ADORA2A C Allele Carriers Exhibit Ergogenic Responses to Caffeine Supplementation
by
 , , , and
, , , and
Jozo Grgic
1,*, 
Craig Pickering
2,
David J. Bishop
1,3,
Juan Del Coso
4 ,
,
Brad J. Schoenfeld
5, 
Grant M. Tinsley
6 and
Zeljko Pedisic
1 1
Institute for Health and Sport (IHES), Victoria University, Melbourne 3011, Australia
2
Institute of Coaching and Performance, School of Sport and Wellbeing, University of Central Lancashire, Preston PR1 2HE, UK
3
School of Medical and Health Sciences, Edith Cowan University, Joondalup, Perth, WA 6027, Australia
4
Centre for Sport Studies, Rey Juan Carlos University, C/Camino del Molino, s/n, 28943 Fuenlabrada, Spain
5
Department of Health Sciences, Lehman College, Bronx, NY 10468, USA
6
Energy Balance & Body Composition Laboratory, Department of Kinesiology & Sport Management, Texas Tech University, Lubbock, TX 79424, USA
*
Author to whom correspondence should be addressed.
Nutrients 2020, 12(3), 741; https://doi.org/10.3390/nu12030741
Submission received: 14 February 2020
/
Revised: 3 March 2020
/
Accepted: 9 March 2020
/
Published: 11 March 2020
(This article belongs to the Section Sports Nutrition)
Abstract
:Caffeine’s ergogenic effects on exercise performance are generally explained by its ability to bind to adenosine receptors. ADORA2A is the gene that encodes A2A subtypes of adenosine receptors. It has been suggested that ADORA2A gene polymorphisms may be responsible for the inter-individual variations in the effects of caffeine on exercise performance. In the only study that explored the influence of variation in ADORA2A—in this case, a common polymorphism (rs5751876)—on the ergogenic effects of caffeine on exercise performance, C allele carriers were identified as “non-responders” to caffeine. To explore if C allele carriers are true “non-responders” to the ergogenic effects of caffeine, in this randomized, double-blind study, we examined the acute effects of caffeine ingestion among a sample consisting exclusively of ADORA2A C allele carriers. Twenty resistance-trained men identified as ADORA2A C allele carriers (CC/CT genotype) were tested on two occasions, following the ingestion of caffeine (3 mg/kg) and a placebo. Exercise performance was evaluated with movement velocity, power output, and muscle endurance during the bench press exercise, countermovement jump height, and power output during a Wingate test. Out of the 25 analyzed variables, caffeine was ergogenic in 21 (effect size range: 0.14 to 0.96). In conclusion, ADORA2A (rs5751876) C allele carriers exhibited ergogenic responses to caffeine ingestion, with the magnitude of improvements similar to what was previously reported in the literature among samples that were not genotype-specific. Therefore, individuals with the CT/CC genotype may still consider supplementing with caffeine for acute improvements in performance.
1. Introduction
The effects of caffeine on exercise have received substantial attention in the scientific literature [1,2,3,4,5,6,7,8]. Currently, it is well established that acute ingestion of caffeine doses in the range from 2 to 6 mg per kilogram of body mass enhances exercise performance [1,2,3,4,5,6,7,8]. Caffeine’s ergogenic effects are apparent in different components of exercise. For example, a recent umbrella review reported that caffeine ingestion enhances muscle strength and endurance, aerobic endurance, power output, and jumping performance [3]. Even though research indicates that caffeine ingestion may be acutely ergogenic for a wide range of exercise tasks, between-person variability in responses to this dietary supplement seems substantial [9,10]. The ergogenic effects of caffeine are generally explained by its interaction with adenosine A1, A2A, and A2B receptors [11,12]. Adenosine concentrations in the brain progressively increase during waking hours, resulting ultimately in sensations of fatigue; the concentrations of adenosine also decrease during sleep. Caffeine’s molecular structure is similar to that of adenosine. Therefore, after ingestion, caffeine binds to adenosine receptors, subsequently resulting in reduced fatigue, increased vigilance, and ergogenic effects on exercise performance [11,12].
Researchers have suggested that the inter-individual variation in caffeine response may be due to polymorphisms within two genes, namely CYP1A2 and ADORA2A [10]. Cytochrome P450 1A2 (an enzyme responsible for up to 95% of caffeine metabolism) is encoded by the CYP1A2 gene [10]. A single nucleotide polymorphism rs762551 within CYP1A2 affects the speed of caffeine metabolism. Specifically, individuals with the AA genotype are commonly classified as “fast caffeine metabolizers”, whereas C allele carriers (AC/CC genotypes) are considered to be “slow caffeine metabolizers”, respectively [13]. The influence of CYP1A2 (rs762551) on the acute effects of caffeine supplementation on exercise performance has been explored in several studies [14,15,16,17,18,19,20,21,22,23]. However, the evidence in these studies remains inconsistent, with some reporting no effect of the polymorphism on the ergogenic effects of caffeine supplementation and others showing a modifying effect, but in different directions [14,15,16,17,18,19,20,21,22,23].
