Hyperhomocysteinemia and Low Folate and Vitamin B12 Are Associated with Vascular Dysfunction and Impaired Nitric Oxide Sensitivity in Morbidly Obese Patients



Abstract
:1. Introduction
2. Methods
2.1. Human Participants
2.2. Physical Measurements and Body Composition
2.3. Cardiometabolic Measurements
2.4. Plasma Hcy, Folate, and Vitamin B12
2.5. Serum NO Measurements
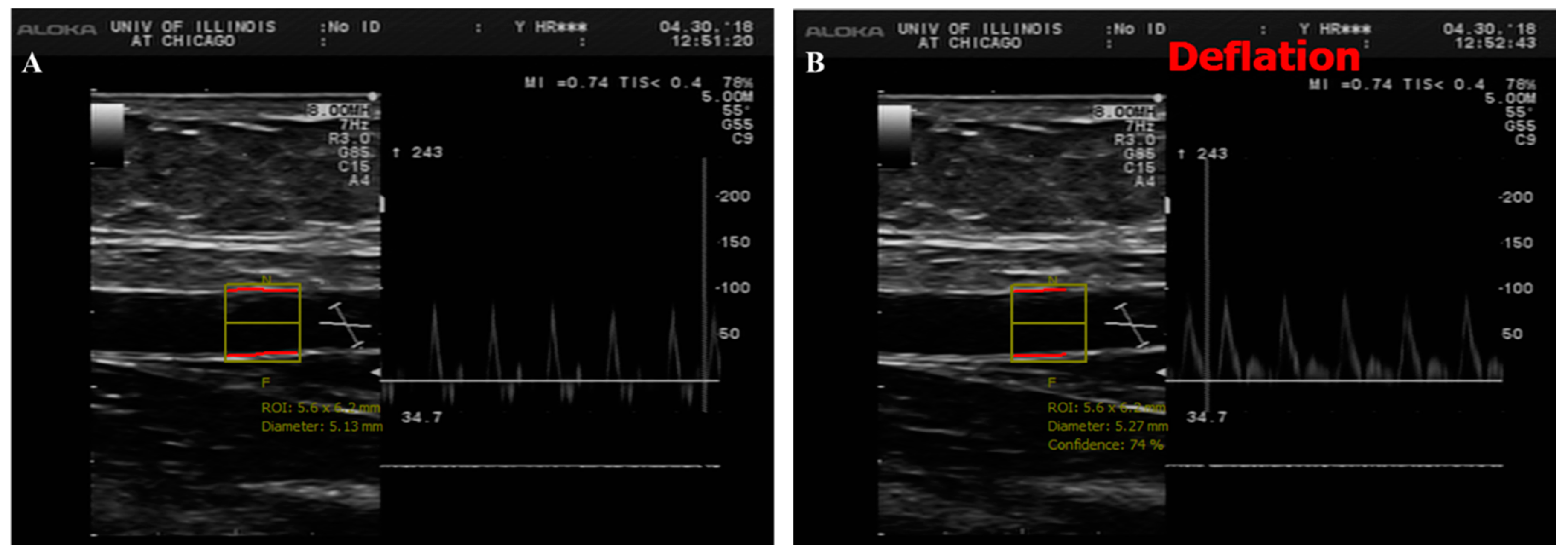
2.6. Brachial Artery Flow-Mediated Dilation (FMD)
2.7. Microvascular Preparations
2.8. Flow-Induced Dilation (FID) Measurements
2.9. Measurements of Arteriolar NO and Reactive Oxygen Species (ROS)
2.10. Statistical Analyses
3. Results
3.1. Physical and Cardiometabolic Parameters
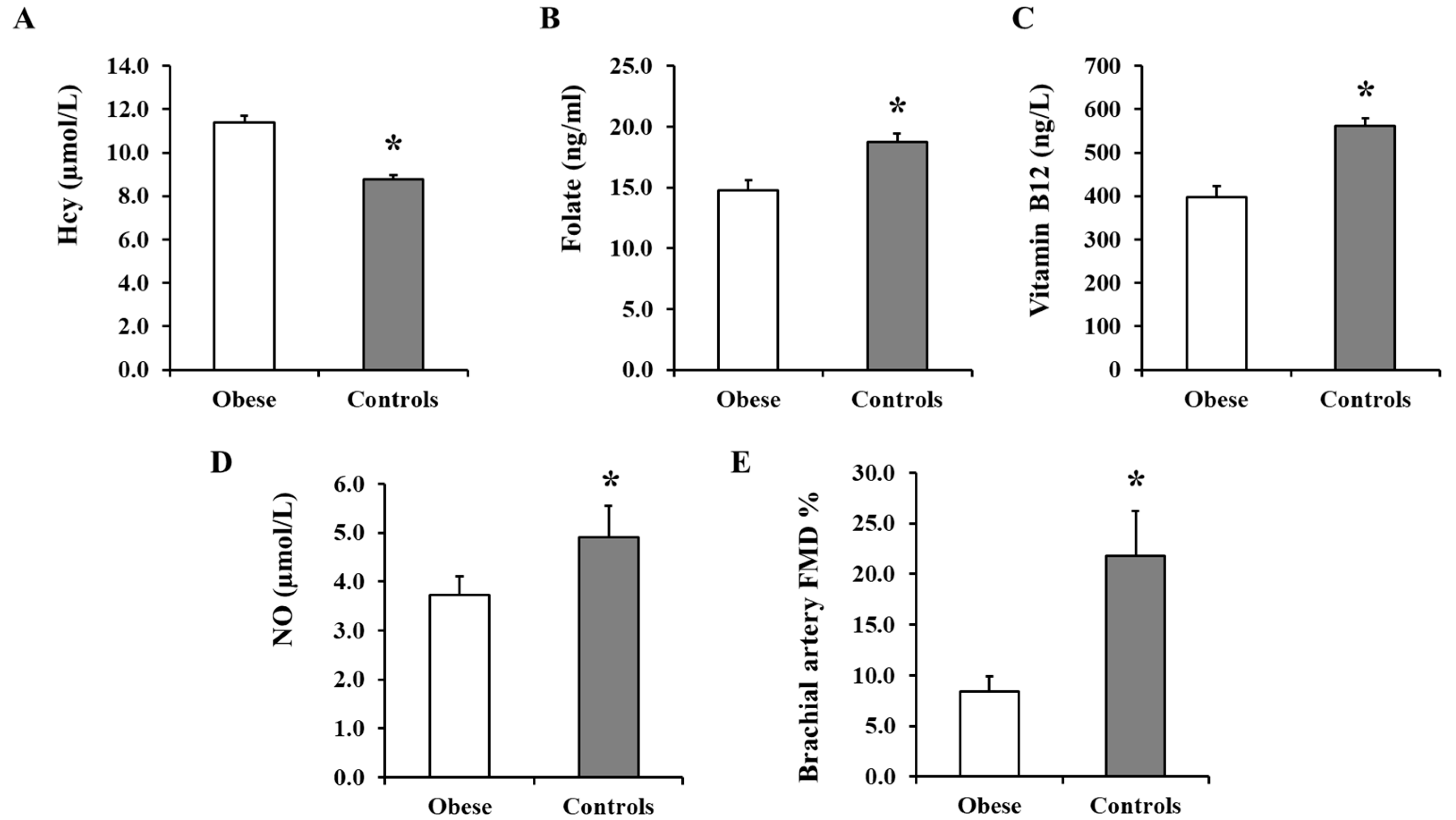
3.2. Plasma Hcy, Folate, and Vitamin B12 and Serum NO Measurements
3.3. Brachial Artery FMD
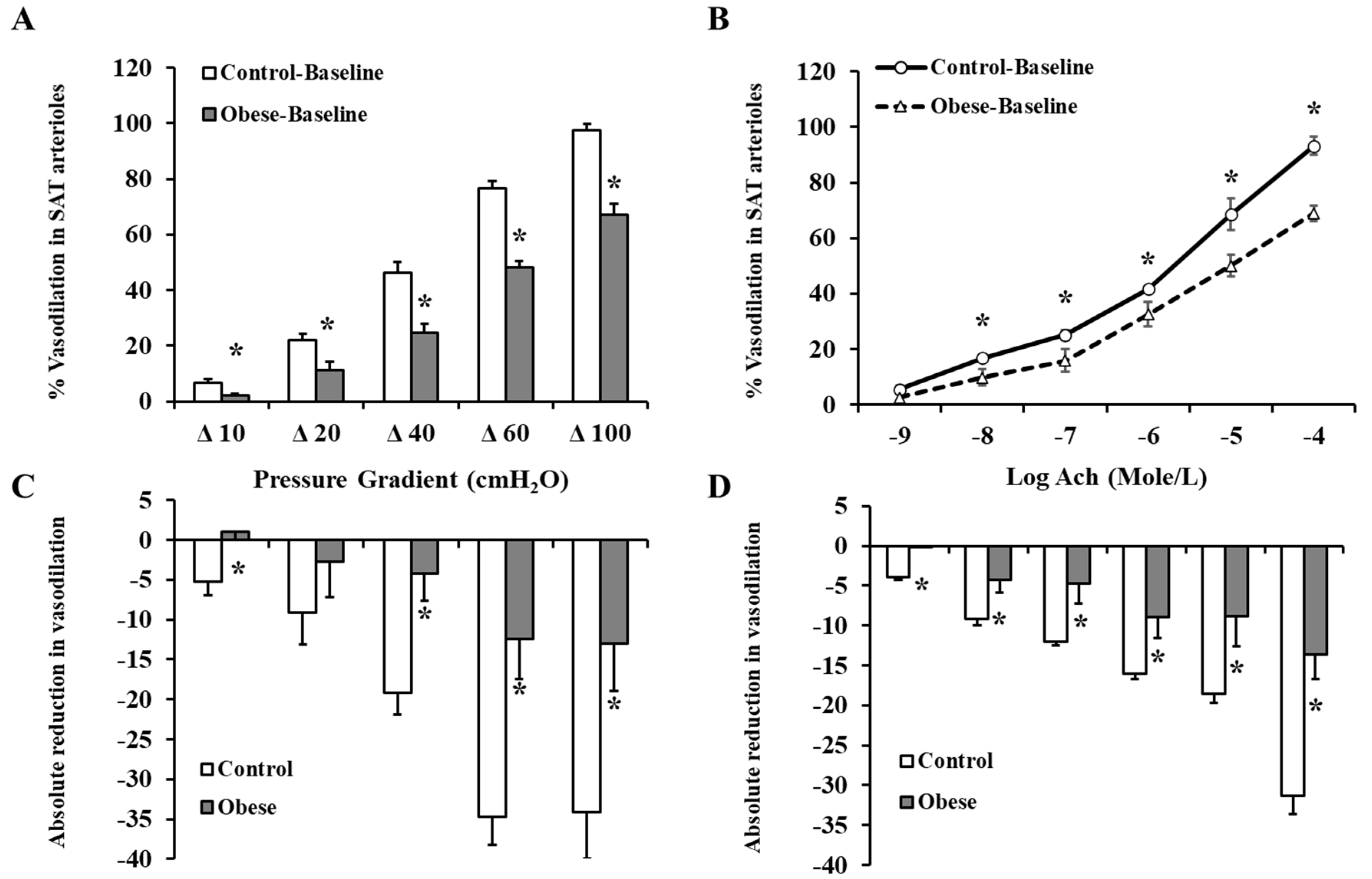
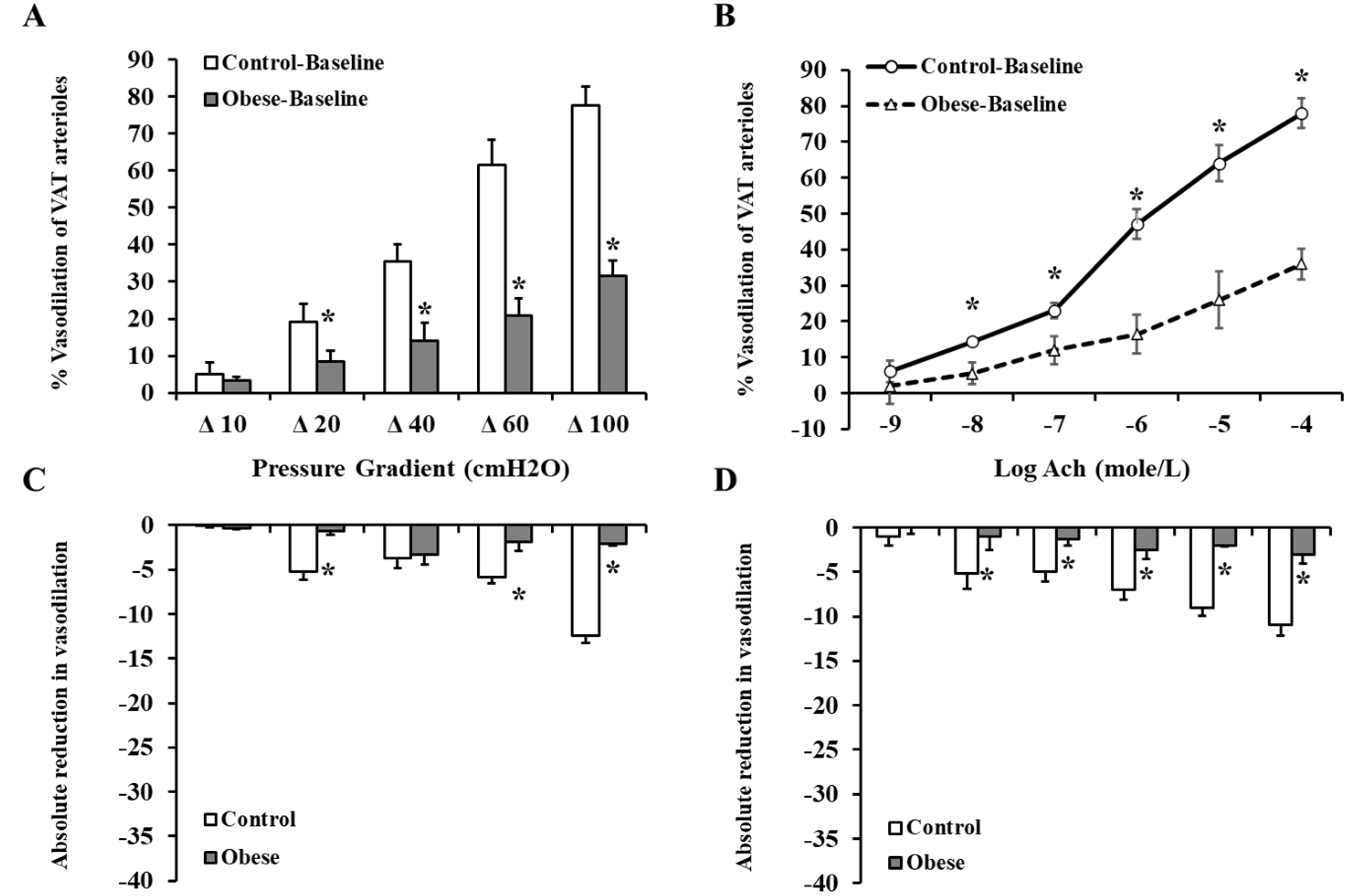
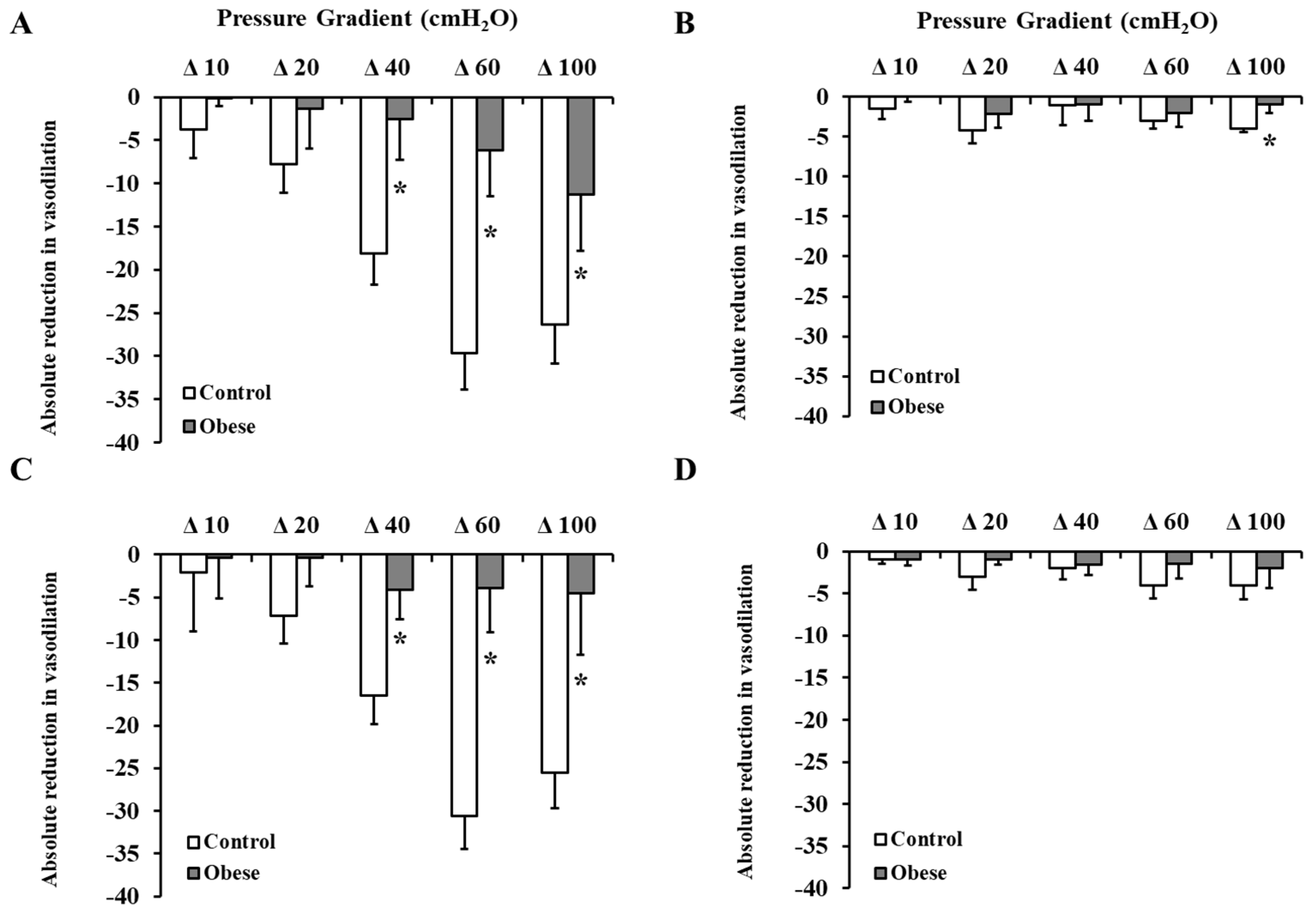
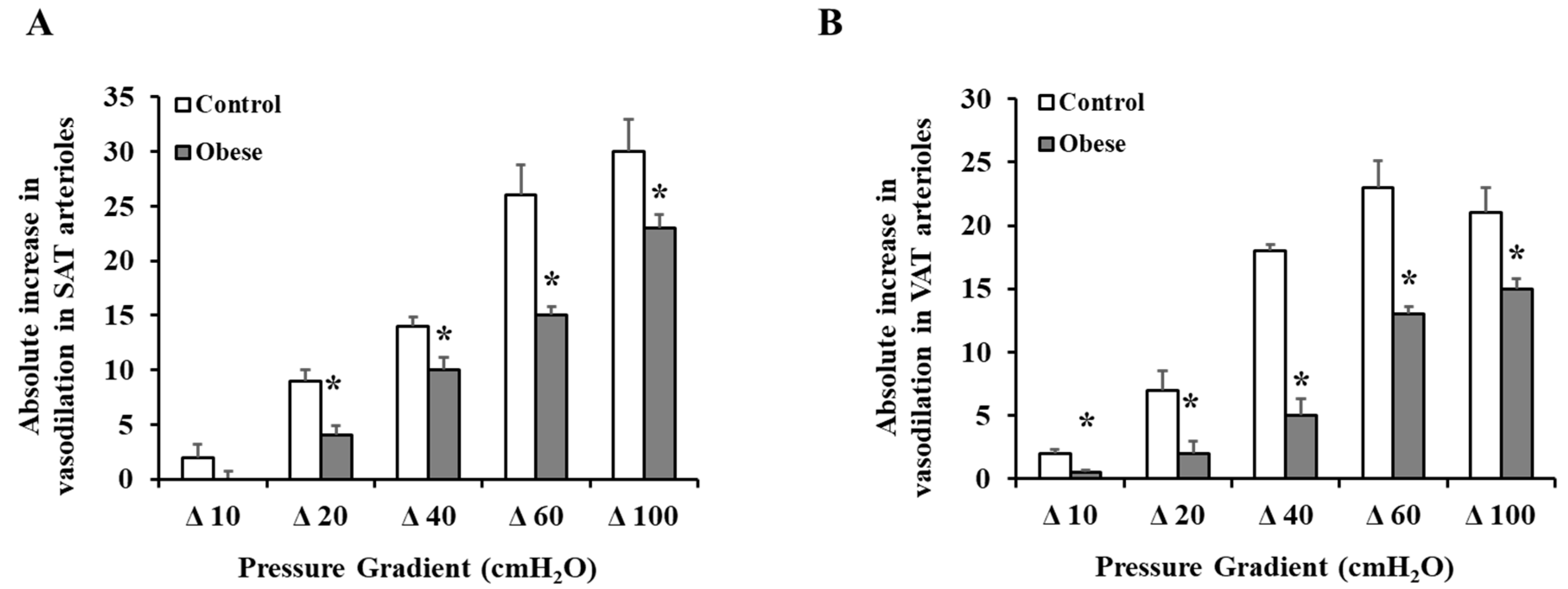
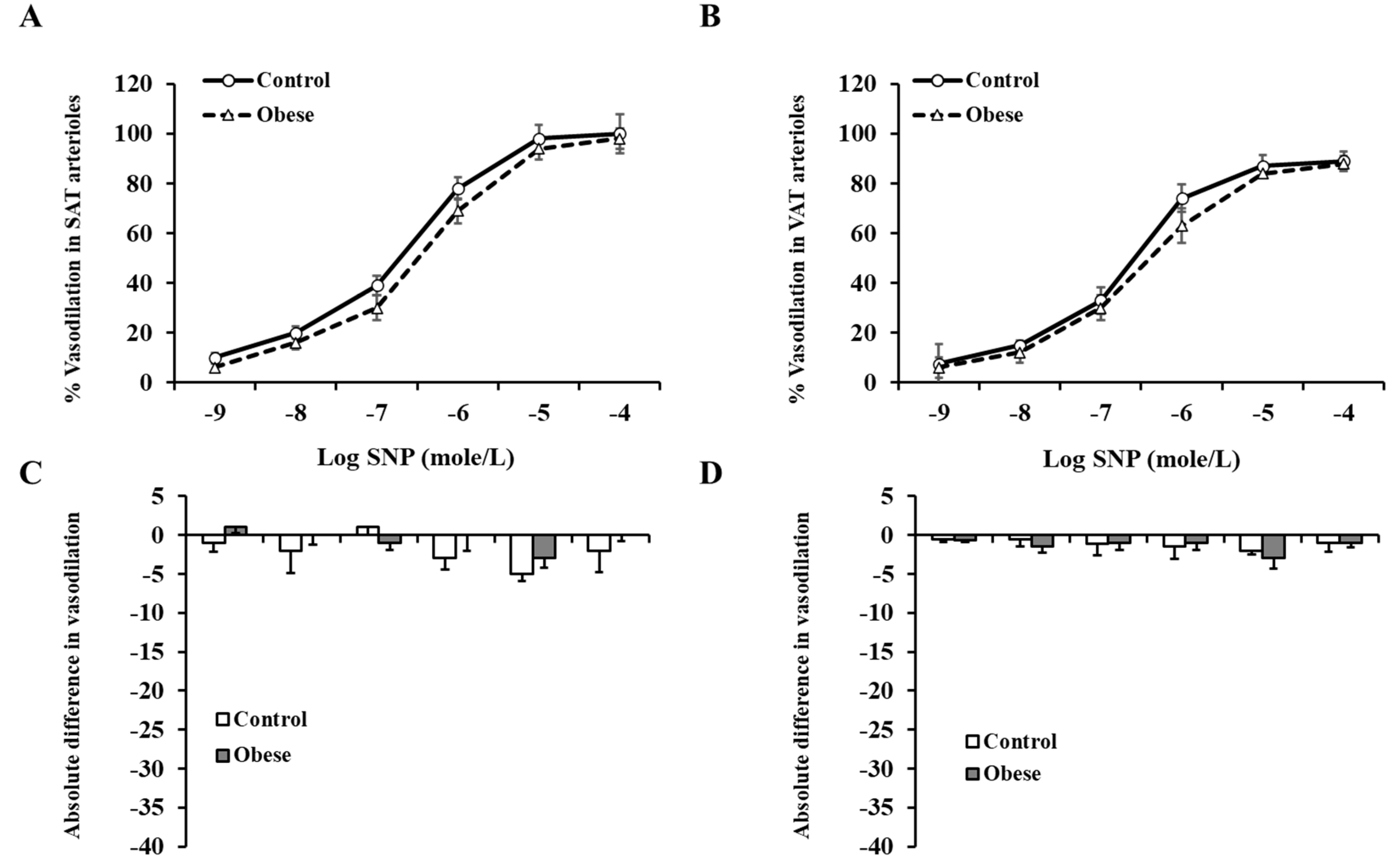
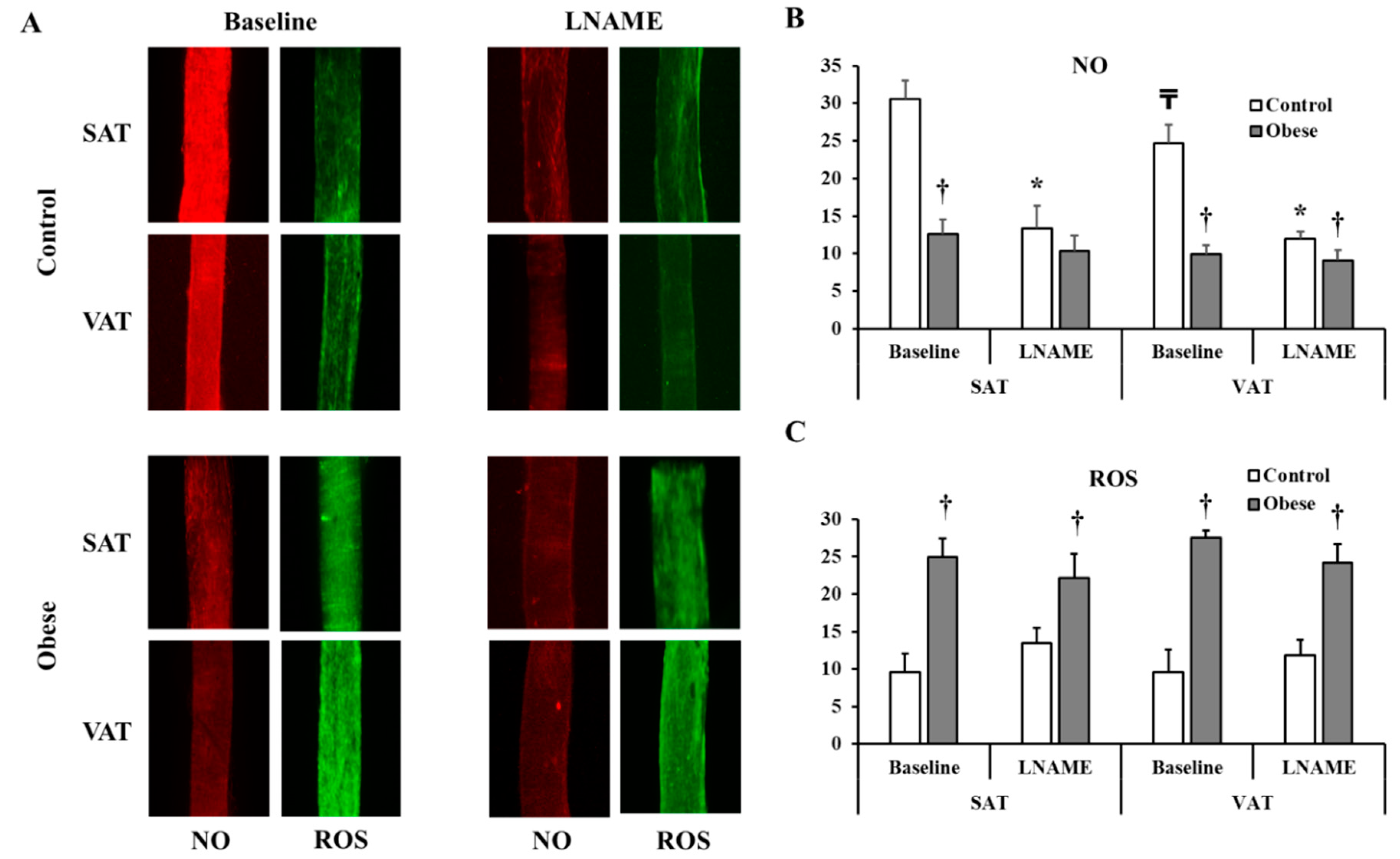
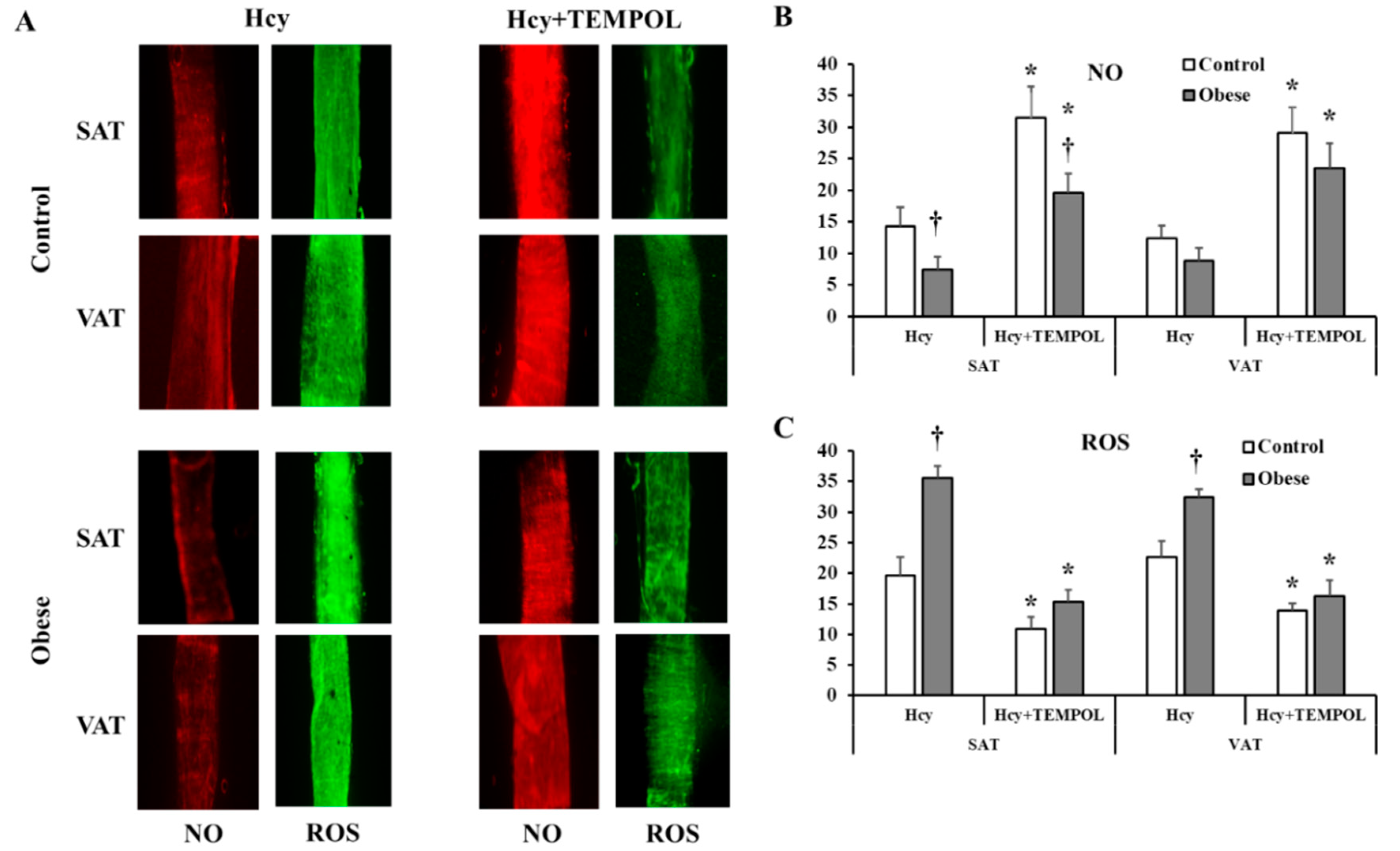
3.4. Arteriolar Vasoreactivity and NO and ROS Production
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mitchell, N.; Catenacci, V.A.; Wyatt, H.R.; Hill, J.O. Obesity: Overview of an epidemic. Psychiatr. Clin. N. Am. 2011, 34, 717–732. [Google Scholar] [CrossRef]
- Poli, V.F.S.; Sanches, R.B.; Moraes, A.D.S.; Fidalgo, J.P.N.; Nascimento, M.A.; Bresciani, P.; Andrade-Silva, S.G.; Cipullo, M.A.T.; Clemente, J.C.; Caranti, D.A. The excessive caloric intake and micronutrient deficiencies related to obesity after a long-term interdisciplinary therapy. Nutrition 2017, 38, 113–119. [Google Scholar] [CrossRef]
- Selhub, J. Folate, vitamin B12 and vitamin B6 and one carbon metabolism. J. Nutr. Health Aging 2002, 6, 39–42. [Google Scholar]
- Maron, B.A.; Loscalzo, J. The treatment of hyperhomocysteinemia. Annu. Rev. Med. 2009, 60, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, H.; Roh, H.; Kwon, Y. Causes of hyperhomocysteinemia and its pathological significance. Arch. Pharm. Res. 2018, 41, 372–383. [Google Scholar] [CrossRef]
- Wierzbicki, A.S. Homocysteine and cardiovascular disease: A review of the evidence. Diab. Vasc. Dis. Res. 2007, 4, 143–150. [Google Scholar] [CrossRef]
