Increased Risk of High Body Fat and Altered Lipid Metabolism Associated to Suboptimal Consumption of Vitamin A Is Modulated by Genetic Variants rs5888 (SCARB1), rs1800629 (UCP1) and rs659366 (UCP2)



Abstract
:1. Introduction
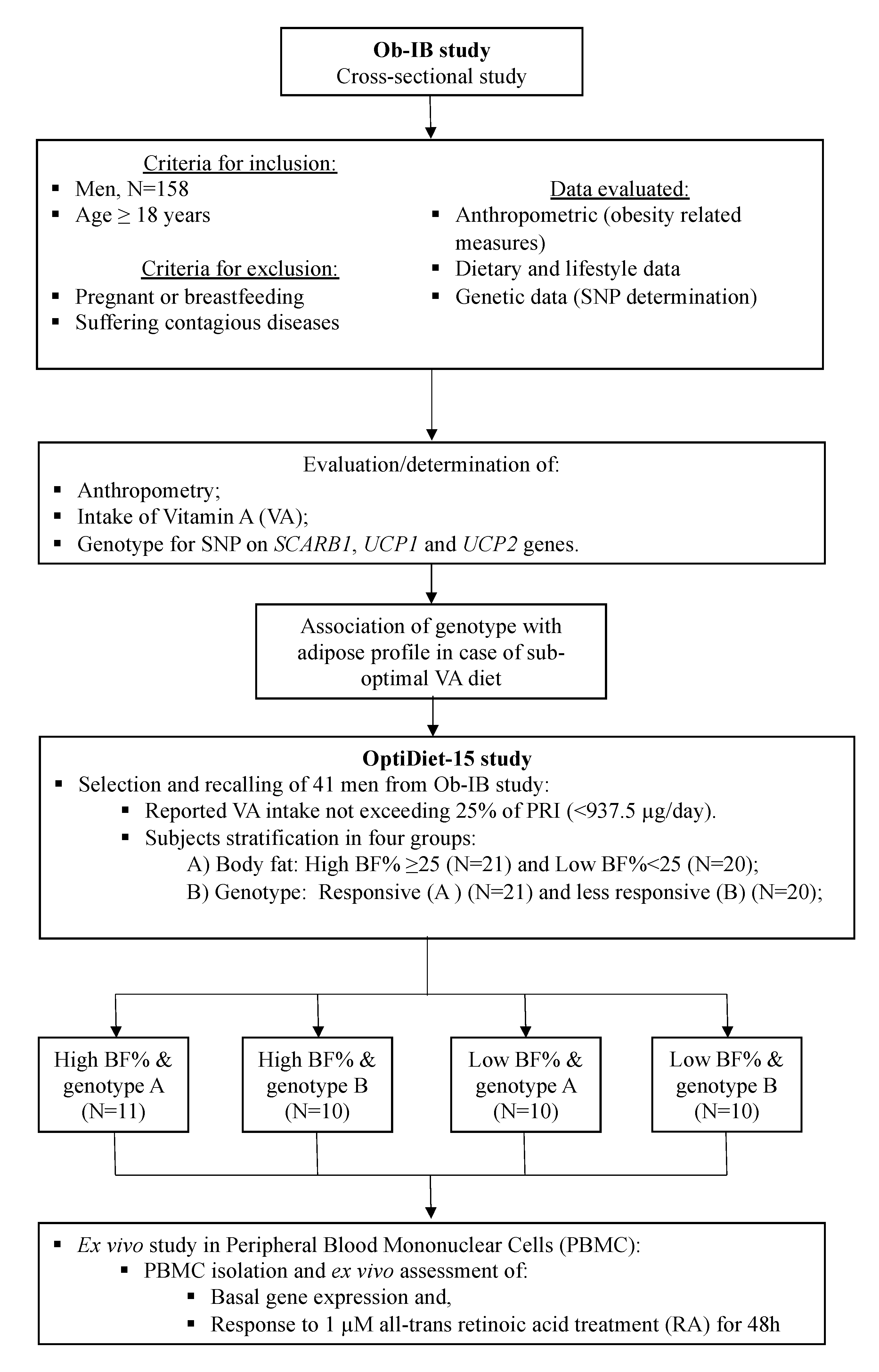
2. Materials and Methods
2.1. Subjects
2.2. Anthropometric Measures
2.3. Estimation of Dietary Intake
2.4. SNP Selection and Population Grouping According to Genotype
2.5. DNA Extraction and Genotype Determination
2.6. Blood Sample Collection, PBMC Isolation, and Ex Vivo Treatment
2.7. RNA Extraction and Gene Expression Analysis
2.8. Statistical Analyses
3. Results
3.1. Evaluation of VA Intake Level and Genotype Impact on Adiposity in Men of the Ob-IB Study
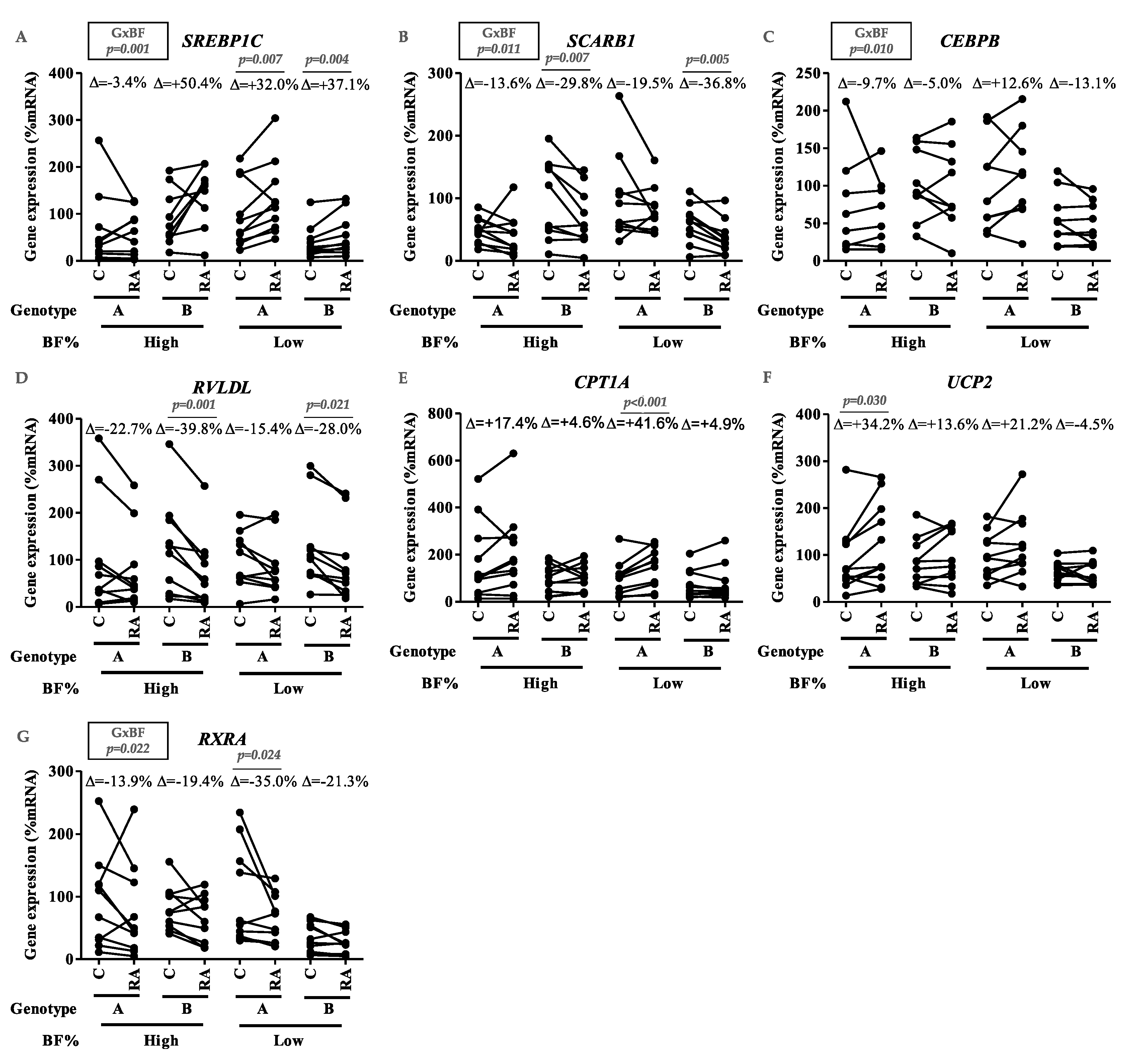
3.2. Assessment of the Influence of Suboptimal VA Intake, Genotype, and Adiposity on PBMC Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seidell, J.C.; Halberstadt, J. The Global Burden of Obesity and the Challenges of Prevention. Ann. Nutr. Metab. 2015, 66, 7–12. [Google Scholar] [CrossRef]
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiseman, E.M.; Bar-El Dadon, S.; Reifen, R. The vicious cycle of vitamin a deficiency: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3703–3714. [Google Scholar] [CrossRef] [PubMed]
- García, O.P. Effect of vitamin A deficiency on the immune response in obesity. Proc. Nutr. Soc. 2012, 71, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, L.A.; Benyshek, D.C.; Martin, J.F. Food acquisition habits, nutrient intakes, and anthropometric data of Havasupai adults. J. Am. Diet. Assoc. 1997, 97, 1275–1282. [Google Scholar] [CrossRef]
- Viroonudomphol, D.; Pongpaew, P.; Tungtrongchitr, R.; Changbumrung, S.; Tungtrongchitr, A.; Phonrat, B.; Vudhivai, N.; Schelp, F.P. The relationships between anthropometric measurements, serum vitamin A and E concentrations and lipid profiles in overweight and obese subjects. Asia Pac. J. Clin. Nutr. 2003, 12, 73–79. [Google Scholar] [PubMed]
- García, O.; Ronquillo, D.; del Carmen Caamaño, M.; Martínez, G.; Camacho, M.; López, V.; Rosado, J. Zinc, Iron and Vitamins A, C and E Are Associated with Obesity, Inflammation, Lipid Profile and Insulin Resistance in Mexican School-Aged Children. Nutrients 2013, 5, 5012–5030. [Google Scholar] [CrossRef] [Green Version]
