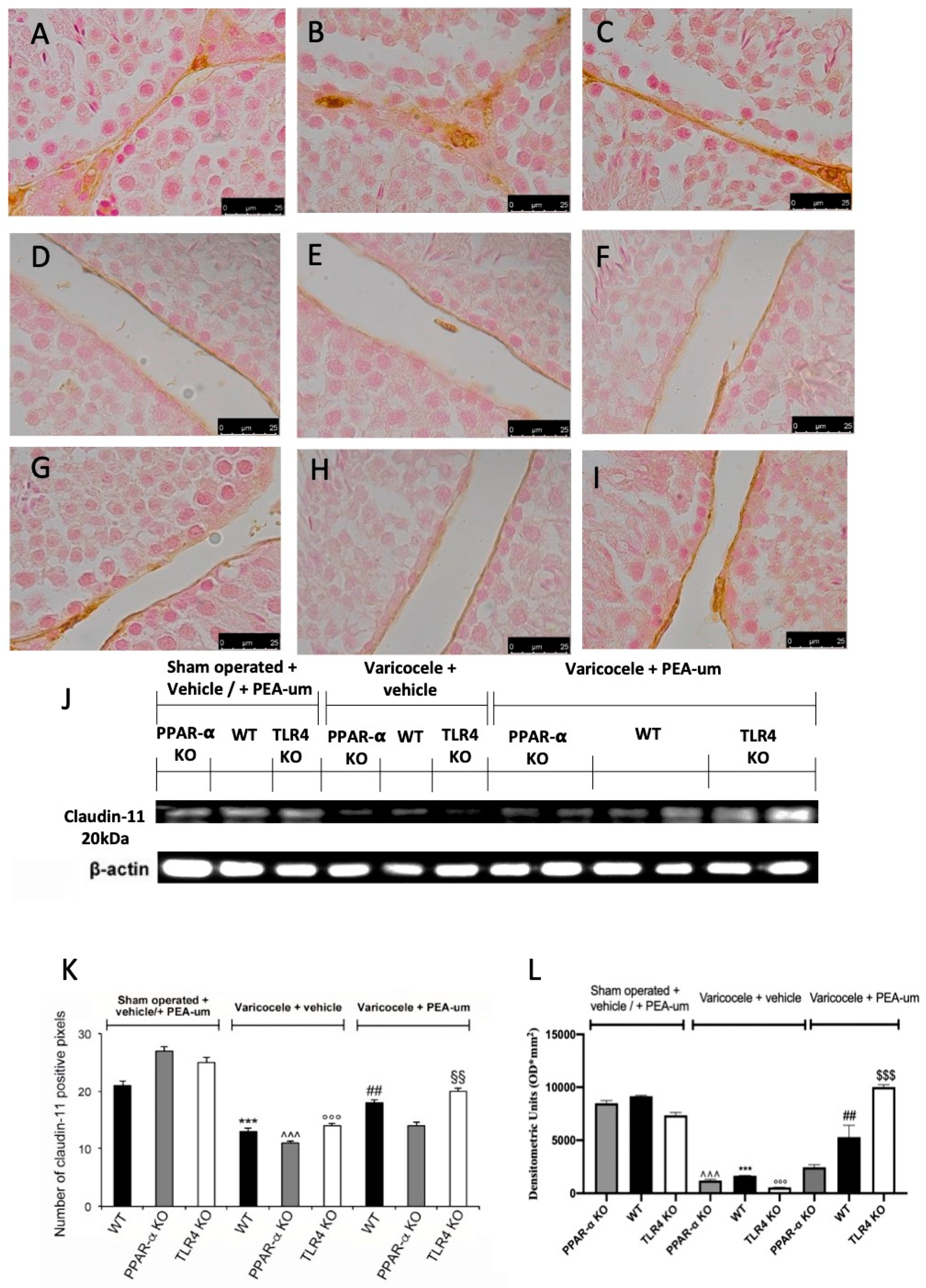
3.3. Effects of the Absence of PPAR-α and TLR4 on Tight Junction Proteins Staining
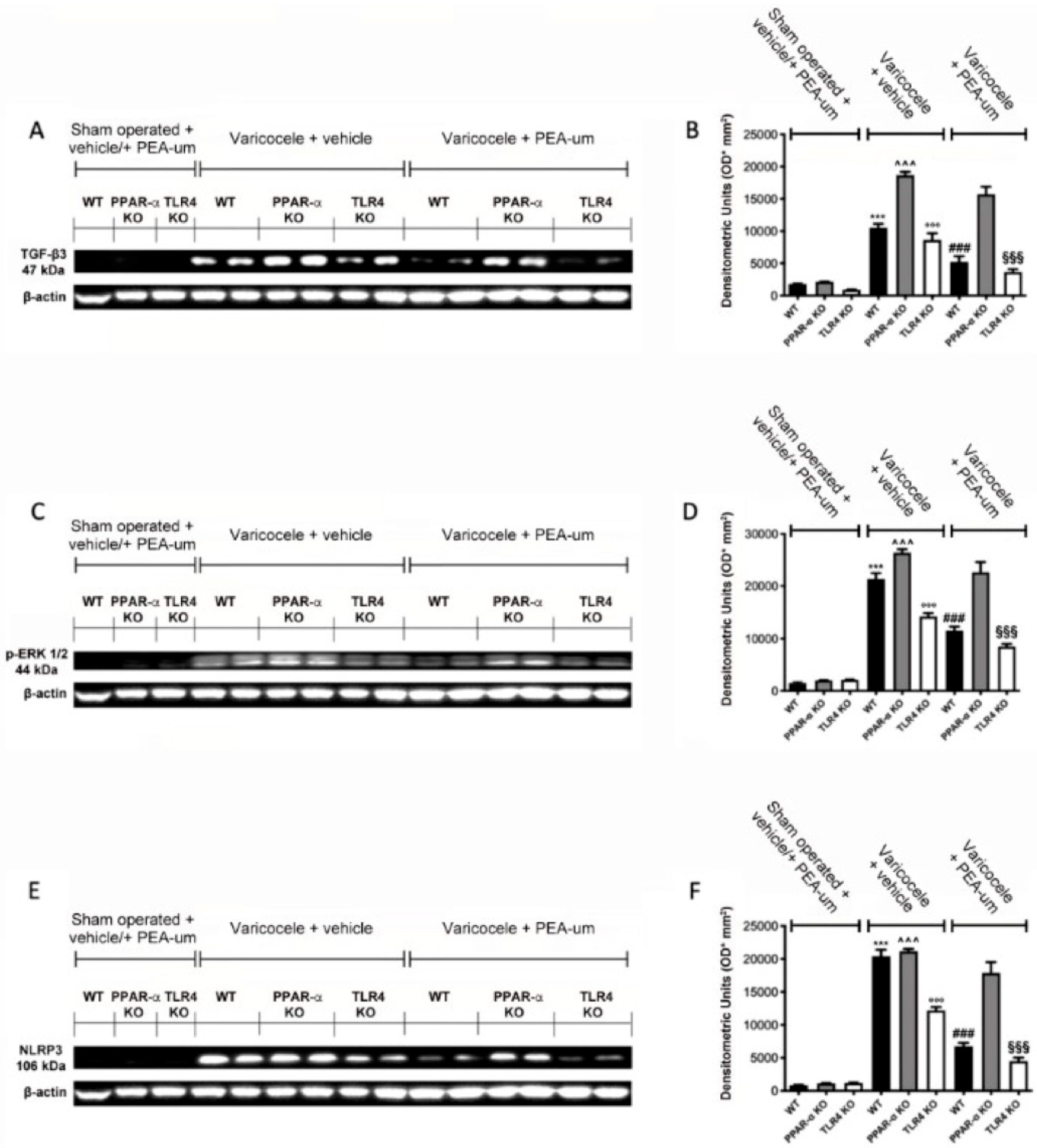
3.4. Effects of the Absence of PPAR-α and TLR4 on TGF-β3, p-ERK 1/2, and NLRP3 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Razi, M.; Malekinejad, H. Varicocele-Induced Infertility in Animal Models. Int. J. Fertil. Steril. 2015, 9, 141–149. [Google Scholar] [CrossRef]
- Hassanin, A.M.; Ahmed, H.H.; Kaddah, A.N. A global view of the pathophysiology of varicocele. Andrology 2018, 6, 654–661. [Google Scholar] [CrossRef]
- Hajipour, E.; Mashayekhi, F.J.; Mosayebi, G.; Baazm, M.; Zendedel, A. Resveratrol decreases apoptosis and NLRP3 complex expressions in experimental varicocele rat model. Iran. J. Basic Med. Sci. 2018, 21, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.K.; Zhang, L.T.; Choi, B.R.; Karna, K.K.; You, J.H.; Shin, Y.S.; Lee, S.W.; Kim, C.Y.; Zhao, C.; Chae, H.J.; et al. Protective effect of MOTILIPERM in varicocele-induced oxidative injury in rat testis by activating phosphorylated inositol requiring kinase 1α (p-IRE1α) and phosphorylated c-Jun N-terminal kinase (p-JNK) pathways. Pharm. Biol. 2018, 56, 94–103. [Google Scholar] [CrossRef]
- Kroese, A.C.; de Lange, N.M.; Collins, J.A.; Evers, J.L. Varicocele surgery, new evidence. Hum. Reprod. Update 2013, 19, 317. [Google Scholar] [CrossRef]
- Busetto, G.M.; Agarwal, A.; Virmani, A.; Antonini, G.; Ragonesi, G.; Del Giudice, F.; Micic, S.; Gentile, V.; De Berardinis, E. Effect of metabolic and antioxidant supplementation on sperm parameters in oligo-astheno-teratozoospermia, with and without varicocele: A double-blind placebo-controlled study. Andrologia 2018, 50, e12927. [Google Scholar] [CrossRef] [PubMed]
- Razi, M.; Tavalaee, M.; Sarrafzadeh-Rezaei, F.; Moazamian, A.; Gharagozloo, P.; Drevet, J.R.; Nasr-Eshafani, M.H. Varicocele and Oxidative Stress: New Perspectives from Animal and Human Studies. Andrology 2020. [Google Scholar] [CrossRef]
- Wang, F.; Chen, R.; Han, D. Innate Immune Defense in the Male Reproductive System and Male Fertility. In Innate Immunity in Health and Disease; Saxena, S.K., Prakash, H., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Politch, J.A.; Tucker, L.; Bowman, F.P.; Anderson, D.J. Concentrations and significance of cytokines and other immunologic factors in semen of healthy fertile men. Hum. Reprod. 2007, 22, 2928–2935. [Google Scholar] [CrossRef]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef] [PubMed]
