A Glycosaminoglycan-Rich Fraction from Sea Cucumber Isostichopus badionotus Has Potent Anti-Inflammatory Properties In Vitro and In Vivo

 , , , , , , , and
, , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sea Cucumber Collection and Processing
2.2. Preliminary Extractions
2.3. Large-Scale Extraction and Characterization of Glycosaminoglycans (GAGs)
2.4. Chemical Characterization of GAGs Fraction
2.5. Infrared Spectroscopy
2.6. NMR Spectroscopy
2.7. Polyacrylamide Gel Electrophoresis
2.8. Action of GAGs on TPA-Stimulated HaCaT (Human Keratinocyte) Cells and Mouse Splenocytes In Vitro
2.9. Action of GAGs on the Viability of Fibroblasts In Vitro
2.10. Experimental Animals
2.11. Mouse Ear Anti-Inflammatory Activity
2.12. Histological Analysis of Ear Samples
2.13. Dextran Sodium Sulfate (DSS)-Colitis
2.14. Gene Expression
2.15. Statistical Analysis
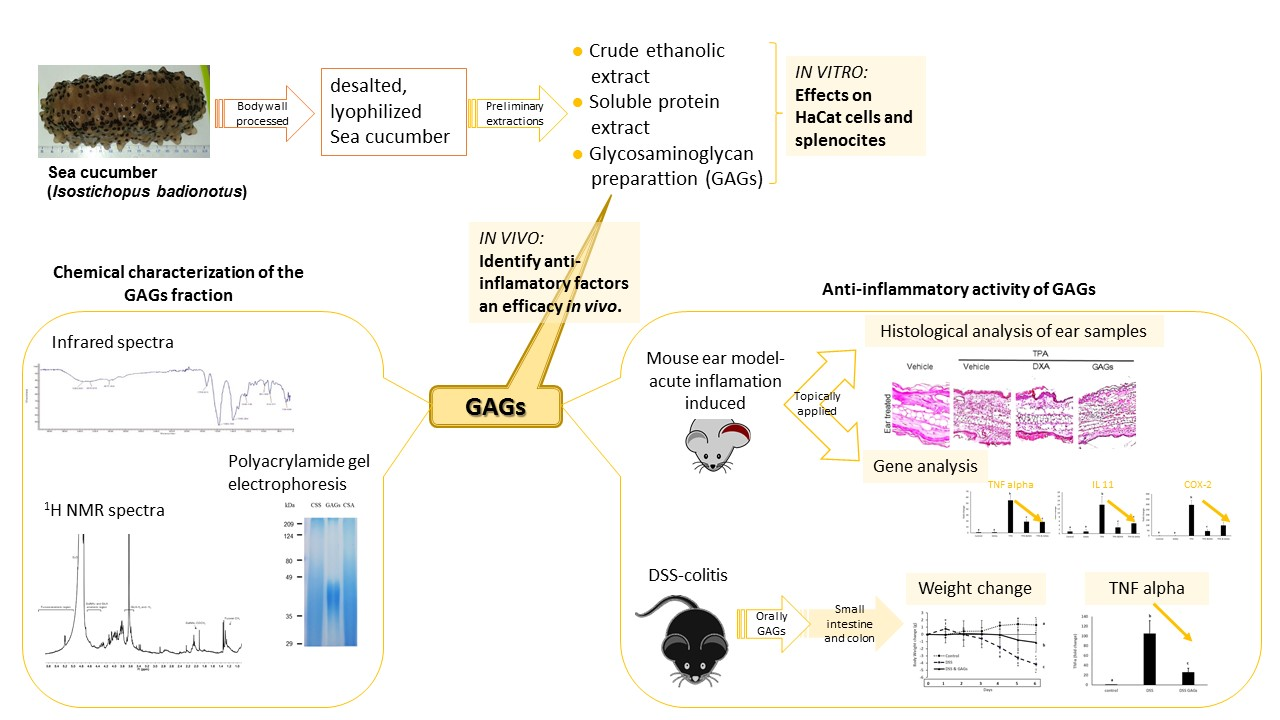
3. Results
3.1. Extracts from the Body-Wall of I. badionotus Inhibit TPA-Induced Proliferation of Keratinocyte and Splenocyte Cells in Culture
3.2. Large-Scale Preparation and Characterization of GAGs
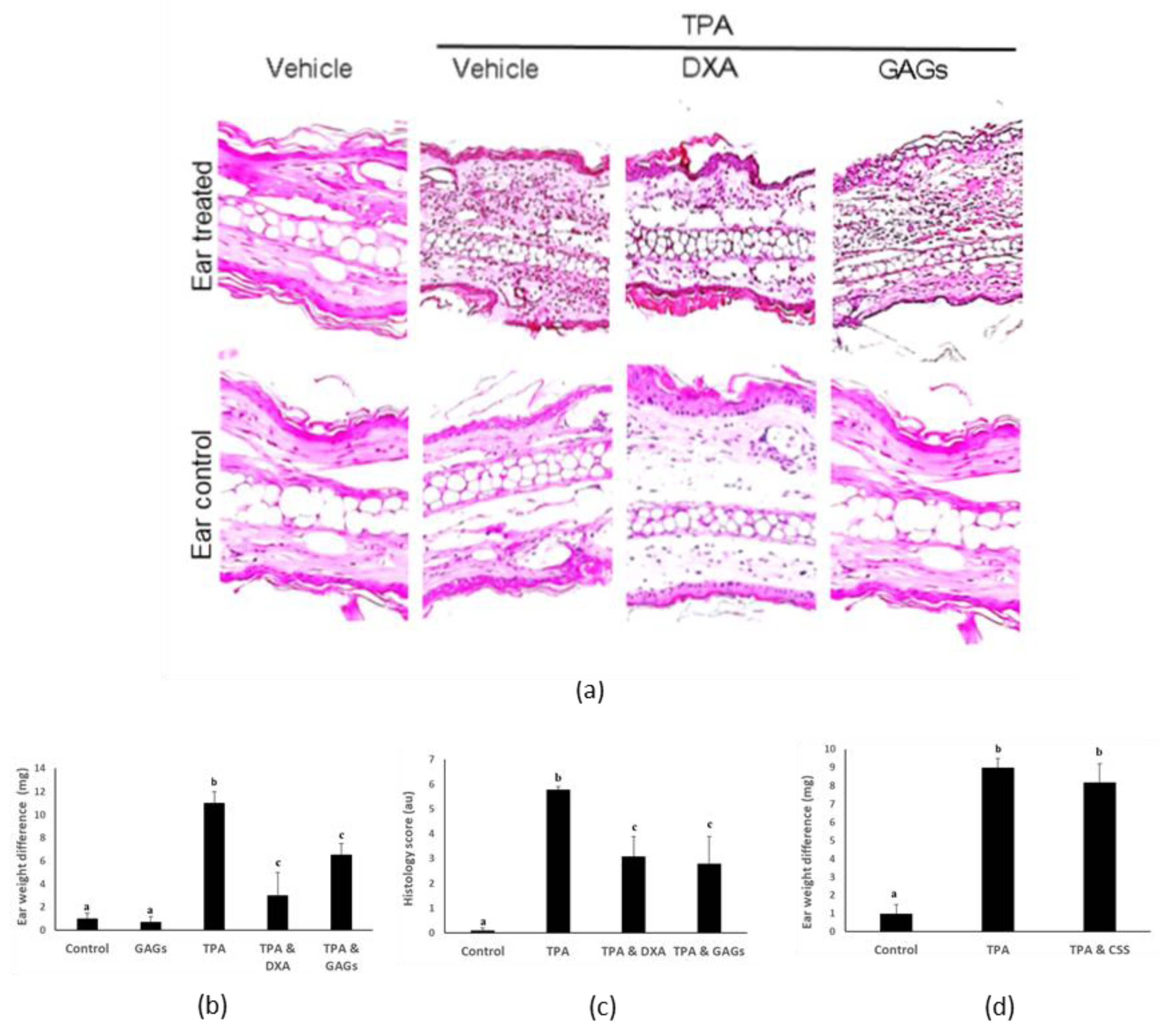
3.3. FCS Exerts Anti-Inflammatory Activity In Vivo
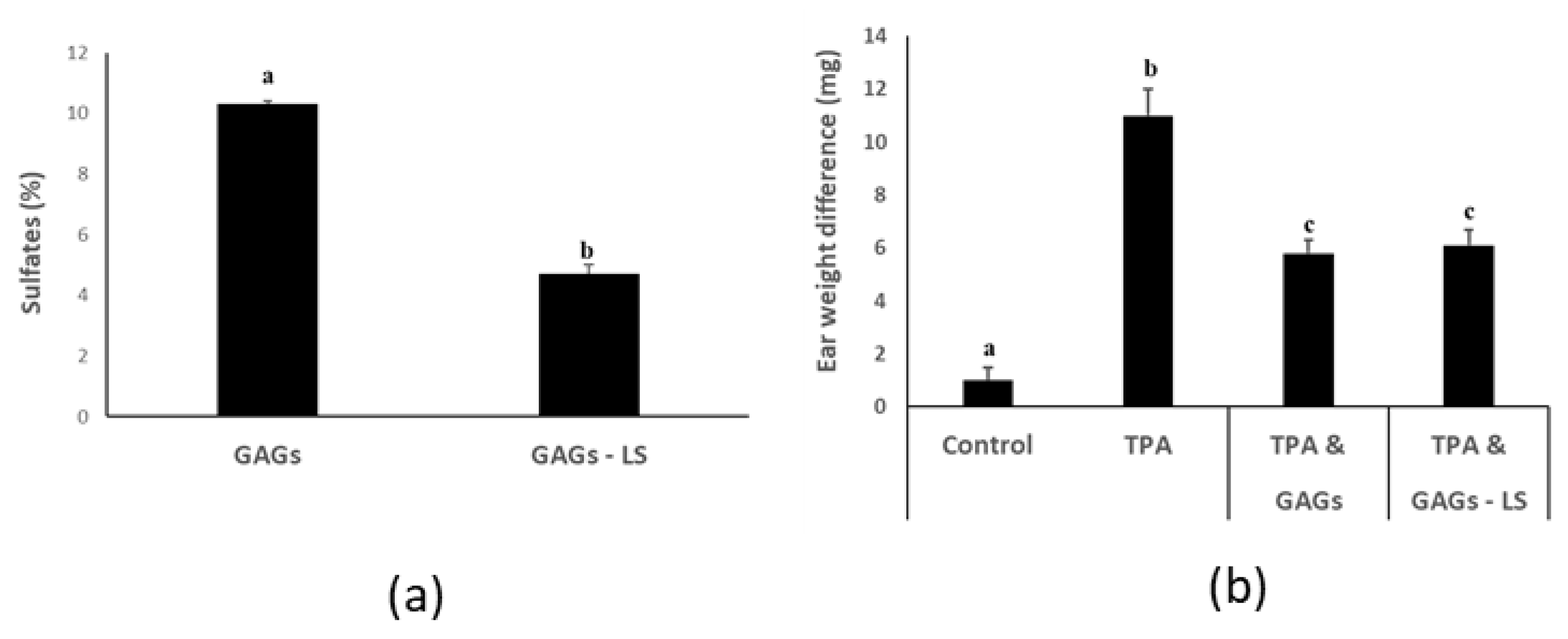
3.4. FCS from I. badionotus with Sulfate Levels of 4.5% or Above Have Similar Anti-Inflammatory Properties
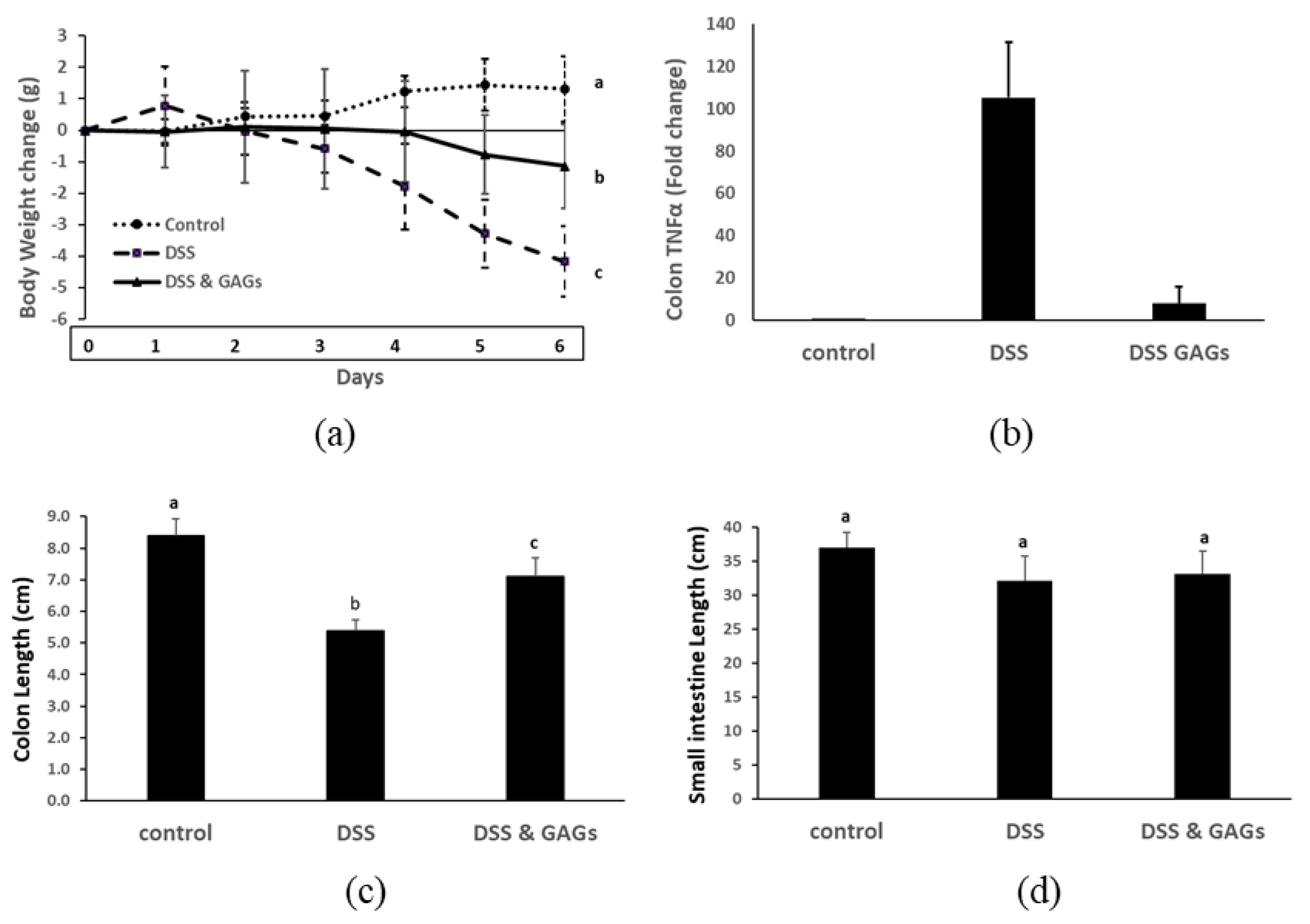
3.5. Orally-Administered FCS from I. badionotus Ameliorate DSS-Colitis in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Khotimchenko, Y. Pharmacological Potential of Sea Cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef] [Green Version]
- Pangestuti, R.; Arifin, Z. Medicinal and health benefit effects of functional sea cucumbers. J. Tradit. Complement. Med. 2018, 8, 341–351. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wu, S.; Wang, L.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber modulates the gut microbiota and its metabolites in normal mice. Int. J. Boil. Macromol. 2018, 120, 502–512. [Google Scholar] [CrossRef]