ADORA2A is the gene that encodes A2A subtypes of adenosine receptors [24]. Previous research has suggested that this receptor represents the primary target of caffeine action in the central nervous system, and thus, polymorphic variations in the ADORA2A gene may impact the responses to acute caffeine ingestion [24]. The rs5751876 polymorphisms in the ADORA2A gene are comprised of a C-to-T substitution at nucleotide position 1083 (rs5751876) (also known as 1976C>T) [24]. Interestingly, as compared to TT homozygotes, ADORA2A C allele carriers have higher habitual caffeine consumption, which may suggest that these individuals need higher doses of caffeine to obtain a pharmacological effect [24].
Only one study has explored the influence of variation in this gene—in this case, a common polymorphism (rs5751876)—on the ergogenic effects of caffeine on exercise performance [25]. The study included 12 participants (6 TT homozygotes and 6 C allele carriers [i.e., CC/CT genotype]). These participants were untrained women who completed 20 min of cycling at a work rate eliciting 60% of VO2peak followed by two 10-min cycling time trials. The exercise task was performed on two occasions, following the ingestion of 5 mg/kg of caffeine or a placebo. Results indicated that caffeine ingestion was ergogenic for TT homozygotes but not for C allele carriers. Based on this study, C allele carriers were identified as “non-responders” to caffeine [25].
Given the limited data on this topic, the aim of this study was to explore the influence of ADORA2A (rs5751876) on the acute effects of caffeine supplementation on exercise performance, by using exercise tests for which caffeine had previously been shown to be ergogenic [3].
2. Materials and Methods
2.1. Experimental Design
In this double-blind, randomized, crossover trial, all participants attended four laboratory sessions (in the morning hours between 07:00 to 12:00 h) that were from 4 to 7 days apart. The first two sessions consisted of familiarization with the exercise protocol. The third and fourth sessions were the main sessions. Twenty-four hours before the main trials, participants were asked the following: (a) to avoid any intense exercise; (b) to track their energy and macronutrient intake; and (c) to refrain from caffeine intake after 6 pm on the day before testing. The participants performed the two main sessions in a fasted state (overnight fast). Caffeine and placebo supplementation was provided on different days. Caffeine (Pure Lean Nutrition, Melbourne, Australia) was administered in a gelatin capsule with a dose of 3 mg/kg of body mass, while the placebo gelatin capsule contained 3 mg/kg of body mass of dextrose. All capsules were of identical appearance. Placebo and caffeine powders were weighed using a high precision electronic digital scale (Precisa, XT 120A, Dietikon, Switzerland) and then packaged into capsules. Capsules were prepared in the laboratory by an experienced researcher while other researchers performed the blinding. Capsules were ingested 60 min before the start of the exercise session under the supervision of the research staff, as in previous research [1,26,27]. The participants’ genotype was determined using a buccal swab. Ethical approval was requested and granted from the Victoria University Human Research Ethics Committee (number: HRE19-019), and every participant signed an informed consent form.
2.2. Participants
The study included a sample of 22 resistance-trained men, defined herein as having a minimum of six months of resistance training experience with a minimum weekly training frequency of two times on most weeks. Exclusion criteria were the existence of any health limitations and prior use of anabolic steroids (self-reported). All participants completed all sessions with no injuries or adverse events. Participants’ characteristics are presented in Table 1.
2.3. Exercise Protocol
Exercises involving the upper body were performed prior to those that predominately activated the lower body, to avoid any transfer of muscle fatigue from one exercise task to another. At the beginning of the exercise protocol, the participants performed the bench press exercise with different loads (i.e., 25%, 50%, 75%, and 90% of one-repetition maximum (1RM)—performed in that order) [28]. 1RM was established during the first familiarization session. At each respective load, the participants performed two sets of one repetition, separated by a 3-min rest interval. The better repetition at each load was used for the analysis. The eccentric phase lasted 2 s, there was no pause at the bottom phase, and the concentric action was performed with maximal velocity. Mean power (W), mean concentric velocity (m/s), peak power (W), and peak concentric velocity (m/s) were measured for each repetition using the GymAware linear position transducer device (GymAware Power Tool, Kinetic Performance Technologies, Canberra, Australia) that was attached to the barbell.
After the second set that was performed with 90% of 1RM, the participants were provided with five minutes of rest. Then, we tested upper-body muscular endurance with a task that involved performing repetitions to momentary muscular failure in the bench press exercise with a load of 85% of 1RM. In this test, we collected data on the total number of repetitions, as well as power and velocity output of each repetition using the linear position transducer attached to the barbell. The tempo was the same as in the previous task. For the statistical analysis, we compared the total number of repetitions between the placebo and caffeine conditions. In addition, to explore the “quality” of performed repetitions, we matched the number of repetitions between the placebo and caffeine conditions and examined their average power and velocity. For example, one participant performed 7 and 8 repetitions following the ingestion of the placebo and caffeine, respectively. In this case, we only examined the velocity and power of the first 7 repetitions in both conditions.