- Girling, J.E.; Hedger, M.P. Toll-like receptors in the gonads and reproductive tract: Emerging roles in reproductive physiology and pathology. Immunol. Cell Biol. 2007, 85, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Lakpour, M.R.; Koruji, M.; Shahverdi, A.; Aghajanpour, S.; Rajabian Naghandar, M.; Sadighi Gilani, M.A.; Sabbaghian, M.; Aflatoonian, R. The Expression of TLR2 and TLR3 in Sertoli Cells of Azoospermic Patients. Cell J. 2017, 19, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Naas, H.; de Oliveira, A.A.; Karpova, T.; Nunes, K.P. Toll-like receptor 4 (TLR4) as a possible pathological mechanism in hyperglycemia-associated testicular dysfunction. Med. Hypotheses 2019, 127, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, H.; Xiong, W.; Chen, S.; Tang, H.; Han, D. Expression patterns and functions of toll-like receptors in mouse Sertoli cells. Endocrinology 2008, 149, 4402–4412. [Google Scholar] [CrossRef]
- Riccioli, A.; Starace, D.; Galli, R.; Fuso, A.; Scarpa, S.; Palombi, F.; De Cesaris, P.; Ziparo, E.; Filippini, A. Sertoli cells initiate testicular innate immune responses through TLR activation. J. Immunol. 2006, 177, 7122–7130. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, T.; Deng, T.; Xiong, W.; Lui, P.; Li, N.; Chen, Y.; Han, D. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Mol. Cell Endocrinol. 2013, 365, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jin, H.; Wu, L.; Shao, J.; Wu, X.; Lu, Y.; Zheng, S. Ligustrazine disrupts lipopolysaccharide-activated NLRP3 inflammasome pathway associated with inhibition of Toll-like receptor 4 in hepatocytes. Biomed. Pharmacother. 2016, 78, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef]
- Hong, F.; Pan, S.; Guo, Y.; Xu, P.; Zhai, Y. PPARs as Nuclear Receptors for Nutrient and Energy Metabolism. Molecules 2019, 24, 2545. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; Duval, C.; Muller, M.; Kersten, S. PPARs, Obesity, and Inflammation. PPAR Res. 2007, 2007, 95974. [Google Scholar] [CrossRef] [PubMed]
- Kadivar, A.; Heidari Khoei, H.; Hassanpour, H.; Ghanaei, H.; Golestanfar, A.; Mehraban, H.; Davoodian, N.; Dehghani Tafti, R. Peroxisome proliferator-activated receptors (PPARalpha, PPARgamma and PPARbeta/delta) gene expression profile on ram spermatozoa and their relation to the sperm motility. Vet. Res. Forum. 2016, 7, 27–34. [Google Scholar]