- Humphrey, L.L.; Fu, R.; Rogers, K.; Freeman, M.; Helfand, M. Homocysteine level and coronary heart disease incidence: A systematic review and meta-analysis. Mayo Clin. Proc. 2008, 83, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Szczurek, M.R.; Blackburn, B.K.; Mey, J.T.; Chen, Z.; Robinson, A.T.; Bian, J.-T.; Unterman, T.G.; Minshall, R.D.; Brown, M.D.; et al. Hyperinsulinemia augments endothelin-1 protein expression and impairs vasodilation of human skeletal muscle arterioles. Physiol. Rep. 2016, 4, e12895. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.A.; Hatoum, O.A.; Gutterman, D.D. The mechanism of flow-induced dilation in human adipose arterioles involves hydrogen peroxide during CAD. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H93–H100. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Hwang, C.-L.; Szczurek, M.R.; Bian, J.-T.; Ranieri, C.; Gutterman, D.D.; Phillips, S.A. Low-Fat Diet Designed for Weight Loss But Not Weight Maintenance Improves Nitric Oxide-Dependent Arteriolar Vasodilation in Obese Adults. Nutrients 2019, 11, 1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, H.; Wachtel, E.R.; Liu, Y.; Loberiza, F.R.; Saito, T.; Miura, M.; Gutterman, D.D. Flow-induced dilation of human coronary arterioles: Important role of Ca(2+)-activated K(+) channels. Circulation 2001, 103, 1992–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, C.-L.; Ranieri, C.; Szczurek, M.R.; Ellythy, A.M.; Elokda, A.; Mahmoud, A.M.; Phillips, S.A. The Effect of Low-Carbohydrate Diet on Macrovascular and Microvascular Endothelial Function is Not Affected by the Provision of Caloric Restriction in Women with Obesity: A Randomized Study. Nutrients 2020, 12, 1649. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Szczurek, M.R.; Hassan, C.; Masrur, M.; Gangemi, A.; Phillips, S.A. Vitamin D Improves Nitric Oxide-Dependent Vasodilation in Adipose Tissue Arterioles from Bariatric Surgery Patients. Nutrients 2019, 11, 2521. [Google Scholar] [CrossRef] [Green Version]
- Bian, J.-T.; Piano, M.; Kotlo, K.U.; Mahmoud, A.M.; Phillips, S.A. MicroRNA-21 Contributes to Reduced Microvascular Function in Binge Drinking Young Adults. Alcohol Clin. Exp. Res. 2018, 42, 278–285. [Google Scholar] [CrossRef]
- Lang, D.; Kredan, M.B.; Moat, S.J.; Hussain, S.A.; Powell, C.A.; Bellamy, M.F.; Powers, H.J.; Lewis, M.J. Homocysteine-induced inhibition of endothelium-dependent relaxation in rabbit aorta: Role for superoxide anions. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.T.; Franklin, N.; Norkeviciute, E.; Bian, J.T.; Babana, J.C.; Szczurek, M.R.; Phillips, S.A. Improved arterial flow-mediated dilation after exertion involves hydrogen peroxide in overweight and obese adults following aerobic exercise training. J. Hypertens. 2016, 34, 1309–1316. [Google Scholar] [CrossRef]
- Clark, W.B.; Hilton, M.E. Alcohol in America: Drinking Practices and Problems; State University of New York Press: Albany, NY, USA, 1991. [Google Scholar]
