Appendix B
Figure A1.
Flow chart of the study selection process: Our search strategy retrieved 17,329 unique hits, of which we included 111 studies reporting on amino acid (AA) supplementation in our systematic review. Of these, 63 studies reported on arginine supplementation, 11 reported on branched-chain amino acids (BCAA) supplementation, and 38 reported on methyl donor supplementation. We pooled data on the effect of arginine supplementation on birth weight (BW) in 57 studies, on maternal blood pressure (BP) in 15 studies, and on small for gestational (SGA) development in risk populations in 8 studies. We were able to pool data on the effect of BCAA on BW in 10 studies and of methyl donor supplementation in 36 studies. Adapted from PRISMA [
29].
Figure A1.
Flow chart of the study selection process: Our search strategy retrieved 17,329 unique hits, of which we included 111 studies reporting on amino acid (AA) supplementation in our systematic review. Of these, 63 studies reported on arginine supplementation, 11 reported on branched-chain amino acids (BCAA) supplementation, and 38 reported on methyl donor supplementation. We pooled data on the effect of arginine supplementation on birth weight (BW) in 57 studies, on maternal blood pressure (BP) in 15 studies, and on small for gestational (SGA) development in risk populations in 8 studies. We were able to pool data on the effect of BCAA on BW in 10 studies and of methyl donor supplementation in 36 studies. Adapted from PRISMA [
29].
Figure A2.
Influential case analysis of studies reporting on arginine family supplementation and fetal/birth weight: A sensitivity analysis revealed Sharkey et al. as an influential case [
51].
Figure A2.
Influential case analysis of studies reporting on arginine family supplementation and fetal/birth weight: A sensitivity analysis revealed Sharkey et al. as an influential case [
51].
Figure A3.
Funnel plot for amino acids of the arginine family and fetal/birth weight in all studies and in studies with only pregnancy complications: These funnel plots and Egger’s regression did not indicate publication bias in all studies or studies in complicated pregnancies. Black dots are the included studies. Sharkey et al., as an influential case, was highlighted by the colour red [
51].
Figure A3.
Funnel plot for amino acids of the arginine family and fetal/birth weight in all studies and in studies with only pregnancy complications: These funnel plots and Egger’s regression did not indicate publication bias in all studies or studies in complicated pregnancies. Black dots are the included studies. Sharkey et al., as an influential case, was highlighted by the colour red [
51].
Figure A4.
Meta-analysis on prenatal supplementation of arginine on maternal blood pressure: Blood pressure was unaffected in normal-growth pregnancies following arginine supplementation, but was reduced in the risk population and complicated pregnancies. The data is ordered within each amino acid (AA) from smallest to largest animal. Blood pressure difference (BP diff) data represent pooled estimates expressed as mean difference (MD) with 95% confidence interval (CI) using a random effect model. Residual is shown. FGR, fetal growth restriction; I2, heterogeneity; PE, preeclampsia; PIH, pregnancy-induced hypertension.
Figure A4.
Meta-analysis on prenatal supplementation of arginine on maternal blood pressure: Blood pressure was unaffected in normal-growth pregnancies following arginine supplementation, but was reduced in the risk population and complicated pregnancies. The data is ordered within each amino acid (AA) from smallest to largest animal. Blood pressure difference (BP diff) data represent pooled estimates expressed as mean difference (MD) with 95% confidence interval (CI) using a random effect model. Residual is shown. FGR, fetal growth restriction; I2, heterogeneity; PE, preeclampsia; PIH, pregnancy-induced hypertension.
Figure A5.
Species meta-regression of amino acids (AA) in arginine family on maternal blood pressure (BP) in pregnancy complications: Meta-regression revealed large interspecies differences, although only two human study cohorts versus seven rat study cohorts reported on the effect of prenatal supplementation of AA of the arginine family in complicated pregnancies. Data represent pooled estimates expressed as a mean difference (MD) with a 95% confidence interval (CI) using a random effect model. I2, heterogeneity.
Figure A5.
Species meta-regression of amino acids (AA) in arginine family on maternal blood pressure (BP) in pregnancy complications: Meta-regression revealed large interspecies differences, although only two human study cohorts versus seven rat study cohorts reported on the effect of prenatal supplementation of AA of the arginine family in complicated pregnancies. Data represent pooled estimates expressed as a mean difference (MD) with a 95% confidence interval (CI) using a random effect model. I2, heterogeneity.
Figure A6.
Dose–response curve of prenatal supplementation of arginine family on blood pressure (BP) in complicated pregnancies: Higher doses of arginine result in lower maternal blood pressure in complicated pregnancies (pslope = 0.0031). However, this dose–response relation is influenced by an interspecies difference as the higher doses are tested in animal studies and the lowest doses are tested in human studies. Animal models for pregnancy complication included adriamycin nephropathy-induced preeclampsia (A), spontaneous hypertension and heart failure (F), hyperinsulinemic-induced PIH/FGR (H), L-NAME-induced fetal growth restriction/preeclampsia (L), magnesium deficiency-induced fetal growth restriction (M), reduced uterine perfusion pressure-induced fetal growth restriction/preeclampsia (R), and sonic stress-induced preeclampsia (S). Daily dose is expressed as mg per kg metabolic body weight. Open dots indicate human studies, and closed dots indicate animal studies. The black line is drawn for all studies, the yellow line is for animal studies only, and no line is drawn for human studies since only 3 studies were available.
Figure A6.
Dose–response curve of prenatal supplementation of arginine family on blood pressure (BP) in complicated pregnancies: Higher doses of arginine result in lower maternal blood pressure in complicated pregnancies (pslope = 0.0031). However, this dose–response relation is influenced by an interspecies difference as the higher doses are tested in animal studies and the lowest doses are tested in human studies. Animal models for pregnancy complication included adriamycin nephropathy-induced preeclampsia (A), spontaneous hypertension and heart failure (F), hyperinsulinemic-induced PIH/FGR (H), L-NAME-induced fetal growth restriction/preeclampsia (L), magnesium deficiency-induced fetal growth restriction (M), reduced uterine perfusion pressure-induced fetal growth restriction/preeclampsia (R), and sonic stress-induced preeclampsia (S). Daily dose is expressed as mg per kg metabolic body weight. Open dots indicate human studies, and closed dots indicate animal studies. The black line is drawn for all studies, the yellow line is for animal studies only, and no line is drawn for human studies since only 3 studies were available.
![Nutrients 12 02535 g0a6]()
Figure A7.
Sensitivity analysis of studies reporting on arginine family supplementation and maternal blood pressure: The sensitivity analysis revealed no clear outlier in studies reporting on arginine family supplementation on maternal blood pressure.
Figure A7.
Sensitivity analysis of studies reporting on arginine family supplementation and maternal blood pressure: The sensitivity analysis revealed no clear outlier in studies reporting on arginine family supplementation on maternal blood pressure.
Figure A8.
Dose–response curve on the prenatal supplementation of arginine on development of small for gestational age (SGA): The available data points do not show a dose–response relation between odd ratio (OR) of development of SGA and daily arginine dose (pslope = 0.73) in a human risk population. Almost all studies clustered at the lower end of the dose spectrum.
Figure A8.
Dose–response curve on the prenatal supplementation of arginine on development of small for gestational age (SGA): The available data points do not show a dose–response relation between odd ratio (OR) of development of SGA and daily arginine dose (pslope = 0.73) in a human risk population. Almost all studies clustered at the lower end of the dose spectrum.
Figure A9.
Meta-analysis on prenatal supplementation of branched chain amino acid on fetal/birth weight (BW): Prenatal branched chain amino acid supplementation did not affect birth weight, neither in normal-growth pregnancies nor in complicated pregnancies. Data are ordered within each amino acid (AA) from smallest to largest animal. Data represent pooled estimates expressed as a ratio of means (ROM) with a 95% confidence interval (CI) using a random effect model. Residual I2 is shown. Some studies had multiple cohorts and are distinguishable in this figure by the following: * in this upper line, the daily dose is lower compared the next line(s) in increasing order; † the upper two lines are in the lower dose compared to the next two lines, and per dose, the outcomes are separately reported for males first and then females. FGR, fetal growth restriction; I2, heterogeneity.
Figure A9.
Meta-analysis on prenatal supplementation of branched chain amino acid on fetal/birth weight (BW): Prenatal branched chain amino acid supplementation did not affect birth weight, neither in normal-growth pregnancies nor in complicated pregnancies. Data are ordered within each amino acid (AA) from smallest to largest animal. Data represent pooled estimates expressed as a ratio of means (ROM) with a 95% confidence interval (CI) using a random effect model. Residual I2 is shown. Some studies had multiple cohorts and are distinguishable in this figure by the following: * in this upper line, the daily dose is lower compared the next line(s) in increasing order; † the upper two lines are in the lower dose compared to the next two lines, and per dose, the outcomes are separately reported for males first and then females. FGR, fetal growth restriction; I2, heterogeneity.
Figure A10.
Dose–response curve on prenatal supplementation of branched chain amino acid (BCAA) on fetal/birth weight in all pregnancies: There was a dose–response effect in which the highest doses resulted in a larger improvement of birth weight (pslope = 0.006). Only animal studies were included. Daily dose is expressed as mg per kg metabolic body weight.
Figure A10.
Dose–response curve on prenatal supplementation of branched chain amino acid (BCAA) on fetal/birth weight in all pregnancies: There was a dose–response effect in which the highest doses resulted in a larger improvement of birth weight (pslope = 0.006). Only animal studies were included. Daily dose is expressed as mg per kg metabolic body weight.
Figure A11.
Sensitivity analysis of studies reporting on branched-chain amino acids supplementation and fetal/birth weight: The sensitivity analysis revealed Viana et al. [
103] as an influential case.
Figure A11.
Sensitivity analysis of studies reporting on branched-chain amino acids supplementation and fetal/birth weight: The sensitivity analysis revealed Viana et al. [
103] as an influential case.
Figure A12.
Dose–response curve on prenatal supplementation of methyl donors on fetal/birth weight in all pregnancies: There was a dose–response relationship between birth weight ratio and daily dose of methyl donors (pslope = 0.0002). Excessive doses of methionine and cysteine resulted in lower birth weight (methionine pslope = 1.09 * 10–5; cysteine pslope = 0.16). Daily dose is expressed as mg per kg metabolic body weight. Open dots indicate human studies, and closed dots indicate animal studies.
Figure A12.
Dose–response curve on prenatal supplementation of methyl donors on fetal/birth weight in all pregnancies: There was a dose–response relationship between birth weight ratio and daily dose of methyl donors (pslope = 0.0002). Excessive doses of methionine and cysteine resulted in lower birth weight (methionine pslope = 1.09 * 10–5; cysteine pslope = 0.16). Daily dose is expressed as mg per kg metabolic body weight. Open dots indicate human studies, and closed dots indicate animal studies.
Figure A13.
Meta-regression of methyl donors on birth weight (BW): Meta-regression on (A) Amino acid (AA), (B) species, (C) pregnancy complication, (D) administration duration, (E) administration scheme, and (F) intervention type (prevention vs. treatment). No specific methyl donor was identified to be the most optimal. Methyl donor supplementation increased birth weight in human and preeclamptic studies (similar studies). Data represent pooled estimates expressed as a ratio of means (ROM) with a 95% confidence interval (CI) using a random effect model. FGR, fetal growth restriction; I2, heterogeneity; PE, preeclampsia; PIH, pregnancy-induced hypertension.
Figure A13.
Meta-regression of methyl donors on birth weight (BW): Meta-regression on (A) Amino acid (AA), (B) species, (C) pregnancy complication, (D) administration duration, (E) administration scheme, and (F) intervention type (prevention vs. treatment). No specific methyl donor was identified to be the most optimal. Methyl donor supplementation increased birth weight in human and preeclamptic studies (similar studies). Data represent pooled estimates expressed as a ratio of means (ROM) with a 95% confidence interval (CI) using a random effect model. FGR, fetal growth restriction; I2, heterogeneity; PE, preeclampsia; PIH, pregnancy-induced hypertension.
Figure A14.
Funnel plot for methyl donors and fetal/birth weight in all studies: The funnel plot and Eggers regression do not indicate publication bias in studies reporting the effect of methyl donor supplementation on fetal or birth weight. Dots are the included studies.
Figure A14.
Funnel plot for methyl donors and fetal/birth weight in all studies: The funnel plot and Eggers regression do not indicate publication bias in studies reporting the effect of methyl donor supplementation on fetal or birth weight. Dots are the included studies.
Figure A15.
Sensitivity analysis of studies reporting on methyl donor supplementation and fetal/birth weight: The sensitivity analysis revealed Mori et al. [
98] as a potential influential case.
Figure A15.
Sensitivity analysis of studies reporting on methyl donor supplementation and fetal/birth weight: The sensitivity analysis revealed Mori et al. [
98] as a potential influential case.
Figure A16.
Reporting quality and risk of bias in animal studies: Reporting of key indicators of study quality and risk of bias in animal studies was assessed for all items, but especially blinding, randomization, and sample size calculation scored as unreported, unclear, or high risk.
Figure A16.
Reporting quality and risk of bias in animal studies: Reporting of key indicators of study quality and risk of bias in animal studies was assessed for all items, but especially blinding, randomization, and sample size calculation scored as unreported, unclear, or high risk.
Figure A17.
Risk of bias in human studies: Risk of bias assessment in human studies appeared to be very unclear for most items.
Figure A17.
Risk of bias in human studies: Risk of bias assessment in human studies appeared to be very unclear for most items.
Figure A18.
Quality assessment of the included human studies by the Cochrane tool. Quality assessment using the Cochrane risk of bias tool. − = high risk of bias; + = low risk of bias; ? = unclear risk of bias.
Figure A18.
Quality assessment of the included human studies by the Cochrane tool. Quality assessment using the Cochrane risk of bias tool. − = high risk of bias; + = low risk of bias; ? = unclear risk of bias.
Table A1.
Search terms in Pubmed.
Table A1.
Search terms in Pubmed.
| 14 amino acids AND administration | (“Arginine/administration and dosage” [Mesh] OR “Arginine/adverse effects” [Mesh] OR “Arginine/deficiency” [Mesh] OR “Arginine/drug effects” [Mesh] OR “Arginine/physiology” [Mesh] OR “Arginine/pharmacology” [Mesh] OR “Arginine/therapeutic use” [Mesh] OR “Arginine/therapy” [Mesh] OR “Leucine/administration and dosage” [Mesh] OR “Leucine/adverse effects” [Mesh] OR “Leucine/deficiency” [Mesh] OR “Leucine/drug effects” [Mesh] OR “Leucine/physiology” [Mesh] OR “Leucine/pharmacology” [Mesh] OR “Leucine/therapeutic use” [Mesh] OR “Isoleucine/administration and dosage” [Mesh] OR “Isoleucine/adverse effects” [Mesh] OR “Isoleucine/deficiency” [Mesh] OR “Isoleucine/drug effects” [Mesh] OR “Isoleucine/physiology” [Mesh] OR “Isoleucine/pharmacology” [Mesh] OR “Isoleucine/therapeutic use” [Mesh] OR “Valine/administration and dosage” [Mesh] OR “Valine/adverse effects” [Mesh] OR “Valine/deficiency” [Mesh] OR “Valine/drug effects” [Mesh] OR “Valine/physiology” [Mesh] OR “Valine/pharmacology” [Mesh] OR “Valine/therapeutic use” [Mesh] OR “Valine/therapy” [Mesh] OR “Cysteine/administration and dosage” [Mesh] OR “Cysteine/adverse effects” [Mesh] OR “Cysteine/deficiency” [Mesh] OR “Cysteine/drug effects” [Mesh] OR “Cysteine/physiology” [Mesh] OR “Cysteine/pharmacology” [Mesh] OR “Cysteine/therapeutic use” [Mesh] OR “Cysteine/therapy” [Mesh] OR “Methionine/administration and dosage” [Mesh] OR “Methionine/adverse effects” [Mesh] OR “Methionine/deficiency” [Mesh] OR “Methionine/drug effects” [Mesh] OR “Methionine/physiology” [Mesh] OR “Methionine/pharmacology” [Mesh] OR “Methionine/therapeutic use” [Mesh] OR “Methionine/therapy” [Mesh] OR “Glutamic Acid/administration and dosage” [Mesh] OR “Glutamic Acid/adverse effects” [Mesh] OR “Glutamic Acid/deficiency” [Mesh] OR “Glutamic Acid/drug effects” [Mesh] OR “Glutamic Acid/pharmacology” [Mesh] OR “Glutamic Acid/physiology” [Mesh] OR “Glutamic Acid/therapeutic use” [Mesh] OR “Glutamine/administration and dosage” [Mesh] OR “Glutamine/adverse effects” [Mesh] OR “Glutamine/deficiency” [Mesh] OR “Glutamine/drug effects” [Mesh] OR “Glutamine/pharmacology” [Mesh] OR “Glutamine/physiology” [Mesh] OR “Glutamine/therapeutic use” [Mesh] OR “Glutamine/therapy” [Mesh] OR “Citrulline/administration and dosage” [Mesh] OR “Citrulline/adverse effects” [Mesh] OR “Citrulline/deficiency” [Mesh] OR “Citrulline/drug effects” [Mesh] OR “Citrulline/pharmacology” [Mesh] OR “Citrulline/physiology” [Mesh] OR “Citrulline/therapeutic use” [Mesh] OR “Asparagine/administration and dosage” [Mesh] OR “Asparagine/adverse effects” [Mesh] OR “Asparagine/deficiency” [Mesh] OR “Asparagine/drug effects” [Mesh] OR “Asparagine/pharmacology” [Mesh] OR “Asparagine/physiology” [Mesh] OR “Asparagine/therapeutic use” [Mesh] OR “Asparagine/therapy” [Mesh] OR “Asparagine/administration and dosage” [Mesh] OR “Asparagine/adverse effects” [Mesh] OR “Asparagine/deficiency” [Mesh] OR “Asparagine/drug effects” [Mesh] OR “Asparagine/pharmacology” [Mesh] OR “Asparagine/physiology” [Mesh] OR “Asparagine/therapeutic use” [Mesh] OR “Asparagine/therapy” [Mesh] OR “Aspartic Acid/administration and dosage” [Mesh] OR “Aspartic Acid/adverse effects” [Mesh] OR “Aspartic Acid/deficiency” [Mesh] OR “Aspartic Acid/drug effects” [Mesh] OR “Aspartic Acid/pharmacology” [Mesh] OR “Aspartic Acid/physiology” [Mesh] OR “Aspartic Acid/therapeutic use” [Mesh] OR “Aspartic Acid/therapy” [Mesh] OR “Proline/administration and dosage” [Mesh] OR “Proline/adverse effects” [Mesh] OR “Proline/deficiency” [Mesh] OR “Proline/drug effects” [Mesh] OR “Proline/pharmacology” [Mesh] OR “Proline/physiology” [Mesh] OR “Proline/therapeutic use” [Mesh] OR “Ornithine/administration and dosage” [Mesh] OR “Ornithine/adverse effects” [Mesh] OR “Ornithine/deficiency” [Mesh] OR “Ornithine/drug effects” [Mesh] OR “Ornithine/pharmacology” [Mesh] OR “Ornithine/physiology” [Mesh] OR “Ornithine/therapeutic use” [Mesh] OR “Choline/administration and dosage” [Mesh] OR “Choline/adverse effects” [Mesh] OR “Choline/deficiency” [Mesh] OR “Choline/drug effects” [Mesh] OR “Choline/pharmacology” [Mesh] OR “Choline/physiology” [Mesh] OR “Choline/therapeutic use” [Mesh] OR “Choline/therapy” [Mesh]) OR ((“Arginine” [Mesh] OR Arginine [tiab] OR L-Arginine [tiab] OR “Leucine” [Mesh] OR “Isoleucine” [Mesh] OR Leucine [tiab] OR L-Leucine [tiab] OR Leucin [tiab] OR Isoleucine [tiab] OR Isoleucin [tiab] OR Alloisoleucine [tiab] OR Alloisoleucin [tiab] OR “Valine” [Mesh] OR Valine [tiab] OR L-Valine [tiab] OR Valsartan [tiab] OR Valerate [tiab] OR “Cysteine” [Mesh] OR Cysteine [tiab] OR L-Cysteine [tiab] OR Cysteinate [tiab] OR Acetylcysteine [tiab] OR Carbocysteine [tiab] OR Cysteinyldopa [tiab] OR Cystine [tiab] OR cystein [tiab] OR cysthion [tiab] OR Selenocysteine [tiab] OR “Methionine” [Mesh] OR Methionine [tiab] OR L-Methionine [tiab] OR Liquimeth [tiab] OR Pedameth [tiab] OR Formylmethionine [tiab] OR Racemethionine [tiab] OR Adenosylmethionine [tiab] OR Selenomethionine [tiab] OR Vitamin U [tiab] OR acimetion [tiab] OR cotameth [tiab] OR lobamine [tiab] OR menin [tiab] OR menine [tiab] OR meonine [tiab] OR methiolate [tiab] OR methionin [tiab] OR methiotrans [tiab] OR methnine [tiab] OR methurine [tiab] OR metione [tiab] OR methidin [tiab] OR neutrodor [tiab] OR oradash [tiab] OR urosamine [tiab] OR Methyl Donor [tiab] OR “Glutamic Acid” [Mesh] OR glutamic acid [tiab] OR glutamate [tiab] OR MSG [tiab] OR vestin [tiab] OR “aminoglutaric acid” [tiab] OR “aminopentanedioic acid” [tiab] OR acidogen [tiab] OR acidoride [tiab] OR acidothym [tiab] OR acidulin [tiab] OR aciglumin [tiab] OR aciglut [tiab] OR aclor [tiab] OR antalka [tiab] OR flanithin [tiab] OR gastuloric [tiab] OR glusate [tiab] OR glutadox [tiab] OR glutamidin [tiab] OR “glutamin acid” [tiab] OR “glutaminic acid” [tiab] OR glutaminol [tiab] OR glutan [tiab] OR glutansin [tiab] OR glutasin [tiab] OR glutaton [tiab] OR hydrionic [tiab] OR hypochylin [tiab] OR levoglutamate [tiab] OR “levoglutamic acid” [tiab] OR muriamic [tiab] OR pepsdol [tiab] OR “Glutamine” [Mesh:NoExp] OR glutamine [tiab] OR “aminoglutaramic acid” [tiab] OR acutil [tiab] OR “adamin G” [tiab] OR glumin [tiab] OR glutamin [tiab] OR levoglutamide [tiab] OR levoglutamine [tiab] OR nutrestore [tiab] OR “Citrulline” [Mesh] OR citrulline [tiab] OR “ureidopentanoic acid” [tiab] OR ureidonorvaline [tiab] OR “ureidovaleric acid” [tiab] OR citrullin [tiab] OR carbamylornithine [tiab] OR “Asparagine” [Mesh] OR asparagine [tiab] OR asparagin [tiab] OR “aminosuccinamic acid” [tiab] OR “Aspartic Acid” [Mesh:NoExp] OR “D-Aspartic Acid” [Mesh] OR “Potassium Magnesium Aspartate” [Mesh] OR “aspartic acid” [tiab] OR aspartate [tiab] OR Magnesiocard [tiab] OR Mg-5-Longoral [tiab] OR Mg 5 Longoral [tiab] OR Mg5 Longoral [tiab] OR panangin [tiab] OR astra 2045 [tiab] OR “aminosuccinic acid” [tiab] OR “asparagic acid” [tiab] OR asparaginate [tiab] OR “asparaginic acid” [tiab] OR aspartyl [tiab] OR aspatofort [tiab] OR “levoaspartic acid” [tiab] OR “Proline” [Mesh:NoExp] OR proline [tiab] OR prolin [tiab] OR levoproline [tiab] OR “pyrrolidinecarboxylic acid” [tiab] OR pyrrolidine carboxylate [tiab] OR “Ornithine” [Mesh:NoExp] OR ornithine [tiab] OR ornithin [tiab] OR “Diaminopentanoic Acid” [tiab] OR “diaminovaleric acid” [tiab] OR “Choline” [Mesh:NoExp] OR choline [tiab] OR bursine [tiab] OR vidine [tiab] OR fagine [tiab] OR trimethylammonium hydroxide [tiab] OR amonita [tiab] OR bilineurine [tiab] OR biocholine [tiab] OR biocolina [tiab] OR cholin [tiab] OR hepacholine [tiab] OR laevocholine [tiab] OR levocholine [tiab] OR lipotril [tiab] OR luridine [tiab] OR sincaline [tiab] OR urocholine [tiab]) AND (“Dietary Supplements” [Mesh] OR “Administration, Oral” [Mesh] OR administration * [tiab] OR administer * [tiab] OR dose [tiab] OR doses [tiab] OR dosage [tiab] OR treatment [tiab] OR treated [tiab] OR supplement * [tiab] OR diet [tiab] OR diets [tiab] OR dietary [tiab] OR intake [tiab] OR intakes [tiab] OR consumption [tiab] OR consumptions [tiab] OR consume [tiab] OR nutraceutical * [tiab] OR nutriceutical * [tiab] OR therapy [tiab] OR therapies [tiab])) |
| AND | AND |
| Healthy pregnancy OR Complicated pregnancy | “Pregnancy” [Mesh] OR “gravidity” [Mesh] OR “Fetus” [Mesh] OR Pregnancy [tiab] OR Pregnancies [tiab] OR Pregnant [tiab] OR Gestation [tiab] OR Gestations [tiab] OR Gestational [tiab] OR gravidity [tiab] OR gravidities [tiab] OR gravid [tiab] OR fetus [tiab] OR foetus [tiab] OR fetal [tiab] OR foetal [tiab] OR childbearing [tiab] OR “child bearing” [tiab] OR “Fetal Growth Retardation” [Mesh] OR “Infant, Low Birth Weight “ [Mesh] OR “Infant, Premature” [Mesh] OR “Premature Birth” [Mesh] OR FGR [tiab] OR Intrauterine growth retardation [tiab] OR Intra-uterine growth retardation [tiab] OR Intrauterine growth restriction [tiab] OR Intra-uterine growth restriction [tiab] OR IUGR [tiab] OR Small for Gestational Age [tiab] OR SGA [tiab] OR Low birth weight [tiab] OR Premature baby [tiab] OR Pre-mature baby [tiab] OR Preterm baby [tiab] OR Pre-term baby [tiab] OR Premature babies [tiab] OR Pre-mature babies [tiab] OR Preterm babies [tiab] OR Pre-term babies [tiab] OR Premature child [tiab] OR Pre-mature child [tiab] OR Preterm child [tiab] OR Pre-term child [tiab] OR Premature children [tiab] OR Pre-mature children [tiab] OR Preterm children [tiab] OR Pre-term children [tiab] OR Premature infant * [tiab] OR Pre-mature infant * [tiab] OR Preterm infant * [tiab] OR Pre-term infant * [tiab] OR Premature newborn * [tiab] OR Pre-mature newborn * [tiab] OR Preterm newborn * [tiab] OR Pre-term newborn * [tiab] OR Premature neonate * [tiab] OR Pre-mature neonate * [tiab] OR Preterm neonate * [tiab] OR Pre-term neonate * [tiab] OR Premature birth * [tiab] OR Pre-mature birth * [tiab] OR Preterm birth * [tiab] OR Pre-term birth * [tiab] OR Neonatal prematurity [tiab] OR prematuritas [tiab] OR “Hypertension, Pregnancy-Induced” [Mesh] OR Pre-Eclampsia [tiab] OR Preeclampsia [tiab] OR pre-eclamptic [tiab] OR preeclamptic [tiab] OR preclampsia [tiab] OR Proteinuria Edema Hypertension Gestosis [tiab] OR Edema Proteinuria Hypertension Gestosis [tiab] OR EPH Gestosis [tiab] OR EPH Toxemia * [tiab] OR EPH Complex [tiab] OR “Placental Insufficiency” [Mesh] OR “placenta insufficiency” [tiab] OR “placental insufficiency” [tiab] OR “placenta insufficiencies” [tiab] OR “placental insufficiencies” [tiab] OR “placenta deficiency” [tiab] OR “placental deficiency” [tiab] OR “placenta deficiencies” [tiab] OR “placental deficiencies” [tiab] OR “placenta failure” [tiab] OR “placental failure” [tiab] OR “Fetal Macrosomia” [Mesh] OR Macrosomia * [tiab] OR high birth weight [tiab] OR “overweight infant” [tiab] OR “overweight infants” [tiab] OR “overweight newborn” [tiab] OR “overweight newborns” [tiab] OR “overweight neonate” [tiab] OR “overweight neonates” [tiab] OR “Diabetes, Gestational” [Mesh] OR “Pregnancy in Diabetics” [Mesh] OR diabetes gravidarum [tiab] |
Table A2.