- Wang, G.; Cheng, S.; Zhang, S.; Zhu, Y.; Xiao, Y.; Ju, L. LPS impairs steroidogenesis and ROS metabolism and induces PPAR transcriptional activity to disturb estrogen/androgen receptor expression in testicular cells. Mol. Biol. Rep. 2020, 47, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Froment, P.; Gizard, F.; Defever, D.; Staels, B.; Dupont, J.; Monget, P. Peroxisome proliferator-activated receptors in reproductive tissues: From gametogenesis to parturition. J. Endocrinol. 2006, 189, 199–209. [Google Scholar] [CrossRef]
- Regueira, M.; Riera, M.F.; Galardo, M.N.; Pellizzari, E.H.; Cigorraga, S.B.; Meroni, S.B. Activation of PPAR α and PPAR β/δ regulates Sertoli cell metabolism. Mol. Cell Endocrinol. 2014, 382, 271–281. [Google Scholar] [CrossRef]
- Hasan, M.M.; El-Shal, A.S.; Mackawy, A.M.H.; Ibrahim, E.M.; Abdelghany, E.M.M.A.; Saeed, A.A.; El-Gendy, J. Ameliorative effect of combined low dose of Pioglitazone and omega-3 on spermatogenesis and steroidogenesis in diabetic rats. J. Cell Biochem. 2020, 121, 1524–1540. [Google Scholar] [CrossRef]
- Cerrato, S.; Brazis, P.; Della Valle, M.F.; Miolo, A.; Petrosino, S.; Di Marzo, V.; Puigdemont, A. Effects of palmitoylethanolamide on the cutaneous allergic inflammatory response in Ascaris hypersensitive Beagle dogs. Vet. J. 2012, 191, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Cristiano, C.; Avagliano, C.; De Caro, C.; La Rana, G.; Raso, G.M.; Canani, R.B.; Meli, R.; Calignano, A. Gut-brain Axis: Role of Lipids in the Regulation of Inflammation, Pain and CNS Diseases. Curr. Med. Chem. 2018, 25, 3930–3952. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Campolo, M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. Effect of a new formulation of micronized and ultramicronized N-palmitoylethanolamine in a tibia fracture mouse model of complex regional pain syndrome. PLoS ONE. 2017, 12, e0178553. [Google Scholar] [CrossRef]
- Di Paola, R.; Cordaro, M.; Crupi, R.; Siracusa, R.; Campolo, M.; Bruschetta, G.; Fusco, R.; Pugliatti, P.; Esposito, E.; Cuzzocrea, S. Protective effects of ultramicronized palmitoylethanolamide (PEA-um) in myocardial ischaemia and reperfusion injury in vivo. Shock 2016, 46, 202–213. [Google Scholar] [CrossRef]