- Gamboa-Álvarez, M.Á.; López-Rocha, J.A.; Poot-López, G.R.; Aguilar-Perera, A.; Villegas-Hernández, H. Rise and decline of the sea cucumber fishery in Campeche Bank, Mexico. Ocean Coast. Manag. 2020, 184, 105011. [Google Scholar] [CrossRef]
- López-Rocha, J.A.; Velázquez-Abunader, I. Fast decline of the sea cucumber Isostichopus badionotus as a consequence of high exploitation in Yucatan, Mexico. Reg. Stud. Mar. Sci. 2019, 27, 100547. [Google Scholar] [CrossRef]
- Olivera-Castillo, L.; Grant, G.; Kantún-Moreno, N.; Acevedo, J.J.; Puc-Sosa, M.; Montero, J.; Olvera-Novoa, M.A.; Negrete-León, E.; Santa-Olalla, J.; Ceballos-Zapata, J.; et al. Sea cucumber (Isostichopus badionotus) body-wall preparations exert anti-inflammatory activity in vivo. PharmaNutrition 2018, 6, 74–80. [Google Scholar] [CrossRef]
- Olivera-Castillo, L.; Davalos, A.; Grant, G.; Valadez-González, N.; Montero, J.; Barrera-Pérez, H.A.M.; Chi, Y.A.C.; Olvera-Novoa, M.A.; Ceja-Moreno, V.; Acereto-Escoffié, P.; et al. Diets Containing Sea Cucumber (Isostichopus badionotus) Meals Are Hypocholesterolemic in Young Rats. PLoS ONE 2013, 8, e79446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Gao, Z.; Zhang, L.; Guo, F.; Chen, Y.; Li, Y.; Huang, C. Saponin-enriched sea cucumber extracts exhibit an antiobesity effect through inhibition of pancreatic lipase activity and upregulation of LXR-β signaling. Pharm. Boil. 2015, 54, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridzwan, B.H.; Farah Hanis, Z.; Daud, J.M.; Althunibat, O.Y. Protein profiles of three species of Malaysian sea cucumber; Holothuria edulis Lesson, H. scabra Jaeger and Stichopus horrens Selenka. Eur. J. Sci. Res. 2012, 75, 255–261. [Google Scholar]