- Boushey, C.J.; Beresford, S.A.; Omenn, G.S.; Motulsky, A.G. A quantitative assessment of plasma homocysteine as a risk factor for vascular disease. Probable benefits of increasing folic acid intakes. JAMA 1995, 274, 1049–1057. [Google Scholar] [CrossRef]
- Van den Bosch, M.A.; Bloemenkamp, D.G.; Willem, P.T.M.; Kemmeren, J.M.; Tanis, B.C.; Algra, A.; Rosendaal, F.R.; van der Graaf, Y. Hyperhomocysteinemia and risk for peripheral arterial occlusive disease in young women. J. Vasc. Surg. 2003, 38, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.V.; Oudit, G.Y.; Evans, M. Homocysteine, vitamins, and coronary artery disease. Comprehensive review of the literature. Can. Fam. Physician 2000, 46, 2236–2245. [Google Scholar]
- Vayá, A.; Rivera, L.; Mijares, A.H.; De La Fuente, M.; Solá, E.; Romagnoli, M.; Alis, R.; Laiz, B. Homocysteine levels in morbidly obese patients: Its association with waist circumference and insulin resistance. Clin. Hemorheol. Microcirc. 2012, 52, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Margalet, V.; Valle, M.; Ruz, F.J.; Gascón, F.; Mateo, J.; Goberna, R. Elevated plasma total homocysteine levels in hyperinsulinemic obese subjects. J. Nutr. Biochem. 2002, 13, 75–79. [Google Scholar] [CrossRef]
- Meigs, J.B.; Jacques, P.F.; Selhub, J.; Singer, D.E.; Nathan, D.M.; Rifai, N.; D’Agostino, R.B.; Wilson, P.W. Fasting plasma homocysteine levels in the insulin resistance syndrome: The Framingham offspring study. Diabetes Care 2001, 24, 1403–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehler, K.M.; Baumgartner, R.N.; Garry, P.J.; Allen, R.H.; Stabler, S.P.; Rimm, E.B. Association of folate intake and serum homocysteine in elderly persons according to vitamin supplementation and alcohol use. Am. J. Clin. Nutr. 2001, 73, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Martos, R.; Valle, M.; Morales, R.; Cañete, R.; Gavilan, M.I.; Sánchez-Margalet, V. Hyperhomocysteinemia correlates with insulin resistance and low-grade systemic inflammation in obese prepubertal children. Metabolism 2006, 55, 72–77. [Google Scholar] [CrossRef]
- Cheng, Z.; Yang, X.; Wang, H. Hyperhomocysteinemia and Endothelial Dysfunction. Curr. Hypertens. Rev. 2009, 5, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Austin, R.C.; Lentz, S.R.; Werstuck, G.H. Role of hyperhomocysteinemia in endothelial dysfunction and atherothrombotic disease. Cell Death Differ. 2004, 11, S56–S64. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.K.; Kan, M.Y. Homocysteine-Induced Endothelial Dysfunction. Ann. Nutr. Metab. 2015, 67, 1–12. [Google Scholar] [CrossRef]