After the muscular endurance test, the participants rested for three minutes. Then the participants performed a short warm-up consisting of one minute of light running, followed by ten bodyweight squats. After the warm-up, participants performed a countermovement jump (CMJ) without an arm swing on a force platform (400S Isotronic Fitness Technology, Skye, Australia). The participants positioned themselves in an upright starting position and received commands from the computer software associated with the force platform that was positioned in front of the platform. This software visually counted down, “3, 2, 1” and provided “Set” and “Go” commands. After the “Go” command, the participants had five seconds to complete the jump. The participants performed a fast knee flexion (where their lowest position was a semi-squat position) [29,30]. Immediately after reaching this point (i.e., no pause at the bottom phase), the participants rapidly extended the hip, knee, and ankle joints with prior instructions to jump as quickly and “explosively” as possible to achieve maximal vertical jump height [29,30]. A total of three attempts was provided with one minute of rest between them. The best jump was used for the analysis. The outcome in the CMJ test was vertical jump height.
After the CMJ, the participants rested for three minutes. Then, the participants performed the Wingate test on an Excalibur Sport Cycle Ergometer (Lode, Groningen, The Netherlands). The Wingate test started with a 5-min warm-up consisting of pedaling at 100 W at 60–80 rpm [31]. Following the warm-up, participants performed a 30-s “all-out” sprint on the bike. The flywheel resistance was set at 0.075 Nm/kg. The participants were instructed to remain seated during the 30-s sprint.
2.4. Assessment of Blinding
In both main trials (i.e., caffeine and placebo), before and after the testing session, participants responded to the following question: “Which supplement do you think you have ingested?” [32]. This question was used to explore the effectiveness of the blinding and had three possible responses: (a) “caffeine”, (b) “placebo”, and (c) “I do not know” [32]. If the participants responded with “a” or “b”, they were also asked to state the reason for choosing their respective response.
2.5. Genetic Testing
Genetic testing was performed using a commercially available testing kit from DNAfit Life Sciences. The procedure used for genetic testing is explained in detail elsewhere [33]. Briefly, the buccal swab sample was collected using OCR-100 kits by DNAGenotek. For the analysis, these samples were sent to IDna Genetics Laboratory (Norwich, UK). DNA was: (a) extracted and purified using the Isohelix Buccalyse DNA extraction kit BEK-50 (Kent, UK); and (b) amplified using polymerase chain reaction (PCR) on an ABI 7900 real-time thermocycler (Applied Biosystem, Waltham, MA, USA). The collected samples were analyzed for the ADORA2A (rs5751876) single-nucleotide polymorphism. Genotype analyses were performed after the exercise performance data collection was finalized. Therefore, researchers and participants were blinded to genotype variations of the sample during the exercise performance data collection.
2.6. Statistical Analysis
Two participants who were ADORA2A TT homozygotes were excluded, leaving a total of 20 C allele carriers (CC and CT) in the analysis. One-way repeated-measures analysis of variance (ANOVA) was used to analyze the exercise performance data. Relative effect sizes (and their 95% confidence intervals; 95% CI) were expressed using Hedges’ g for repeated measures. The effect sizes were classified as follows: trivial (<0.20); small (0.20–0.49); moderate (0.50–0.79); and large (≥0.80). The effectiveness of blinding was examined using the Bang’s Blinding Index, as explained elsewhere [29]. All analyses were performed using the Statistica software (version 13.0; StatSoft; Tulsa, OK, USA). The significance level was set at p < 0.05.