- Vieira, R.P.; A Mourão, P. Occurrence of a unique fucose-branched chondroitin sulfate in the body wall of a sea cucumber. J. Boil. Chem. 1988, 263, 18176–18183. [Google Scholar]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Bitter, T.; Muir, H. A modified uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Craigie, J.S.; Wen, Z.C.; Van Der Meer, J.P.; Wen, J.S.C.Z.C. Interspecific, Intraspecific and Nutritionally-Determined Variations in the Composition of. Agars from Gracilaria spp. Bot. Mar. 1984, 27, 55–62. [Google Scholar] [CrossRef]
- Dische, Z. New color reactions for determination of sugars in polysaccharides. Methods Biochem. Anal. 1955, 2, 313–358. [Google Scholar] [PubMed]
- Magcwebeba, T.U.; Swart, A.C.; Swanvelder, S.; Joubert, E.; Gelderblom, W. Anti-Inflammatory Effects of Aspalathus linearis and Cyclopia spp. Extracts in a UVB/Keratinocyte (HaCaT) Model Utilising Interleukin-1α Accumulation as Biomarker. Molecules 2016, 21, 1323. [Google Scholar] [CrossRef] [Green Version]
- Medina-Contreras, O.; Harusato, A.; Nishio, H.; Flannigan, K.L.; Ngo, V.L.; Leoni, G.; Neumann, P.-A.; Geem, D.; Lili, L.N.; Ramadas, R.A.; et al. Cutting Edge: IL-36 Receptor Promotes Resolution of Intestinal Damage. J. Immunol. 2015, 196, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Caamal-Herrera, I.O.; Carrillo-Cocom, L.M.; Escalante-Réndiz, D.Y.; Araiz-Hernández, D.; Azamar-Barrios, J.A. Antimicrobial and antiproliferative activity of essential oil, aqueous and ethanolic extracts of Ocimum micranthum Willd leaves. BMC Complement. Altern. Med. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Gábor, M. Models of Acute Inflammation in the Ear. Inflamm. Protoc. 2003, 225, 129–138. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Delday, M.; Mulder, I.; Logan, E.T.; Grant, G. Bacteroides thetaiotaomicronAmeliorates Colon Inflammation in Preclinical Models of Crohn’s Disease. Inflamm. Bowel Dis. 2018, 25, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Mao, G.; Wu, T.; Lin, D.; Hu, Y.; Ye, X.; Tian, D.; Chai, W.; Linhardt, R.J.; et al. Fucosylated chondroitin sulfate from Isostichopus badionotus alleviates metabolic syndromes and gut microbiota dysbiosis induced by high-fat and high-fructose diet. Int. J. Boil. Macromol. 2018, 124, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Mao, G.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. Fucosylated chondroitin sulfate oligosaccharides from Isostichopus badionotus regulates lipid disorder in C57BL/6 mice fed a high-fat diet. Carbohydr. Polym. 2018, 201, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Mao, G.; Yan, L.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. Effect of the sulfation pattern of sea cucumber-derived fucoidan oligosaccharides on modulating metabolic syndromes and gut microbiota dysbiosis caused by HFD in mice. J. Funct. Foods 2019, 55, 193–210. [Google Scholar] [CrossRef]
- Yin, J.; Yang, X.; Xia, B.; Yang, Z.; Wang, Z.; Wang, J.; Li, T.; Lin, P.; Song, X.; Guo, S. The fucoidan from sea cucumber Apostichopus japonicus attenuates lipopolysaccharide-challenged liver injury in C57BL/6J mice. J. Funct. Foods 2019, 61, 103493. [Google Scholar] [CrossRef]
- Xu, X.; Chang, Y.; Xue, C.; Wang, J.; Shen, J. Gastric Protective Activities of Sea Cucumber Fucoidans with Different Molecular Weight and Chain Conformations: A Structure–Activity Relationship Investigation. J. Agric. Food Chem. 2018, 66, 8615–8622. [Google Scholar] [CrossRef]