- Di Paola, R.; Impellizzeri, D.; Fusco, R.; Cordaro, M.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Ultramicronized palmitoylethanolamide (PEA-um®) in the treatment of idiopathic pulmonary fibrosis. Pharmacol. Res. 2016, 111, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; Evangelista, M.; Granese, R.; Cuzzocrea, S. Co-micronized palmitoylethanolamide/polydatin treatment causes endometriotic lesion regression in a rodent model of surgically induced endometriosis. Front. Pharmacol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Britti, D.; Crupi, R.; Impellizzeri, D.; Gugliandolo, E.; Fusco, R.; Schievano, C.; Morittu, V.M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. A novel composite formulation of palmitoylethanolamide and quercetin decreases inflammation and relieves pain in inflammatory and osteoarthritic pain models. BMC Vet. Res. 2017, 13, 229. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Inferrera, A.; Esposito, E.; Di Paola, R.; Cuzzocrea, S. Effects of a co-micronized composite containing palmitoylethanolamide and polydatin in an experimental model of benign prostatic hyperplasia. Toxicol. Appl. Pharmacol. 2017, 329, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The nuclear receptor peroxisome proliferator-activated receptor-α mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; D’Amico, R.; Fusco, R.; Evangelista, M.; Cuzzocrea, S. The neuroprotective effects of micronized PEA (PEA-m) formulation on diabetic peripheral neuropathy in mice. FASEB J. 2019, 33, 11364–11380. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Arena, S.; Antonuccio, P.; Romeo, C.; Bitto, A.; Magno, C.; Rinaldi, M.; Micali, A.; Irrera, N.; Pizzino, G.; et al. Role of Inhibitors of Apoptosis Proteins in Testicular Function and Male Fertility: Effects of Polydeoxyribonucleotide Administration in Experimental Varicocele. Biomed. Res. Int. 2015, 2015, 248976. [Google Scholar] [CrossRef]
- Antonuccio, P.; Micali, A.; Puzzolo, D.; Romeo, C.; Vermiglio, G.; Squadrito, V.; Freni, J.; Pallio, G.; Trichilo, V.; Righi, M.; et al. Nutraceutical Effects of Lycopene in Experimental Varicocele: An “In Vivo” Model to Study Male Infertility. Nutrients 2020, 12, 1536. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Arena, S.; Bonvissuto, G.; Bitto, A.; Polito, F.; Irrera, N.; Arena, F.; Fragalà, E.; Romeo, C.; Nicotina, P.A.; et al. Activation of adenosine A2A receptors by polydeoxyribonucleotide increases vascular endothelial growth factor and protects against testicular damage induced by experimental varicocele in rats. Fertil. Steril. 2011, 95, 1510–1513. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, S.G. Testicular biopsy score count—A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Mizuno, K.; Hayashi, Y.; Kojima, Y.; Kurokawa, S.; Sasaki, S.; Kohri, K. Influence for testicular development and histological peculiarity in the testes of flutamide-induced cryptorchid rat model. Int. J. Urol. 2007, 14, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Fusco, R.; Biundo, F.; D’Amico, R.; Benedetto, F.; Di Paola, R.; Cuzzocrea, S. Palmitoylethanolamide and Polydatin combination reduces inflammation and oxidative stress in vascular injury. Pharmacol. Res. 2017, 123, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; D’amico, R.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Absence of formyl peptide receptor 1 causes endometriotic lesion regression in a mouse model of surgically-induced endometriosis. Oncotarget 2018, 9, 31355–31366. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; D’Amico, R.; Peditto, M.; Oteri, G.; Di Paola, R.; Cuzzocrea, S.; Navarra, M. Treatment with a Flavonoid-Rich Fraction of Bergamot Juice Improved Lipopolysaccharide-Induced Periodontitis in Rats. Front. Pharmacol. 2019, 9, 1563. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; Ginestra, G.; D’amico, R.; Bisignano, C.; Mandalari, G.; Cuzzocrea, S.; Di Paola, R. Involvement of TLR4 and PPAR-α Receptors in Host Response and NLRP3 Inflammosome Activation, Against Pulmonary Infection with Pseudomosas Aeruginosa. Shock 2019, 15, 221–227. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Campolo, M.; Latteri, S.; Carughi, A.; Mandalari, G.; Cuzzocrea, S. The Antioxidant Activity of Pistachios Reduces Cardiac Tissue Injury of Acute Ischemia/Reperfusion (I/R) in Diabetic Streptozotocin (STZ)-Induced Hyperglycaemic Rats. Front. Pharmacol. 2018, 9, 51. [Google Scholar] [CrossRef]
- Cho, K.S.; Seo, J.T. Effect of varicocelectomy on male infertility. Korean J. Urol. 2014, 55, 703–799. [Google Scholar] [CrossRef] [PubMed]
- Keppel Hesselink, J.M.; Kopsky, D.J. Palmitoylethanolamide, a neutraceutical, in nerve compression syndromes: Efficacy and safety in sciatic pain and carpal tunnel syndrome. J. Pain Res. 2015, 8, 729–734. [Google Scholar] [CrossRef]