3. Results
3.1. Exercise Performance
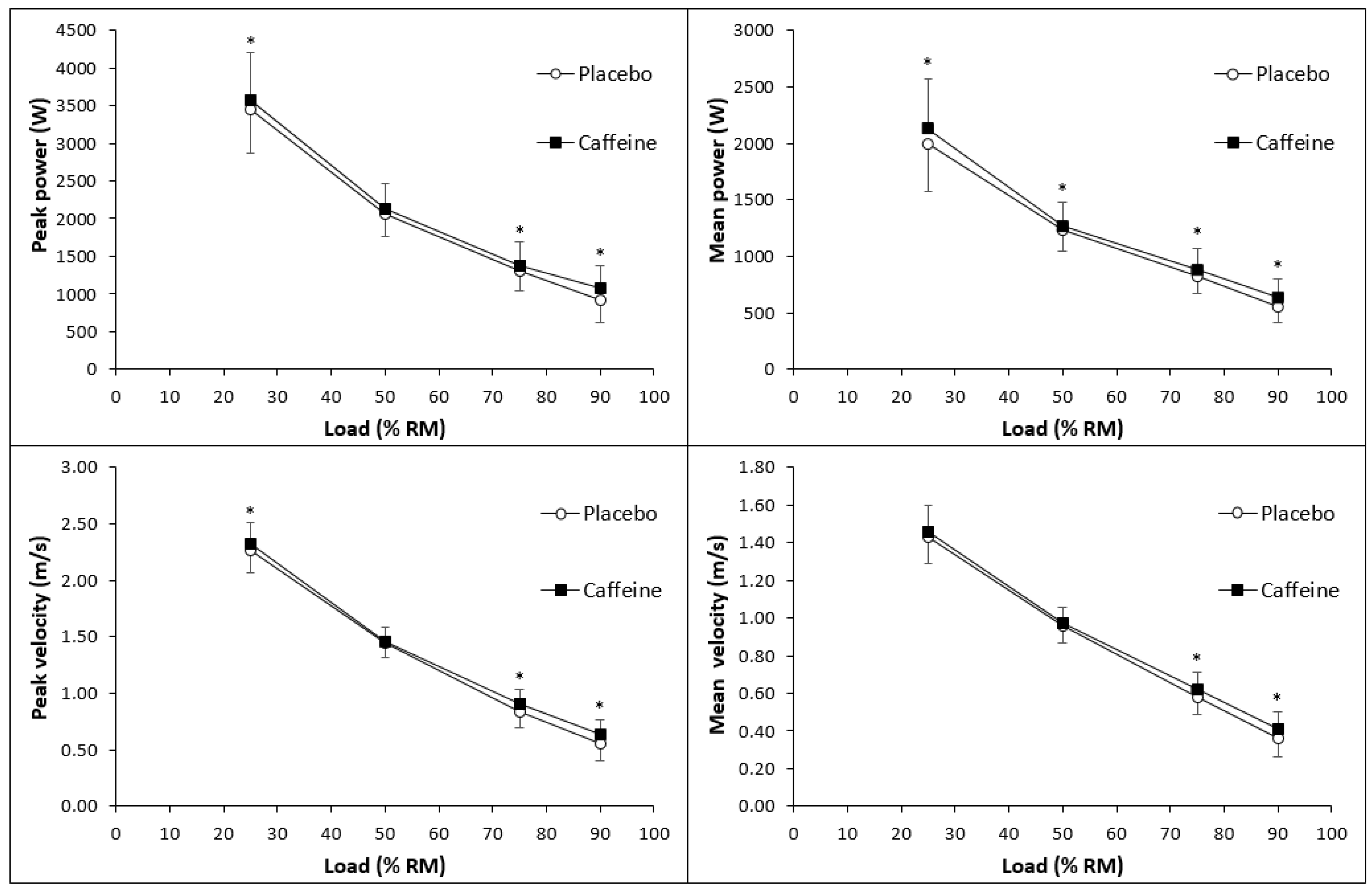
For movement velocity and power, we found significant effects of caffeine ingestion for all outcomes except for mean velocity at 25% of 1RM, and mean velocity, peak power, and peak velocity at 50% of 1RM (Figure 1). The significant effect sizes ranged from 0.16 to 0.53. For muscular endurance, we found significant effects of caffeine ingestion on the total number of performed repetitions and the quality of repetitions when matched for repetitions between the conditions. Here, the effect sizes ranged from 0.27 to 0.96 (Table 2). We also found a significant effect of caffeine ingestion on vertical jump height with an effect size of 0.13. For power output in the Wingate test, we found significant effects of caffeine ingestion on peak, mean, and minimum power. The effect sizes ranged from 0.34 to 0.41.
3.2. Assessment of Blinding
Before the start of the exercise session, 50% and 65% of the participants correctly guessed (beyond chance) the placebo and caffeine conditions, respectively. After finishing the exercise session, 65% and 75% of the participants correctly guessed the placebo and caffeine conditions beyond chance, respectively. Participants who correctly identified caffeine reported “feeling more energized” and/or “more alert”, or they associated the improvements in exercise performance with caffeine ingestion.
4. Discussion
The main finding of this study is that caffeine ingestion may be ergogenic for ADORA2A (rs5751876) C allele carriers in a range of exercise performance outcomes. Therefore, these results do not support the theoretical supposition that ADORA2A C allele carriers do not experience improvements in exercise performance following caffeine ingestion.
Our findings are not in accord with the Loy et al. [25] study, which proposed that ADORA2A C allele carriers do not experience an ergogenic response to caffeine supplementation. The main differences between our study and Loy et al. [25] are the sex of the participants and the exercise tests employed. Specifically, we included male participants, whereas Loy and colleagues included females. Therefore, it may be that female ADORA2A C allele carriers experience a different response to caffeine ingestion as compared to their male counterparts. However, this explanation is perhaps less plausible because recent evidence suggests that female and male participants experience similar ergogenic responses to caffeine ingestion in aerobic-, anaerobic- and strength-based exercise tasks [34,35,36]. Importantly, the present study and the work by Loy et al. [25] also differed in the selection of performance tests; while we assessed changes in power, muscular endurance, and sprinting performance, Loy and colleagues focused on aerobic endurance. It may be that caffeine affects performance in these components of exercise performance through different mechanisms. The possible impact of genetic variations may be more expressed in some tests and less in others. Given the scarce evidence on the influence of polymorphisms in ADORA2A on the individual variation in responses to caffeine, this topic certainly requires further research. Finally, given that we report here that ADORA2A C allele carriers improve performance following caffeine ingestion, this may suggest that other genotypes that were not tested herein (e.g., CYP1A2 AA and AC/CC genotypes) are more important for the individual responses to caffeine ingestion.