- Panagos, C.G.; Thomson, D.S.; Moss, C.; Hughes, A.D.; Kelly, M.S.; Liu, Y.; Chai, W.; Venkatasamy, R.; Spina, D.; Page, C.P.; et al. Fucosylated chondroitin sulfates from the body wall of the sea cucumber Holothuria forskali: Conformation, selectin binding, and biological activity. J. Boil. Chem. 2014, 289, 28284–28298. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, J.; Zhi, Z.; Wei, C.; Wang, W.; Ding, T.; Ye, X.; Hu, Y.; Linhardt, R.J.; Chen, S. Macromolecular properties and hypolipidemic effects of four sulfated polysaccharides from sea cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef]
- Pomin, V.H.; Mourão, P.A.D.S. Specific sulfation and glycosylation—A structural combination for the anticoagulation of marine carbohydrates. Front. Microbiol. 2014, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Wu, M.; Xu, L.; Lian, W.; Xiang, J.; Lu, F.; Zhao, J. Comparison of physicochemical characteristics and anti-coagulant activities of polysaccharides from three sea cucumbers. Mar. Drugs 2013, 11, 399–417. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Zhang, Y.; Ye, X.; Hu, Y.; Ding, T.; Chen, S. Sulfation pattern of fucose branches affects the anti-hyperlipidemic activities of fucosylated chondroitin sulfate. Carbohydr. Polym. 2016, 147, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustyuzhanina, N.E.; Bilan, M.I.; Nifantiev, N.E.; Usov, A.I. New insight on the structural diversity of holothurian fucosylated chondroitin sulfates. Pure Appl. Chem. 2019, 91, 1065–1071. [Google Scholar] [CrossRef]
- Lee, J.; Kang, U.; Seo, E.K.; Kim, Y.S. Heme oxygenase-1-mediated anti-inflammatory effects of tussilagonone on macrophages and 12-O-tetradecanoylphorbol-13-acetate-induced skin inflammation in mice. Int. Immunopharmacol. 2016, 34, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.S.; Chen, B.C.; Yu, M.T.; Sheu, J.R.; Chen, T.F.; Lin, C.H. Phorbol 12-myristate 13-acetate up-regulates cyclooxygenase-2 expression in human pulmonary epithelial cells via Ras, Raf-1, ERK, and NF-κB, but not p38 MAPK, pathways. Cell. Signal. 