- Faraci, F.M.; Lentz, S.R. Hyperhomocysteinemia, oxidative stress, and cerebral vascular dysfunction. Stroke 2004, 35, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Chambers, J.C.; McGregor, A.; Jean-Marie, J.; Obeid, A.O.; Kooner, J.S. Demonstration of rapid onset vascular endothelial dysfunction after hyperhomocysteinemia: An effect reversible with vitamin C therapy. Circulation 1999, 99, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Lentz, S.R.; Rodionov, R.N.; Dayal, S. Hyperhomocysteinemia, endothelial dysfunction, and cardiovascular risk: The potential role of ADMA. Atheroscler. Suppl. 2003, 4, 61–65. [Google Scholar] [CrossRef]
- McDowell, I.F.; Lang, D. Homocysteine and endothelial dysfunction: A link with cardiovascular disease. J. Nutr. 2000, 130, 369S–372S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Guldener, C. Why is homocysteine elevated in renal failure and what can be expected from homocysteine-lowering? Nephrol. Dial. Transplant. 2006, 21, 1161–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinzari, F.; Iantorno, M.; Campia, U.; Mores, N.; Rovella, V.; Tesauro, M.; Di Daniele, N.; Cardillo, C. Vasodilator responses and endothelin-dependent vasoconstriction in metabolically healthy obesity and the metabolic syndrome. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E787–E792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jongh, R.T.; Serné, E.H.; Ijzerman, R.G.; De Vries, G.; Stehouwer, C.D. Impaired microvascular function in obesity: Implications for obesity-associated microangiopathy, hypertension, and insulin resistance. Circulation 2004, 109, 2529–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldeweg, S.; Pink, A.; Yudkin, J.; Coppack, S. The relationship between obesity, vascular reactivity and endothelial dysfunction in subjects with non-insulin dependent diabetes mellitus. Int. J. Obes. Relat. Metab. Disord. 2000, 24, S134–S135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patik, J.C.; Christmas, K.M.; Hurr, C.; Brothers, R.M. Impaired endothelium independent vasodilation in the cutaneous microvasculature of young obese adults. Microvasc. Res. 2016, 104, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.Y.; Dudman, N.P.; Perry, M.A.; Wang, X. Homocysteine attenuates hemodynamic responses to nitric oxide in vivo. Atherosclerosis 2002, 161, 169–176. [Google Scholar] [CrossRef]
- Schlaich, M.P.; John, S.; Jacobi, J.; Lackner, K.J.; Schmieder, R.E. Mildly elevated homocysteine concentrations impair endothelium dependent vasodilation in hypercholesterolemic patients. Atherosclerosis 2000, 153, 383–389. [Google Scholar] [CrossRef]