Interestingly, the effects of caffeine on exercise performance in this study were very similar in size to the effects previously reported in the literature. For example, the increases in muscular endurance in our study are similar to the performance benefits of caffeine recorded in a previous study that included individuals with CYP1A2 (rs762551) AA genotype—which are suggested to experience the most profound ergogenic benefits of caffeine [22]. Furthermore, the increases in movement velocity, vertical jump height, and power output in the Wingate test are comparable to the improvements reported in meta-analyses of these outcomes among samples that were not genotype-specific [5,7,37]. For example, one meta-analysis [7] reported that caffeine ingestion acutely enhanced Wingate peak power by an effect size of 0.27 (95% CI: 0.08, 0.47), which is very similar to the effect size of 0.37 (95% CI: 0.21, 0.55) observed in this study.
Strengths and Limitations
The main strength of the present study was the use of a randomized, double-blind study design, which is identified as the gold standard in sports nutrition [38]. Additionally, the strength of the present study was in the use of exercise tests for which caffeine had been shown to be ergogenic.
The main limitation of this study was that 50% to 75% of the participants were able to identify caffeine and placebo conditions beyond chance. However, these results were not a likely explanation of the differences in findings between our study and the Loy et al. [25] study, given that the majority of participants (>75%) in the Loy et al. study were able to guess the content of the capsules correctly. Additionally, given the small number of ADORA2A TT homozygotes in our sample, we could not assess whether TT homozygotes experience different responses to caffeine ingestion compared with C allele carriers, an area that should be explored in future research. The low number of participants classified as TT homozygotes could be explained by the estimate that around 85% of the population possess the CC/CT genotype at rs5751876 [39].
Finally, to avoid any potential confounding by prior food and caffeine ingestion [40,41], we opted to test the participants in a fasted state. This needs to be acknowledged as a limitation given that caffeine supplementation and exercise in a fasted state is likely not a “real-life” practice of many individuals, and is not in line with the current sports nutrition recommendations [42]. Future studies may consider further exploring this topic by using caffeine supplementation protocols that mirror those more commonly observed in practice.
5. Conclusions
Our findings suggest that ADORA2A (rs5751876) C allele carriers respond positively to caffeine supplementation. Therefore, individuals with the CT/CC genotype may still consider supplementing with caffeine for acute improvements in performance. Future research is needed to explore if ADORA2A TT homozygotes experience different responses to caffeine supplementation than C allele carriers.
Author Contributions
Conceptualization, J.G., C.P., D.J.B., and Z.P.; Data curation, J.G.; Methodology, J.G., C.P., D.J.B., and Z.P.; Project administration, D.J.B. and Z.P.; Resources, C.P., D.J.B., J.D.C., and Z.P.; Visualization, J.G. and J.D.C.; Formal analysis, J.G.; Investigation, J.G.; Supervision, D.J.B. and Z.P.; Data interpretation, J.G., C.P., D.J.B., J.D.C., B.J.S., G.M.T., and Z.P. Writing—original draft, J.G., C.P., D.J.B., J.D.C., B.J.S., G.M.T., and Z.P.; Writing—review and editing, J.G., C.P., D.J.B., J.D.C., B.J.S., G.T., and Z.P. Revisions and responses to reviewers, J.G., C.P., D.J.B., J.D.C., B.J.S., G.M.T., and Z.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
Craig Pickering is a former employee of DNAFit Life Sciences, a genetic testing company. He received no financial incentives for the preparation of this manuscript. All other authors declare no conflict of interest.