2005, 17, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, D.; Wang, S.; Tao, L.; Wang, A.; Chen, W.; Zhu, Z.; Zheng, S.; Gao, X.; Lu, Y. Holothurian Glycosaminoglycan Inhibits Metastasis and Thrombosis via Targeting of Nuclear Factor-κB/Tissue Factor/Factor Xa Pathway in Melanoma B16F10 Cells. PLoS ONE 2013, 8, e56557. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Jiang, W.; Hu, S.; Song, W.; Ji, L.; Wang, Y.; Cai, L. Fucosylated chondroitin sulphate from Cusumaria frondosa mitigates hepatic endoplasmic reticulum stress and inflammation in insulin resistant mice. Food Funct. 2015, 6, 1547–1556. [Google Scholar] [CrossRef]
- Luo, Y.; Zheng, S. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms. Front. Immunol. 2016, 7, 503. [Google Scholar] [CrossRef]
- Zhang, R.; Yuan, S.; Ye, J.; Wang, X.; Zhang, X.; Shen, J.; Yuan, M.; Liao, W. Polysaccharide from flammuliana velutipes improves colitis via regulation of colonic microbial dysbiosis and inflammatory responses. Int. J. Boil. Macromol. 2020, 149, 1252–1261. [Google Scholar] [CrossRef]
- Wu, H.; Rao, Q.; Ma, G.-C.; Yu, X.-H.; Zhang, C.-E.; Ma, Z.-J. Effect of Triptolide on Dextran Sodium Sulfate-Induced Ulcerative Colitis and Gut Microbiota in Mice. Front. Pharmacol. 2020, 10, 1652. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Xu, Y.; Yang, H.; Wang, J.; Xue, C.; Yan, X.; Su, L. Anti-inflammation effects of fucosylated chondroitin sulphate from Acaudina molpadioides by altering gut microbiota in obese mice. Food Funct. 