- Refsum, H.; Ueland, P.M.; Nygård, O.; Vollset, S.E. Homocysteine and cardiovascular disease. Annu. Rev. Med. 1998, 49, 31–62. [Google Scholar] [CrossRef]
- Wald, D.S.; Law, M.; Morris, J.K. Homocysteine and cardiovascular disease: Evidence on causality from a meta-analysis. BMJ 2002, 325, 1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.J.; Gona, P.; Larson, M.G.; Tofler, G.H.; Levy, D.; Newton-Cheh, C.; Jacques, P.F.; Rifai, N.; Selhub, J.; Robins, S.J.; et al. Multiple biomarkers for the prediction of first major cardiovascular events and death. N. Engl. J. Med. 2006, 355, 2631–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Peng, D.; Liu, C.; Huang, C.; Luo, J. Serum high concentrations of homocysteine and low levels of folic acid and vitamin B12 are significantly correlated with the categories of coronary artery diseases. BMC Cardiovasc. Disord. 2017, 17, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scazzone, C.; Bono, A.; Tornese, F.; Arsena, R.; Schillaci, R.; Butera, D.; Cottone, S. Correlation between low folate levels and hyperhomocysteinemia, but not with vitamin B12 in hypertensive patients. Ann. Clin. Lab. Sci. 2014, 44, 286–290. [Google Scholar]
- Karatela, R.A.; Sainani, G.S. Plasma homocysteine in obese, overweight and normal weight hypertensives and normotensives. Indian Heart J. 2009, 61, 156–159. [Google Scholar]
- Han, L.; Liu, Y.; Wang, C.; Tang, L.; Feng, X.; Astell-Burt, T.; Wen, Q.; Duan, D.; Lu, N.; Xu, G.; et al. Determinants of hyperhomocysteinemia in healthy and hypertensive subjects: A population-based study and systematic review. Clin. Nutr. 2017, 36, 1215–1230. [Google Scholar] [CrossRef]
- Jacobsen, D.W. Hyperhomocysteinemia and oxidative stress: Time for a reality check? Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1182–1184. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Chen, Y.; Li, J.; Yang, X.; Zhang, H.; Qin, X.; Hu, Y.; Mo, Z. Serum Homocysteine Concentration Is Significantly Associated with Inflammatory/Immune Factors. PLoS ONE 2015, 10, e0138099. [Google Scholar] [CrossRef] [Green Version]
- Yakub, M.; Iqbal, M.P.; Iqbal, R. Dietary patterns are associated with hyperhomocysteinemia in an urban Pakistani population. J. Nutr. 2010, 140, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Deminice, R.; Ribeiro, D.F.; Frajacomo, F.T. The Effects of Acute Exercise and Exercise Training on Plasma Homocysteine: A Meta-Analysis. PLoS ONE 2016, 11, e0151653. [Google Scholar] [CrossRef]