- Bonet, M.L.; Ribot, J.; Galmés, S.; Serra, F.; Palou, A. Carotenoids and carotenoid conversion products in adipose tissue biology and obesity: Pre-clinical and human studies. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158676. [Google Scholar] [CrossRef]
- Blomhoff, R.; Blomhoff, H.K. Overview of retinoid metabolism and function. J. Neurobiol. 2006, 66, 606–630. [Google Scholar] [CrossRef]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids and their conversion products in the control of adipocyte function, adiposity and obesity. Arch. Biochem. Biophys. 2015, 572, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Bonet, M.L.; Ribot, J.; Felipe, F.; Palou, A. Vitamin A and the regulation of fat reserves. Cell. Mol. Life Sci. 2003, 60, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Bonet, M.L.; Ribot, J.; Palou, A. Lipid metabolism in mammalian tissues and its control by retinoic acid. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 1821, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Bonet, M.L.; Mercader, J.; Palou, A. A nutritional perspective on UCP1-dependent thermogenesis. Biochimie 2017, 134, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.; Petrov, P.; Bonet, M.L.; Ribot, J.; Palou, A. Induction of carnitine palmitoyl transferase 1 and fatty acid oxidation by retinoic acid in HepG2 cells. Int. J. Biochem. Cell Biol. 2012, 44, 2019–2027. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Canas, J.A.; Lochrie, A.; McGowan, A.G.; Hossain, J.; Schettino, C.; Balagopal, P.B. Effects of mixed carotenoids on adipokines and abdominal adiposity in children: A pilot study. J. Clin. Endocrinol. Metab. 2017, 102, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Beydoun, M.A.; Chen, X.; Jha, K.; Beydoun, H.A.; Zonderman, A.B.; Canas, J.A. Carotenoids, vitamin A, and their association with the metabolic syndrome: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 32–45. [Google Scholar] [CrossRef]
- Borel, P.; Desmarchelier, C.; Nowicki, M.; Bott, R. A Combination of Single-Nucleotide Polymorphisms Is Associated with Interindividual Variability in Dietary -Carotene Bioavailability in Healthy Men. J. Nutr. 2015, 145, 1740–1747. [Google Scholar] [CrossRef] [Green Version]
- Gueguen, S.; Leroy, P.; Gueguen, R.; Siest, G.; Visvikis, S.; Herbeth, B. Genetic and environmental contributions to serum retinol and alpha-tocopherol concentrations: The Stanislas Family Study. Am. J. Clin. Nutr. 2005, 81, 1034–1044. [Google Scholar] [CrossRef]