Search terms in Embase.
Table A2.
Search terms in Embase.
| 14 amino acids AND administration | (exp Arginine/OR (Arginine OR L-Arginine).ti,ab.) OR (exp Leucine/OR exp Isoleucine/OR (Leucine OR L-Leucine OR Leucin OR Isoleucine OR Isoleucin OR Alloisoleucine OR Alloisoleucin).ti,ab.) OR (exp Valine/OR (Valine OR L-Valine OR Valsartan OR Valerate).ti,ab.) OR (exp Cysteine/OR (Cysteine OR L-Cysteine OR Cysteinate OR Acetylcysteine OR Carbocysteine OR Cysteinyldopa OR Cystine OR cystein OR cysthion OR Selenocysteine).ti,ab,kw.) OR (exp Methionine/OR (Methionine OR L-Methionine OR Liquimeth OR Pedameth OR Formylmethionine OR Racemethionine OR Adenosylmethionine OR Selenomethionine OR Vitamin U OR acimetion OR cotameth OR lobamine OR menin OR menine OR meonine OR methiolate OR methionin OR methiotrans OR methnine OR methurine OR metione OR methidin OR neutrodor OR oradash OR urosamine OR Methyl Donor).ti,ab,kw.) OR (exp glutamic acid/OR (glutamic acid OR glutamate OR MSG OR vestin OR aminoglutaric acid OR aminopentanedioic acid OR acidogen OR acidoride OR acidothym OR acidulin OR aciglumin OR aciglut OR aclor OR antalka OR flanithin OR gastuloric OR glusate OR glutadox OR glutamidin OR glutamin acid OR glutaminic acid OR glutaminol OR glutan OR glutansin OR glutasin OR glutaton OR hydrionic OR hypochylin OR levoglutamate OR levoglutamic acid OR muriamic OR pepsdol).ti,ab,kw.) OR (exp glutamine/OR (glutamine OR aminoglutaramic acid OR acutil OR adamin G OR glumin OR glutamin OR levoglutamide OR levoglutamine OR nutrestore).ti,ab,kw.) OR (exp Citrulline/OR (citrulline OR ureidopentanoic acid OR ureidonorvaline OR ureidovaleric acid OR citrullin OR carbamylornithine).ti,ab,kw.) OR (exp asparagine/OR (asparagine OR asparagin OR aminosuccinamic acid).ti,ab,kw.) OR (exp aspartic acid/OR (aspartic acid OR aspartate OR Magnesiocard OR Mg-5-Longoral OR Mg 5 Longoral OR Mg5 Longoral OR panangin OR astra 2045 OR aminosuccinic acid OR asparagic acid OR asparaginate OR asparaginic acid OR aspartyl OR aspatofort OR levoaspartic acid).ti,ab,kw.) OR (exp Proline/OR (proline OR prolin OR levoproline OR pyrrolidinecarboxylic acid OR pyrrolidine carboxylate).ti,ab,kw.) OR (exp ornithine/OR (ornithine OR ornithin OR Diaminopentanoic Acid OR diaminovaleric acid).ti,ab,kw.) OR (exp Choline/OR (choline OR bursine OR vidine OR fagine OR trimethylammonium hydroxide OR amonita OR bilineurine OR biocholine OR biocolina OR cholin OR hepacholine OR laevocholine OR levocholine OR lipotril OR luridine OR sincaline OR urocholine).ti,ab,kw.) AND (exp dietary supplement/OR exp nutrition supplement/OR (administration * OR administer * OR dose OR doses OR dosage OR treatment OR treated OR supplement * OR diet OR diets OR dietary OR intake OR intakes OR consumption OR consumptions OR consume OR nutraceutical * OR nutriceutical * OR therapy OR therapies).ti,ab,kw.) |
| AND | AND |
| Healthy pregnancy OR Complicated pregnancy | (exp pregnancy/OR exp fetus/OR (Pregnancy OR Pregnancies OR Pregnant OR Gestation OR Gestations OR Gestational OR gravidity OR gravidities OR gravid OR fetus OR foetus OR fetal OR foetal OR childbearing OR child bearing).ti,ab,kw.) OR (exp intrauterine growth retardation/OR exp Low Birth Weight/OR exp prematurity/OR (FGR OR Intrauterine growth retardation OR Intra-uterine growth retardation OR Intrauterine growth restriction OR Intra-uterine growth restriction OR IUGR OR Small for Gestational Age OR SGA OR Low birth weight OR Premature baby OR Pre-mature baby OR Preterm baby OR Pre-term baby OR Premature babies OR Pre-mature babies OR Preterm babies OR Pre-term babies OR Premature child OR Pre-mature child OR Preterm child OR Pre-term child OR Premature children OR Pre-mature children OR Preterm children OR Pre-term children OR Premature infant OR Premature infants OR Pre-mature infant OR Pre-mature infants OR Preterm infant OR Preterm infants OR Pre-term infant OR Pre-term infants OR Premature newborn OR Premature newborns OR Pre-mature newborn OR Pre-mature newborns OR Preterm newborn OR Preterm newborns OR Pre-term newborn OR Pre-term newborns OR Premature neonate OR Premature neonates OR Pre-mature neonate OR Pre-mature neonates OR Preterm neonate OR Preterm neonates OR Pre-term neonate OR Pre-term neonates OR Premature birth OR Premature births OR Pre-mature birth OR Pre-mature births OR Preterm birth OR Preterm births OR Pre-term birth OR Pre-term births OR Neonatal prematurity OR prematuritas).ti,ab,kw.) OR (exp maternal hypertension/OR (Pre-Eclampsia OR Preeclampsia OR pre-eclamptic OR preeclamptic OR preclampsia OR Proteinuria Edema Hypertension Gestosis OR Edema Proteinuria Hypertension Gestosis OR EPH Gestosis OR EPH Toxemia * OR EPH Complex).ti,ab,kw.) OR (exp Placenta Insufficiency/OR (placenta insufficiency OR placental insufficiency OR placenta insufficiencies OR placental insufficiencies OR placenta deficiency OR placental deficiency OR placenta deficiencies OR placental deficiencies OR placenta failure OR placental failure).ti,ab,kw.) OR (exp Macrosomia/OR (Macrosomia * OR high birth weight OR overweight infant OR overweight infants OR overweight newborn OR overweight newborns OR overweight neonate OR overweight neonates).ti,ab,kw.) OR (exp pregnancy diabetes mellitus/OR diabetes gravidarum.ti,ab,kw) |
Table A3.
Search terms in Cochrane.
Table A3.
Search terms in Cochrane.
| 14 amino acids AND administration | (MeSH descriptor: [Glutamine] explode all trees and with qualifier(s): [Administration & dosage-AD, Adverse effects-AE, Deficiency-DF, Drug effects-DE, Pharmacology-PD, Physiology-PH, Therapeutic use-TU] OR MeSH descriptor: [Glutamic Acid] explode all trees OR MeSH descriptor: [Citrulline] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects-AE, Deficiency—DF, Drug effects—DE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Asparagine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Drug effects—DE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Aspartic Acid] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Drug effects—DE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Proline] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Drug effects—DE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Ornithine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Drug effects—DE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Choline] explode all trees and with qualifier(s): Administration & dosage—AD, Adverse effects—AE, Pharmacology—PD, Physiology—PH, Therapeutic use—TU]MeSH descriptor: [Arginine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Leucine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Isoleucine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Valine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Cysteine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU] OR MeSH descriptor: [Methionine] explode all trees and with qualifier(s): [Administration & dosage—AD, Adverse effects—AE, Deficiency—DF, Pharmacology—PD, Physiology—PH, Therapeutic use—TU]) OR ((MeSH descriptor: [Dietary Supplements] explode all trees OR MeSH descriptor: [Administration, Oral] explode all trees OR administration * OR administer * OR dose OR doses OR dosage OR treatment OR treated OR supplement * OR diet OR diets OR dietary OR intake OR intakes OR consumption OR consumptions OR consume OR nutraceutical * OR nutriceutical * OR therapy OR therapies:ti,ab,kw ) AND (MeSH descriptor: [Glutamic Acid] explode all trees OR MeSH descriptor: [Glutamine] this term only OR MeSH descriptor: [Citrulline] explode all trees OR MeSH descriptor: [Asparagine] explode all trees OR MeSH descriptor: [Aspartic Acid] this term only OR MeSH descriptor: [D-Aspartic Acid] explode all trees OR MeSH descriptor: [Proline] this term only OR MeSH descriptor: [Choline] this term only OR MeSH descriptor: [Ornithine] this term only OR glutamic acid OR glutamate OR MSG OR vestin OR “aminoglutaric acid” OR “aminopentanedioic acid” OR acidogen or acidoride OR acidothym OR acidulin OR aciglumin OR aciglut OR aclor OR antalka OR flanithin OR gastuloric OR glusate OR glutadox OR glutamidin OR “glutamin acid” OR “glutaminic acid” OR glutaminol OR glutan OR glutansin OR glutasin OR glutaton OR hydrionic OR hypochylin OR levoglutamate OR “levoglutamic acid” OR muriamic OR pepsdol or glutamine OR “aminoglutaramic acid” or acutil OR “adamin G” OR glumin OR glutamin OR levoglutamide OR levoglutamine OR nutrestore or citrulline OR “ureidopentanoic acid” OR ureidonorvaline OR “ureidovaleric acid” OR citrullin OR carbamylornithine OR asparagine OR asparagin OR “aminosuccinamic acid” OR “aspartic acid” OR aspartate OR Magnesiocard OR Mg-5-Longoral OR Mg 5 Longoral OR Mg5 Longoral OR panangin OR astra 2045 OR “aminosuccinic acid” OR “asparagic acid” OR asparaginate OR “asparaginic acid” OR aspartyl OR aspatofort OR “levoaspartic acid” OR proline OR prolin OR levoproline OR “pyrrolidinecarboxylic acid”OR pyrrolidine carboxylate OR ornithine OR ornithin OR “Diaminopentanoic Acid” OR “diaminovaleric acid” OR choline OR bursine OR vidine OR fagine OR trimethylammonium hydroxide OR amonita OR bilineurine OR biocholine OR biocolina OR cholin OR hepacholine OR laevocholine OR levocholine OR lipotril OR luridine OR sincaline OR urocholine:ti,ab,kw OR MeSH descriptor: [Arginine] explode all trees OR MeSH descriptor: [Leucine] explode all trees OR MeSH descriptor: [Isoleucine] explode all trees OR MeSH descriptor: [Valine] explode all trees OR MeSH descriptor: [Cysteine] explode all trees OR MeSH descriptor: [Methionine] explode all trees OR Arginine OR L-Arginine OR Leucine OR L-Leucine OR Leucin OR Isoleucine OR Isoleucin OR Alloisoleucine OR Alloisoleucin OR Valine or L-Valine OR Valsartan OR Valerate OR Cysteine OR L-Cysteine OR Cysteinate OR Acetylcysteine OR Carbocysteine OR Cysteinyldopa OR Cystine OR cystein OR cysthion OR Selenocysteine OR Methionine OR L-Methionine OR Liquimeth OR Pedameth OR formylmethionine OR Racemethionine OR Adenosylmethionine OR Selenomethionine OR Vitamin U OR acimetion OR cotameth OR lobamine OR menin OR menine OR meonine OR methiolate OR methionin OR methiotrans OR methnine OR methurine OR metione OR methidin OR neutrodor OR oradash OR urosamine OR Methyl Donor:ti,ab,kw)) |
| AND | AND |
| Healthy pregnancy OR Complicated pregnancy | MeSH descriptor: [Pregnancy] explode all trees OR MeSH descriptor: [Gravidity] explode all trees OR MeSH descriptor: [Fetus] explode all trees OR MeSH descriptor: [Infant, Low Birth Weight] explode all trees OR MeSH descriptor: [Infant, Premature] explode all trees OR MeSH descriptor: [Premature Birth] explode all trees OR MeSH descriptor: [Hypertension, Pregnancy-Induced] explode all trees OR MeSH descriptor: [Fetal Macrosomia] explode all trees OR MeSH descriptor: [Diabetes, Gestational] explode all trees OR MeSH descriptor: [Pregnancy in Diabetics] explode all trees OR MeSH descriptor: [Placental Insufficiency] explode all trees OR Pregnancy OR Pregnancies OR Pregnant OR Gestation OR Gestations OR Gestational OR gravidity OR gravidities OR gravid or fetus OR foetus OR fetal OR foetal OR childbearing OR “child bearing” OR FGR OR Intrauterine growth retardation OR Intra-uterine growth retardation OR Intrauterine growth restriction OR Intra-uterine growth restriction OR IUGR OR Small for Gestational Age OR SGA OR Low birth weight OR Premature baby OR Pre-mature baby OR Preterm baby OR Pre-term baby OR Premature babies OR Pre-mature babies OR Preterm babies OR Pre-term babies OR Premature child OR Pre-mature child OR Preterm child OR Pre-term child OR Premature children OR Pre-mature children or Preterm children OR Pre-term children OR Premature infant * OR Pre-mature infant * OR Preterm infant * OR Pre-term infant * OR Premature newborn * OR Pre-mature newborn * OR Preterm newborn * OR Pre-term newborn * OR Premature neonate * OR Pre-mature neonate * OR Preterm neonate * OR Pre-term neonate * OR Premature birth * OR Pre-mature birth * PR Preterm birth * OR Pre-term birth * OR Neonatal prematurity OR prematuritas OR Pre-Eclampsia OR Preeclampsia OR pre-eclamptic OR preeclamptic OR preclampsia OR Proteinuria Edema Hypertension Gestosis OR Edema Proteinuria Hypertension Gestosis OR EPH Gestosis or EPH Toxemia * OR EPH Complex OR “placenta insufficiency” OR “placental insufficiency” OR “placenta insufficiencies” OR “placental insufficiencies” OR “placenta deficiency” OR “placental deficiency” OR “placenta deficiencies” OR “placental deficiencies” OR “placenta failure” OR “placental failure” OR Macrosomia * OR high birth weight OR “overweight infant” OR “overweight infants” OR “overweight newborn” OR “overweight newborns” OR “overweight neonate” OR “overweight neonates” OR diabetes gravidarum:ti,ab,kw |
Table A4.
Main study characteristics and reported outcome of included studies.
Table A4.
Main study characteristics and reported outcome of included studies.
| Author (Year) | Species | Animal Model | Pregnancy Complication | Dose (g/kg/day) | Fetal/Birth Weight | Maternal BP | Glucose | GWG | Dev. Compl |
|---|
| Arginine | | | | | | | | | |
| Greene (2012) [33] | Mouse | Control | Normal | 2.664 a | X | | | X | |
| Alexander (2004) [34] | Rat | RUPP | FGR/PE | 2.000 a | X | X | | | |
| | | Control | Normal | 2.000 a | X | X | | | |
| Altun (2008) [35] | Rat | Sonic stress-induction | PE | 0.021 | X | X | | | |
| Bourdon (2016) [44] | Rat | Protein restriction | FGR | 1.500 | X | | | X | |
| Bursztyn (2003) [45] | Rat | Hyperinsulinemia | FGR/PIH | 0.217 | X | X | X | X | |
| Chin (1991) [46] | Rat | Control | Normal | 0.159 | X | | | X | |
| Da Costa (2014) [47] | Rat | Control | Normal | 0.021 | X | | | | |
| Helmbrecht (1996) [48] | Rat | L-NAME | FGR/PE | 0.021 | X | X | | | |
| | | Control | Normal | 0.021 | X | X | | | |
| Podjarny (1993) [90] | Rat | Control | Normal | 0.322 | | X | | X | |
| | | Adriamycin nephropathy | PE | 0.322 | | X | | X | |
| Podjarny (1997) [92] | Rat | Adriamycin nephropathy | PE | 0.230 | | X | | | |
| Podjarny (2001a) [49] | Rat | Hyperinsulinemia | PIH | 0.220 | X | X | X | X | |
| Podjarny (2001b) [91] | Rat | Control | Normal | 0.260 | | X | | X | |
| | | L-NAME | FGR/PE | 0.260 | | X | | X | |
| Schooley (2002) [50] | Rat | Mg deficiency | FGR | 1.000 | X | X | | | |
| | | Control | No | 1.000 | X | X | | | |
| Sharkey (2001) [51] | Rat | Genetic | FGR/PIH | 0.100 a,b | X | X | | | |
| Vosatka (1998) [36] | Rat | Hypobaric hypoxia | FGR | 0.200 a | X | | | | |
| | | | | 2.000 a | X | | | X | |
| Zeng (2008) [37] | Rat | Control | Normal | 0.216 | X | | | X | |
| | | | | | X | | | X | |
| Crane (2016) [52] | Sheep | Control | Normal | 0.150 | X | | | | |
| Sun (2017) [54] | Sheep | Diet restriction twin bearing | FGR | 0.250 | X | | | X | |
| Sun (2018) [55] | Sheep | Diet restriction twin bearing | FGR | 0.249 | X | | | X | |
| Zhang (2016a) [56] | Sheep | Diet restriction twin bearing | FGR | 0.250 | X | | | | |
| Zhang (2016b) [57] | Sheep | Diet restriction twin bearing | FGR | 0.250 | X | | X | X | |
| Peine (2018) [53] | Sheep | Diet restriction | FGR | 0.180 | X | | | X | |
| Bass (2017) [58] | Pig | Control | Normal | 0.150 | X | | | X | |
| | | | | 0.170 | X | | | X | |
| Bérard (2010) [59] | Pig | Control | Normal | 0.160 | X | | | X | |
| Che (2013) [70] | Pig | Control | Normal | 0.082 a | X | | | | |
| Dallanora (2017) [69] | Pig | Control | Normal | 0.115 | X | | | X | |
| Gao (2012) [71] | Pig | Control | Normal | 0.140 a | X | | | | |
| Garbossa (2015) [72] | Pig | Control | Normal | 0.113 a | X | | | | |
| | | Ractopamine | Over-growth | 0.113 a | X | | | | |
| Guo (2016) [73] | Pig | Control | Normal | 0.015 a | X | | | | |
| Li (2010) [74] | Pig | Control | Normal | 0.070 | X | | X | X | |
| | | | | 0.140 | X | | X | X | |
| Li (2014) [75] | Pig | Control | Normal | 0.070 | X | | | X | |
| | | | | 0.140 | X | | | X | |
| Li (2015) [76] | Pig | Control | Normal | 0.210 | X | | | X | |
| | | | | 0.140 | X | | | X | |
| | | | | 0.130 | X | | | X | |
| Liu (2012) [77] | Pig | Control | Normal | 0.107 | X | | X | | |
| Madsen (2017) [60] | Pig | UOL | Normal | 0.157 a | X | | | | |
| | | Intact (relatively crowded) | FGR | 0.157 a | X | | | | |
| Mateo (2007) [61] | Pig | Control | Normal | 0.120 | X | | | | |
| Mateo (2008) [62] | Pig | Control | Normal | 0.120 | X | | | | |
| Wu (2012) [63] | Pig | Control | Normal | 0.110 | X | | | | |
| Quesnel (2014) [64] | Pig | Control | Normal | 0.110 | X | | | | |
| Camarena Pulido (2016) [78] | Human | Increased risk of PE | Risk | 0.042 a | X | X | | | X |
| Dera (2007) [95] | Human | FGR < p10; gestational HTN | Risk | 0.043 a | | | | X | X |
| Facchinetti (2007) [79] | Human | HTN after 20th wk | Risk | 0.110 a | X | | | | X |
| Hladenuwich (2006) [82] | Human | PE | PE | 0.200 a | X | | | | |
| Neri (2010) [83] | Human | Chronic HTN | Risk | 0.047 a | X | X | | | X |
| Ropacka (2007) [84] | Human | EFW < p10 | FGR | 0.043 a | X | | | | |
| Rytlewski (2005) [93] | Human | PE; BW < p10 | FGR/PE | 0.040 | | X | | | |
| Rytlewski (2006) [85] | Human | PE; BW < p10 | FGR/PE | 0.041 | X | | | | X |
| Rytlewski (2008) [86] | Human | Preterm labor sign | Preterm | 0.041 a | X | | | | X |
| Sieroszewski (2004) [87] | Human | EFW < p10 | FGR | 0.040 a | X | | | | X |
| Singh (2015) [88] | Human | EFW < p10 | FGR | 0.065 a | X | | | | |
| Staff (2004) [89] | Human | PE | FGR/PE | 0.150 a | X | X | | | |
| Valdivia-silva (2009) [96] | Human | PE | PE | 0.039 | | | | | X |
| Winer (2009) [80] | Human | AC < p3 and abnormal UA flow | FGR | 0.190 | X | X | | | |
| Zhang (2007) [81] | Human | PIH and FGR | FGR/PIH | 0.333 | X | | | | |