- Petrosino, S.; Di Marzo, V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, B.; Crupi, R.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Beneficial Effects of Co-Ultramicronized Palmitoylethanolamide/ Luteolin in a Mouse Model of Autism and in a Case Report of Autism. CNS Neurosci. Ther. 2017, 23, 87–98. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Cordaro, M.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Crupi, R.; Genovese, T.; Evangelista, M.; Di Paola, R.; et al. The Protective Effects of Pre- and Post-Administration of Micronized Palmitoylethanolamide Formulation on Postoperative Pain in Rats. Int. J. Mol. Sci. 2020, 21, 7700. [Google Scholar] [CrossRef] [PubMed]
- Peritore, A.F.; Siracusa, R.; Crupi, R.; Cuzzocrea, S. Therapeutic Efficacy of Palmitoylethanolamide and Its New Formulations in Synergy with Different Antioxidant Molecules Present in Diets. Nutrients 2019, 11, 2175. [Google Scholar] [CrossRef]
- Venables, B.J.; Waggoner, C.A.; Chapman, K.D. N-acylethanolamines in seeds of selected legumes. Phytochemistry 2005, 66, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.D.; Venables, B.; Markovic, R.; Blair, R.W., Jr.; Bettinger, C. N-acylethanolamines in seeds. Quantification of molecular species and their degradation upon imbibition. Plant Physiol. 1999, 120, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Rawyler, A.J.; Braendle, R.A. N-acylphosphatidylethanolamine accumulation in potato cells upon energy shortage caused by anoxia or respiratory inhibitors. Plant Physiol. 2001, 127, 240–251. [Google Scholar] [CrossRef]
- Lam, P.M.; Marczylo, T.H.; Konje, J.C. Simultaneous measurement of three n-acylethanolamides in human bio-matrices using ultra performance liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Micali, A.; Pisani, A.; Puzzolo, D.; Bitto, A.; Rinaldi, M.; Pizzino, G.; Irrera, N.; Galfo, F.; Arena, S.; et al. Flavocoxid Protects Against Cadmium-Induced Disruption of the Blood-Testis Barrier and Improves Testicular Damage and Germ Cell Impairment in Mice. Toxicol. Sci. 2015, 148, 311–329. [Google Scholar] [CrossRef]
- Oh, Y.S.; Jo, N.H.; Park, J.K.; Gye, M.C. Changes in Inflammatory Cytokines Accompany Deregulation of Claudin-11, Resulting in Inter-Sertoli Tight Junctions in Varicocele Rat Testes. J. Urol. 2016, 196, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- McCabe, M.J.; Tarulli, G.A.; Meachem, S.J.; Robertson, D.M.; Smooker, P.M.; Stanton, P.G. Gonadotropins regulate rat testicular tight junctions in vivo. Endocrinology 2010, 151, 2911–2922. [Google Scholar] [CrossRef] [PubMed]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-induced Hyperpermeability of the Human Gut In Vitro and In Vivo-A Randomized, Placebo-controlled, Double-blind Controlled Trial. Inflamm. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef]