2019, 10, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Guo, C.; Li, X. Chitosan Ameliorates DSS-Induced Ulcerative Colitis Mice by Enhancing Intestinal Barrier Function and Improving Microflora. Int. J. Mol. Sci. 2019, 20, 5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Chen, Y.; Wang, G.; Yang, Y.; Song, X.; Xiong, Z.; Zhang, H.; Lai, P.; Wang, S.; Ai, L. Lactobacillus plantarum AR113 alleviates DSS-induced colitis by regulating the TLR4/MyD88/NF-κB pathway and gut microbiota composition. J. Funct. Foods 2020, 67, 103854. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAGs | CSS | |
|---|---|---|
| Total CHO | 24.77 ± 2.5 | 25.36 ± 0.06 |
| Fucose | 6.40 ± 0.54 | ND |
| Uronic acid | 8.55 ± 0.48 | 20.6 ± 0.04 |
| Sulfates | 11.12 ± 0.60 | 15.06 ± 0.001 |
| Soluble proteins | 4.15 ± 0.01 | 5.01 ± 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivera-Castillo, L.; Grant, G.; Kantún-Moreno, N.; Barrera-Pérez, H.A.; Montero, J.; Olvera-Novoa, M.A.; Carrillo-Cocom, L.M.; Acevedo, J.J.; Puerto-Castillo, C.; May Solís, V.; et al. A Glycosaminoglycan-Rich Fraction from Sea Cucumber Isostichopus badionotus Has Potent Anti-Inflammatory Properties In Vitro and In Vivo. Nutrients 2020, 12, 1698. https://doi.org/10.3390/nu12061698
Olivera-Castillo L, Grant G, Kantún-Moreno N, Barrera-Pérez HA, Montero J, Olvera-Novoa MA, Carrillo-Cocom LM, Acevedo JJ, Puerto-Castillo C, May Solís V, et al. A Glycosaminoglycan-Rich Fraction from Sea Cucumber Isostichopus badionotus Has Potent Anti-Inflammatory Properties In Vitro and In Vivo. Nutrients. 2020; 12(6):1698. https://doi.org/10.3390/nu12061698
Chicago/Turabian StyleOlivera-Castillo, Leticia, George Grant, Nuvia Kantún-Moreno, Hirian A. Barrera-Pérez, Jorge Montero, Miguel A. Olvera-Novoa, Leydi M. Carrillo-Cocom, Juan J. Acevedo, Cesar Puerto-Castillo, Victor May Solís, and et al. 2020. "A Glycosaminoglycan-Rich Fraction from Sea Cucumber Isostichopus badionotus Has Potent Anti-Inflammatory Properties In Vitro and In Vivo" Nutrients 12, no. 6: 1698. https://doi.org/10.3390/nu12061698