- Mondul, A.M.; Yu, K.; Wheeler, W.; Zhang, H.; Weinstein, S.J.; Major, J.M.; Cornelis, M.C.; Männistö, S.; Hazra, A.; Hsing, A.W.; et al. Genome-wide association study of circulating retinol levels. Hum. Mol. Genet. 2011, 20, 4724–4731. [Google Scholar] [CrossRef]
- Valacchi, G.; Sticozzi, C.; Lim, Y.; Pecorelli, A. Scavenger receptor class B type I: A multifunctional receptor. Ann. N. Y. Acad. Sci. 2011, 1229, E1–E7. [Google Scholar] [CrossRef] [PubMed]
- Mercader, J.; Ribot, J.; Murano, I.; Felipe, F.; Cinti, S.; Bonet, M.L.; Palou, A. Remodeling of White Adipose Tissue after Retinoic Acid Administration in Mice. Endocrinology 2006, 147, 5325–5332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felipe, F.; Bonet, M.L.; Ribot, J.; Palou, A. Up-regulation of muscle uncoupling protein 3 gene expression in mice following high fat diet, dietary vitamin A supplementation and acute retinoic acid-treatment. Int. J. Obes. 2003, 27, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borel, P.; Moussa, M.; Reboul, E.; Lyan, B.; Defoort, C.; Vincent-Baudry, S.; Maillot, M.; Gastaldi, M.; Darmon, M.; Portugal, H.; et al. Human Plasma Levels of Vitamin E and Carotenoids Are Associated with Genetic Polymorphisms in Genes Involved in Lipid Metabolism. J. Nutr. 2007, 137, 2653–2659. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimzadeh Attari, V.; Asghari Jafarabadi, M.; Zemestani, M.; Ostadrahimi, A. Effect of Zingiber officinale Supplementation on Obesity Management with Respect to the Uncoupling Protein 1 -3826A > G and β3-adrenergic Receptor Trp64Arg Polymorphism. Phyther. Res. 2015, 29, 1032–1039. [Google Scholar] [CrossRef]
- Snitker, S.; Fujishima, Y.; Shen, H.; Ott, S.; Pi-Sunyer, X.; Furuhata, Y.; Sato, H.; Takahashi, M. Effects of novel capsinoid treatment on fatness and energy metabolism in humans: Possible pharmacogenetic implications. Am. J. Clin. Nutr. 2009, 89, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergman, R.N.; Stefanovski, D.; Buchanan, T.A.; Sumner, A.E.; Reynolds, J.C.; Sebring, N.G.; Xiang, A.H.; Watanabe, R.M. A Better Index of Body Adiposity. Obesity 2011, 19, 1083–1089. [Google Scholar] [CrossRef]
- Ortega, R.M.; Andrés, P.; Requejo, A.M.; Aparicio, A.; Molinero, L.M.; López-Sobaler, A.M. DIAL software for assessing diets and food calculations (for Windows, version 2.0). Department of Nutrition (UCM) & Alce Ingeniería, S.L. Madrid, Spain. Available online: https://www.alceingenieria.net/infodial.htm (accessed on 20 July 2020).