- Minutoli, L.; Antonuccio, P.; Polito, F.; Bitto, A.; Squadrito, F.; Irrera, N.; Nicotina, P.A.; Fazzari, C.; Montalto, A.S.; Di Stefano, V.; et al. Peroxisome proliferator activated receptor beta/delta activation prevents extracellular regulated kinase 1/2 phosphorylation and protects the testis from ischemia and reperfusion injury. J. Urol. 2009, 181, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Baazm, M.; Ghafarizadeh, A.A.; Noshad Kamran, A.R.; Beyer, C.; Zendedel, A. Presence of The NLRP3 Inflammasome Components in Semen of Varicocele Patients. Int. J. Fertil. Steril. 2020, 14, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Antonuccio, P.; Irrera, N.; Rinaldi, M.; Bitto, A.; Marini, H.; Pizzino, G.; Romeo, C.; Pisani, A.; Santoro, G.; et al. NLRP3 Inflammasome Involvement in the Organ Damage and Impaired Spermatogenesis Induced by Testicular Ischemia and Reperfusion in Mice. J. Pharmacol. Exp. Ther. 2015, 355, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Benvenga, S.; Micali, A.; Pallio, G.; Vita, R.; Malta, C.; Puzzolo, D.; Irrera, N.; Squadrito, F.; Altavilla, D.; Minutoli, L. Effects of Myo-inositol Alone and in Combination with Seleno-L-methionine on Cadmium-Induced Testicular Damage in Mice. Curr. Mol. Pharmacol. 2019, 12, 311–323. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonuccio, P.; Marini, H.R.; Micali, A.; Romeo, C.; Granese, R.; Retto, A.; Martino, A.; Benvenga, S.; Cuzzocrea, S.; Impellizzeri, D.; et al. The Nutraceutical N-Palmitoylethanolamide (PEA) Reveals Widespread Molecular Effects Unmasking New Therapeutic Targets in Murine Varicocele. Nutrients 2021, 13, 734. https://doi.org/10.3390/nu13030734
Antonuccio P, Marini HR, Micali A, Romeo C, Granese R, Retto A, Martino A, Benvenga S, Cuzzocrea S, Impellizzeri D, et al. The Nutraceutical N-Palmitoylethanolamide (PEA) Reveals Widespread Molecular Effects Unmasking New Therapeutic Targets in Murine Varicocele. Nutrients. 2021; 13(3):734. https://doi.org/10.3390/nu13030734
Chicago/Turabian StyleAntonuccio, Pietro, Herbert Ryan Marini, Antonio Micali, Carmelo Romeo, Roberta Granese, Annalisa Retto, Antonia Martino, Salvatore Benvenga, Salvatore Cuzzocrea, Daniela Impellizzeri, and et al. 2021. "The Nutraceutical N-Palmitoylethanolamide (PEA) Reveals Widespread Molecular Effects Unmasking New Therapeutic Targets in Murine Varicocele" Nutrients 13, no. 3: 734. https://doi.org/10.3390/nu13030734