| Citrulline | | | | | | | | | |
| Bourdon (2016) [44] | Rat | Protein restriction | FGR | 2.000 | X | X | | X | |
| Koeners (2017) [38] | Rat | Pre-existent HTN | PIH | 0.438 b | X | | | | |
| Tran (2017) [39] | Rat | Protein restriction | FGR | 2.000 | X | | | | |
| Powers (2015) [94] | Human | Obese | Normal | 0.035 a | | X | | | |
| Glutamate | | | | | | | | | |
| Beaudoin (1981) [40] | Rat | Control | Normal | 2.600 a | X | | | | |
| Navarro (2005) [148] | Rat | Control | Normal | 0.110 | | | | X | |
| Zeng (2012) [41] | Rat | Control | Normal | 0.074 b | X | | | | |
| | | | | 0.149 b | X | | | | |
| Sun (2017) [54] | Sheep | Diet restriction twin bearing | FGR | 0.060 | X | | | X | |
| Sun (2018) [55] | Sheep | Diet restriction twin bearing | FGR | 0.062 | X | | | X | |
| Zhang (2016a) [56] | Sheep | Diet restriction twin bearing | FGR | 0.060 | X | | | | |
| Zhang (2016b) [57] | Sheep | Diet restriction twin bearing | FGR | 0.060 | X | | X | | |
| Liu (2012) [77] | Pig | Control | Normal | 0.011 | X | | X | | |
| Wu (2012) [63] | Pig | Control | Normal | 0.011 | X | | | | |
| Zhang (2014) [65] | Pig | Control | Normal | 0.007 | X | | | | |
| | | | | 0.015 | X | | | | |
| | | | | 0.022 | X | | | | |
| | | | | 0.030 | X | | | | |
| Zhu (2015) [149] | Pig | Diet restriction twin bearing | FGR | 0.060 | | | | X | |
| Cai (2018) [66] | Swine | Control | Normal | 0.003 | X | | | X | |
| | | | | | X | | | X | |
| | | | | | X | | | X | |
| Glutamine | | | | | | | | | |
| Caporossi (2014) [42] | Rat | Control | Normal | 2.000 | X | | | | |
| Zhu (2018) [67] | Pig | Control | Normal | 0.126 | X | | X | X | |
| Proline | | | | | | | | | |
| Gonzalez-Anover (2017) [68] | Pig | Control | Normal | 0.120 a | X | | | | |
| Aspartic acid | | | | | | | | | |
| Brunner (1978) [43] | Rat | PKU-induction | FGR | 1.570 b | X | | | X | |
| | | Control | Normal | 1.570 b | X | | | X | |
| Valine | | | | | | | | | |
| Brunner (1978) [43] | Rat | PKU-induction | FGR | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| | | Control | Normal | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| Matsueda (1982) [97] | Rat | Control | Normal | 2.710 | X | | | X | |
| Mori (1999) [98] | Rat | Control | Normal | 2.368 | X | | | X | |
| Xu (2017) [104] | Pig | Control | Normal | 0.037 | X | | | | |
| | | | | 0.087 | X | | | | |
| Leucine | | | | | | | | | |
| Viana (2013) [103] | Mouse | Control | Normal | 3.000 a,b | X | | | | |
| Brunner (1978) [43] | Rat | PKU-induction | FGR | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| | | Control | Normal | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| Cruz (2014) [99] | Rat | Control | Normal | NA | X | | | | |
| Cruz (2016) [100] | Rat | Control | Normal | NA | X | | | | |
| Matsueda (1982) [97] | Rat | Control | Normal | 1.500 | X | | | X | |
| Mawatari (2004) [102] | Rat | Control | Normal | 0.300 | X | | | X | |
| | | Control | Normal | 1.000 | X | | | X | |
| Mori (1999) [98] | Rat | Control | Normal | 2.368 | X | | | X | |
| Ventrucci (2001) [101] | Rat | Control | Normal | 7.180 b | X | | | X | |
| Ventrucci (2002) [150] | Rat | Control | Normal | 7.180 b | | | X | | |
| Wang (2018) [105] | Pig | Control | Normal | 0.046 | X | | | | |
| | | | | 0.092 | X | | | | |
| | | | | 0.138 | X | | | | |
| Isoleucine | | | | | | | | | |
| Brunner (1978) [43] | Rat | PKU-induction | FGR | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| | | Control | Normal | 1.570 b | X | | | X | |
| | | | | 4.720 b | X | | | X | |
| Matsueda (1982) [97] | Rat | Control | Normal | 3.150 | X | | | X | |
| Mori (1999) [98] | Rat | Control | Normal | 2.368 | X | | | X | |
| Cysteine | | | | | | | | | |
| Balansky (2009) [106] | Mouse | Control | Normal | 1.000 | X | | | | |
| Miller (2014) [108] | Mouse | Control | Normal | 0.150 | X | | | X | |
| Moazzen (2014) [107] | Mouse | Streptozotocin | Pre-existent DM | 1.000 | X | | X | | |
| | | Control | Normal | 1.000 | X | | X | | |
| Lin (2011) [113] | Rat | High-fat diet | Over-growth | 0.075 a,b | X | | X | | |
| Soto-Blanco (2001) [114] | Rat | Control | Normal | 0.510 | X | | | X | |
| Hashimoto (2012) [120] | Guinea pig | Hypoxia | FGR | 0.550 | X | | | | |
| | | Normoxia | Normal | 0.550 | X | | | | |
| Herrera (2017) [121] | Guinea pig | Control | Normal | 0.500 | X | | | | |
| | | Uterine artery occlusion | FGR | 0.500 | X | | | | |
| Thompson (2011) [122] | Guinea pig | Nicotine exposed | Risk | 0.496 | X | | | | |
| Motawei (2016) [133] | Human | PE | PE | 0.005 | X | X | | | |
| Roes (2006) [134] | Human | Early onset severe PE/HELLP | PE | 0.025 a | X | | | | X |
| Shahin (2009) [137] | Human | Previous preterm labor | Risk | 0.008 a | X | | | | X |
| Methionine | | | | | | | | | |
| Abdel-Wanhab (1999) [115] | Rat | Control | Normal | 0.043 | X | | | | |
| Brunner (1978) [43] | Rat | PKU-induction | FGR | 1.570 b | X | | | X | |
| | | Control | Normal | 1.570 b | X | | | X | |
| Chandrashekar (1977) [116] | Rat | Control | Normal | NA | X | | | | |
| Fujii (1971) [117] | Rat | Control | Normal | 0.029 | X | | | | |
| Koz (2010) [118] | Rat | Control | Normal | 1.000 | X | | | | |
| Matsueda (1982) [97] | Rat | Control | Normal | 1.150 | X | | | X | |
| Viau (1973) [119] | Rat | Control | Normal | NA | X | | | | |
| Gauthier (2009) [123] | Guinea pig | Ethanol exposed | FGR | 0.096 | X | | | | |
| Othmani Mecif (2017) [124] | Rabbit | Control | Normal | 0.172 | X | | | | |
| Liu (2016) [125] | Sheep | Control | Normal | 0.047 | X | | | X | |
| Tsiplakou (2017) [126] | Sheep | Control | Normal | 0.041 | X | | | | |
| Mori (1999) [98] | Rat | Control | Normal | 2.368 | X | | | X | |
| Batistel (2017) [128] | Cow | Control | Normal | 0.012 a | X | | | | |
| Clements (2017) [129] | Cow | Control | Normal | 0.016 | X | | | | |
| Jacometo (2016) [130] | Cow | Control | Normal | 0.010 b | X | | | | |
| Xu (2018) [131] | Cow | Control | Normal | 0.012 a,b | X | | | | |
| Choline | | | | | | | | | |
| Bai (2012) [109] | Rat | Protein restriction | FGR | 0.670 a | X | | | X | |
| Thomas (2009) [110] | Rat | Ethanol exposed | FGR | 0.250 | X | | | | |
| | | Control | Normal | 0.250 | X | | | | |
| Yang (2000) [111] | Rat | Control | Normal | 0.390 a,b | X | | | | |
| Zhang (2018) [112] | Rat | LPS infusion | FGR | 0.875 b | X | X | | X | |
| | | Control | Normal | 0.875 b | X | X | | X | |
| Birch (2016) [127] | Sheep | Ethanol infusion | FGR | 0.010 | X | | | | |
| | | Control | Normal | 0.010 | X | | | | |
| Janovick Guretzky (2006) [151] | Cow | Control (Holstein) | Normal | 0.021 | | | | X | |
| | | Control (Jersey) | Normal | 0.030 | | | | X | |
| Zenobi (2018a) [132] | Cow | Excess energy diet | Risk overgrowth | 0.024 | X | | X | | |
| | | Maintenance diet | Normal | 0.024 | X | | X | | |
| Zenobi (2018b) [152] | Cow | Diet restriction | FGR | 0.002 | | | X | | |
| | | | | 0.003 | | | X | | |
| | | | | 0.005 | | | X | | |
| | | | | 0.007 | | | X | | |
| | | Ad libitum diet | Normal | 0.002 | | | X | | |
| | | | | 0.003 | | | X | | |
| | | | | 0.005 | | | X | | |
| | | | | 0.007 | | | X | | |
| Jacobson (2018) [138] | Human | Alcohol | Risk | 0.035 | X | X | | X | X |
| Ross (2013) [135] | Human | Control | Normal | 0.012 a | X | | | X | X |
| Yan (2012) [136] | Human | Control | Normal | 0.007 a | X | | | X | |
Table A5.
Data-extraction of included animal and human studies on birth weight.
Table A5.
Data-extraction of included animal and human studies on birth weight.
| Author (Year) | Species (Strain or Coumtry) | Animal Model | Pregnancy Complication | Maternal Weight (kg) | Dietary Protein Intake (%) | Supplementation Scheme a; intervention Type b | Supplementation Duration (GD) | Daily Dose (g/kg Body wt) | Measurement Day (GD) | AA BW ± SD (g) | n Offspring (Sex c) | n Mother | CON BW ± SD (g) | n Offspring (Sex c) | n Mother |
|---|
| Arginine | | | | | | | | | | | | | | | |
| Greene (2012) [33] | Mouse (FNVB/N × vegfr2-luc) | Control | Normal | 0.03 | 24 | C; T d | 1–18 | 2.660 e | P0 | 1.18 ± 0.28 | 49 | 6 | 1.32 ± 0.25 | 25 | 5 |
| Alexander (2004) [34] | Rat (SD) | RUPP | FGR/PE | 0.25 | NA | C; P | 10–19 | 2.000 e | 19 | 2.30 ± 0.60 | 117 | 9 | 2.00 ± 0.60 | 117 | 9 |
| | | Control | Normal | 0.25 | NA | C; P | 10–19 | 2.000 e | 19 | 2.80 ± 0.60 | 117 | 9 | 2.40 ± 0.35 | 156 | 12 |
| Altun (2008) [35] | Rat (Wistar) | Sonic stress–induction | PE | 0.20 | NA | C; T | 18–19 | 0.021 | 19 | 5.18 ± 0.58 | 67 | 6 | 4.41 ± 0.46 | 55 | 6 |
| Bourdon (2016) [44] | Rat (SD) | Protein restriction | FGR | 0.29 | 4 | C; T d | 7–21 | 1.500 | 21 | 4.13 ± 0.30 | 111 | 9 | 4.05 ± 0.15 | 110 | 9 |
| Bursztyn (2003) [45] | Rat (Wistar) | Hyperinsulinemia | FGR/PIH | 0.24 | NA | C; T | 11–22 | 0.217 | 22 | 4.10 ± 0.20 | 65 | 5 | 3.00 ± 1.60 | 55 | 5 |
| Chin (1991) [46] | Rat (Fisher F344/NTacfBR) | Control | Normal | 0.17 | NA | C; P d | 1-P0 | 0.159 | P0 | 4.60 ± 1.88 | 40 | 4 | 4.90 ± 1.88 | 28 | 4 |
| Da Costa (2014) [47] | Rat (Wistar) | Control | Normal | 0.20 | NA | I; P | 4–18 | 0.021 | 18 | 469.33 ± 124.55 | NA | 5 | 368.00 ± 138.12 | NA | 5 |
| Helmbrecht (1996) [48] | Rat (SD) | L–NAME | FGR/PE | 0.31 | NA | C; T | 16–21 | 0.021 | P0 | 5.60 ± 0.32 | 126 | 6 | 5.00 ± 0.17 | 120 | 6 |
| | | Control | Normal | 0.31 | NA | C; P | 16–21 | 0.021 | P0 | 6.11 ± 0.37 | 78 | 6 | 6.00 ± 0.27 | 80 | 6 |
| Podjarny (2001a) [49] | Rat (Wistar) | Hyperinsulinaemia | PIH | 0.24 | NA | C; P | 11–22 | 0.220 | 22 | 5.60 ± 0.39 | 180 | 15 | 4.30 ± 1.34 | 200 | 20 |
| Schooley (2002) [50] | Rat (SD) | Mg deficiency | FGR | 0.20 | NA | C; T | 10–21 | 1.000 | 21 | 3.97 ± 0.40 | 146 | 13 | 3.64 ± 0.42 | 133 | 12 |
| | | Control | Normal | 0.20 | NA | C; P | 10–21 | 1.000 | 21 | 4.10 ± 0.40 | 134 | 13 | 3.80 ± 0.45 | 129 | 14 |
| Sharkey (2001) [51] | Rat (SHHF/Mcc-facp) | Genetic | FGR/PIH | 0.20 | NA | C; P | 1–20 | 0.100 e,f | 20 | 1.57 ± 0.07 | NA | 3 | 2.68 ± 0.12 | NA | 4 |
| Vosatka (1998) [36] | Rat (Wistar) | Hypobaric hypoxia | FGR | 0.25 | 4 g/day | C; P | 9–21 | 0.200 e | 21 | 4.19 ± 0.41 | 38 | 4 | 3.30 ± 0.65 | 137 | 14 |
| | | | | | | | | 2.000 e | 21 | 4.27 ± 0.80 | 116 | 10 | 3.30 ± 0.65 | 137 | 14 |
| Zeng (2008) [37] | Rat (SD) | Control | Normal | 0.23 | 22 | C; P d | 0-P0 | 0.216 | P0 | 6.73 ± 0.87 | 174 | 12 | 6.43 ± 1.01 | 136 | 12 |
| Crane (2016) [52] | Sheep (Rambouillet) | Control | Normal | 65.0 | 16 | C; P | 0–14 | 0.150 | P0 | 5300 ± 1073 | 20 | 20 | 5400 ± 1200 | 29 | 25 |
| Peine (2018) [53] | Sheep (Rambouillet-cross) | Diet restriction | FGR | 67.7 | 16 | C; P | 54-P0 | 0.180 | P0 | 4603.00 ± 1454 | 11 | 11 | 4449 ± 1454 | 11 | 11 |
| Sun (2017) [54] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | NA | C; P | 35–110 | 0.250 | 110 | 1660 ± 877 | 16 | 8 | 1400 ± 396 | 16 | 8 |
| Sun (2018) [55] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; T | 35–110 | 0.249 | 110 | 1703 ± 154 | 16 | 8 | 1431 ± 156 | 16 | 8 |
| Zhang (2016a) [56] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; P | 35–110 | 0.250 | 110 | 1660 ± 2998 | 16 | 8 | 1410 ± 2998 | 16 | 8 |
| Zhang (2016b) [57] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; P | 35–110 | 0.250 | 110 | 1659 ± 130 | 16 | 8 | 1401.00 ± 130.11 | 16 | 8 |
| Bass (2017) [58] | Pig (GPK-35) | Control | Normal | 180 | 18.7 | C; P d | 93-P0 | 0.150 | P0 | 1420 ± 197 | 682 | 49 | 1410 ± 197 | 652 | 48 |
| | Pig (PIC Camborough 1050 and 1055) | Control | Normal | 165 | 12 | C; P d | 81–116 | 0.170 | P0 | 1410 ± 391 | 2515 | 195 | 1450 ± 391 | 2388 | 188 |
| Bérard (2010) [59] | Pig (Swiss Large White) | Control | Normal | 159 | NA | C; P | 14–28 | 0.160 | 75 | 373 ± 63 | 29 (F) | 10 | 365 ± 63 | 20 (F) | 7 |
| Che (2013) [70] | Pig (Landrace × large White) | Control | Normal | 236 | 14 | C; P d | 30–90 | 0.082 e | P0 | 13,900 ± 268 | 212 | 20 | 1480 ± 224 | 204 | 20 |
| | | | | | | | 30–114 | 0.092 e | P0 | 1500 ± 224 | 236 | 20 | 1480 ± 224 | 204 | 20 |
| Dallanora (2017) [69] | Pig (Landrace × Large white) | Control | Normal | 148 | 17 | C; P | 25–112 | 0.115 | 80–112 | 1170 ± 282 | 755 | 51 | 1160 ± 282 | 796 | 51 |
| Gao (2012) [71] | Pig (Yorkshire × Landrace) | Control | Normal | 160 | 13 | C; P d | 22-P0 | 0.140 e | P0 | 1450 ± 624 | 642 | 52 | 1410 ± 624 | 630 | 56 |
| Garbossa (2015) [72] | Pig | Control | Normal | 160 | 14 | C; P | 25–53 | 0.113 e | 110 | 1430 ± 379 | 334 | 23 | 1380 ± 379 | 305 | 23 |
| | | Ractopamine | Overgrowth | 160 | 14 | C; T | 25–53 | 0.113 e | 110 | 1510 ± 379 | 334 | 22 | 1500 ± 379 | 305 | 22 |
| Guo (2016) [73] | Pig (Yorkshire × Landrace) | Control | Normal | 150 | 16 | C; P d | 30–110 | 0.015 e | P0 | 1250 ± 319 | 60 | 60 | 1250 ± 319 | 53 | 53 |
| Li (2010) [74] | Pig (Yorkshire × Landrace) | Control | Normal | 112 | 12 | C; P d | 0–25 | 0.070 | 25 | 8.90 ± 3.08 | 111 | 9 | 8.30 ± 4.08 | 114 | 9 |
| | | | | | | | | 0.140 | 25 | 5.50 ± 4.08 | 77 | 8 | 8.30 ± 4.08 | 114 | 9 |
| Li (2014) [75] | Pig (Yorkshire × Landrace) | Control | Normal | 115 | 12 | C; P d | 14–25 | 0.070 | 25 | 6.27 ± 1.18 | 191 | 15 | 5.58 ± 1.18 | 147 | 14 |
| | | | | | | | | 0.140 | 25 | 5.83 ± 1.18 | 171 | 14 | 5.58 ± 1.18 | 147 | 14 |
| Li (2015) [76] | Pig (Landrace) | Control | Normal | 125 | 16 | C; P d | 1–30 | 0.210 | P0 | 1440 ± 340 | 354 | 32 | 1460 ± 397 | 306 | 30 |
| | | | | 187 | 16 | C; P d | 1–30 | 0.140 | P0 | 1440 ± 397 | 756 | 57 | 1510 ± 397 | 651 | 56 |
| | | | | 196 | 16 | C; P d | 1–30 | 0.130 | P0 | 1400 ± 374 | 481 | 37 | 1440 ± 374 | 123 | 37 |
| Liu (2012) [77] | Pig (Landrace x Large White) | Control | Normal | 187 | 15 | C; P | 1-P0 | 0.107 | P0 | 1510 ± 104 | 98 | 9 | 1450 ± 104 | 88 | 9 |
| Madsen (2017) [60] | Pig (Swiss Large White) | UOL | Normal | 159 | 15 | C; P d | 14–28 | 0.157 e | P0 | 1440 ± 411 | NA (F) | 5 | 1500 ± 411 | NA (F) | 5 |
| | | | | | | | | | | | NA (M) | 5 | 1740 ± 353 | NA (M) | 5 |
| | | Intact (relatively crowded) | FGR | 159 | 15 | C; T d | 14–28 | 0.157 e | P0 | 1510 ± 411 | NA (F) | 5 | 1260 ± 411 | NA (F) | 5 |
| | | | | | | | | | | | NA (M) | 5 | 1310 ± 353 | NA (M) | 5 |
| Mateo (2007) [61] | Pig (Camborough 22) | Control | Normal | 166 | 12 | C; P d | 30-P0 | 0.120 | P0 | 1460 ± 288 | 273 | 24 | 1410 ± 288 | 262 | 28 |
| Mateo (2008) [62] | Pig (Camborough 22) | Control | Normal | 166 | 12 | C; P d | 30-P0 | 0.120 | P0 | 1434 ± 154 | NA | 21 | 1430 ± 154 | | 17 |
| Wu (2012) [63] | Pig (Landrace × Large White) | Control | Normal | 187 | 15 | C; P | 90-P0 | 0.110 | P0 | 16,200 ± 104 | 97 | 9 | 1460 ± 104 | 83 | 9 |
| Quesnel (2014) [64] | Pig (Landrace × Large White) | Control | Normal | 236 | 13 | C; P | 70-P0 | 0.110 | P0 | 1520 ± 376 | 357 | 24 | 1460 ± 379 | 317 | 23 |
| Camarena Pulido (2016) [78] | Human (Mexico) | Increased risk of PE | Risk | 71 | NA | I; P | Wk 20-P0 | 0.042 e | P0 | 3144 ± 454 | 49 | 49 | 2937 ± 491 | 47 | 47 |
| Facchinetti (2007) [79] | Human (Italy) | HTN after 20th wk | Risk | 80 | NA | I; P | 2 wks | 0.110 e | P0 | 2753 ± 857 | 39 | 39 | 2523 ± 803 | 35 | 35 |
| Hladenuwich (2006) [82] | Human (USA) | PE | PE | 70 | NA | I; T | 6 days | 0.200 e | P0 | 1734 ± 680 | 10 | 10 | 1653 ± 602 | 10 | 10 |
| Neri (2010) [83] | Human (Italy) | Chronic HTN < 16 wk | Risk | 85 | NA | I; P | 10–12 wks duration | 0.047 e | P0 | 3094 ± 719 | 39 | 39 | 2836 ± 946 | 40 | 40 |
| Ropacka (2007) [84] | Human (Poland) | FGR (EFW < p10) | FGR | 70 | NA | I; T | Until P0 (±35 days) | 0.043 e | P0 | 2526 ± 844 | 24 | 24 | 1996 ± 928 | 17 | 17 |
| Rytlewski (2006) [85] | Human (Poland) | PE; BW < p10 | FGR/PE | 74 | NA | I; T | Wk 29-P0 | 0.041 | P0 | 2358 ± 901 | 30 | 30 | 2066 ± 917 | 31 | 31 |
| Rytlewski (2008) [86] | Human (Poland) | Preterm labor sign | Preterm | 74 | NA | I; T | 1–4 wks after admission-delivery | 0.041 e | P0 | 2956 ± 538 | 25 | 25 | 2987 ± 474 | 20 | 20 |
| Sieroszewski (2004) [87] | Human (Poland) | EFW < p10 | FGR | 75 | NA | I; T | 20 days (start wk 32) | 0.040 e | P0 | 2823 ± 751 | 78 | 78 | 2495 ± 805 | 30 | 30 |
| Singh (2015) [88] | Human (India) | EFW < p10 | FGR | 48 | NA | I; T | 21 days | 0.065 e | P0 | 1900 ± 380 | 30 | 30 | 1770 ± 530 | 30 | 30 |
| Staff (2004) [89] | Human (Norway) | PE | FGR/PE | 80 | NA | I; T | 5 days | 0.150 e | P0 | 2264 ± 833 | 15 | 15 | 1986 ± 905 | 15 | 15 |
| Winer (2009) [80] | Human (France) | AC < p3 and abnormal UA flow | FGR | 75 | NA | I; T | Until P0 (±3 wk) | 0.190 | P0 | 1042 ± 476 | 21 | 21 | 1068 ± 452 | 22 | 22 |
| Zhang (2007) [81] | Human (China) | PIH and FGR | FGR/PIH | 60 | NA | I; T | Wk 28–30 | 0.333 | P0 | 2900 ± 300 | 35 | 35 | 2700 ± 300 | 33 | 33 |
| Citrulline | | | | | | | | | | | | | | | |
| Bourdon (2016) [44] | Rat (SD) | Protein restriction | FGR | 0.29 | 4 | C; T | 7–21 | 2.000 | | 4.15 ± 0.81 | 102 | 9 | 4.05 ± 0.15 | 110 | 9 |
| Koeners (2007) [38] | Rat (SHR) | Pre-existent HTN | PIH | 0.20 | NA | C; T | 7-P0 | 0.438 f | P0 | 5.10 ± 0.56 | 14 (F) | 7 | 4.88 ± 0.74 | 16 (F) | 5 |
| | | | | | | | | | | 5.32 ± 0.80 | 14 (M) | 7 | 5.50 ± 0.52 | 17 (M) | 5 |
| Tran (2017) [39] | Rat (SD) | Protein restriction | FGR | 0.20 | 4 | C; P | 2–15 | 2.000 | 15 | 0.29 ± 0.03 | 18 | 3 | 0.40 ± 0.02 | 24 | 3 |
| | | | | | | | 2–21 | 2.000 | 21 | 4.88 ± 0.42 | 30 | 3 | 4.62 ± 0.33 | 46 | 3 |
| Glutamate | | | | | | | | | | | | | | | |
| Beaudoin (1981) [40] | Rat (Wistar Albino) | Control | Normal | 0.23 | NA | I; P | 6–10 | 2.600 e | 20 | 3.96 ± 0.02 | 54 | 4 | 3.97 ± 0.04 | 57 | 5 |
| Zeng (2012) [41] | Rat (SD) | Control | Normal | 0.24 | NA | C; P | 1-P0 | 0.074 f | P0 | 6.37 ± 0.29 | 1171 | 96 | 6.38 ± 0.29 | 1094 | 96 |
| | | | | | | | | 0.149 f | P0 | 6.36 ± 0.20 | 1248 | 96 | 6.38 ± 0.29 | 1094 | 96 |