References
- Graham, T.E. Caffeine and exercise: Metabolism, endurance and performance. Sports. Med. 2001, 31, 785–807. [Google Scholar] [CrossRef] [PubMed]
- McLellan, T.M.; Caldwell, J.A.; Lieberman, H.R. A review of caffeine’s effects on cognitive, physical and occupational performance. Neurosci. Biobehav. Rev. 2016, 71, 294–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grgic, J.; Grgic, I.; Pickering, C.; Schoenfeld, B.J.; Bishop, D.J.; Pedisic, Z. Wake up and smell the coffee: Caffeine supplementation and exercise performance-an umbrella review of 21 published meta-analyses. Br. J. Sports. Med. 2019. [Google Scholar] [CrossRef]
- Salinero, J.J.; Lara, B.; Del Coso, J. Effects of acute ingestion of caffeine on team sports performance: A systematic review and meta-analysis. Res. Sports Med. 2019, 27, 238–256. [Google Scholar] [CrossRef]
- Grgic, J.; Trexler, E.T.; Lazinica, B.; Pedisic, Z. Effects of caffeine intake on muscle strength and power: A systematic review and meta-analysis. J. Int. Soc. Sports Nutr. 2018, 15, 11. [Google Scholar] [CrossRef] [Green Version]
- Grgic, J.; Pickering, C. The effects of caffeine ingestion on isokinetic muscular strength: A meta-analysis. J. Sci. Med. Sport 2019, 22, 353–360. [Google Scholar] [CrossRef]
- Grgic, J. Caffeine ingestion enhances Wingate performance: A meta-analysis. Eur. J. Sport. Sci. 2018, 18, 219–225. [Google Scholar] [CrossRef]
- Grgic, J.; Mikulic, P.; Schoenfeld, B.J.; Bishop, D.J.; Pedisic, Z. The Influence of Caffeine Supplementation on Resistance Exercise: A Review. Sports Med. 2019, 49, 17–30. [Google Scholar] [CrossRef]
- Grgic, J. Are There Non-Responders to the Ergogenic Effects of Caffeine Ingestion on Exercise Performance? Nutrients 2018, 10, 1736. [Google Scholar] [CrossRef] [Green Version]
- Pickering, C.; Kiely, J. Are the Current Guidelines on Caffeine Use in Sport Optimal for Everyone? Inter-individual Variation in Caffeine Ergogenicity, and a Move Towards Personalised Sports Nutrition. Sports Med. 2018, 48, 7–16. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Yang, J.; Wang, Y. Low, but not high, dose caffeine is a readily available probe for adenosine actions. Mol. Asp. Med. 2017, 55, 20–25. [Google Scholar] [CrossRef]
- Davis, J.M.; Zhao, Z.; Stock, H.S.; Mehl, K.A.; Buggy, J.; Hand, G.A. Central nervous system effects of caffeine and adenosine on fatigue. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R399–R404. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, N.; Ghotbi, R.; Jankovic, S.; Aklillu, E. Induction of CYP1A2 by heavy coffee consumption is associated with the CYP1A2 -163C>A polymorphism. Eur. J. Clin. Pharmacol. 2010, 66, 697–703. [Google Scholar] [CrossRef]
- Algrain, H.A.; Thomas, R.M.; Carrillo, A.E.; Ryan, E.J.; Kim, C.H.; Lettan, R.B.; Ryan, E.J. The effects of a polymorphism in the cytochrome P450 CYP1A2 gene on performance enhancement with caffeine in recreational cyclists. J. Caffeine Res. 2016, 6, 34–39. [Google Scholar] [CrossRef]
- Giersch, G.E.; Boyett, J.C.; Hargens, T.A.; Luden, N.D.; Saunders, M.J.; Daley, H.; Hughey, C.A.; El-Sohemy, A.; Womack, C.J. The effect of the CYP1A2-163 C>A polymorphism on caffeine metabolism and subsequent cycling performance. J. Caffeine Adenosine Res. 2018, 8, 65–70. [Google Scholar] [CrossRef]
- Guest, N.; Corey, P.; Vescovi, J.; El-Sohemy, A. Caffeine, CYP1A2 genotype, and endurance performance in athletes. Med. Sci. Sports. Exerc. 2018, 50, 1570–1578. [Google Scholar] [CrossRef]
- Klein, C.S.; Clawson, A.; Martin, M.; Saunders, M.J.; Flohr, J.A.; Bechtel, M.K.; Dunham, W.; Hancock, M.; Womack, C.J. The effect of caffeine on performance in collegiate tennis players. J. Caffeine Res. 2012, 2, 111–116. [Google Scholar] [CrossRef]
- Pataky, M.W.; Womack, C.J.; Saunders, M.J.; Goffe, J.L.; D’lugos, A.C.; El-Sohemy, A.; Lunden, N.D. Caffeine and 3-km cycling performance: Effects of mouth rinsing, genotype, and time of day. Scand. J. Med. Sci. Sports 2016, 26, 613–619. [Google Scholar] [CrossRef]
- Puente, C.; Abián-Vicén, J.; Del Coso, J.; Lara, B.; Salinero, J.J. The CYP1A2-163C > A polymorphism does not alter the effects of caffeine on basketball performance. PLoS ONE 2018, 13, e0195943. [Google Scholar] [CrossRef] [Green Version]
- Salinero, J.J.; Lara, B.; Ruiz-Vicente, D.; Areces, F.; Puente-Torres, C.; Gallo-Salazar, C.; Pascual, T.; Del Coso, J. CYP1A2 genotype variations do not modify the benefits and drawbacks of caffeine during exercise: A pilot study. Nutrients 2017, 9, 269. [Google Scholar] [CrossRef]