- European Food Safety Authority (EFSA). Dietary Reference Values: Vitamin A Advice Published|European Food Safety Authority. Available online: https://www.efsa.europa.eu/en/press/news/150305 (accessed on 12 June 2017).
- Shen, W.-J.; Azhar, S.; Kraemer, F.B. SR-B1: A Unique Multifunctional Receptor for Cholesterol Influx and Efflux. Annu. Rev. Physiol. 2018, 80, 95–116. [Google Scholar] [CrossRef]
- Borel, P.; Lietz, G.; Goncalves, A.; Szabo de Edelenyi, F.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C.; et al. CD36 and SR-BI Are Involved in Cellular Uptake of Provitamin A Carotenoids by Caco-2 and HEK Cells, and Some of Their Genetic Variants Are Associated with Plasma Concentrations of These Micronutrients in Humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Murholm, M.; Isidor, M.S.; Basse, A.L.; Winther, S.; Sørensen, C.; Skovgaard-Petersen, J.; Nielsen, M.M.; Hansen, A.S.; Quistorff, B.; Hansen, J.B. Retinoic acid has different effects on UCP1 expression in mouse and human adipocytes. BMC Cell Biol. 2013, 14, 41. [Google Scholar] [CrossRef] [Green Version]
- Pierelli, G.; Stanzione, R.; Forte, M.; Migliarino, S.; Perelli, M.; Volpe, M.; Rubattu, S. Uncoupling protein 2: A key player and a potential therapeutic target in vascular diseases. Oxid. Med. Cell. Longev. 2017, 2017, 7348372. [Google Scholar] [CrossRef] [PubMed]
- Serra, F.; Bonet, M.L.; Puigserver, P.; Oliver, J.; Palou, A. Stimulation of uncoupling protein 1 expression in brown adipocytes by naturally occurring carotenoids. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 650–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larose, M.; Cassard-Doulcier, A.M.; Fleury, C.; Serra, F.; Champigny, O.; Bouillaud, F.; Ricquier, D. Essential cis-acting elements in rat uncoupling protein gene are in an enhancer containing a complex retinoic acid response domain. J. Biol. Chem. 1996, 271, 31533–31542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonet, M.L.; Puigserver, P.; Serra, F.; Ribot, J.; Vázquez, F.; Pico, C.; Palou, A. Retinoic acid modulates retinoid X receptor alpha and retinoic acid receptor alpha levels of cultured brown adipocytes. FEBS Lett. 1997, 406, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Cadenas, S. Mitochondrial uncoupling, ROS generation and cardioprotection. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 940–950. [Google Scholar] [CrossRef]
- Constantineau, J.; Greason, E.; West, M.; Filbin, M.; Kieft, J.S.; Carletti, M.Z.; Christenson, L.K.; Rodriguez, A. A synonymous variant in scavenger receptor, class B, type I gene is associated with lower SR-BI protein expression and function. Atherosclerosis 2010, 210, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Nagai, N.; Sakane, N.; Tsuzaki, K.; Moritani, T. UCP1 genetic polymorphism (–3826 A/G) diminishes resting energy expenditure and thermoregulatory sympathetic nervous system activity in young females. Int. J. Obes. 2011, 35, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Yoneshiro, T.; Ogawa, T.; Okamoto, N.; Matsushita, M.; Aita, S.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Impact of UCP1 and β3AR gene polymorphisms on age-related changes in brown adipose tissue and adiposity in humans. Int. J. Obes. 2013, 37, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, P.; Ma, L.; Hanson, R.L.; Franks, P.; Stumvoll, M.; Bogardus, C.; Baier, L.J. Genetic variation in UCP2 (uncoupling protein-2) is associated with energy metabolism in Pima Indians. Diabetologia 2005, 48, 2292–2295. [Google Scholar] [CrossRef] [Green Version]