| Sun (2017) [54] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | NA | C; P | 35–110 | 0.060 | 220 | 1680 ± 1188 | 16 | 8 | 1400 ± 396 | 16 | 8 |
| Sun (2018) [55] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; T | 35–110 | 0.062 | 110 | 1718 ± 110 | 16 | 8 | 1431 ± 156 | 16 | 8 |
| Zhang (2016a) [56] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; P | 35–110 | 0.060 | 110 | 1680 ± 2998 | 16 | 8 | 1410 ± 2998 | 16 | 8 |
| Zhang (2016b) [57] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40.1 | 10–14 | C; P | 35–110 | 0.060 | 110 | 1682 ± 130 | 16 | 8 | 1401 ± 130 | 16 | 8 |
| Liu (2012) [77] | Pig (Landrace × Large White) | Control | Normal | 187 | 15 | I; P | 1-P0 | 0.011 | P0 | 1490 ± 104 | 97 | 9 | 1450 ± 104 | 88 | 9 |
| Wu (2012) [63] | Pig (Landrace × Large White) | Control | Normal | | NA | I; P | 90-P0 | 0.011 | P0 | 1590 ± 104 | 95 | 9 | 1460 ± 104 | 83 | 9 |
| Zhang (2014) [65] | Pig (Landrace × Large White) | Control | Normal | 136 | 15 | C; P | 1-P0 | 0.007 | P0 | 1460 ± 67 | 99 | 9 | 1370 ± 67 | 89 | 9 |
| | | | | | | | | 0.015 | P0 | 1410 ± 67 | 97 | 9 | 1370 ± 67.08 | 89 | 9 |
| | | | | | | | | 0.022 | P0 | 1430 ± 67 | 98 | 9 | 1370.00 ± 67 | 89 | 9 |
| | | | | | | | | 0.030 | P0 | 1360 ± 67 | 95 | 9 | 1370 ± 67 | 89 | 9 |
| Cai (2018) [66] | Swine (Landrace × Yorkshire) | Control | Normal | 210 | 13 | C; P | 1–8 | 0.003 | P0 | 1450 ± 415 | 177 | 18 | 1440 ± 415 | 173 | 18 |
| | | | | | | | 9–28 | 0.003 | P0 | 1430 ± 415 | 166 | 16 | 1440 ± 415 | 173 | 18 |
| | | | | | | | 1–28 | 0.003 | P0 | 1400 ± 415 | 207 | 17 | 1440 ± 415 | 173 | 18 |
| Glutamine | | | | | | | | | | | | | | | |
| Caporossi (2014) [42] | Rat (Wistar) | Control | Normal | 0.24 | NA | I; P | 1–21 | 2.000 | 21 | 3.53 ± 2.11 | 46 | 6 | 3.61 ± 2.23 | 45 | 6 |
| Zhu (2018) [67] | Pig (Landrace × Large White) | Control | Normal | 272 | NA | C; P d | 85–114 | 0.126 | P0 | 1390 ± 110 | 372 | 30 | 1340 ± 164 | 367 | 30 |
| Proline | | | | | | | | | | | | | | | |
| Gonzalez-Anover (2017) [68] | Pig (Landrace × Yorkshire) | Control | Normal | 115 | 14 | C; P | 11–30 | 0.120 e | P0 | 1400 ± 755 | 844 | 57 | 1400 ± 762 | 835 | 58 |
| Aspartic acid | | | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 2.60 ± 0.18 | 36 | 4 | 2.57 ± 0.36 | 36 | 4 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 f | 20 | 2.96 ± 0.24 | 36 | 4 | 3.21 ± 0.24 | 36 | 4 |
| Valine | | | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 2.52 ± 0.26 | 28 | 3 | 2.57 ± 0.36 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 2.42 ± 0.26 | 27 | 3 | 2.54 ± 0.31 | 108 | 12 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 f | 20 | 2.86 ± 0.26 | 28 | 3 | 3.21 ± 0.24 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 3.15 ± 0.42 | 27 | 3 | 3.07 ± 0.21 | 108 | 12 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | 6 | C; P | 1–22 | 2.710 | 22 | 3.49 ± 0.29 | 53 | 5 | 3.69 ± 0.21 | 53 | 5 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 22 | 5.20 ± 0.10 | 66 | 6 | 5.40 ± 0.40 | 99 | 9 |
| Xu (2017) [104] | Pig (Landrace × Large White) | Control | Normal | 220 | 18 | C; P | 107-P0 | 0.037 | P0 | 1520 ± 396 | 78 | 8 | 1540 ± 396 | 82 | 8 |
| | | | | | | | | 0.087 | P0 | 1410 ± 396 | 80 | 8 | 1540 ± 396 | 82 | 8 |
| Leucine | | | | | | | | | | | | | | | |
| Viana (2013) [103] | Mouse (NMRI) | Control | Normal | 0.03 | 18 | C; P d | 1–19 | 3.000 e,f | 19 | 0.95 ± 0.07 | 6 | 6 | 0.81 ± 0.03 | 6 | 6 |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 2.49 ± 0.30 | 36 | 4 | 2.57 ± 0.36 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 2.70 ± 0.21 | 27 | 3 | 2.54 ± 0.311 | 108 | 12 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 f | 20 | 3.10 ± 0.24 | 36 | 4 | 3.21 ± 0.24 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 3.04 ± 0.21 | 27 | 3 | 3.07 ± 0.21 | 108 | 12 |
| Cruz (2014) [99] | Rat (Wistar) | Control | Normal | 0.25 | 18 | C; P | 10–20 | NA | 20 | 3.34 ± 0.24 | 24 | 8 | 3.67 ± 0.20 | 24 | 8 |
| Cruz (2016) [100] | Rat (Wistar) | Control | Normal | 0.25 | 18 | C; P | 2–20 | NA | 20 | 3.65 ± 0.19 | 101 | 10 | 3.92 ± 0.22 | 100 | 10 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | 6 | C; P | 1–22 | 1.500 | 22 | 2.79 ± 0.31 | 49 | 5 | 2.05 ± 0.62 | 73 | 7 |
| Mawatari (2004) [102] | Rat (SD) | Control | Normal | 0.30 | NA | I; P | 7–17 | 0.300 | 20 | 4.11 ± 0.22 | 148 (M) | 19 | 4.13 ± 0.29 | 128 (M) | 19 |
| | | | | | | | | | | 3.89 ± 0.27 | 136 (F) | 19 | 3.89 ± 0.35 | 115 (F) | 19 |
| | | | | | | | | 1.000 | 20 | 4.12 ± 0.22 | 141 (M) | 20 | 4.13 ± 0.29 | 128 (M) | 19 |
| | | | | | | | | | | 3.83 ± 0.17 | 151 (F) | 20 | 3.89 ± 0.35 | 115 (F) | 19 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 22 | 5.10 ± 0.40 | 45 | 5 | 5.40 ± 0.40 | 99 | 9 |
| Ventrucci (2001) [101] | Rat (Wistar) | Control | Normal | 0.15 | 18 | C; P | 1–20 | 7.180 f | 20 | 3.45 ± 0.51 | | 10 | 3.73 ± 0.41 | | 10 |
| Wang (2018) [105] | Pig (Landrace × Large White) | Control | Normal | 260 | 15 | C; P | 70-P0 | 0.046 | P0 | 1470 ± 49 | 70 | 6 | 1490 ± 49 | 68 | 6 |
| | | | | | | | | 0.092 | P0 | 1550 ± 49 | 63 | 6 | 1490 ± 49 | 68 | 6 |
| | | | | | | | | 0.138 | P0 | 1460 ± 49 | 70 | 6 | 1490 ± 49 | 68 | 6 |
| Isoleucine | | | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 2.53 ± 0.26 | 28 | 3 | 2.57 ± 0.36 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 2.88 ± 0.31 | 27 | 3 | 2.54 ± 0.31 | 108 | 12 |
| | | Control | Normal | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 3.24 ± 0.24 | 36 | 4 | 3.21 ± 0.24 | 36 | 4 |
| | | | | | | | | 4.720 f | 20 | 2.95 ± 0.20 | 27 | 3 | 3.07 ± 0.21 | 108 | 12 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | 6 | C; P | 1–22 | 3.150 | 22 | 3.81 ± 0.47 | 54 | 5 | 4.11 ± 0.22 | 47 | 5 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 22 | 5.30 ± 0.30 | 52 | 4 | 5.40 ± 0.40 | 99 | 9 |
| Cysteine (NAC) | | | | | | | | | | | | | | | |
| Balansky (2009) [106] | Mouse (Swiss-Albino) | Control | Normal | 0.03 | NA | C; P | 1-P0 | 1.000 | P0 | 1.20 ± 0.04 | 47 | 5 | 1.30 ± 0.09 | 94 | 9 |
| Moazzen (2014) [107] | Mouse (C57BL/6) | Streptozotocin | pre-existent DM | 0.03 | NA | C; P | 0.5–18.5 | 1.000 | P0 | 1.15 ± 0.07 | 28 | 8 | 0.99 ± 0.08 | 28 | 8 |
| | | Control | Normal | 0.03 | NA | C; P | 0.5–18.5 | 1.000 | P0 | 1.24 ± 0.08 | 28 | 8 | 1.24 ± 0.08 | 28 | 8 |
| Miller (2014) [108] | Mouse (CD-1) | Control | Normal | NA | 19 | I; P | 6–13 | 0.150 | 17 | 1.07 ± 0.38 | 353 | 25 | 1.00 ± 0.37 | 349 | 25 |
| Lin (2011) [113] | Rat (SD) | HF diet | Overgrowth | 0.23 | 17 | C; P | 1–19.5 | 0.075 e,f | 19.5 | 2.39 ± 0.19 | 213 | 20 | 2.38 ± 0.10 | 200 | 20 |
| Soto-Blanco (2001) [114] | Rat (Wistar) | Control | Normal | 0.27 | 16 | C; P | 6–21 | 0.510 | 21 | 4.37 ± 0.24 | 100 | 12 | 4.44 ± 0.24 | 87 | 12 |
| Hashimoto (2012) [120] | Guinea pig (Dunkin-Hartley) | Hypoxia | FGR | NA | NA | C; T | 52–62 | 0.550 | 63 | 59.8 ± 5.88 | | 6 | 59.5 ± 5.9 | | 6 |
| | | Normoxia | Normal | NA | NA | C; P | 52–62 | 0.550 | 63 | 76.6 ± 19.1 | | 6 | 88.8 ± 11.9 | | 7 |
| Herrera (2017) [121] | Guinea pig (Pirbright White) | Control | Normal | NA | 19 | C; P | 34–67 | 0.500 | 67 | 79.1 ± 19.8 | 13 | | 83.7 ± 12.4 | 16 | NA |
| | | UA occlusion | FGR | NA | 19 | C; P | 34–67 | 0.500 | 67 | 74.1 ± 13.5 | 9 | | 57.9 ± 15.3 | 9 | |
| Thompson (2011) [122] | Guinea pig (Dunkin-Hartley) | Nicotine exposed | Risk | NA | NA | C; P | 52–62 | 0.496 | 62 | 65.8 ± 5.4 | NA | 2 | 78.2 ± 6.4 | NA | 2 |
| Motawei (2016) [133] | Human (Egypt) | PE | PE | 85 | NA | I; T | Mo 5-term (<4 months) | 0.005 | P0 | 2560 ± 590 | 50 | 50 | 198 ± 420 | 50 | 50 |
| Roes (2006) [134] | Human (The Netherlands) | Early onset severe PE or HELLP | PE | 72 | NA | I; T | inclusion-delivery (±6 days) | 0.025 e | P0 | 9712 ± 419 | 19 | 19 | 1070 ± 399 | 19 | 19 |
| Shahin (2009) [137] | Human (Egypt) | Previous preterm labor | Risk | 80 | NA | I; P | Wk 17–36 or P0 | 0.008 e | P0 | 3107 ± 232 | 140 | 140 | 2715 ± 357 | 140 | 140 |
| Methionine | | | | | | | | | | | | | | | |
| Abdel-Wanhab (1999) [115] | Rat (SD) | Control | Normal | 0.20 | NA | C; P | 6–15 | 0.043 | 20 | 4.40 ± 0.33 | 69 | 8 | 4.50 ± 0.42 | 71 | 8 |
| Brunner (1978) [43] | Rat (SD) | PKU-inducing diet | FGR | 0.22 | NA | C; T | 10–20 | 1.570 f | 20 | 2.56 ± 0.18 | 36 | 4 | 2.57 ± 0.36 | 36 | 4 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 f | 20 | 3.19 ± 0.24 | 36 | 4 | 3.21 ± 0.24 | 36 | 4 |
| Chandrashekar (1977) [116] | Rat (SD) | Control | Normal | 0.20 | 18 | C; P | 1–21 | NA | 21 | 2.30 ± 0.41 | 172 | 17 | 3.20 ± 0.59 | 368 | 35 |
| Fujii (1971) [117] | Rat (McCollum) | Control | Normal | 0.25 | 20 | C; P | 1-P0 | 0.029 | P0 | 6.40 ± 0.16 | NA (M) | 4 | 6.40 ± 0.18 | NA (M) | 5 |
| | | | | | | | | | | 6.10 ± 0.20 | NA (F) | 4 | 6.10 ± 0.13 | NA (F) | 5 |
| Koz (2010) [118] | Rat (Wistar) | Control | Normal | 0.25 | NA | C; P | 1-P0 | 1.000 | P0 | 5.90 ± 0.30 | 25 | 5 | 6.10 ± 0.35 | 25 | 5 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | 6 | C; P | 1–22 | 1.130 | 22 | 1.63 ± 0.27 | 45 | 5 | 2.05 ± 0.62 | 73 | 7 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 22 | 2.60 ± 0.10 | 88 | 8 | 5.40 ± 0.40 | 99 | 9 |
| Viau (1973) [119] | Rat (SD) | Control | Normal | 0.19 | 18 | C; P | 1–20 | NA | 20 | 2.50 ± 0.32 | 101 | 10 | 3.60 ± 0.28 | 82 | 8 |
| Gauthier (2009) [123] | Guinea pig | Ethanol exposed | FGR | 0.70 | NA | C; P | 35–71 | 0.096 | 71 | 99.0 ± 39.2 | 6 | NA | 98.0 ± 36.7 | 6 | NA |
| Othmani Mecif (2017) [124] | Rabbit (White) | Control | Normal | 2.90 | 30 | C; P | 0–29 | 0.172 | 29 | 36.0 ± 32.8 | 22 | 8 | 50.0 ± 26.8 | 45 | 8 |
| Liu (2016) [125] | Sheep (Merino) | Control | Normal | 63 | NA | I; P | 111-P0 | 0.047 | P0 | 4620.0 ± 1929.6 | 79 | 60 | 4170.00 ± 1929.61 | 73 | 60 |
| Tsiplakou (2017) [126] | Sheep (Chios) | Control | Normal | 66 | 16 | C; P | Last 15 days | 0.041 | P0 | 4.49 ± 0.66 | 16 | 15 | 4.51 ± 0.90 | 19 | 15 |
| Batistel (2017) [128] | Cow (Holstein) | Control | Normal | 735 | 16 | C; P | Last 28 days | 0.012 e | P0 | 44,062 ± 5995 | 42 | 30 | 41,947 ± 5776 | 39 | 30 |
| Clements (2017) [129] | Cow (Angus × Simmental) | Control | Normal | 635 | 12 | C; P | Last 23 days (±7 days) | 0.016 | P0 | 35,000 ± 4157 | NA | 6 | 35,000 ± 4156 | NA | 6 |
| Jacometo (2016) [130] | Cow (Holstein) | Control | Normal | 773 | NA | C; P | Last 21 days | 0.010 f | P0 | 44,200 ± 5543 | 12 | 12 | 42,900 ± 5196 | 12 | 12 |
| Xu (2018) [131] | Cow (Holstein) | Control | Normal | 735 | 16 | C; P | Last 28 days | 0.012 e,f | P0 | 41,760 ± 4432 | NA | 21 | 40,936 ± 6190 | NA | 18 |
| Choline | | | | | | | | | | | | | | | |
| Bai (2012) [109] | Rat (Wistar) | Protein restriction | FGR | 0.30 | 9 | C; P | 1-P0 | 0.670 e | P0 | 5.72 ± 0.17 | 32 (M) | 8 | 5.62 ± 0.20 | 32 (M) | 8 |
| | | | | | | | | | | 5.58 ± 0.17 | 32 (F) | 8 | 5.35 ± 0.20 | 32 (F) | 8 |
| Thomas (2009) [110] | Rat (SD) | Ethanol exposed | FGR | 0.20 | NA | I; P | 5–20 | 0.250 | P0 | 6.90 ± 0.35 | 154 | 12 | 6.50 ± 0.33 | 160 | 11 |
| | | Control | Normal | 0.20 | NA | I; P | 5–20 | 0.250 | P0 | 6.90 ± 0.32 | 144 | 10 | 6.70 ± 0.37 | 200 | 14 |
| Yang (2000) [111] | Rat (SD) | Control | Normal | NA | NA | C; P | 11–17 | 0.390 e,f | P0 | 6.35 ± 0.27 | 177 | 15 | 6.28 ± 0.24 | 155 | 12 |
| Zhang (2018) [112] | Rat (SD) | LPS | FGR | 0.20 | NA | C; P | 1–20 | 0.875 f | 20 | 4.02 ± 0.08 | 125 | 9 | 3.76 ± 0.13 | 105 | 9 |
| | | Control | Normal | 0.20 | NA | C; P | 1–20 | 0.875 f | 20 | 4.35 ± 0.10 | 68 | 6 | 4.08 ± 0.06 | 79 | 6 |
| Birch (2016) [127] | Sheep (Suffolk) | Ethanol infusion | FGR | 75 | 13 | C; P | 4–148 | 0.010 | P0 | 4940 ± 764 | NA | 8 | 4340 ± 1347 | NA | 14 |
| | | Control | Normal | 75 | 13 | C; P | 4–148 | 0.010 | P0 | 6150 ± 1592 | NA | 6 | 5740 ± 933 | NA | 8 |
| Zenobi (2018a) [132] | Cow (Holstein) | Excess energy diet | Risk overgrowth | 735 | 14 | C; T | Last 21 days | 0.024 | P0 | 37,400 ± 7099 | 9 | 25 | 40,800 ± 7099 | 9 | 22 |
| | | Maintenance energy diet | Normal | 735 | 14 | C; P | Last 21 days | 0.024 | P0 | 39,300 ± 7099 | 9 | 21 | 40,100 ± 2099 | 8 | 25 |
| Jacobson (2018) [138] | Human (South Africa) | Alcohol use | Risk | 57 | NA | I; T | Enrollment-P0 | 0.034 | P0 | 2853 ± 451 | 31 | 31 | 2844 ± 658 | 31 | 31 |
| Ross (2013) [135] | Human (USA) | Control | Normal | 78 | NA | I; P | Wk 17-P0 | 0.012 e | P0 | 3114 ± 636 | 46 | 46 | 3193 ± 540 | 47 | 47 |
| Yan (2012) [136] | Human (USA) | Control | Normal | 63 | NA | I; P | Wk 27–39 | 0.007 e | P0 | 3500 ± 300 | 13 | 13 | 3400 ± 400 | 13 | 13 |
Table A6.
Data extraction of included animal and human studies on maternal blood pressure.
Table A6.
Data extraction of included animal and human studies on maternal blood pressure.
| Author (Year) | Species (Strain or Strain) | Model | Pregnancy Complication | Maternal Weight (kg) | Dietary Protein Intake (%) | Supplementation Scheme a; intervention type b | Daily Dose (g/kg Body wt) | Supplementation Duration (GD) | Technique (Office/Restrained vs. 24 h/Unrestrained) | Measurement Day (GD) | BP Type | AA BP ± SD (mmHg) | n | CON BP ± SD (mmHg) | n |
|---|
| Arginine | | | | | | | | | | | | | | | |
| Alexander (2004) [34] | Rat (SD) | RUPP | FGR/PE | 0.25 | NA | C; P | 2.00 c | 10–19 | Telemetry (unrestrained) | 19 | MAP | 113 ± 6 | 9 | 132 ± 6 | 9 |
| | | Control | Normal | 0.25 | NA | C; P | 2.00 c | 10–19 | Telemetry (unrestrained) | 19 | MAP | 97 ± 9 | 9 | 109 ± 7 | 12 |
| Altun (2008) [35] | Rat (Wistar) | Sonic stress-induction | PE | 0.20 | NA | C; T | 0.02 | 18–19 | Tail cuff (restrained) | 20 | MAP | 112 ± 10 | 6 | 145 ± 10 | 6 |
| Bursztyn (2003) [45] | Rat (Wistar) | Hyper-insulinemia | FGR/PIH | 0.24 | NA | C; T | 0.22 | 11-P0 | Tail cuff (restrained) | 19/20 | SBP | 77 ± 8 | 5 | 119 ± 14 | 5 |
| Helmbrecht (1996) [48] | Rat (SD) | L-NAME | FGR/PE | 0.31 | NA | C; T | 0.02 | 16–21 | Tail cuff (restrained) | 21 | SBP | 125 ± 7 | 9 | 153 ± 13 | 9 |
| | | Control | Normal | 0.31 | NA | C; P | 0.02 | 16–21 | Tail cuff (restrained) | 21 | SBP | 117 ± 14 | 6 | 115 ± 7 | 6 |
| Podjarny (1993) [90] | Rat (Wistar) | Control | Normal | 0.23 | 20 | C; T | 0.32 | 12–22 | Tail artery cannulation (restrained) | 22 | MAP | 90 ± 7 | 10 | 91 ± 4 | 10 |
| | | Adriamycin nephropathy | PE | 0.23 | 20 | C; T | 0.32 | 12–22 | Tail artery cannulation (restrained) | 22 | MAP | 90 ± 11 | 10 | 135 ± 13 | 10 |
| Podjarny (2001a) [49] | Rat (Wistar) | Hyper-insulinemia | PIH | 0.24 | 20 | C; P | 0.22 | 11–22 | Tail cuff (restrained) | 19/20 | SBP | 83 ± 9 | 15 | 104 ± 13 | 20 |
| Podjarny (2001b) [91] | Rat (Wistar) | Control | Normal | 0.23 | 20 | C; P | 0.26 | 11–22 | Tail cuff (restrained) | 22 | SBP | 87 ± 1 | 4 | 83 ± 6 | 4 |
| | | L-NAME | FGR/PE | 0.23 | 20 | C; T | 0.26 | 11–22 | Tail cuff (restrained) | 22 | SBP | 89 ± 4 | 8 | 129 ± 4 | 8 |
| Podjarny (1997) [92] | Rat (Wistar) | Adriamycin nephropathy | PE | 0.22 | 20 | C; T | 0.23 | 11–22 | Tail cuff (restrained) | 22 | SBP | 106 ± 7 | 5 | 122 ± 7 | 8 |
| | | | | | | | | | | | MAP | 91 ± 5 | 5 | 124 ± 7 | 8 |
| Schooley (2002) [50] | Rat (SD) | Mg deficiency | FGR | 0.20 | NA | C; T | 1.00 | 10–21 | Tail cuff (restrained) | 21 | SBP | 111 ± 11 | 13 | 160 ± 10 | 12 |
| | | Control | Normal | 0.20 | NA | C; P | 1.00 | 10–21 | Tail cuff (restrained) | 21 | SBP | 121 ± 11 | 13 | 148 ± 11 | 14 |
| Sharkey (2001) [51] | Rat (SHHF) | Genetic | FGR/PIH | 0.20 | NA | C; P | 0.10 c,d | 1–20 | Tail cuff (restrained) | 15–19 | SBP | 142 ± 14 | 3 | 188 ± 14 | 4 |
| Camarena Pulido (2016) [78] | Human (Mexico) | Increased risk of PE | Risk | 71 | NA | I; P | 0.04 c | wk 20-P0 | Sphygmo-manometer (office) | wk 39 | SBP | 118 ± 12 | 49 | 126 ± 14 | 47 |
| | | | | | | | | | | | DBP | 73 ± 11 | 49 | 79 ± 14 | 47 |
| | | | | | | | | | | | MAP | 88 ± 11 | 49 | 95 ± 5 | 47 |
| Rytlewski (2005) [93] | Human (Poland) | PE; BW < p10 | FGR/PE | 75 | NA | I; T | 0.04 | wk 29-P0 | Sphygmo-manometer (office) | After 3 wks | SBP | 134 ± 3 | 30 | 143 ± 3 | 31 |
| | | | | | | | | | | | DBP | 82 ± 2 | 30 | 87 ± 1 | 31 |
| | | | | | | | | | | | MAP | 102 ± 2 | 30 | 108 ± 1 | 31 |
| Staff (2004) [89] | Human (Norway) | PE | PE | 69 | NA | I; T | 0.18 c | 5 days | NA | After suppl. | SBP | 147 ± 5 | 10 | 148 ± 5 | 8 |
| | | | | | | | | | | | DBP | 98 ± 3 | 10 | 102 ± 4 | 8 |
| Neri (2010) [83] | Human (Italy) | Chronic HTN | Risk | 90 | NA | I; P | 0.04 c | 10–12 wks | Sphygmo-manometer (24 h) | Aftersuppl. | SBP | 129 ± 8 | 39 | 130 ± 14 | 40 |
| | | | | | | | | | Sphygmo-manometer (24 h) | After suppl. | DBP | 80 ± 7 | 39 | 79 ± 10 | 40 |
| Winer (2009) [80] | Human (France) | AC < p3 and abnormal UA flow | FGR | 75 | NA | I; T | 0.19 | Until P0 (3.5 wks) | NA | After suppl. | SBP | 133 ± 19 | 21 | 124 ± 18 | 22 |
| | | | | | | | | | | | DBP | 78 ± 13 | 21 | 74 ± 12 | 22 |
| Citrulline | | | | | | | | | | | | | | | |
| Powers (2015) [94] | Human (USA) | Obese | Normal | 85 | NA | I; T | 0.04 c | 16–19 wks | NA (office) | Wk 19 | SBP | 108 ± 4 | 12 | 116 ± 6 | 12 |
| | | | | | | | | | | | DBP | 66 ± 7 | 12 | 72 ± 6 | 12 |
| Cysteine | | | | | | | | | | | | | | | |
| Motawei (2016) [133] | Human (Egypt) | PE | PE | 85 | NA | I; T | 0.01 | Mo 5-term (<4 mo) | NA (office) | After 6 wks suppl. | SBP | 127 ± 8 | 50 | 133 ± 7 | 50 |
| Choline | | | | | | | | | | | | | | | |
| Zhang (2018) [112] | Rat (SD) | LPS infusion | FGR | 0.20 | NA | C; P | 0.88 d | 1–20 | Tail cuff (restrained) | 18 | SBP | 116 ± 2 | 9 | 130 ± 3 | 9 |
| | | Control | Normal | 0.20 | NA | C; P | 0.88 d | 1–20 | Tail cuff (restrained) | 18 | SBP | 104 ± 2 | 6 | 103 ± 5 | 6 |
| Jacobson (2018) [138] | Human (South Africa) | Alcohol | FGR | 57 | NA | I; T | 0.03 | wk 19-P0 | Sphygmo-manometer (office) | 12 wks after start | MAP | 81 ± 9 | 34 | 85 ± 0 | 35 |
Table A7.