- Gibson, A.; Woodside, J.V.; Young, I.S.; Sharpe, P.; Mercer, C.; Patterson, C.; McKinley, M.; Kluijtmans, L.; Whitehead, A.; Evans, A. Alcohol increases homocysteine and reduces B vitamin concentration in healthy male volunteers—A randomized, crossover intervention study. QJM 2008, 101, 881–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffer, A.; Verdoia, M.; Barbieri, L.; Cassetti, E.; Suryapranata, H.; De Luca, G. Impact of Diabetes on Homocysteine Levels and Its Relationship with Coronary Artery Disease: A Single-Centre Cohort Study. Ann. Nutr. Metab. 2016, 68, 180–188. [Google Scholar] [CrossRef] [PubMed]
- De Bree, A.; van Mierlo, L.A.; Draijer, R. Folic acid improves vascular reactivity in humans: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2007, 86, 610–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olthof, M.R.; Bots, M.L.; Katan, M.B.; Verhoef, P. Acute effect of folic acid, betaine, and serine supplements on flow-mediated dilation after methionine loading: A randomized trial. PLoS Clin. Trials. 2006, 1, e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, A.; Kim, E.-K.; Kim, H.; Choi, Y.J.; Huh, K.B.; Chang, N. Effects of folic acid supplementation on serum homocysteine levels, lipid profiles, and vascular parameters in post-menopausal Korean women with type 2 diabetes mellitus. Nutr. Res. Pract. 2017, 11, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, X.; Zhou, Y.; Jiang, D.; Li, X.; Guo, Y.; Jiang, X. Efficacy of folic acid supplementation on endothelial function and plasma homocysteine concentration in coronary artery disease: A meta-analysis of randomized controlled trials. Exp. Ther. Med. 2014, 7, 1100–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoch, A.Z.; Pajewski, N.M.; Hoffmann, R.G.; Schimke, J.E.; Gutterman, D.D. Possible relationship of folic Acid supplementation and improved flow-mediated dilation in premenopausal, eumenorrheic athletic women. J. Sports Sci. Med. 2009, 8, 123–129. [Google Scholar]
- Piano, M.R. Alcohol’s Effects on the Cardiovascular System. Alcohol Res. 2017, 38, 219–241. [Google Scholar]
- Halsted, C.H.; Villanueva, J.A.; Devlin, A.M.; Chandler, C.J. Metabolic interactions of alcohol and folate. J. Nutr. 2002, 132, 2367S–2372S. [Google Scholar] [CrossRef] [Green Version]
- Laufer, E.M.; Hartman, T.J.; Baer, D.J.; Gunter, E.W.; Dorgan, J.F.; Campbell, W.S.; Clevidence, A.B.; Brown, E.D.; Albanes, D.; Judd, J.T.; et al. Effects of moderate alcohol consumption on folate and vitamin B(12) status in postmenopausal women. Eur. J. Clin. Nutr. 2004, 58, 1518–1524. [Google Scholar] [CrossRef] [Green Version]