- Galmés, S.; Cifre, M.; Palou, A.; Oliver, P.; Serra, F. A Genetic Score of Predisposition to Low-Grade Inflammation Associated with Obesity May Contribute to Discern Population at Risk for Metabolic Syndrome. Nutrients 2019, 11, 298. [Google Scholar] [CrossRef] [Green Version]
- Reynés, B.; Díaz-Rúa, R.; Cifre, M.; Oliver, P.; Palou, A. Peripheral blood mononuclear cells as a potential source of biomarkers to test the efficacy of weight-loss strategies. Obesity 2014, 23, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Cifre, M.; Díaz-Rua, R.; Varela Calviño, R.; Reynés, B.; Pericás-Beltrán, J.; Palou, A.; Oliver, P. Human peripheral blood mononuclear cell in vitro system to test the efficacy of food bioactive compounds: Effects of polyunsaturated fatty acids and their relation with BMI. Mol. Nutr. Food Res. 2016, 61, 1600353. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Dietary Reference Values for nutrients Summary report. EFSA Support. Publ. 2017, 14, e15121. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, E.; Ávila, J.M.; Valero, T.; del Pozo, S.; Rodriguez, P.; Aranceta-Bartrina, J.; Gil, Á.; González-Gross, M.; Ortega, R.M.; Serra-Majem, L.; et al. Energy Intake, Profile, and Dietary Sources in the Spanish Population: Findings of the ANIBES Study. Nutrients 2015, 7, 4739–4762. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on Dietary Reference Values for energy. EFSA J. 2013, 11, 3333. [Google Scholar] [CrossRef] [Green Version]
- Harari, A.; Coster, A.C.; Jenkins, A.; Xu, A.; Greenfield, J.R.; Harats, D.; Shaish, A.; Samocha-Bonet, D. Obesity and Insulin Resistance Are Inversely Associated with Serum and Adipose Tissue Carotenoid Concentrations in Adults. J. Nutr. 2020, 150, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Borel, P.; Desmarchelier, C. Genetic Variations Associated with Vitamin A Status and Vitamin A Bioavailability. Nutrients 2017, 9, 246. [Google Scholar] [CrossRef]
- Morabia, A.; Ross, B.M.; Costanza, M.C.; Cayanis, E.; Flaherty, M.S.; Alvin, G.B.; Das, K.; James, R.; Yang, A.-S.; Evagrafov, O.; et al. Population-based study of SR-BI genetic variation and lipid profile. Atherosclerosis 2004, 175, 159–168. [Google Scholar] [CrossRef]
- Ma, R.; Zhu, X.; Yan, B. SCARB1 rs5888 gene polymorphisms in coronary heart disease: A systematic review and a meta-analysis. Gene 2018, 678, 280–287. [Google Scholar] [CrossRef]
- Ye, L.-F.; Zheng, Y.-R.; Zhang, Q.-G.; Yu, J.-W.; Wang, L.-H. Meta-analysis of the association between SCARB1 polymorphism and fasting blood lipid levels. Oncotarget 2017, 8, 81145–81153. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Xu, K.; Xu, X.; Gu, R.; Liu, X.; Shan, S.; Yang, T. UCP2 -866G/A, Ala55Val and UCP3 -55C/T polymorphisms in association with obesity susceptibility—A meta-analysis study. PLoS ONE 2013, 8, e58939. [Google Scholar] [CrossRef] [Green Version]
- Andersen, G.; Dalgaard, L.T.; Justesen, J.M.; Anthonsen, S.; Nielsen, T.; Thørner, L.W.; Witte, D.; Jørgensen, T.; Clausen, J.O.; Lauritzen, T.; et al. The frequent UCP2 −866G > A polymorphism protects against insulin resistance and is associated with obesity: A study of obesity and related metabolic traits among 17 636 Danes. Int. J. Obes. 2013, 37, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Chae, S.; Cha, Y.; Park, Y. Supplementation of Korean fermented soy paste doenjang reduces visceral fat in overweight subjects with mutant uncoupling protein-1 allele. Nutr. Res. 2012, 32, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.; O’Brien, P. Obesity and the White Blood Cell Count: Changes with Sustained Weight Loss. Obes. Surg. 2006, 16, 251–257. [Google Scholar] [CrossRef]