Data extraction on development of pregnancy complication in risk population in human studies.
Table A7.
Data extraction on development of pregnancy complication in risk population in human studies.
| AA | Author (Year) | Country | Daily Dose (g/kgBW) | Pregnancy Complication | Definition | AA | CON |
|---|
| N Yes (%) | N No (%) | N Yes (%) | N No (%) |
|---|
| Arginine | Dera (2007) [95] | Poland | 0.04 a | SGA | BW < p10 | 24 (57) | 18 (43) | 15 (52) | 13 (48) |
| | Camarena Pulido (2016) [78] | Mexico | 0.04 a | SGA | Undefined | 1 (2) | 48 (98) | 3 (6) | 44 (94) |
| | | | | PE | ≥140/90 mmHg b | 3 (6) | 46 (94) | 11 (23) | 36 (77) |
| | | | | Preterm | Undefined | 1 (2) | 48 (98) | 7 (15) | 40 (85) |
| | | | | GDM | Undefined | 2 (4) | 47 (96) | 2 (4) | 45 (96) |
| | Facchinetti (2007) [79] | Italy | 0.11 a | SGA | BW < 2500 g | 11 (28) | 28 (72) | 15 (43) | 20 (57) |
| | | | | PE | ≥140/90 mmHg + proteinuria > 300 mg/24 h | 3 (11) | 24 (89) | 7 (37) | 12 (63) |
| | | | | Preterm | GA < 37 wks | 13 (16) | 67 (84) | 18 (25) | 51 (74) |
| | Neri (2010) [83] | Italy | 0.05 a | SGA | BW < p10 | 7 (19) | 32 (81) | 10 (25) | 30 (75) |
| | | | | SGA | BW < 2500 g | 7 (18) | 32 (82) | 11 (28) | 29 (72) |
| | | | | SGA | BW < 1500 g | 1 (3) | 38 (97) | 5 (13) | 35 (88) |
| | | | | Preterm | GA < 37 wks | 10 (26) | 29 (74) | 14 (35) | 26 (65) |
| | | | | Preterm | GA < 34 wks | 2 (5) | 37 (95) | 7 (18) | 33 (82) |
| | Rytlewski (2006) [85] | Poland | 0.04 | SGA | BW < p10 | 7 (23) | 23 (77) | 14 (45) | 17 (55) |
| | Rytlewski (2008) [86] | Poland | 0.04 a | SGA | EFW < p10 | 8 (32) | 17 (68) | 8 (40) | 12 (60) |
| | Sieroszewski (2004) [87] | Poland | 0.04 a | SGA | BW < p10 | 23 (29) | 55 (71) | 22 (75) | 8 (25) |
| | Valdivia-silva (2009) [96] | Mexico | 0.04 | SGA | BW < p10 | 6 (6) | 44 (94) | 14 (30) | 32 (70) |
| Cysteine (NAC) | Roes (2006) [134] | The Netherlands | 0.03 a | SGA | BW < p10 | 7 (37) | 12 (63) | 9 (47) | 10 (52) |
| | Shahin (2009) [137] | Egypt | 0.01 a | Preterm | GA < 36 wks | 8 (6) | 132 (94) | 62 (44) | 78 (56) |
| | | | | SGA | Undefined | 4 (3) | 136 (97) | 12 (9) | 128 (91) |
| Choline | Jacobson (2018) [138] | South Afrika | 0.03 a | SGA | BW < 2500 g | 8 (25) | 25 (75) | 10 (32) | 21 (68) |
| | | | | PE | Undefined | 2 (6) | 32 (94) | 3 (9) | 32 (91) |
| | | | | PIH | Undefined | 3 (9) | 31 (91) | 4 (11) | 31 (89) |
| | | | | GDM | Undefined | 4 (12) | 30 (88) | 0 (0) | 35 (100) |
| | Ross (2013) [135] | USA | 0.01 a | SGA | Undefined | 7 (15) | 39 (85) | 1 (2) | 46 (98) |
| | | | | PIH | Undefined | 11 (24) | 35 (76) | 3 (6) | 44 (94) |
| | | | | Preterm | Undefined | 9 (20) | 37 (80) | 7 (15) | 40 (85) |
| | | | | GDM | Undefined | 3 (7) | 43 (93) | 2 (4) | 45 (96) |
| | | | | LGA | Undefined | 2 (4) | 43 (96) | 1 (2) | 45 (98) |
Table A8.
Data extraction included animal and human studies on gestational weight gain.
Table A8.
Data extraction included animal and human studies on gestational weight gain.
| Author (Year) | Species (Strain or Country) | Animal Model | Pregnancy Complication | Mean Maternal Weight (kg)(kg) | Dietary Protein Intake (%) | Supplementation Scheme a; Intervention Type b | Supplementation Duration (GD) | Daily Dose (g/kg Body wt) | Measurement Day (GD) | AA Gestational Weight Gain (g) | n | CON Gestational Weight Gain (g) | n |
|---|
| Arginine | | | | | | | | | | | | | |
| Greene (2011) [33] | Mouse (FNVB/N × vegfr2-luc) | Control | Normal | 0.03 | 24 | C; T c | 1–18 | 2.660 d | 12–18 | 7 ± 1 | 6 | 6 ± 1 | 5 |
| Bourdon (2016) [44] | Rat (SD) | Protein restriction | FGR | 0.29 | 4 | C; T c | 7–21 | 1.500 | 7–21 | 42 ± 17 | 9 | 42.5 ± 17 | 9 |
| Bursztyn (2003) [45] | Rat (Wistar) | Hyper-insulinemia | FGR/PIH | 0.24 | NA | C; T | 11-P0 | 0.217 | 0–22 | 88 ± 40 | 5 | 69 ± 35 | 5 |
| Chin (1991) [46] | Rat (Fisher F344/NTacfBR) | Control | Normal | 0.17 | NA | C; P c | 1-P0 | 0.159 | 1-P0 | 18 ± 41 | 4 | 23 ± 41 | 4 |
| Podjarny (1993) [90] | Rat (Wistar) | Control | Normal | 0.23 | 20 | C; T | 12–22 | 0.322 | 0–22 | 151 ± 25 | 10 | 161 ± 43 | 10 |
| | | Adriamycin nephropathy | Risk | 0.23 | 20 | C; T | 12–22 | 0.322 | 0–22 | 72 ± 23 | 10 | 76 ± 22 | 10 |
| Podjarny (2001a) [49] | Rat (Wistar) | Hyper-insulinemia | PIH | 0.24 | 20 | C; P | 11–22 | 0.220 | 0–22 | 78 ± 7 | 15 | 66 ± 8 | 20 |
| Podjarny (2001b) [91] | Rat (Wistar) | Control | Normal | 0.23 | 20 | C; P | 11–22 | 0.260 | 0–22 | 83 ± 11 | 4 | 86 ± 7 | 4 |
| | | L-NAME | FGR/PE | 0.23 | 20 | C; T | 11–22 | 0.260 | 0–22 | 78 ± 15 | 8 | 64 ± 15 | 8 |
| Vosatka (1998) [36] | Rat (Wistar) | Hypobaric hypoxia | FGR | 0.25 | 4 g/day | C; P | 9–21 | 2.000 d | 9–21 | 59 ± 10 | 7 | 39 ± 10 | 10 |
| Zeng (2008) [37] | Rat (SD) | Control | Normal | 0.23 | 22 | C; P c | 0-P0 | 0.216 | 1–21 | 85 ± 22 | 12 | 79 ± 23 | 12 |
| | | | | | | | 1–7 | 0.216 | 1–21 | 75 ± 16 | 20 | 68 ± 17 | 20 |
| Peine (2018) [53] | Sheep (Rambouillet-cross) | Diet restricted | FGR | 68 | 16 | C; P | 54-P0 | 0.18 | 54–152 | −7760 ± 2388 | 10 | −7230 ± 2504 | 11 |
| Sun (2017) [54] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | NA | C; P | 35–110 | 0.250 | 35–110 | 2600 ± 489 | 8 | 2600 ± 369 | 8 |
| Sun (2018) [55] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | 10–14 | C; T | 35–110 | 0.249 | 1–110 | 4494 ± 3225 | 8 | 3983 ± 2518 | 8 |
| Bérard (2010) [59] | Pig (Swiss Large White) | Control | Normal | 159 | NA | C; P | 14–28 | 0.160 | 0–75 | 54,300 ± 9648 | 10 | 48,700 ± 9648 | 7 |
| Bass (2017) [58] | Pig (GPK-35) | Control | Normal | 180 | 19 | C; P c | 93-P0 | 0.150 | 93–110 | 14,400 ± 6580 | 49 | 12,000 ± 6443 | 48 |
| | | Nulliparous | Normal | 165 | 12 | C; P c | 81–116 | 0.170 | 93–110 | 15,000 ± 6103 | 19 | 12,700 ± 64,416 | 21 |
| | | Primiparous | Normal | 165 | 12 | C; P c | 81–116 | 0.170 | 93–110 | 17,400 ± 5889 | 12 | 11,800 ± 6129 | 13 |
| | | Multiparous | Normal | 165 | 12 | C; P c | 81–116 | 0.170 | 93–110 | 11,500 ± 5940 | 18 | 12,700 ± 5613 | 14 |
| Dallanora (2017) [69] | Pig (Landrace × Large white) | Control | Normal | 148 | 17 | C; P | 25–112 | 0.115 | 80–112 | 22,900 ± 32,446 | 51 | 20,300 ± 32,446 | 51 |
| Li (2010) [74] | Pig (Yorkshire × Landrace) | Control | Normal | 113 | 12 | C; P c | 0–25 | 0.070 | 0–25 | 4900 ± 15,973 | 9 | 7000 ± 15,973 | 9 |
| | | | | | | | 0–25 | 0.140 | 0–25 | 2700 ± 15,973 | 8 | 2700 ± 15,973 | 9 |
| Li (2014) [75] | Pig (Yorkshire × Landrace) | Control | Normal | 115 | 12 | C; Pc | 14–25 | 0.070 | 0–25 | 1100.0 ± 65,557 | 15 | 1100 ± 6557 | 14 |
| | | | | | | | 14–25 | 0.140 | 0–25 | 1200.0 ± 6557 | 14 | 1100 ± 6557 | 14 |
| Li (2015) [76] | Pig (Landrace) | Control | Normal | 125 | 16 | C; P c | 1–30 | 0.21 | 1-P0 | 53,920 ± 1098 | 32 | 52,100 ± 10,980 | 30 |
| | | | | 187 | 16 | C; P c | 1–30 | 0.14 | 1-P0 | 47,800 ± 10,980 | 57 | 47,430 ± 10,980 | 56 |
| | | | | 196 | 16 | C; P c | 1–30 | 0.13 | 1-P0 | 46,380 ± 8964 | 37 | 45,640 ± 8964 | 37 |
| Dera (2007) [95] | Human (Poland) | FGR < p10; gestational HTN | Risk | 70 | NA | I’T | Until P0 | 0.043 d | Entry-P0 | 11,630 ± 4830 | 42 | 10,070 ± 3500 | 27 |
| Citrulline | | | | | | | | | | | | | |
| Bourdon (2016) [44] | Rat (SD) | Protein restriction | FGR | 0.29 | 4 | C; T | 7–21 | 2.000 | 7–21 | 54 ± 16 | 9 | 43 ± 17 | 9 |
| Glutamate (NCG) | | | | | | | | | | | | | |
| Navarro (2005) [148] | Rat (Wistar) | Control | Normal | 0.26 | NA | C; P | 2–20 | 0.110 | 2–20 | 116 ± 21 | 11 | 132 ± 16 | 7 |
| Sun (2017) [54] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | NA | C; P | 35–110 | 0.125 | 35–110 | 4370 ± 433 | 8 | 2550 ± 369 | 8 |
| Sun (2018) [55] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | 10–14 | C; T | 35–110 | 0.062 | 35–110 | 4200 ± 2605 | 8 | 2600 ± 369 | 8 |
| Zhu (2015) [149] | Pig (Landrace × Yorkshire) | Control | Normal | 132 | NA | C; P | 0–28 | 0.008 | 0–28 | 21,000 ± 7483 | 7 | 16,000 ± 10,198 | 8 |
| Cai (2018) [66] | Swine (Landrace × Yorkshire) | Control | Normal | 210 | 13 | C; P | 1–8 | 0.003 | 0–28 | 7810 ± 515 | 18 | 8390 ± 515 | 18 |
| | | | | | | | 9–28 | 0.003 | 0–28 | 8390 ± 515 | 16 | 8390 ± 515 | 18 |
| | | | | | | | 1–28 | 0.003 | 0–28 | 9350 ± 515 | 17 | 8390 ± 515 | 18 |
| Glutamine | | | | | | | | | | | | | |
| Zhu (2018) [67] | Pig (Landrace × Large White) | Control | Normal | 272 | NA | C; Pc | 85–114 | 0.126 | 84–110 | 40,700 ± 39,894 | 30 | 42,800 ± 31,773 | 30 |
| Aspartatic acid | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 e | 10–20 | 24 ± 22 | 4 | 30 ± 8 | 4 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 e | 10–20 | 52 ± 18 | 4 | 44 ± 10 | 4 |
| Valine | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 e | 10–20 | 39 ± 11 | 3 | 30 ± 8 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 16 ± 4 | 3 | 30 ± 18 | 12 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 e | 10–20 | 50 ± 17 | 3 | 44 ± 10 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 32 ± 2 | 3 | 55 ± 42 | 12 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | NA | C; P | 1–22 | 2.71 | 1–22 | 40 ± 16 | 5 | 26 ± 12 | 5 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 12–22 | 95 ± 16 | 6 | 93 ± 34 | 9 |
| Leucine | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 e | 10–20 | 21 ± 22 | 4 | 30 ± 8 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 57 ± 6 | 3 | 29 ± 18 | 12 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 e | 10–20 | 55 ± 28 | 4 | 44 ± 10 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 50 ± 17 | 3 | 55 ± 42 | 12 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.18 | NA | C; P | 1–22 | 1.500 | 1–22 | −7.0 ± 11 | 5 | −20 ± 17 | 7 |
| Mawatari (2004) [102] | Rat (SD) | Control | Normal | 0.30 | NA | I; P | 7–17 | 0.300 | 0–20 | 178 ± 31 | 19 | 158 ± 43 | 19 |
| | | | | | | | | 1.000 | 0–20 | 179 ± 32 | 20 | 160 ± 43 | 19 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 12–22 | 82 ± 25 | 5 | 93 ± 34 | 9 |
| Ventrucci (2001) [101] | Rat (Wistar) | Control | Normal | 0.15 | 18 | C; P | 1–20 | 7.180 e | 1–20 | 61 ± 31 | 10 | 90 ± 36 | 10 |
| Isoleucine | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 e | 10–20 | 42 ± 19 | 3 | 30 ± 8 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 32 ± 4 | 3 | 29 ± 18 | 12 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 e | 10–20 | 66 ± 28 | 4 | 44 ± 10 | 4 |
| | | | | | | | | 4.720 e | 10–20 | 23 ± 19 | 3 | 55 ± 42 | 12 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | 6 | C; P | 1–22 | 3.150 | 1–22 | 56 ± 11 | 5 | 44 ± 22 | 5 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 12–22 | 92 ± 9 | 4 | 93 ± 34 | 9 |
| Cysteine (NAC) | | | | | | | | | | | | | |
| Miller (2014) [108] | Mouse (CD-1) | Control | Normal | 0.03 | 19 | I; P | 6–13 | 0.150 | 6–17 | 14 ± 3 | 25 | 13 ± 3 | 25 |
| Soto-Blanco (2001) [114] | Rat (Wistar) | Control | Normal | 0.27 | 26 | C; P | 6–21 | 0.510 | 6–21 | 72 ± 16 | 12 | 68 ± 19 | 12 |
| Methionine | | | | | | | | | | | | | |
| Brunner (1978) [43] | Rat (SD) | PKU-induction | FGR | 0.22 | NA | C; T | 10–20 | 1.570 e | 10–20 | 23 ± 4 | 4 | 30 ± 8 | 4 |
| | | Control | Normal | 0.22 | NA | C; P | 10–20 | 1.570 e | 10–20 | 44 ± 22 | 4 | 44 ± 10 | 4 |
| Matsueda (1982) [97] | Rat (SD) | Control | Normal | 0.19 | NA | C; P | 1–22 | 1.150 | 1–22 | −54 ± 9 | 5 | −20 ± 17 | 7 |
| Mori (1999) [98] | Rat (SD) | Control | Normal | 0.19 | 20 | I; P | 12–22 | 2.368 | 12–22 | −16 ± 36 | 8 | 93 ± 34 | 9 |
| Liu (2016) [125] | Sheep (Merino) | Control | Normal | 63 | NA | I; P | 111-P0 | 0.047 | 111–1 wk pre-lambing | −1100 ± 1200 | 60 | 1500 ± 1200 | 60 |
| Choline | | | | | | | | | | | | | |
| Bai (2012) [109] | Rat (Wistar) | Protein restriction | FGR | 0.30 | 9 | C; P | 1-P0 | 0.670 d | 1–22 | 57 ± 7 | 8 | 59 ± 9 | 8 |
| Zhang (2018) [112] | Rat (SD) | LPS infusion | FGR | 0.20 | NA | C; P | 1–20 | 0.875 e | 1–20 | 175 ± 46 | 9 | 170 ± 24 | 9 |
| Janovick Guretzky (2006) [151] | Cow (Holstein) | Control | Normal | 715 | NA | C; P | 3 wks before P0 | 0.021 | Last 2 wks | 32,500 ± 49,041 | 5 | 5600 ± 58,138 | 5 |
| | Cow (Jersey) | Control | Normal | 506 | NA | C; P | 3 wks before P0 | 0.030 | Last 2 wks | 18,500 ± 47,518 | 16 | 12,800 ± 49,115 | 16 |
| Jacobson (2018) [138] | Human (South Africa) | Alcohol use | Risk | 57 | NA | I; T | Enrollment-P0 | 0.035 | NA | 5960 ± 19,040 | 28 | 2020 ± 19,463 | 24 |
| Ross (2013) [135] | Human (USA) | Control | Normal | 78 | NA | I; P | wk 17-P0 | 0.012 d | NA | 14,832 ± 8165 | 46 | 14,787 ± 6713 | 47 |
| Yan (2012) [136] | Human (USA) | Control | Normal | 63 | NA | I; P | wk 27–39 | 0.007 d | 189–273 | 6000 ± 2200 | 13 | 6400 ± 2700 | 13 |
Table A9.
Data extraction of included animal and human studies on maternal glucose metabolism.
Table A9.
Data extraction of included animal and human studies on maternal glucose metabolism.
| Author (Year) | Species (Strain) | Animal Model | Pregnancy Complication | Mean Maternal Weight (kg)(kg) | Dietary Protein Intake (%) | Supplementation Scheme a; Intervention Type b | Supplementation Duration (GD) | Daily Dose (g/kg Body wt) | Measurement Day (GD) | AA glucose ± SD (mmol/l) | n | CON Glucos ± SD (mmol/l) | n |
|---|
| Arginine | | | | | | | | | | | | | |
| Bursztyn (2003) [45] | Rat (Wistar) | Hyper-insulinemia | FGR/PIH | 0.24 | NA | C; T | 11-P0 | 0.217 | 22 | 4.6 ± 1.6 | 5 | 4.4 ± 1.1 | 5 |
| Podjarny (2001) [49] | Rat (Wistar) | Hyper-insulinemia | PIH | 0.24 | 20 | C; P | 11–22 | 0.220 | 22 | 3.7 ± 0.6 | 15 | 4.1 ± 1.0 | 20 |
| Zhang (2016b) [57] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | NA | I; P | 35–110 | 0.250 | 110 | 2.1 ± 1.0 | 8 | 1.7 ± 1.0 | 8 |
| Li (2010) [74] | Pig (Yorkshire × Landrace) | Control | Normal | 113 | 12 | C; P c | 0–25 | 0.070 | 25 | 4.1 ± 0.8 | 9 | 4.1 ± 0.8 | 9 |
| | | | | | | | | 0.140 | 25 | 4.0 ± 0.8 | 8 | 4.1 ± 0.8 | 9 |
| Liu (2012) [77] | Pig (Landrace × Large White) | Control | Normal | 187 | 15 | C; P | 1-P0 | 0.107 | | 3.8 ± 0.7 | 9 | 3.5 ± 0.7 | 9 |
| Glutamate (NCG) | | | | | | | | | | | | | |
| Zhang (2016) [57] | Sheep (Hu) | Diet restricted twin bearing | FGR | 40 | NA | I; P | 35–110 | 0.060 | 110 | 2.2 ± 1.0 | 8 | 1.7 ± 1.0 | 8 |
| Liu (2012) [77] | Pig (Landrace × Large White) | Control | Normal | 187 | 15 | I; P | 1-P0 | 0.011 | 110 | 3.9 ± 0.7 | 9 | 3.5 ± 0.7 | 9 |
| Glutamine | | | | | | | | | | | | | |
| Zhu (2018) [67] | Pig (Landrace × Large White) | Control | Normal | 272 | NA | C; P c | 85–114 | 0.126 | 100 | 4.0 ± 1.6 | 6 | 4.4 ± 1.6 | 6 |
| | | | | | | | | | 112 | 4.8 ± 1.6 | 6 | 4.7 ± 1.6 | 6 |
| Leucine | | | | | | | | | | | | | |
| Ventrucci (2002) [150] | Rat (Wistar) | Control | Normal | 0.15 | 18 | C; P | 1–20 | 7.180 e | 20 | 5.3 | 10 | 4.2 ± 0.7 | 10 |
| Cysteine (NAC) | | | | | | | | | | | | | |
| Moazzen (2014) [107] | Mouse (C57BL/6) | Streptozotocin | Pre-existent DM | 0.03 | NA | C; P | 0.5–18.5 | 1.000 | 18.5 | 23.9 ± 1.7 | 7 | 27.4 ± 5.3 | 7 |
| | | Control | Normal | 0.03 | NA | C; P | 0.5–18.5 | 1.000 | 18.5 | 8.3 ± 0.5 | 7 | 7.6 ± 1.3 | 7 |
| Lin (2011) [113] | Rat (SD) | High-fat diet | Over-growth | 0.23 | 17 | C; P | 1–19.5 | 0.075 d,e | 19.5 | 6.2 ± 1.1 | 20 | 4.5 ± 0.8 | 20 |
| Choline | | | | | | | | | | | | | |
| Zenobi (2018a) [132] | Cow (Holstein) | Excess energy diet | Risk overgrowth | 735 | 14 | C; T | Last 21 days | 0.024 | Mean of −12 and −7 prepartum | 3.7 ± 0.6 | 25 | 3.6 ± 0.6 | 22 |
| | | Maintenance energy diet | Normal | 735 | 14 | C; P | Last 21 days | 0.024 | Mean of −12 and −7 prepartum | 3.5 ± 0.6 | 21 | 3.6 ± 0.6 | 25 |
| Zenobi (2018b) [152] | Cow (Holstein) | Diet restricted | FGR | 732 | 10 | C; P | Last 64 days | 0.002 | 9 | 3.6 ± 0.7 | 15 | 3.5 ± 0.7 | 15 |
| | | | | | | | | 0.003 | 10 | 3.5 ± 0.7 | 14 | 3.5 ± 0.7 | 15 |
| | | | | | | | | 0.005 | 11 | 3.5 ± 0.7 | 14 | 3.5 ± 0.7 | 15 |
| | | | | | | | | 0.007 | 12 | 3.3 ± 0.7 | 17 | 3.5 ± 0.7 | 15 |
| | | Ad libitum diet | Normal | 732 | 15 | C; P | Last 64 days | 0.002 | 5 | 4.3 ± 0.5 | 15 | 4.2 ± 0.5 | 15 |
| | | | | | | | | 0.003 | 6 | 4.2 ± 0.5 | 14 | 4.2 ± 0.5 | 15 |
| | | | | | | | | 0.005 | 7 | 4.2 ± 0.5 | 14 | 4.2 ± 0.5 | 15 |
| | | | | | | | | 0.007 | 8 | 4.1 ± 0.5 | 17 | 4.2 ± 0.5 | 15 |
Table A10.
Quality assessment of included animal studies.
Table A10.