- Womack, C.J.; Saunders, M.J.; Bechtel, M.K.; Bolton, D.J.; Martin, M.; Luden, N.D.; Hancock, M. The influence of a CYP1A2 polymorphism on the ergogenic effects of caffeine. J. Int. Soc. Sports Nutr. 2012, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, R. The effect of CYP1A2 genotype on the ergogenic properties of caffeine during resistance exercise: A randomized, double-blind, placebo-controlled, crossover study. Ir. J. Med. Sci. 2019, 188, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Grgic, J. Caffeine and Exercise: What Next? Sports Med. 2019, 49, 1007–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelis, M.C.; El-Sohemy, A.; Campos, H. Genetic polymorphism of the adenosine A2A receptor is associated with habitual caffeine consumption. Am. J. Clin. Nutr. 2007, 86, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loy, B.D.; O’Connor, P.J.; Lindheimer, J.B.; Covert, S.F. Caffeine is ergogenic for adenosine A2A receptor gene (ADORA2A) T allele homozygotes: A pilot study. J. Caffeine Res. 2015, 5, 73–81. [Google Scholar] [CrossRef]
- Grgic, J.; Mikulic, P. Caffeine ingestion acutely enhances muscular strength and power but not muscular endurance in resistance-trained men. Eur. J. Sport Sci. 2017, 17, 1029–1036. [Google Scholar] [CrossRef]
- Grgic, J.; Sabol, F.; Venier, S.; Mikulic, I.; Bratkovic, N.; Schoenfeld, B.J.; Pickering, C.; Bishop, D.J.; Pedisic, Z.; Mikulic, P. What Dose of Caffeine to Use: Acute Effects of 3 Doses of Caffeine on Muscle Endurance and Strength. Int. J. Sports Physiol. Perform. 2019. [Google Scholar] [CrossRef]
- Pallarés, J.G.; Fernández-Elías, V.E.; Ortega, J.F.; Muñoz, G.; Muñoz-Guerra, J.; Mora-Rodríguez, R. Neuromuscular responses to incremental caffeine doses: Performance and side effects. Med. Sci. Sports Exerc. 2013, 45, 2184–2192. [Google Scholar] [CrossRef]
- Venier, S.; Grgic, J.; Mikulic, P. Acute Enhancement of Jump Performance, Muscle Strength, and Power in Resistance-Trained Men After Consumption of Caffeinated Chewing Gum. Int. J. Sports. Physiol. Perform. 2020, 14, 1415–1421. [Google Scholar] [CrossRef]
- Venier, S.; Grgic, J.; Mikulic, P. Caffeinated Gel Ingestion Enhances Jump Performance, Muscle Strength, and Power in Trained Men. Nutrients 2019, 11, 937. [Google Scholar] [CrossRef] [Green Version]
- Frikha, M.; Chaâri, N.; Mezghanni, N.; Souissi, N. Influence of warm-up duration and recovery interval prior to exercise on anaerobic performance. Biol. Sport 2016, 33, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, B.; de Oliveira, L.F.; da Silva, R.P.; de Salles Painelli, V.; Gonçalves, L.S.; Yamaguchi, G.; Mutti, T.; Maciel, E.; Roschel, H.; Artioli, G.G.; et al. Placebo in sports nutrition: A proof-of-principle study involving caffeine supplementation. Scand. J. Med. Sci. Sports 2017, 27, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Kiely, J.; Suraci, B.; Collins, D. The magnitude of Yo-Yo test improvements following an aerobic training intervention are associated with total genotype score. PLoS ONE 2018, 13, e0207597. [Google Scholar] [CrossRef]
- Skinner, T.L.; Desbrow, B.; Arapova, J.; Schaumberg, M.A.; Osborne, J.; Grant, G.D.; Anoopkumar-Dukie, S.; Leveritt, M.D. Women Experience the Same Ergogenic Response to Caffeine as Men. Med. Sci. Sports Exerc. 2019, 51, 1195–1202. [Google Scholar] [CrossRef] [Green Version]
- Sabblah, S.; Dixon, D.; Bottoms, L. Sex differences on the acute effects of caffeine on maximal strength and muscular endurance. Comp. Exerc. Physiol. 2015, 11, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Mielgo-Ayuso, J.; Marques-Jiménez, D.; Refoyo, I.; Del Coso, J.; León-Guereño, P.; Calleja-González, J. Effect of Caffeine Supplementation on Sports Performance Based on Differences Between Sexes: A Systematic Review. Nutrients 2019, 11, 2313. [Google Scholar] [CrossRef] [Green Version]
- Raya-González, J.; Rendo-Urteaga, T.; Domínguez, R.; Castillo, D.; Rodríguez-Fernández, A.; Grgic, J. Acute Effects of Caffeine Supplementation on Movement Velocity in Resistance Exercise: A Systematic Review and Meta-analysis. Sports Med. 2019. [Google Scholar] [CrossRef]
- Burke, L.M. Caffeine and sports performance. Appl. Physiol. Nutr. Metab. 2008, 33, 1319–1334. [Google Scholar] [CrossRef]
- Erblang, M.; Drogou, C.; Gomez-Merino, D.; Metlaine, A.; Boland, A.; Deleuze, J.F.; Thomas, C.; Sauvet, F.; Chennaoui, M. The Impact of Genetic Variations in ADORA2A in the Association between Caffeine Consumption and Sleep. Genes 2019, 10, 1021. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.P.; Stokes, K.A.; Trewartha, G.; Doyle, J.; Hogben, P.; Thompson, D. Effects of carbohydrate and caffeine ingestion on performance during a rugby union simulation protocol. J. Sports Sci. 2010, 28, 833–842. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.E.; Jentjens, R.L.; Wallis, G.A.; Jeukendrup, A.E. Caffeine increases exogenous carbohydrate oxidation during exercise. J. Appl. Physiol. 2005, 99, 844–850. [Google Scholar] [CrossRef] [Green Version]
- Aird, T.P.; Davies, R.W.; Carson, B.P. Effects of fasted vs fed-state exercise on performance and post-exercise metabolism: A systematic review and meta-analysis. Scand. J. Med. Sci. Sports 2018, 28, 1476–1493. [Google Scholar] [CrossRef]
Figure 1.