- Koca, T.T. Does obesity cause chronic inflammation? The association between complete blood parameters with body mass index and fasting glucose. Pak. J. Med. Sci. 2017, 33, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Reynés, B.; Priego, T.; Cifre, M.; Oliver, P.; Palou, A. Peripheral Blood Cells, a Transcriptomic Tool in Nutrigenomic and Obesity Studies: Current State of the Art. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1006–1020. [Google Scholar] [CrossRef] [Green Version]
- Picó, C.; Serra, F.; Rodríguez, A.M.; Keijer, J.; Palou, A. Biomarkers of Nutrition and Health: New Tools for New Approaches. Nutrients 2019, 11, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurencikiene, J.; Rydé, M. Liver X receptors and fat cell metabolism. Int. J. Obes. 2012, 3621, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.M.; Stahl, A. SLC27 fatty acid transport proteins. Mol. Asp. Med. 2013, 34, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Caimari, A.; Oliver, P.; Rodenburg, W.; Keijer, J.; Palou, A. Slc27a2 expression in peripheral blood mononuclear cells as a molecular marker for overweight development. Int. J. Obes. 2010, 34, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Becuwe, P.; Ennen, M.; Klotz, R.; Barbieux, C.; Grandemange, S. Manganese superoxide dismutase in breast cancer: From molecular mechanisms of gene regulation to biological and clinical significance. Free Radic. Biol. Med. 2014, 77, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Grdina, D.J.; Murley, J.S.; Miller, R.C.; Mauceri, H.J.; Sutton, H.G.; Thirman, M.J.; Li, J.J.; Woloschak, G.E.; Weichselbaum, R.R. A Manganese Superoxide Dismutase (SOD2)-Mediated Adaptive Response. Radiat. Res. 2013, 179, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulven, S.M.; Dalen, K.T.; Gustafsson, J.-Å.; Nebb, H.I. LXR is crucial in lipid metabolism. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yeh, I.; Chou, S.; Yen, M.; Kuo, P. Regulatory mechanism of fatty acid-CoA metabolic enzymes under endoplasmic reticulum stress in lung cancer. Oncol. Rep. 2018, 40, 2674–2682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Spann, N.J.; Link, V.M.; Muse, E.D.; Strid, T.; Edillor, C.; Kolar, M.J.; Matsuzaka, T.; Hayakawa, S.; Tao, J.; et al. SREBP1 Contributes to Resolution of Pro-inflammatory TLR4 Signaling by Reprogramming Fatty Acid Metabolism. Cell Metab. 2017, 25, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.M.; Janssen, R.C.; Choudhury, M.; Baquero, K.C.; Aikens, R.M.; De La Houssaye, B.A.; Friedman, J.E. CCAAT/enhancer-binding protein β (C/EBPβ) expression regulates dietary-induced inflammation in macrophages and adipose tissue in mice. J. Biol. Chem. 2012, 287, 34349–34360. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, Y.; Li, R.; Chen, G. Transcriptional Factors Mediating Retinoic Acid Signals in the Control of Energy Metabolism. Int. J. Mol. Sci. 2015, 16, 14210–14244. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Male Subjects (Ob-IB Study) (n = 158) | Mean | SD | |
|---|---|---|---|
| Age (years) | 37 | 17 | |