Quality assessment of included animal studies.
| Author (Year) | Any Randomization | Any Blinding | Sample Size Calculation | Conflict of Interest statement | Free of Experimental Unit of Analysis Errors | Random Group Allocation (Selection) | Groups Similar at Baseline (Selection) | Blinded Group Allocation (Selection) | Random Housing (Performance) | Blinded Interventions (Performance) | Random Outcome Ass. (Detection) | Blinded Outcome Ass. (Detection) | Reporting of Drop-Outs (Attrition) | Selective Outcome Reporting (Reporting) | Other Biases |
|---|
| Abdel-Wanhab (1999) [115] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Alexander (2004) [34] | N | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Altun (2008) [35] | N | N | N | N | Y | ? | ? | ? | H | H | ? | ? | ? | ? | ? |
| Bai (2012) [109] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Balansky (2009) [106] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Bass (2017) [58] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Batistel (2017) [128] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Beaudoin (1981) [40] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Bérard (2010) [59] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Birch (2016) [127] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Bourdon (2016) [44] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | H | ? | L |
| Brunner (1978) [43] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Bursztyn (2003) [45] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Cai (2018) [66] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | H | ? | L |
| Caporossi (2014) [42] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Chandrashekar (1977) [116] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Che (2013) [70] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Chin (1991) [46] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Clements (2017) [129] | Y | N | N | N | Y | ? | ? | ? | L | ? | ? | ? | L | ? | ? |
| Crane (2016) [52] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Cruz (2014) [99] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Cruz (2016) [100] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Da Costa (2014) [47] | Y | N | N | N | Y | ? | ? | ? | H | ? | ? | ? | L | ? | ? |
| Dallanora (2017) [69] | Y | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Fuji (1971) [117] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Gao (2012) [71] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Garbossa (2015) [72] | N | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | H | ? | ? |
| Gauthier (2009) [123] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Gonzalez-Anover (2017) [68] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | ? | ? | ? |