- Bleich, S.; Bleich, K.; Kropp, S.; Bittermann, H.-J.; Degner, D.; Sperling, W.; Rüther, E.; Kornhuber, J. Moderate alcohol consumption in social drinkers raises plasma homocysteine levels: A contradiction to the ‘French Paradox’? Alcohol Alcohol. 2001, 36, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Rimm, E.B.; Willett, W.C.; Hu, F.B.; Sampson, L.; Colditz, G.A.; Manson, J.E.; Hennekens, C.; Stampfer, M.J. Folate and vitamin B6 from diet and supplements in relation to risk of coronary heart disease among women. JAMA 1998, 279, 359–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halsted, C.H. Lifestyle effects on homocysteine and an alcohol paradox. Am. J. Clin. Nutr. 2001, 73, 501–502. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Non-Obese Controls | Obese Bariatric Patients | p-Value |
|---|---|---|---|
| n | 40 (18 ♂) | 40 (12 ♂) | |
| Age, y | 35.4 ± 1.3 | 36.2 ± 1 | 0.339 |
| Weight, kg | 74.4 ± 1.6 | 142.4 ± 3.7 * | <0.001 |
| BMI, kg/ m2 | 24.9 ± 0.5 | 50.6 ± 1.1 * | <0.001 |
| WC, cm | 131.5 ± 4 | 91.5 ± 2 * | <0.001 |
| Body fat, % | 32.2 ± 2.5 | 52.3 ± 1.0 * | <0.001 |
| Body lean, % | 65.3 ± 2.4 | 46.5 ± 0.9 * | <0.001 |
| VAT mass, kg | 0.7 ± 0.1 | 2.0 ± 0.2 * | 0.0002 |
| HR, bpm | 74 ± 2 | 81 ± 1 * | 0.004 |
| SBP, mmHg | 118 ± 2 | 132 ± 2 * | <0.001 |
| DBP, mmHg | 75 ± 1 | 80 ± 1 * | 0.013 |
| FPG, mg/dL | 92 ± 2 | 103 ± 5 | 0.112 |
| FPI, µU/mL | 8.4 ± 1.4 | 14.9 ± 2.8 * | <0.001 |
| HOMA-IR | 1.9 ± 0.1 | 4.1 ± 0.4 * | 0.002 |
| HbA1c, % | 5.3 ± 0.1 | 5.8 ± 0.2 | 0.148 |
| Total chol, mg/dL | 155 ± 9 | 165 ± 4 | 0.171 |
| HDL, mg/dL | 56 ± 6 | 43 ± 1 * | <0.001 |
| LDL, mg/dL | 87 ± 7 | 99 ± 4 | 0.116 |
| TG, mg/dL | 92 ± 11 | 115 ± 8 | 0.136 |
| IL6, pg/mL | 5.2 ± 0.7 | 21.6 ± 3.6 * | 0.001 |
| IL8, pg/mL | 3.0 ± 0.1 | 4.9 ± 0.4 * | <0.001 |
| CRP, mg/L | 0.7 ± 0.1 | 3.8 ± 0.2 * | <0.0001 |
| SAT Arteriole FID at Δ 60 | VAT Arteriole FID at Δ 60 | |||
|---|---|---|---|---|
| Pearson Correlation | p Value | Pearson Correlation | p Value | |
| Weight | −0.856 | <0.0001 | −0.873 | <0.0001 |
| WC | −0.761 | <0.0001 | −0.761 | <0.0001 |
| BMI | −0.916 | <0.0001 | −0.937 | <0.0001 |
| Fat% | −0.839 | <0.0001 | −0.843 | <0.0001 |
| Lean% | 0.833 | <0.0001 | 0.837 | <0.0001 |
| VAT Mass | −0.625 | <0.0001 | −0.635 | <0.0001 |
| HR | −0.185 | 0.045 | −0.187 | 0.043 |
| SBP | −0.357 | <0.0001 | −0.355 | <0.0001 |
| DBP | −0.181 | 0.048 | −0.172 | 0.057 |
| FPI | −0.516 | <0.0001 | −0.500 | <0.0001 |
| HOMA-IR | −0.293 | 0.006 | −0.289 | 0.006 |
| HDL | 0.350 | 0.004 | 0.373 | 0.002 |
| Hcy | −0.427 | 0.003 | −0.420 | 0.004 |
| Folate | 0.344 | 0.002 | 0.341 | 0.002 |
| VitB12 | 0.432 | <0.0001 | 0.421 | <0.0001 |