| Anthropometric measures | |||
| Height (cm) | 175 | 7.59 | |
| Weight (kg) | 80.9 | 15.5 | |
| Hip (cm) | 98.6 | 10.1 | |
| Waist (cm) | 92.0 | 14.8 | |
| WHR | 0.93 | 0.07 | |
| BAI | 24.8 | 5.03 | |
| BMI (kg/m2) | 26.5 | 5.04 | |
| BF% | 24.5 | 8.13 | |
| Skinfolds (mm) | |||
| Bicipital | 7.27 | 4.52 | |
| Tricipital | 11.1 | 5.62 | |
| Subscapular | 14.1 | 6.59 | |
| Supraspinatus | 17.2 | 8.98 | |
| Abdominal | 21.7 | 9.84 | |
| Dietary parameters | Recommendation | ||
| Energy intake (kcal/day) | 2231 | 521 | 2000–2600 |
| Carbohydrate (g/day) | 237 (42.6%) | 80.5 | 45–60% * |
| Fat (g/day) | 92.1 (37.2%) | 31.5 | 20–35% * |
| Proteins (g/day) | 88.4 (15.9%) | 26.1 | 15–20% |
| Fiber (g/day) | 22.8 | 9.08 | > 25 g/day * |
| Vitamin A (µg/day) | 1113 | 3718 | 750 # |
| Genetic features | Ob-IB (%) | 1000genomes (%) | |
| rs5888 (SCARB1) | TT + TC | 66.5 | 70.6 |
| CC | 33.5 | 29.4 | |
| rs659366 (UCP2) | TT + TC | 65.2 | 61.0 |
| CC | 34.8 | 39.0 | |
| rs1800592 (UCP1) | AA | 51.3 | 58.1 |
| AG + GG | 48.7 | 41.9 | |
| Variables | Genotype VA Responsive (A) (n = 106) | p-Value | Genotype Less VA Responsive (B) (n = 52) | p-Value | GxVA Interaction (p-Value) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low VA Intake (LI) (n = 60) | Recommended VA Intake (RI) (n = 46) | Low VA Intake (LI) (n = 30) | Recommended VA Intake (RI) (n = 22) | ||||||||||||||||
| Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | ||||
| Weight (kg) | 81.8 | 17.3 | 49.5 | 140 | 79.3 | 14.1 | 47.9 | 122 | 0.352 | 81.2 | 15.3 | 60.0 | 133 | 81.1 | 14.1 | 61.9 | 127 | 0.611 | 0.383 |
| WHR | 0.93 | 0.08 | 0.70 | 1.12 | 0.92 | 0.08 | 0.72 | 1.11 | 0.430 | 0.95 | 0.08 | 0.81 | 1.12 | 0.91 | 0.05 | 0.85 | 1.01 | 0.466 | 0.905 |
| BMI (kg/m2) | 27.0 | 5.51 | 19.2 | 45.1 | 25.5 | 4.47 | 18.7 | 36.8 | 0.076 | 27.2 | 5.39 | 19.8 | 42.9 | 26.3 | 4.24 | 20.5 | 38.3 | 0.737 | 0.249 |
| BF% | 25.9 | 8.12 | 5.30 | 43.7 | 22.1 | 8.44 | 6.70 | 38.8 | 0.006 | 25.8 | 8.24 | 11.3 | 47.6 | 23.8 | 6.39 | 13.0 | 36.6 | 0.972 | 0.114 |
| BAI | 25.5 | 5.30 | 17.2 | 44.8 | 23.7 | 4.66 | 15.1 | 34.6 | 0.033 | 25.8 | 5.67 | 18.9 | 39.6 | 24.1 | 3.59 | 16.7 | 30.5 | 0.799 | 0.271 |
| Bicipital SF | 8.41 | 5.59 | 2.60 | 30.0 | 5.83 | 2.85 | 2.20 | 13.0 | 0.002 | 7.97 | 4.67 | 2.80 | 19.6 | 6.33 | 2.87 | 2.90 | 13.5 | 0.509 | 0.187 |
| Energy (kcal/day) | 2229 | 538 | 1253 | 374 | 2276 | 536 | 1384 | 4256 | 0.551 | 2058 | 456 | 1061 | 3132 | 2377 | 497 | 1615 | 3666 | 0.040 | 0.167 |
| Carbohydrate (% EC) | 43.7 | 10.5 | 21.8 | 73.7 | 41.1 | 9.64 | 20.5 | 57.6 | 0.194 | 43.2 | 9.18 | 30.0 | 64.3 | 42.1 | 10.2 | 19.9 | 63.3 | 0.508 | 0.739 |
| Fat (% EC) | 35.9 | 9.60 | 16.8 | 61.5 | 38.6 | 8.33 | 24.3 | 57.9 | 0.135 | 36.4 | 10.2 | 12.9 | 57.8 | 38.7 | 10.6 | 13.9 | 63.5 | 0.479 | 0.853 |
| Proteins (g/day) | 86.9 | 25.7 | 37.0 | 180 | 94.5 | 29.3 | 55.8 | 182 | 0.122 | 76.2 | 18.5 | 30.0 | 104 | 96.6 | 23.7 | 57.9 | 154 | 0.002 | 0.224 |
| Fiber (g/day) | 22.7 | 11.0 | 4.50 | 63.8 | 23.2 | 7.24 | 13.0 | 39.4 | 0.324 | 20.9 | 5.86 | 10.4 | 36.7 | 24.7 | 10.5 | 11.0 | 56.8 | 0.104 | 0.570 |
| Vitamin A (µg/day) | 458 | 148 | 110 | 744 | 2368 | 6762 | 751 | 45239 | 0.000 | 498 | 131 | 196 | 722 | 1118 | 342 | 764 | 1887 | 0.000 | 0.138 |
| Retinol (µg/day) | 238 | 115 | 2.10 | 527 | 1703 | 6829 | 22.3 | 45116 | 0.000 | 258 | 126 | 19.0 | 518 | 479 | 320 | 35.2 | 1625 | 0.020 | 0.558 |