| Greene (2011) [33] | Y | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Guo (2016) [73] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Hashimoto (2012) [120] | N | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | L |
| Helmbrecht (1996) [48] | Y | Y | N | N | N | ? | ? | ? | ? | L | ? | L | ? | ? | ? |
| Herrera (2017) [121] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Jacometo (2016) [130] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Janovick Guretzky (2006) [151] | Y | N | N | N | Y | ? | H | ? | ? | ? | ? | ? | ? | ? | ? |
| Koeners (2007) [38] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Koz (2010) [118] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Li (2010) [74] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Li (2014) [75] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Li (2015) [76] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Lin (2011) [113] | N | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Liu (2012) [77] | Y | N | N | N | N | ? | L | ? | ? | ? | ? | ? | ? | ? | ? |
| Liu (2016) [125] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Madsen (2017) [60] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Mateo (2007) [61] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Mateo (2008) [62] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Matsueda (1982) [97] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | H | ? | ? |
| Mawatari (2004) [102] | N | N | N | N | Y | L | L | ? | ? | ? | ? | ? | H | ? | ? |
| Miller (2014) [108] | Y | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Moazzen (2014) [107] | N | N | N | Y | N | ? | L | ? | ? | ? | ? | ? | ? | ? | L |
| Mori (1999) [98] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Navarro (2005) [148] | N | N | N | N | N | ? | N | ? | ? | ? | ? | ? | ? | ? | ? |
| Othmani Mecif (2017) [124] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Peine (2018) [53] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Podjarny (2001a) [49] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | ? | ? | ? |
| Podjarny (2001b) [91] | N | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Podjarny (1993) [90] | N | N | N | N | N | ? | N | ? | ? | ? | ? | ? | ? | ? | ? |
| Podjarny (1997) [92] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | ? | ? | ? |
| Quesnel (2014) [64] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | H | ? | ? |
| Schooley (2002) [50] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | H | ? | ? |
| Sharkey (2001) [51] | Y | N | N | N | N | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Soto-Blanco (2001) [114] | N | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Sun (2018) [55] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Sun (2017) [54] | Y | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Thomas (2009) [110] | Y | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Thompson (2011) [122] | N | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Tran (2017) [39] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | L |