| Alcohol | −0.212 | 0.035 | −0.206 | 0.039 |
| FMD% | 0.434 | 0.004 | 0. 349 | 0.017 |
| % Reduction in FID at Δ 60 cmH2O Following L-NAME Incubation (NO Sensitivity) | ||||
|---|---|---|---|---|
| SAT Arteriole | VAT Arteriole | |||
| Pearson Correlation | p Value | Pearson Correlation | p Value | |
| Weight | −0.844 | <0.0001 | −0.428 | <0.0001 |
| WC | −0.649 | <0.0001 | −0.079 | 0.317 |
| BMI | −0.914 | <0.0001 | −0.264 | 0.007 |
| Fat% | −0.715 | <0.0001 | −0.173 | 0.126 |
| Lean% | 0.711 | <0.0001 | 0.172 | 0.126 |
| VAT Mass | −0.529 | 0.002 | −0.051 | 0.399 |
| SBP | −0.332 | 0.001 | −0.220 | 0.021 |
| HbA1c | −0.241 | 0.043 | −0.326 | 0.009 |
| FPI | −0.506 | <0.0001 | −0.363 | 0.001 |
| HOMA-IR | −0.248 | 0.017 | −0.266 | 0.011 |
| HDL | 0.385 | 0.002 | 0.392 | 0.001 |
| Triglycerides | −0.245 | 0.036 | −0.043 | 0.379 |
| Hcy | −0.437 | 0.003 | −0.298 | 0.033 |
| NO | 0.273 | 0.047 | 0.100 | 0.273 |
| Folate | 0.496 | 0.001 | 0.210 | 0.100 |
| VitB12 | 0.435 | <0.0001 | 0.231 | 0.029 |
| Supplement | Non-Obese Controls (n = 40) | Obese Bariatric Patients (n = 40) |
| Folate (1 mg/d) | 0 (0%) | 3 (7.5%) |
| Vitamin B12 (250–5000 mcg/d) | 0 (0%) | 12 (30%) |
| Alcohol | Non-Obese Controls (n = 40) | Obese Bariatric Patients (n = 40) |
| Abstainers | 25 (63%) | 22 (55%) |
| Light drinkers | 8 (20%) | 13 (32.5%) |
| Moderate drinkers | 7 (17%) | 5 (12.5%) |
| Heavy drinkers | 0 (0%) | 0 (0%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haloul, M.; Vinjamuri, S.J.; Naquiallah, D.; Mirza, M.I.; Qureshi, M.; Hassan, C.; Masrur, M.; Bianco, F.M.; Frederick, P.; Cristoforo, G.P.; et al. Hyperhomocysteinemia and Low Folate and Vitamin B12 Are Associated with Vascular Dysfunction and Impaired Nitric Oxide Sensitivity in Morbidly Obese Patients. Nutrients 2020, 12, 2014. https://doi.org/10.3390/nu12072014
Haloul M, Vinjamuri SJ, Naquiallah D, Mirza MI, Qureshi M, Hassan C, Masrur M, Bianco FM, Frederick P, Cristoforo GP, et al. Hyperhomocysteinemia and Low Folate and Vitamin B12 Are Associated with Vascular Dysfunction and Impaired Nitric Oxide Sensitivity in Morbidly Obese Patients. Nutrients. 2020; 12(7):2014. https://doi.org/10.3390/nu12072014
Chicago/Turabian StyleHaloul, Mohamed, Smita Jagdish Vinjamuri, Dina Naquiallah, Mohammed Imaduddin Mirza, Maryam Qureshi, Chandra Hassan, Mario Masrur, Francesco M. Bianco, Patrice Frederick, Giulianotti P. Cristoforo, and et al. 2020. "Hyperhomocysteinemia and Low Folate and Vitamin B12 Are Associated with Vascular Dysfunction and Impaired Nitric Oxide Sensitivity in Morbidly Obese Patients" Nutrients 12, no. 7: 2014. https://doi.org/10.3390/nu12072014