| ß-carotene (µg/day) | 951 | 681 | 10.4 | 2999 | 3090 | 1930 | 15.6 | 9264 | 0.000 | 1140 | 788 | 120 | 3153 | 2795 | 1481 | 318 | 5166 | 0.000 | 0.682 |
| Genotype A | Beta | SE | p-Value |
| BF% | −4.11 | 1.47 | 0.006 |
| BAI | −1.99 | 0.92 | 0.033 |
| Genotype B | Beta | SE | p-Value |
| BF% | −0.10 | 1.75 | 0.972 |
| BAI | −0.28 | 1.10 | 0.799 |
| Variables | High Body Fat % (n = 21) | p-Value | Low Body Fat% (n = 20) | p-Value | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype A (n = 11) | Genotype B (n = 10) | Genotype A (n = 10) | Genotype B (n = 10) | |||||||||||||||
| Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | |||
| Weight (kg) | 93.4 | 23.0 | 63.6 | 152 | 82.0 | 10.8 | 68.8 | 106 | 0.179 | 68.7 | 9.92 | 52.7 | 85.0 | 74.0 | 8.14 | 63.3 | 86.7 | 0.176 |
| WHR | 0.93 | 0.10 | 0.78 | 1.11 | 0.85 | 0.09 | 0.66 | 1.01 | 0.077 | 0.87 | 0.09 | 0.78 | 1.11 | 0.83 | 0.06 | 0.73 | 0.90 | 0.249 |
| BMI (kg/m2) | 31.1 | 6.17 | 24.9 | 46.6 | 28.0 | 4.88 | 21.7 | 39.0 | 0.199 | 22.6 | 2.52 | 19.7 | 27.1 | 23.8 | 2.42 | 19.5 | 27.1 | 0.246 |
| BF% | 32.2 | 4.92 | 26.5 | 43.6 | 29.6 | 2.66 | 26.2 | 35.1 | 0.152 | 19.0 | 4.20 | 10.1 | 24.0 | 18.8 | 4.50 | 12.6 | 24.8 | 0.794 |
| Bicipital SF | 11.2 | 9.95 | 3.90 | 40.0 | 11.1 | 3.31 | 6.00 | 15.8 | 0.915 | 4.77 | 1.54 | 2.70 | 8.00 | 6.38 | 3.85 | 3.60 | 15.8 | 0.123 |
| Energy (kcal/day) | 2311 | 415 | 1880 | 3052 | 1860 | 212 | 1530 | 2130 | 0.007 | 2238 | 424 | 1253 | 2853 | 2047 | 276 | 1695 | 2500 | 0.226 |
| Carbohydrate (% EC) | 38.9 | 5.92 | 30.9 | 47.4 | 40.9 | 7.43 | 30.0 | 51.2 | 0.472 | 46.2 | 8.48 | 38.4 | 60.4 | 42.5 | 8.02 | 32.8 | 56.4 | 0.326 |
| Fat (% EC) | 40.7 | 4.72 | 33.8 | 47.8 | 35.6 | 9.81 | 12.9 | 46.4 | 0.190 | 33.9 | 7.41 | 20.9 | 46.0 | 39.0 | 8.33 | 25.1 | 48.3 | 0.208 |
| Proteins (g/day) | 94.1 | 21.4 | 54.1 | 122 | 72.1 | 11.6 | 44.5 | 85.4 | 0.014 | 80.2 | 20.9 | 53.6 | 112 | 82.0 | 10.3 | 66.1 | 94.30 | 0.922 |
| Fiber (g/day) | 19.9 | 5.78 | 14.7 | 35.5 | 19.4 | 5.53 | 8.10 | 25.7 | 0.599 | 21.6 | 7.51 | 12.0 | 32.0 | 18.6 | 5.75 | 10.1 | 25.50 | 0.511 |
| Vitamin A (µg/day) | 583 | 165 | 372 | 794 | 543 | 148 | 386 | 819 | 0.711 | 532 | 141 | 287 | 754 | 618 | 207 | 365 | 934 | 0.218 |
| Retinol (µg/day) | 231 | 140 | 59.9 | 429 | 254 | 121 | 28.3 | 397 | 0.227 | 250 | 86.4 | 132 | 436 | 280 | 106 | 111 | 497 | 0.553 |
| ß-carotene (µg/day) | 1612 | 1065 | 480 | 4300 | 1540 | 993 | 303 | 3450 | 0.791 | 1295 | 912 | 241 | 2999 | 1741 | 1160 | 308 | 4333 | 0.217 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galmés, S.; Palou, A.; Serra, F. Increased Risk of High Body Fat and Altered Lipid Metabolism Associated to Suboptimal Consumption of Vitamin A Is Modulated by Genetic Variants rs5888 (SCARB1), rs1800629 (UCP1) and rs659366 (UCP2). Nutrients 2020, 12, 2588. https://doi.org/10.3390/nu12092588
Galmés S, Palou A, Serra F. Increased Risk of High Body Fat and Altered Lipid Metabolism Associated to Suboptimal Consumption of Vitamin A Is Modulated by Genetic Variants rs5888 (SCARB1), rs1800629 (UCP1) and rs659366 (UCP2). Nutrients. 2020; 12(9):2588. https://doi.org/10.3390/nu12092588
Chicago/Turabian StyleGalmés, Sebastià, Andreu Palou, and Francisca Serra. 2020. "Increased Risk of High Body Fat and Altered Lipid Metabolism Associated to Suboptimal Consumption of Vitamin A Is Modulated by Genetic Variants rs5888 (SCARB1), rs1800629 (UCP1) and rs659366 (UCP2)" Nutrients 12, no. 9: 2588. https://doi.org/10.3390/nu12092588