| Tsiplakou (2017) [126] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Ventrucci (2001) [101] | N | N | N | N | N | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Ventrucci (2002) [150] | N | N | N | Y | N | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Viana (2013) [103] | Y | N | N | N | Y | ? | ? | ? | ? | ? | L | ? | L | ? | ? |
| Viau (1973) [119] | N | N | N | N | Y | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Vosatka (1998) [36] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Wang (2018) [105] | Y | N | N | Y | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | L |
| Wu (2012) [63] | Y | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Xu (2017) [104] | N | N | N | N | Y | ? | L | ? | ? | ? | ? | ? | L | ? | ? |
| Xu (2018) [131] | N | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Yang (2000) [111] | Y | Y | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Zeng (2008) [37] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Zeng (2012) [41] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Zenobi (2018a) [132] | Y | Y | N | N | Y | ? | ? | ? | ? | ? | ? | ? | H | ? | ? |
| Zenobi (2018b) [152] | Y | N | N | N | N | ? | ? | ? | ? | ? | ? | ? | L | ? | ? |
| Zhang (2014) [65] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Zhang (2016a) [56] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Zhang (2016b) [57] | Y | N | N | Y | N | ? | L | ? | ? | ? | ? | ? | L | ? | L |
| Zhang (2018) [112] | Y | N | N | Y | N | ? | ? | ? | ? | ? | ? | ? | L | ? | L |
| Zhu (2018) [67] | Y | N | N | Y | Y | ? | L | ? | ? | ? | L | ? | L | ? | L |
| Zhu (2015) [149] | Y | N | N | Y | Y | ? | L | ? | ? | ? | ? | ? | L | ? | L |

 ,
, 


















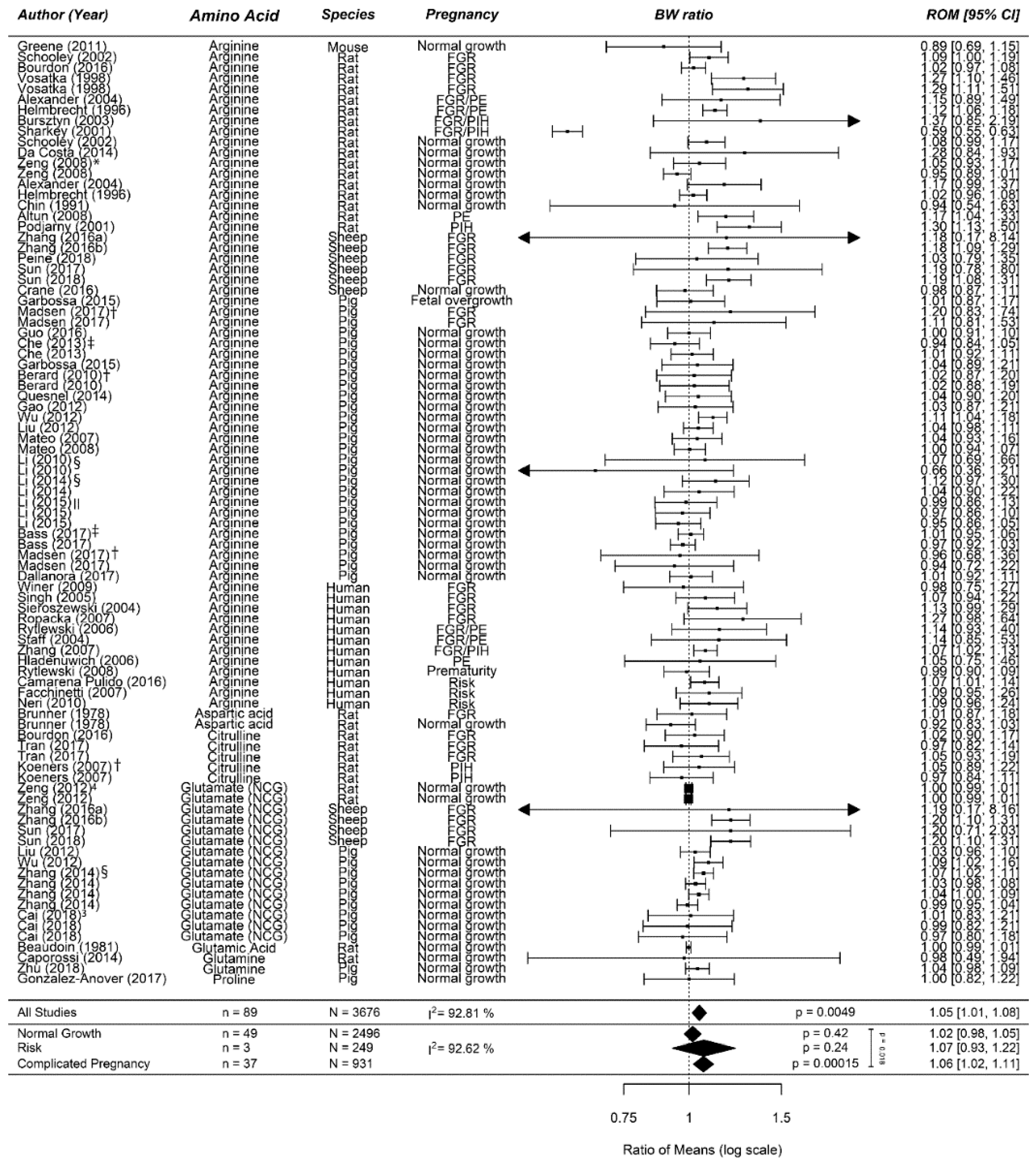{kind=link}
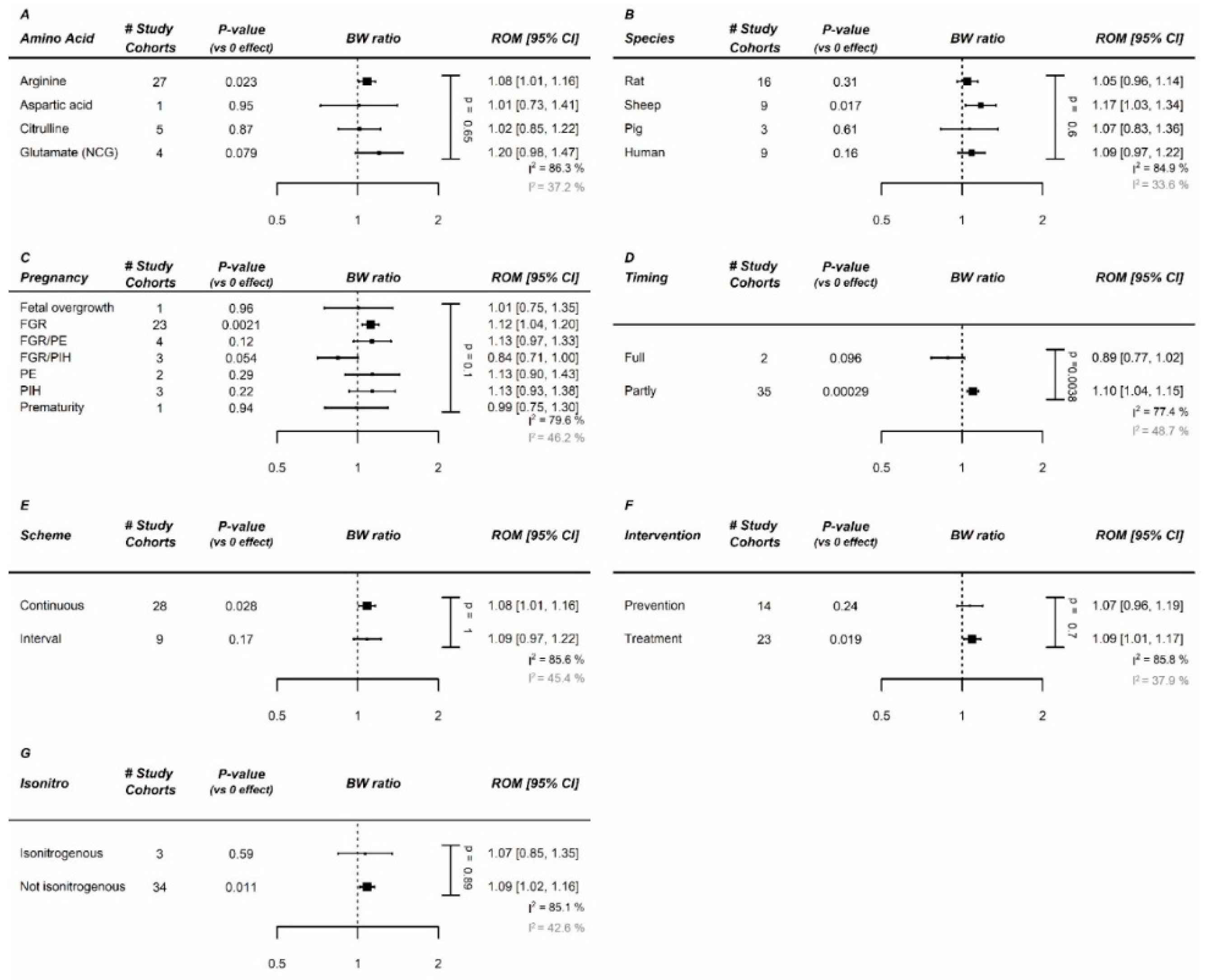{kind=link}
{kind=link}
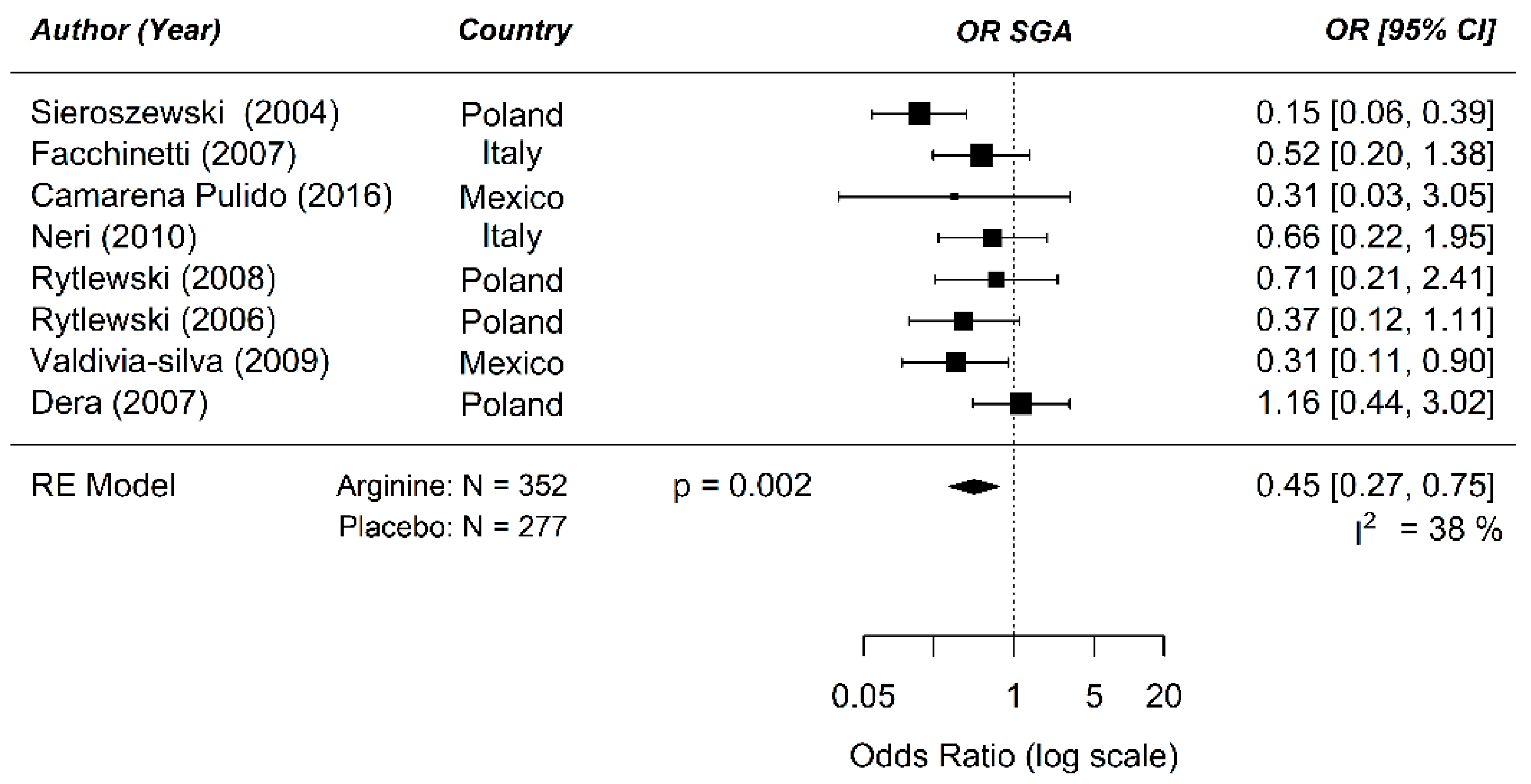{kind=link}
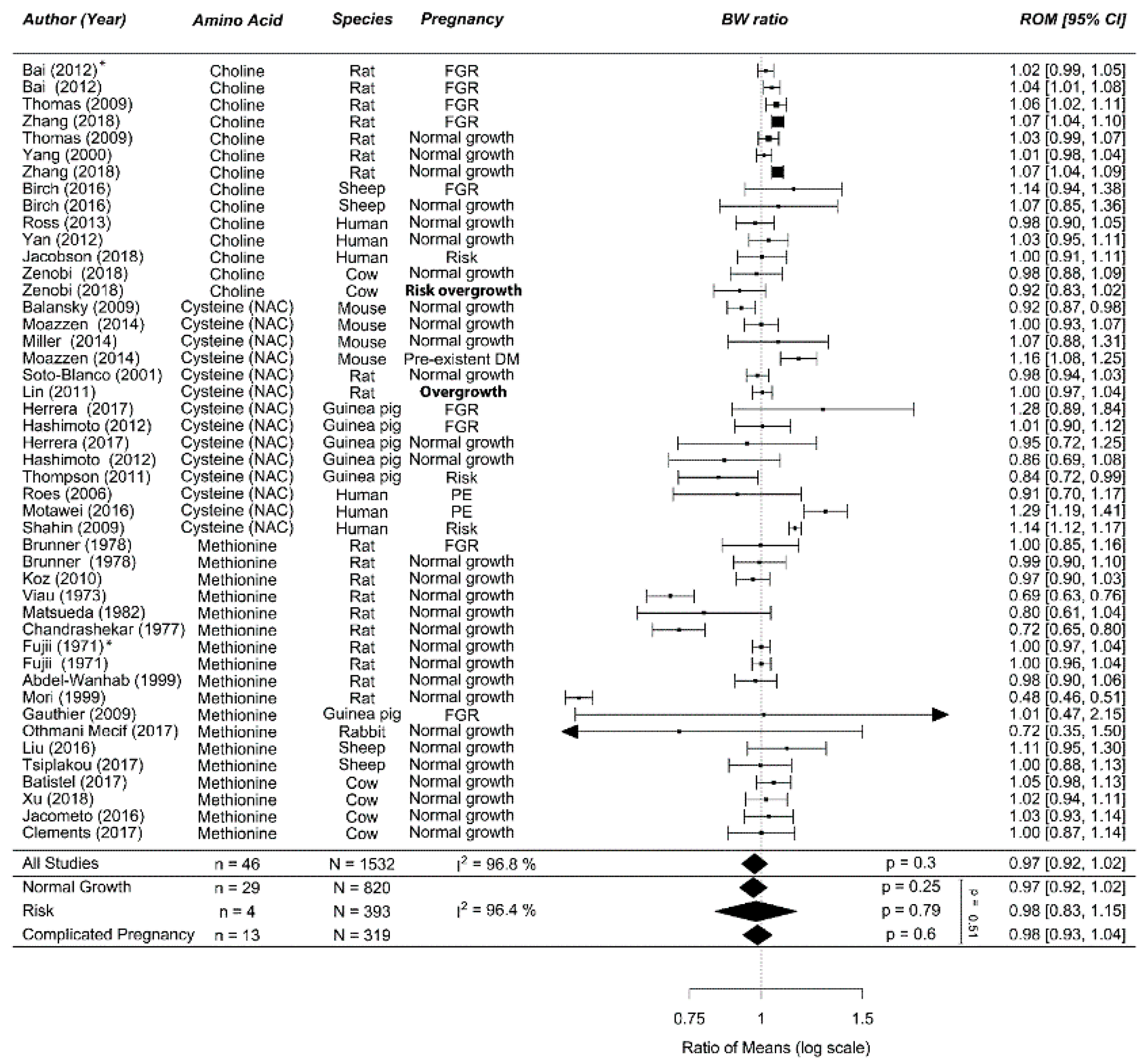{kind=link}
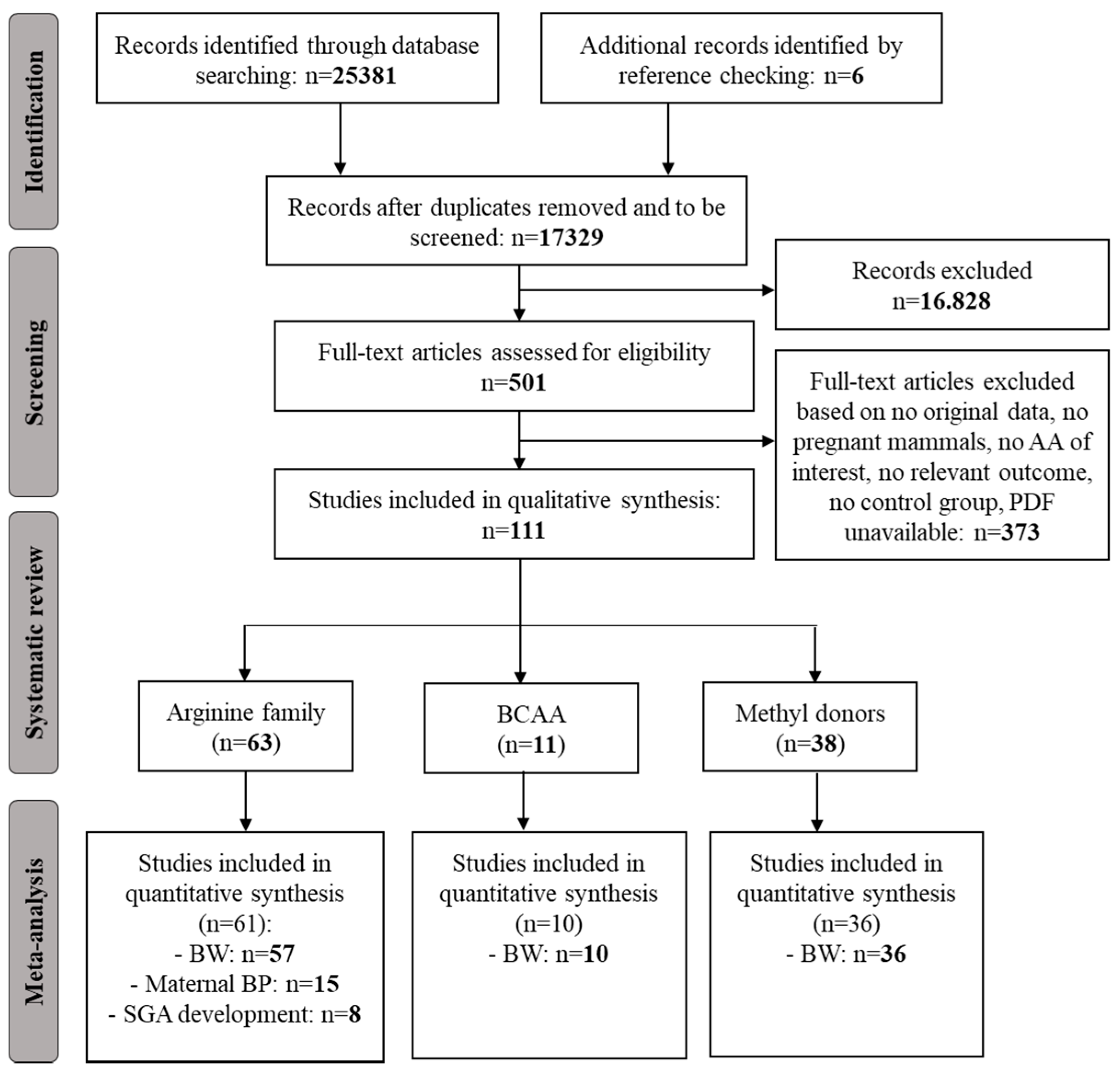{kind=link}
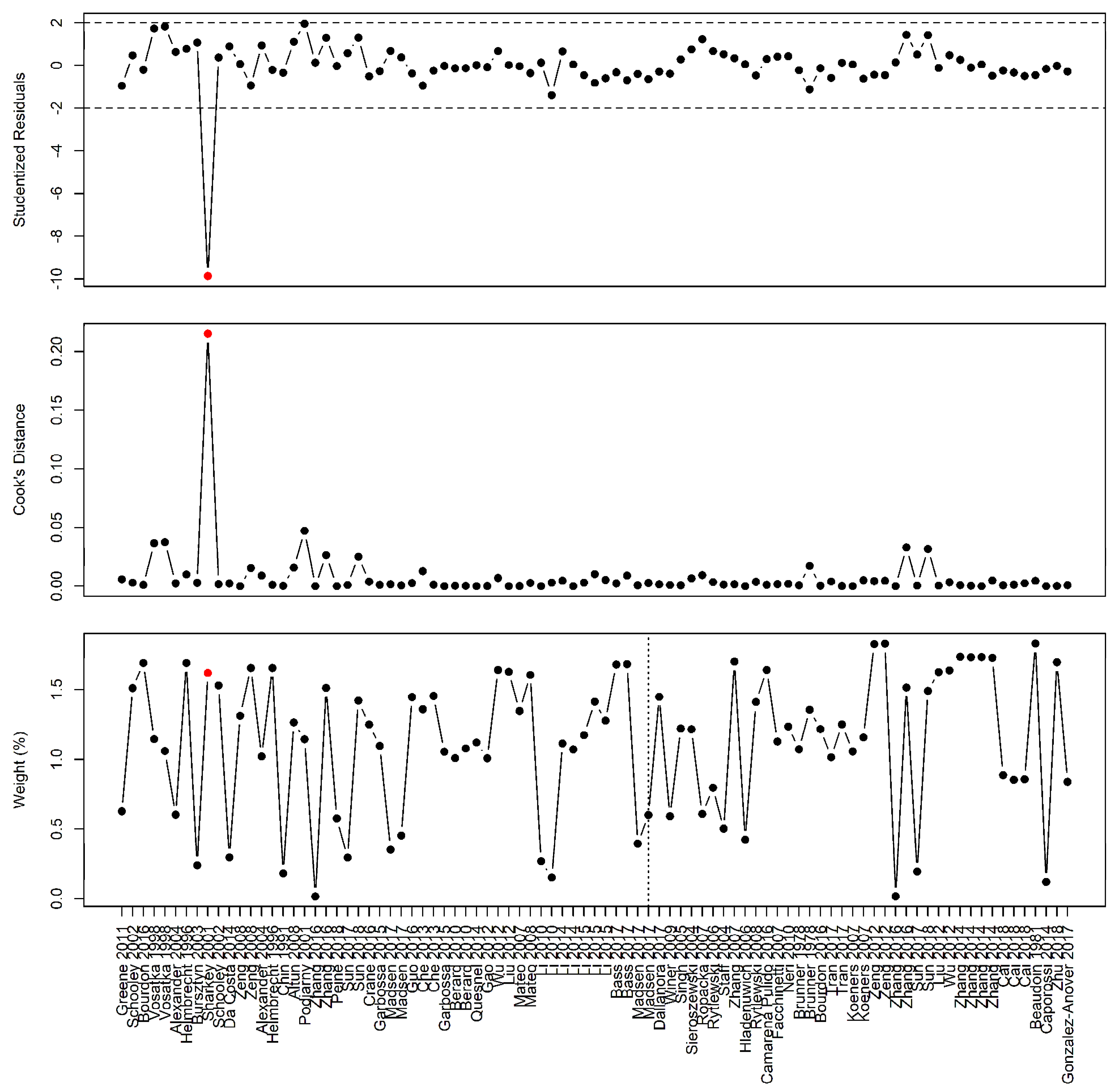{kind=link}
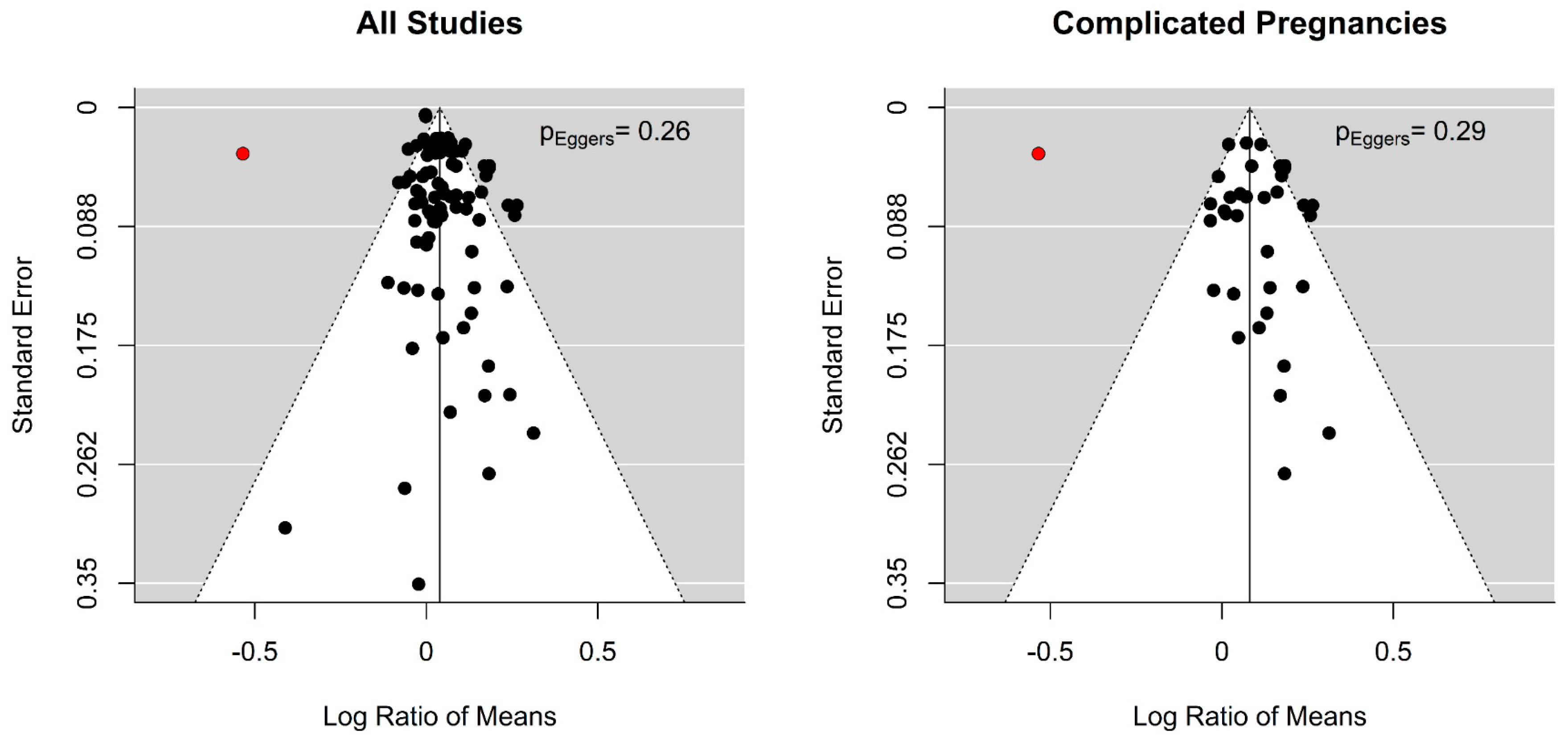{kind=link}
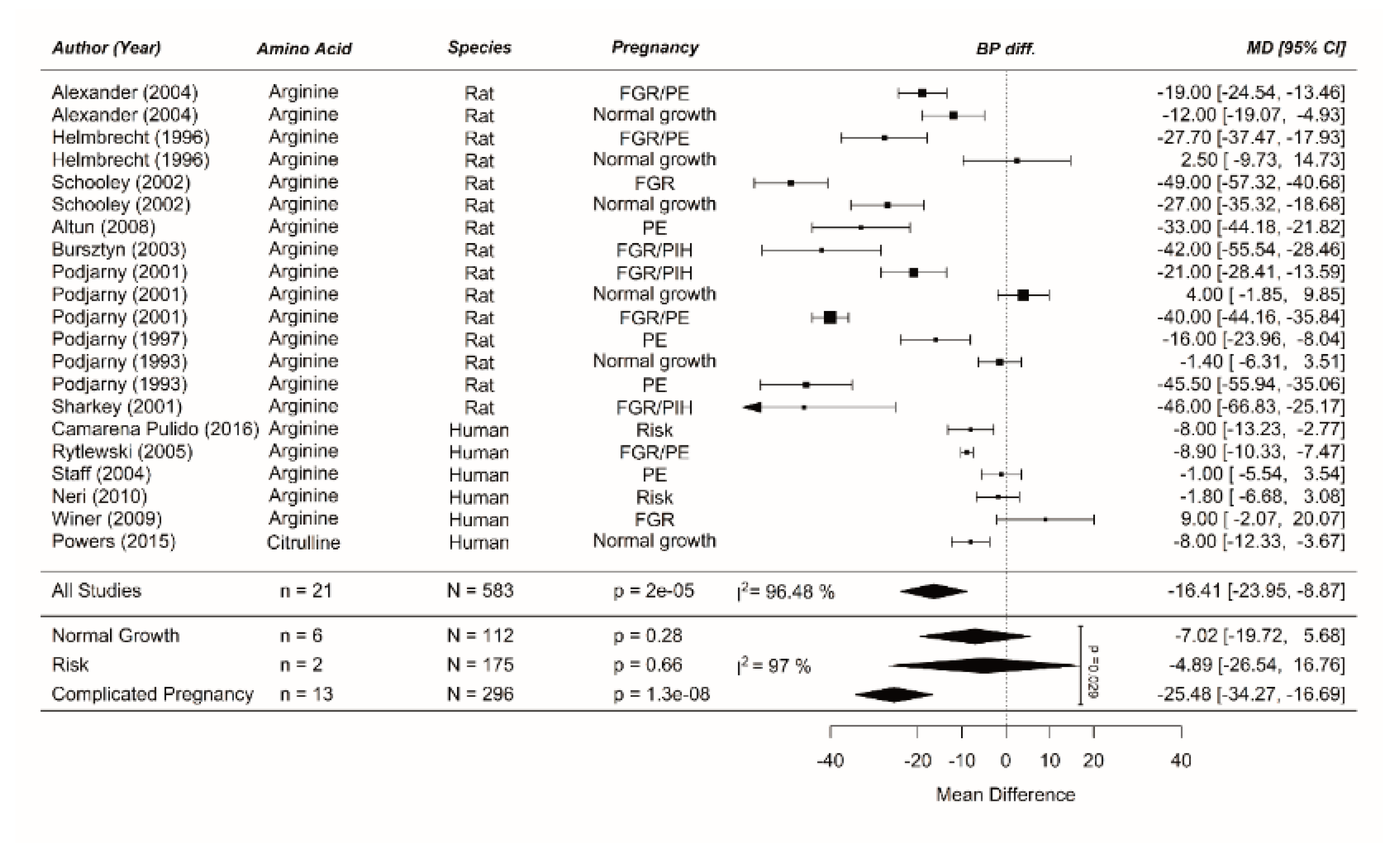{kind=link}
{kind=link}
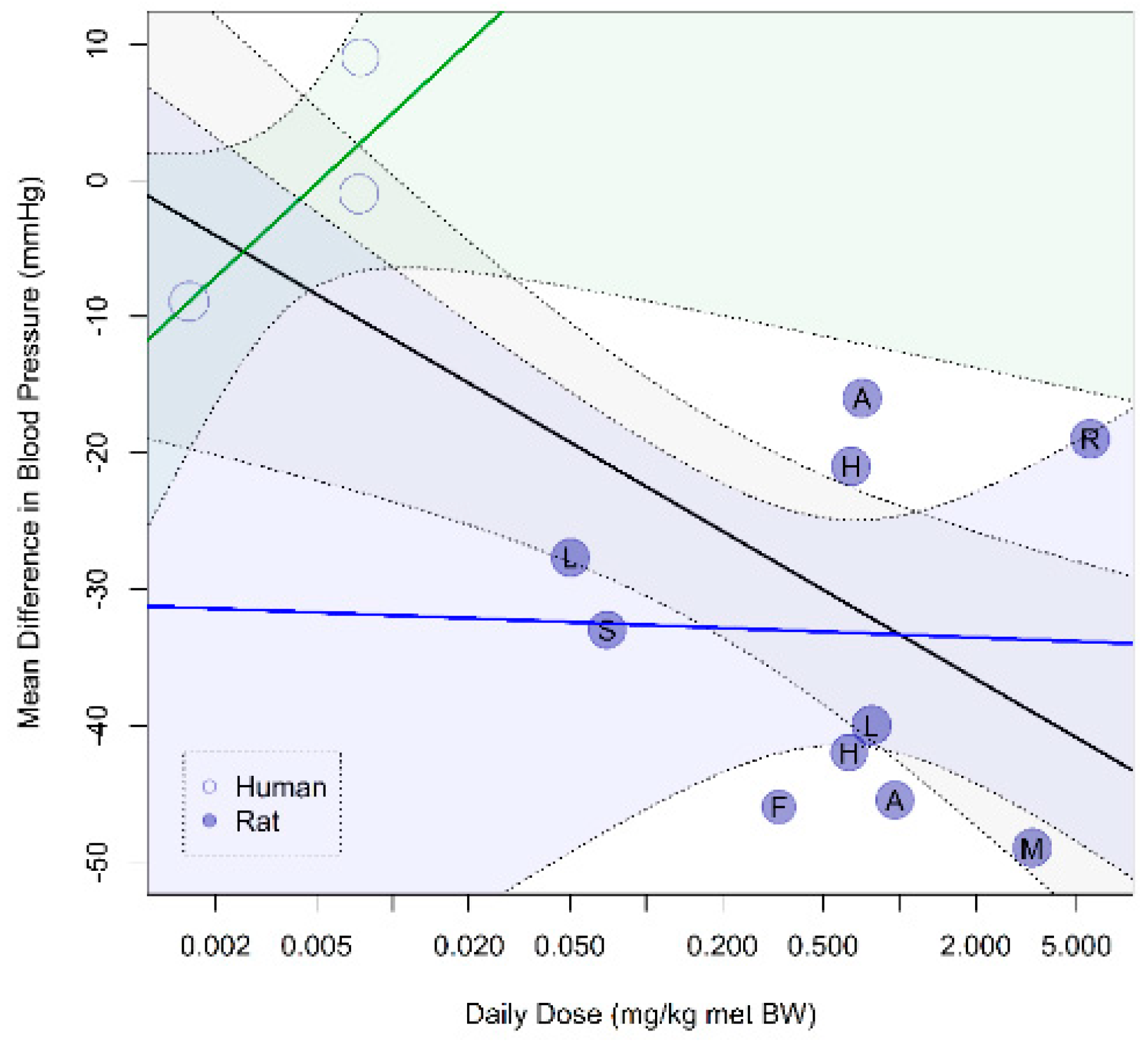{kind=link}
{kind=link}
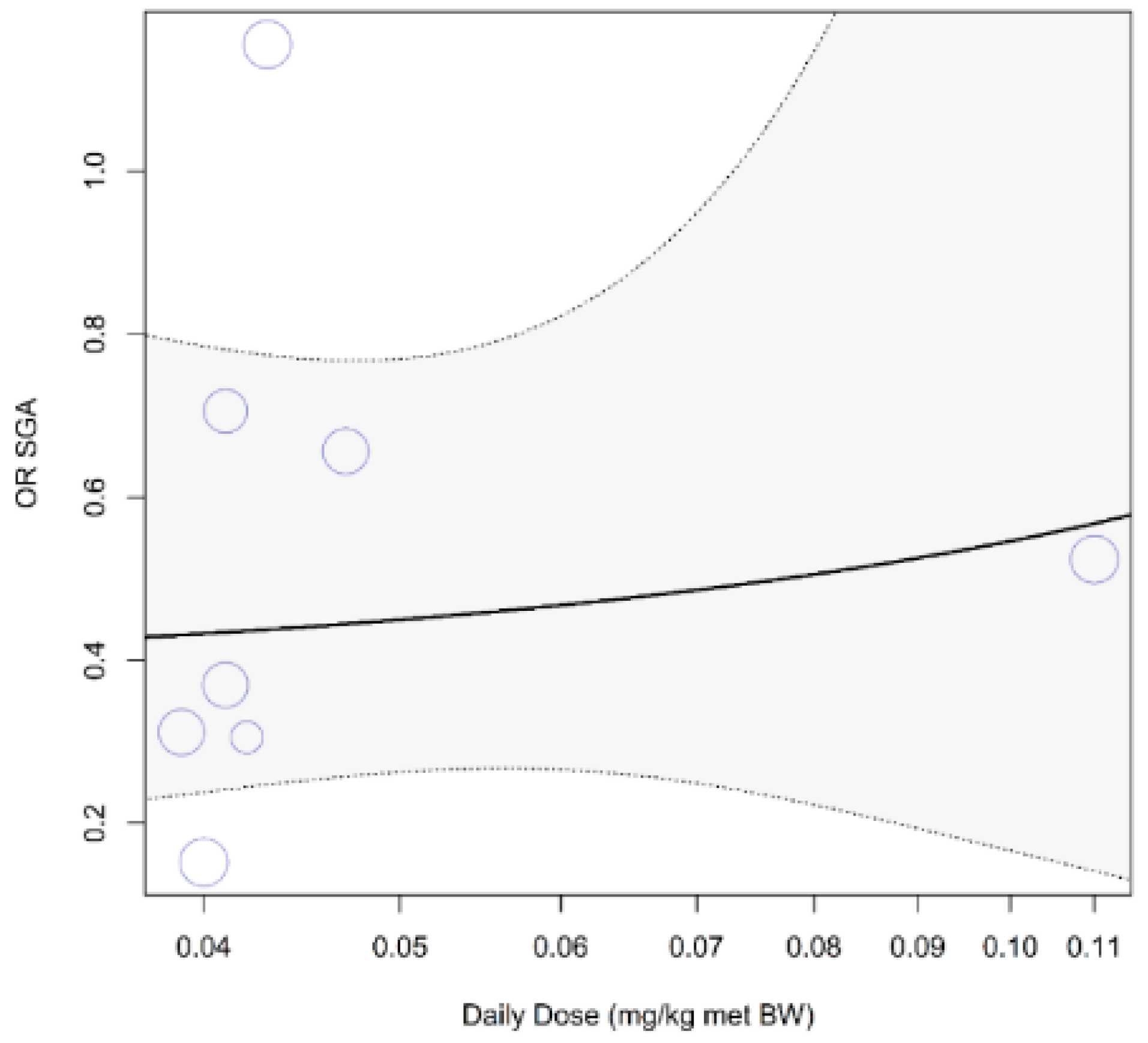{kind=link}
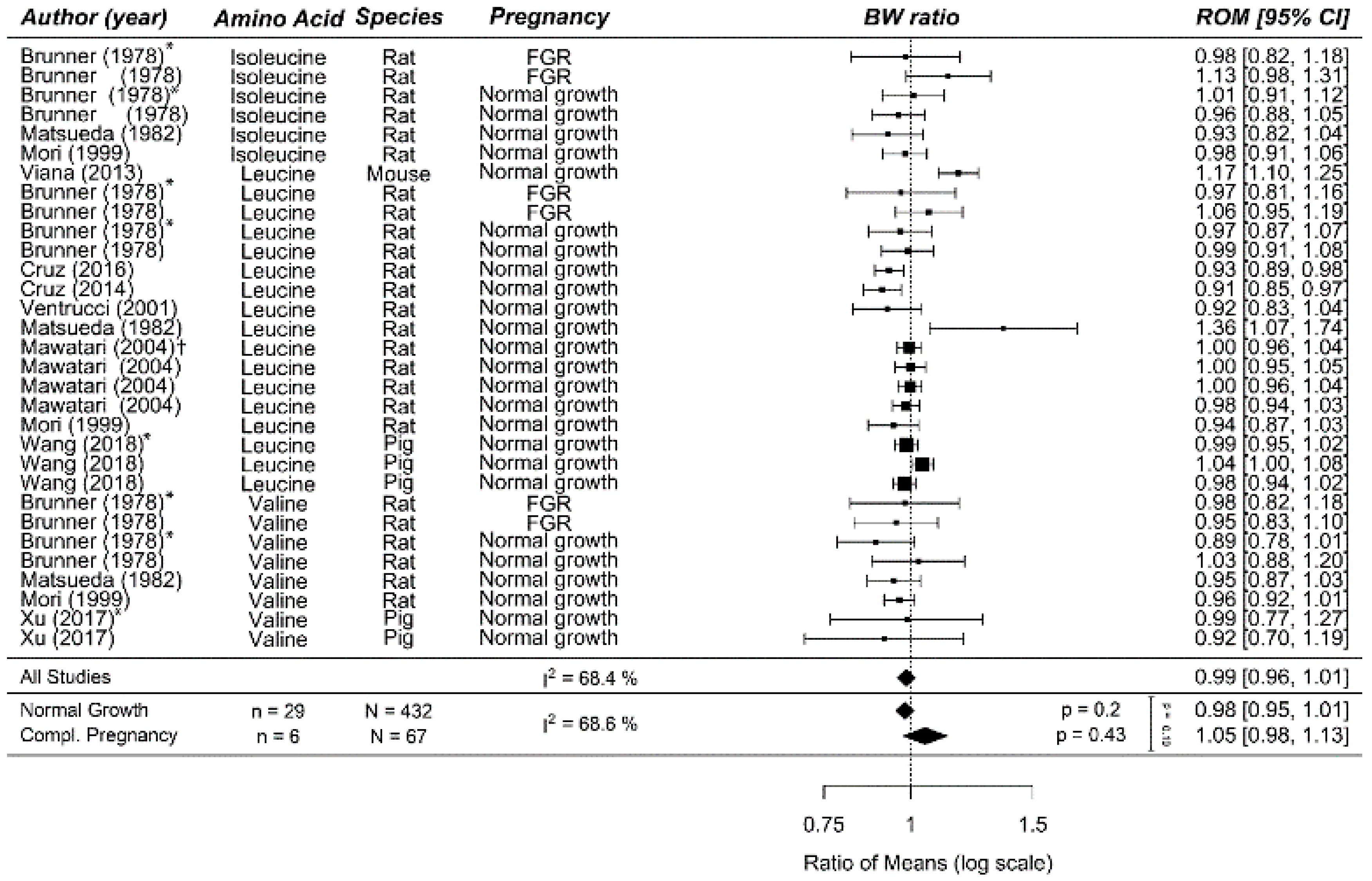{kind=link}
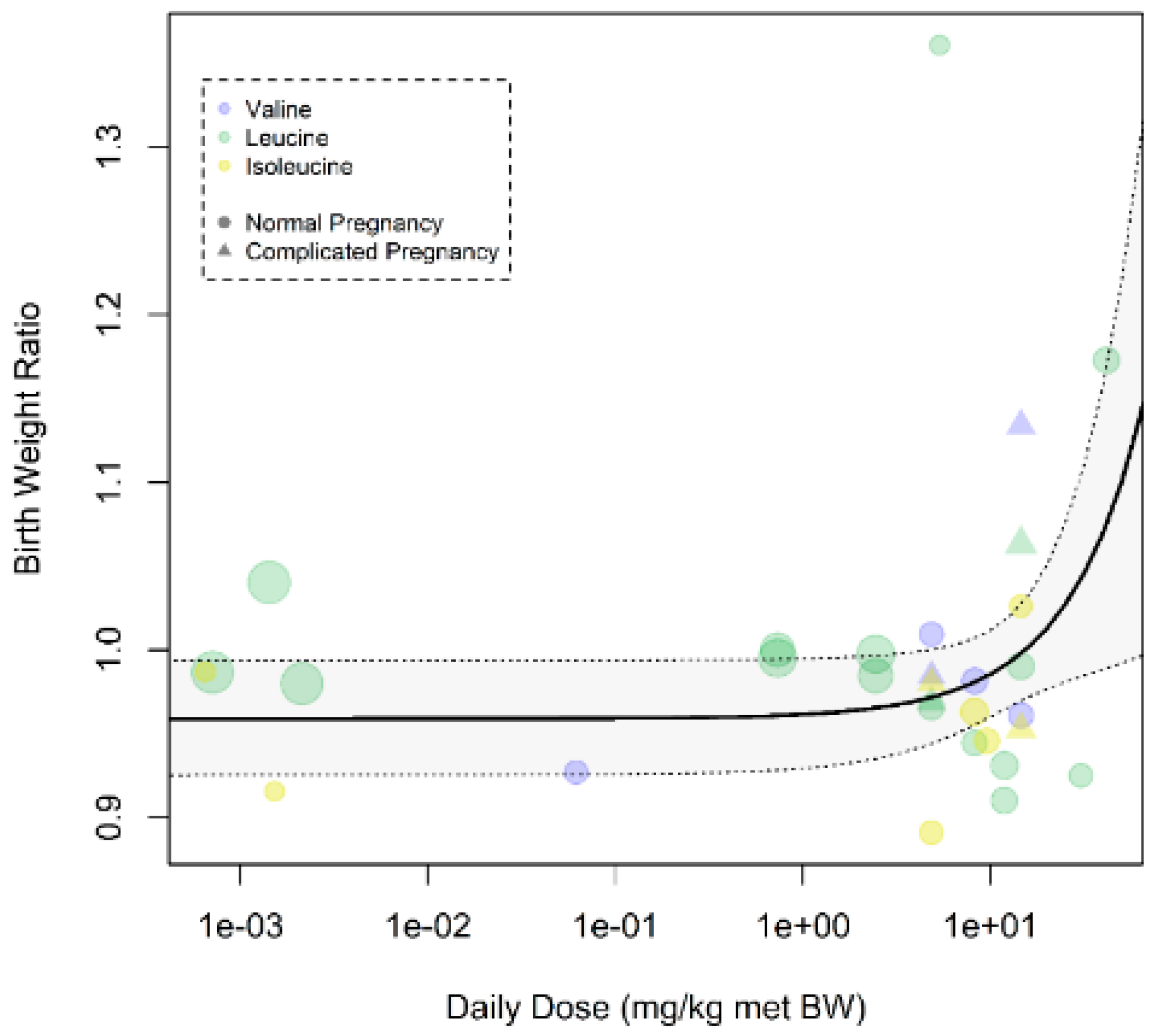{kind=link}
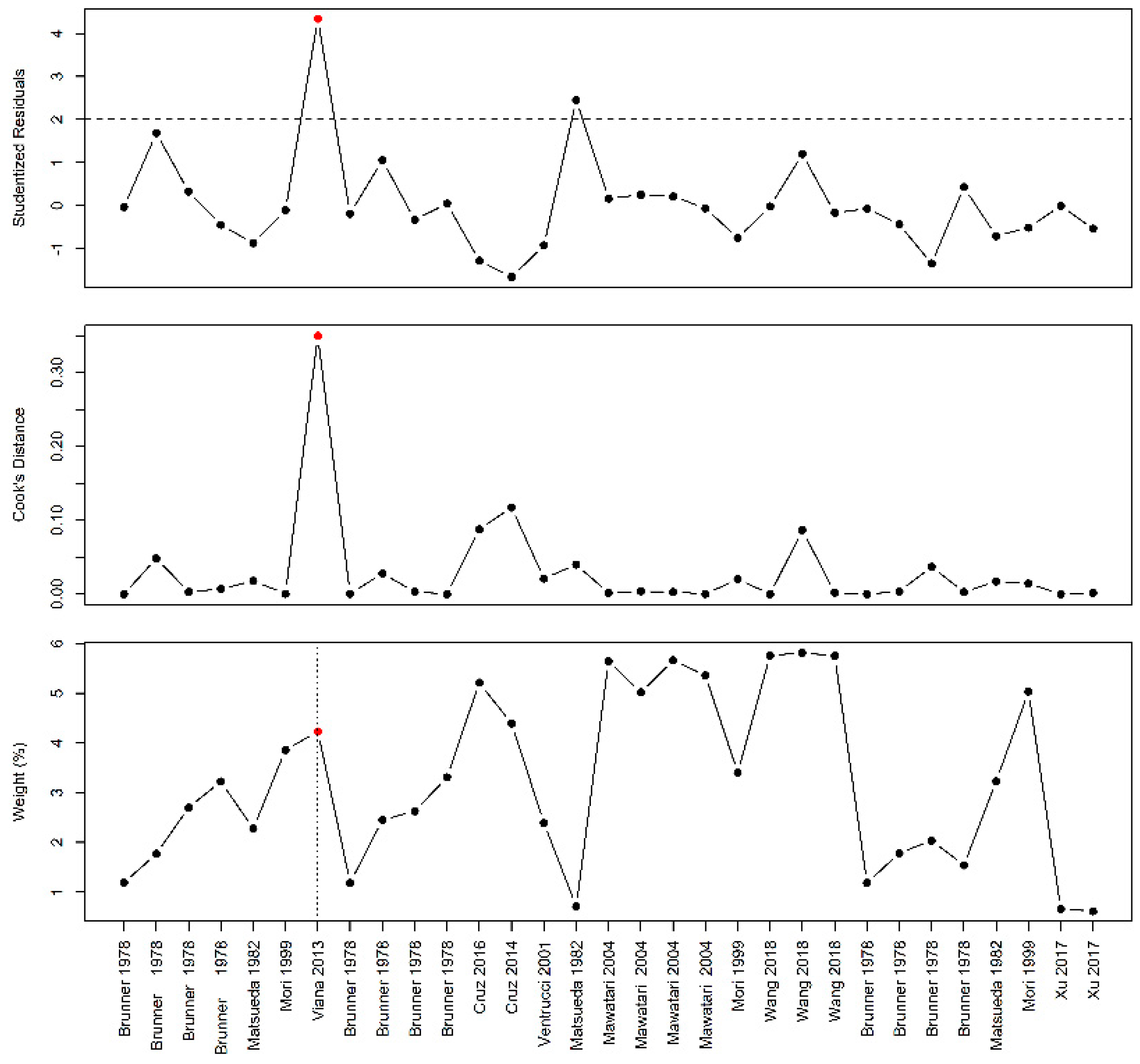{kind=link}
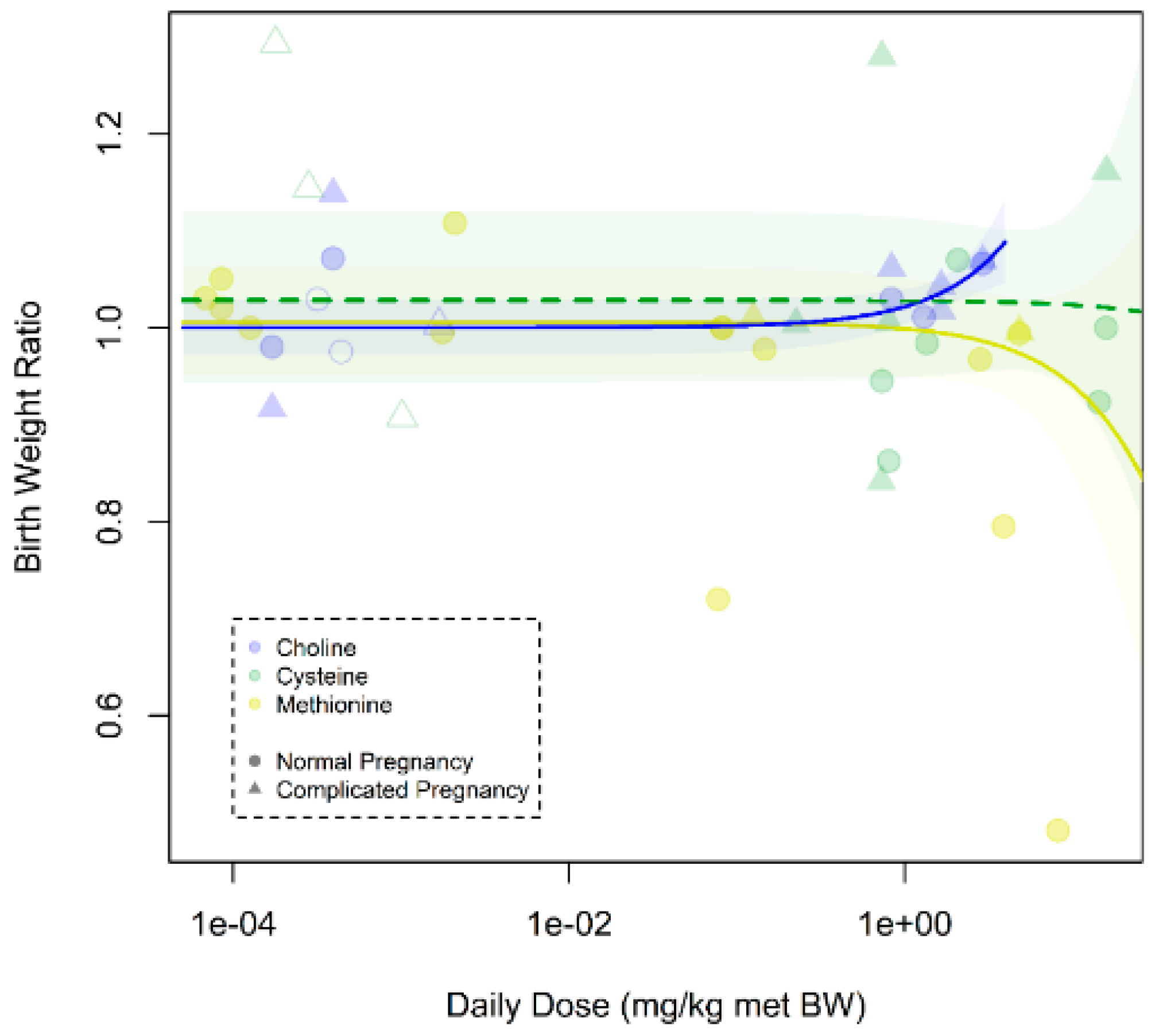{kind=link}
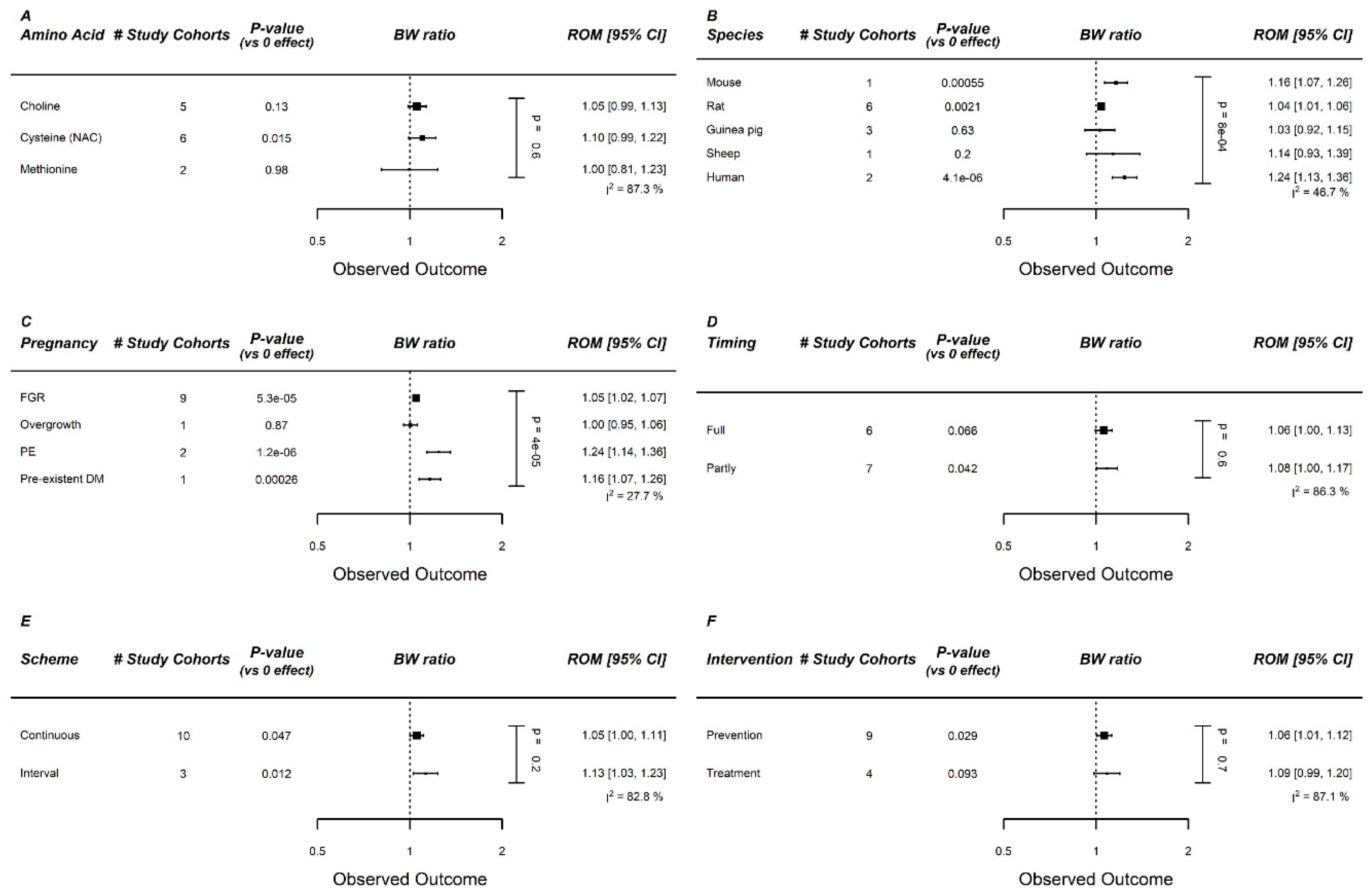{kind=link}
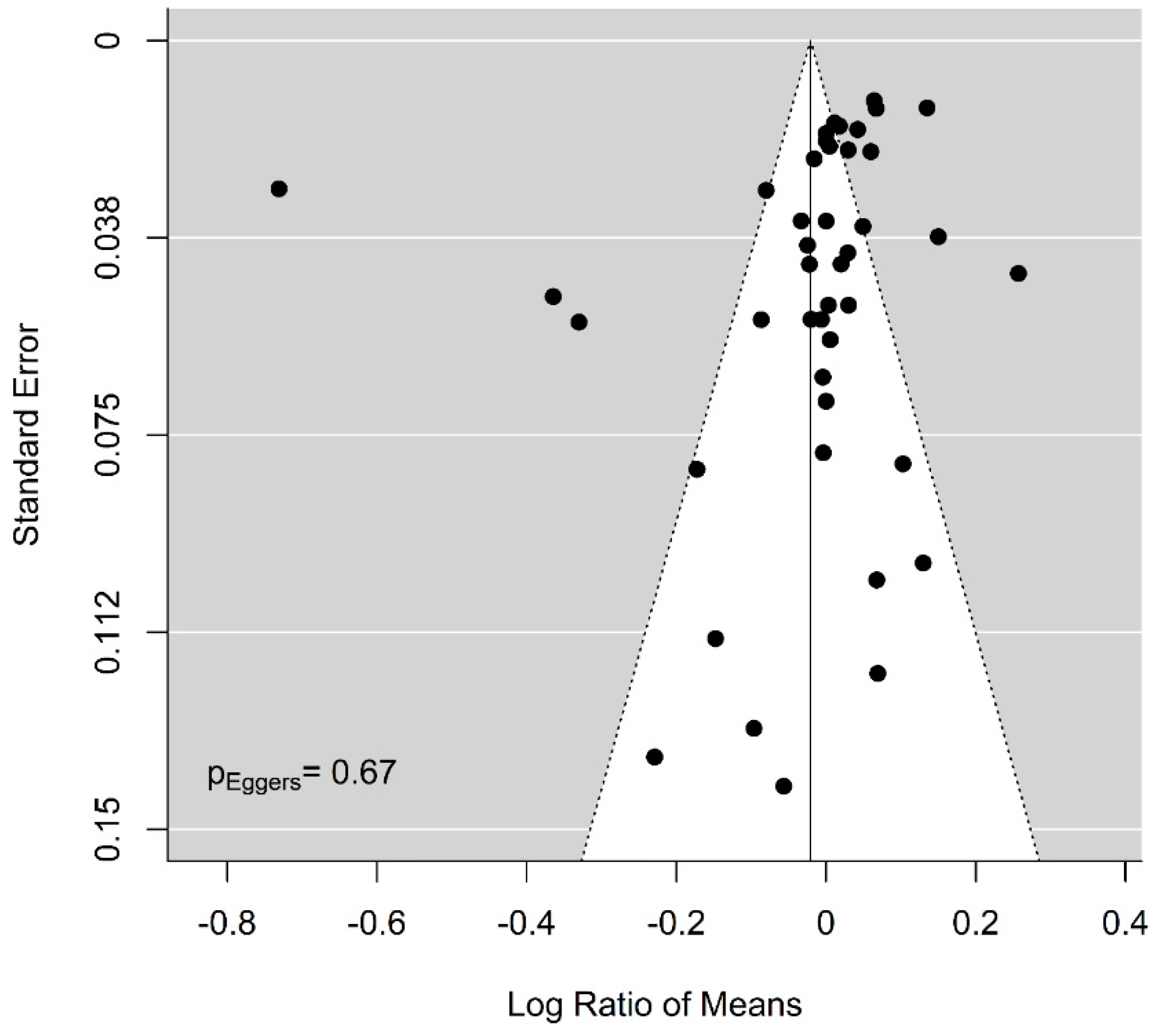{kind=link}
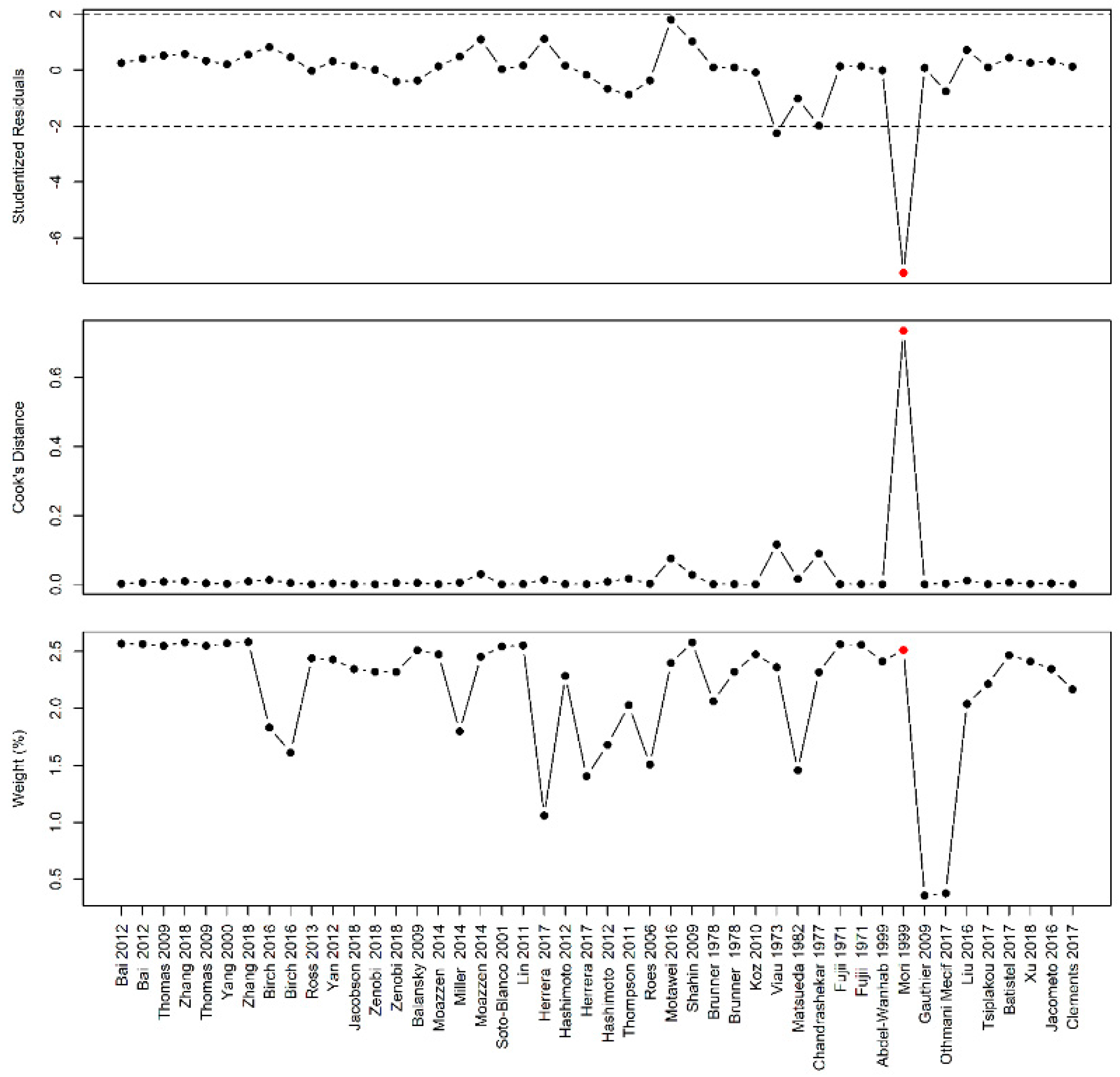{kind=link}
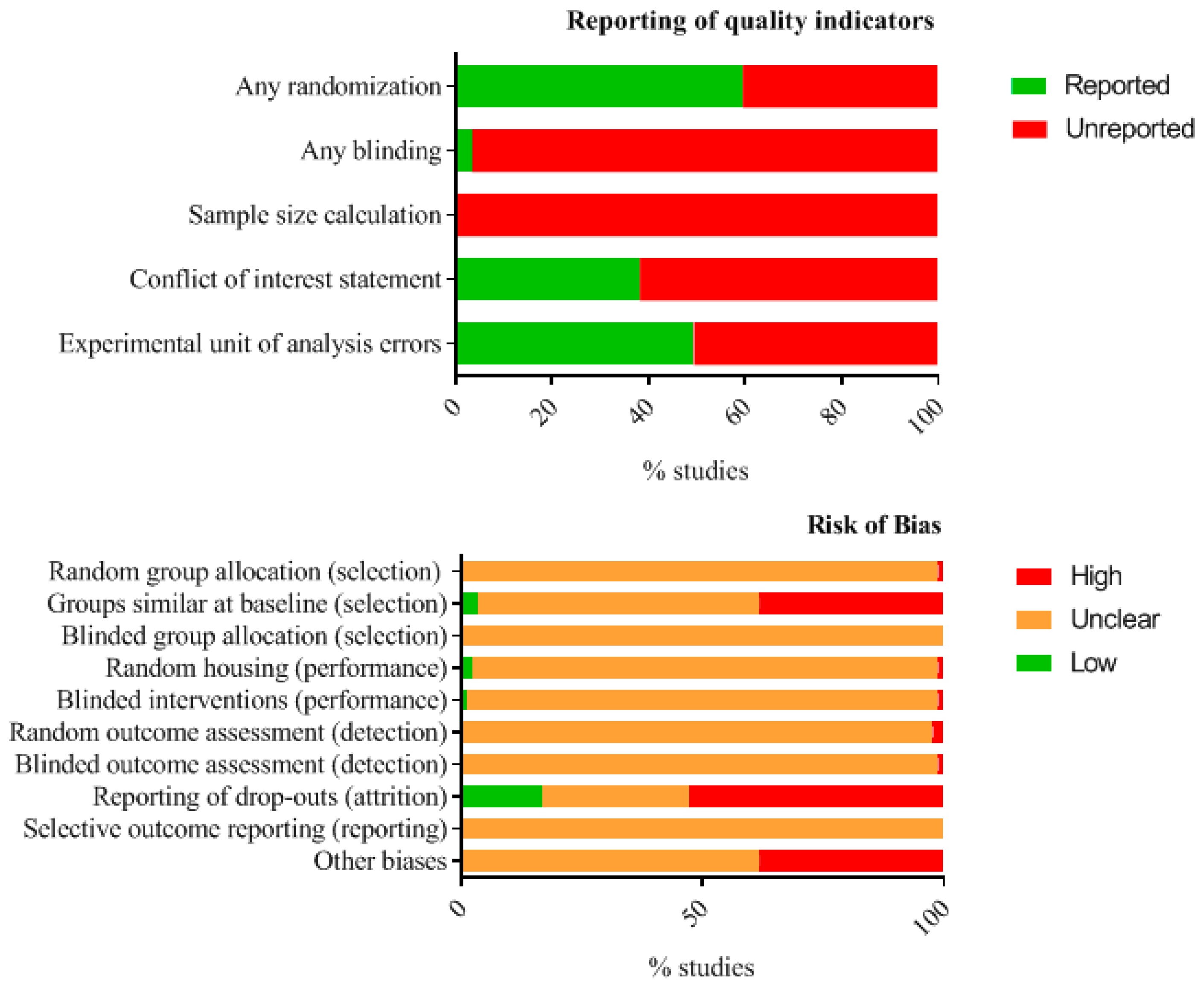{kind=link}
{kind=link}
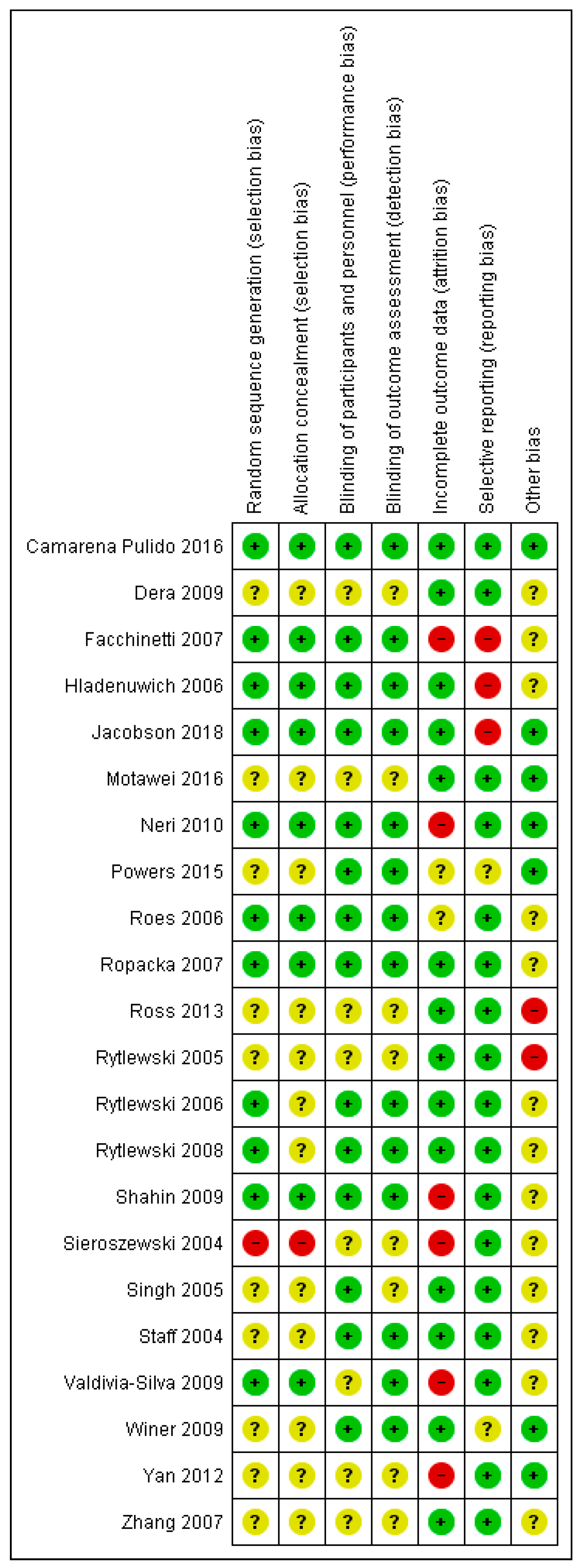{kind=link}