The effects of caffeine vs. placebo on peak power (upper left section), peak velocity (lower left section), mean power (upper right section), and mean velocity (lower right section) in the bench press with 25%, 50%, 75%, and 90% of one repetition maximum (1RM). Data are presented as mean ± standard deviation. * denotes significant differences between the conditions.
Figure 1.
The effects of caffeine vs. placebo on peak power (upper left section), peak velocity (lower left section), mean power (upper right section), and mean velocity (lower right section) in the bench press with 25%, 50%, 75%, and 90% of one repetition maximum (1RM). Data are presented as mean ± standard deviation. * denotes significant differences between the conditions.


{kind=link}
Table 1.
Characteristics of the participants.
| Variable | Mean ± Standard Deviation |
|---|---|
| Age (years) | 29.3 ± 4.8 |
| Body mass (kg) | 80.3 ± 11.2 |
| Height (cm) | 183.1 ± 5.9 |
| 1RM in the bench press (normalized per body mass) | 1.1 ± 0.2 |
| Habitual caffeine intake (mg/day) | 143 ± 113 |
1RM: one repetition maximum.
Table 2.
Effects of caffeine ingestion on performance in the muscular endurance test, countermovement jump, and Wingate: results from a series of one-way repeated measures analyses of variance.
Table 2.
Effects of caffeine ingestion on performance in the muscular endurance test, countermovement jump, and Wingate: results from a series of one-way repeated measures analyses of variance.
| Variable | Placebo | Caffeine | Hedges’ g and 95% CI | p-Value |
|---|---|---|---|---|
| Muscular endurance test | ||||
| Maximum repetitions at 85% 1RM | 6.9 ± 2.2 | 8.2 ± 2.1 | 0.58 (0.29, 0.91) | <0.001 |
| Mean power matched for repetitions (W) | 418 ± 116 | 492 ± 138 | 0.56 (0.32, 0.83) | <0.001 |
| Mean velocity matched for repetitions (m/s) | 0.27 ± 0.05 | 0.32 ± 0.05 | 0.96 (0.58, 1.41) | <0.001 |
| Peak power matched for repetitions (W) | 669 ± 250 | 740 ± 258 | 0.27 (0.14, 0.42) | <0.001 |
| Peak velocity matched for repetitions (m/s) | 0.41 ± 0.08 | 0.46 ± 0.07 | 0.64 (0.38, 0.94) | <0.001 |
| CMJ | ||||
| Vertical jump height (cm) | 35.0 ± 6.1 | 35.8 ± 5.9 | 0.13 (0.02, 0.25) | 0.034 |
| Wingate test | ||||
| Peak power in the Wingate test (W) | 859 ± 237 | 948 ± 229 | 0.37 (0.21, 0.55) | <0.001 |
| Mean power in the Wingate test (W) | 598 ± 101 | 634 ± 100 | 0.34 (0.17, 0.54) | <0.001 |
| Minimum power in the Wingate test (W) | 349 ± 103 | 392 ± 96 | 0.41 (0.07, 0.78) | 0.020 |
1RM: one repetition maximum: CMJ: countermovement jump; CI: confidence interval.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grgic, J.; Pickering, C.; Bishop, D.J.; Del Coso, J.; Schoenfeld, B.J.; Tinsley, G.M.; Pedisic, Z. ADORA2A C Allele Carriers Exhibit Ergogenic Responses to Caffeine Supplementation. Nutrients 2020, 12, 741. https://doi.org/10.3390/nu12030741
AMA Style
Grgic J, Pickering C, Bishop DJ, Del Coso J, Schoenfeld BJ, Tinsley GM, Pedisic Z. ADORA2A C Allele Carriers Exhibit Ergogenic Responses to Caffeine Supplementation. Nutrients. 2020; 12(3):741. https://doi.org/10.3390/nu12030741
Chicago/Turabian StyleGrgic, Jozo, Craig Pickering, David J. Bishop, Juan Del Coso, Brad J. Schoenfeld, Grant M. Tinsley, and Zeljko Pedisic. 2020. "ADORA2A C Allele Carriers Exhibit Ergogenic Responses to Caffeine Supplementation" Nutrients 12, no. 3: 741. https://doi.org/10.3390/nu12030741
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.