Specific Wheat Fractions Influence Hepatic Fat Metabolism in Diet-Induced Obese Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Diets
2.2. Gene Expression
2.3. Quantification of Plasma Lipid Species
2.4. Analysis of Liver Lipids
2.4.1. Liver Triglyceride Analysis
2.4.2. Long-Chain Fatty Acid Analysis
2.5. Microbiota
2.5.1. DNA Extraction
2.5.2. 16S Amplicon Sequencing
2.5.3. Bioinformatic Analysis of Microbiome Data
2.5.4. Data Availability
2.6. SCFA Analysis
2.7. Histology
2.8. Statistical Analysis
3. Results
3.1. Impact of Different Wheat Fractions on Food Intake, Body Weight, and Organ Weights
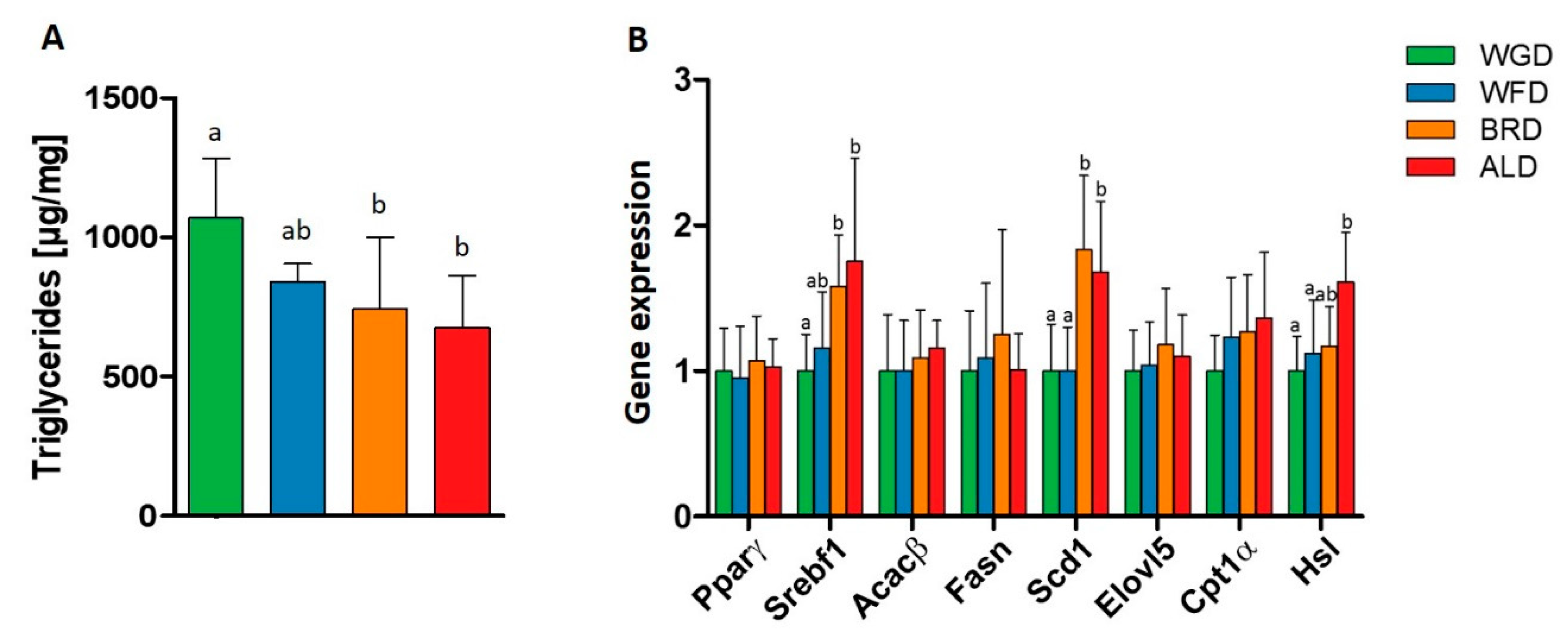
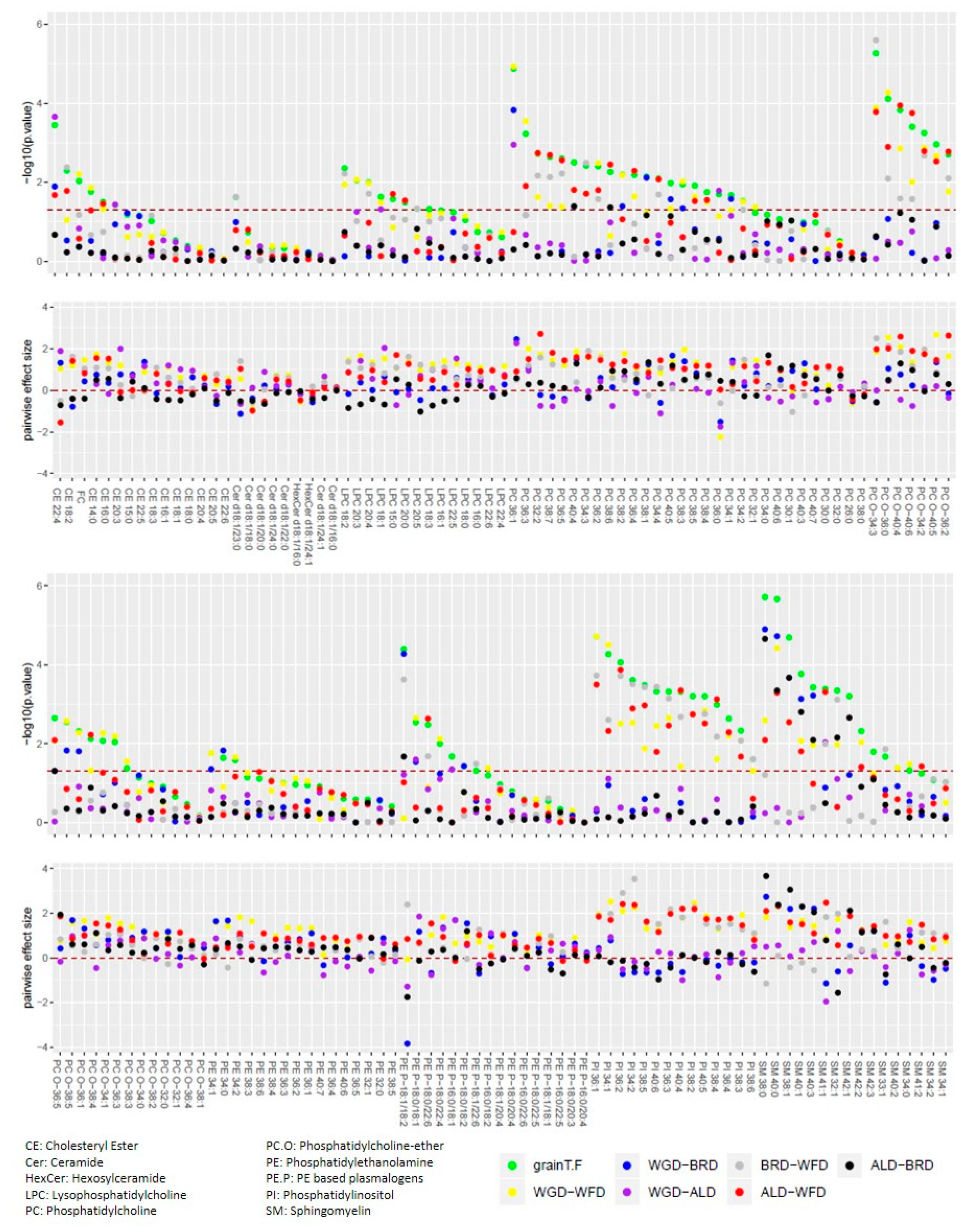
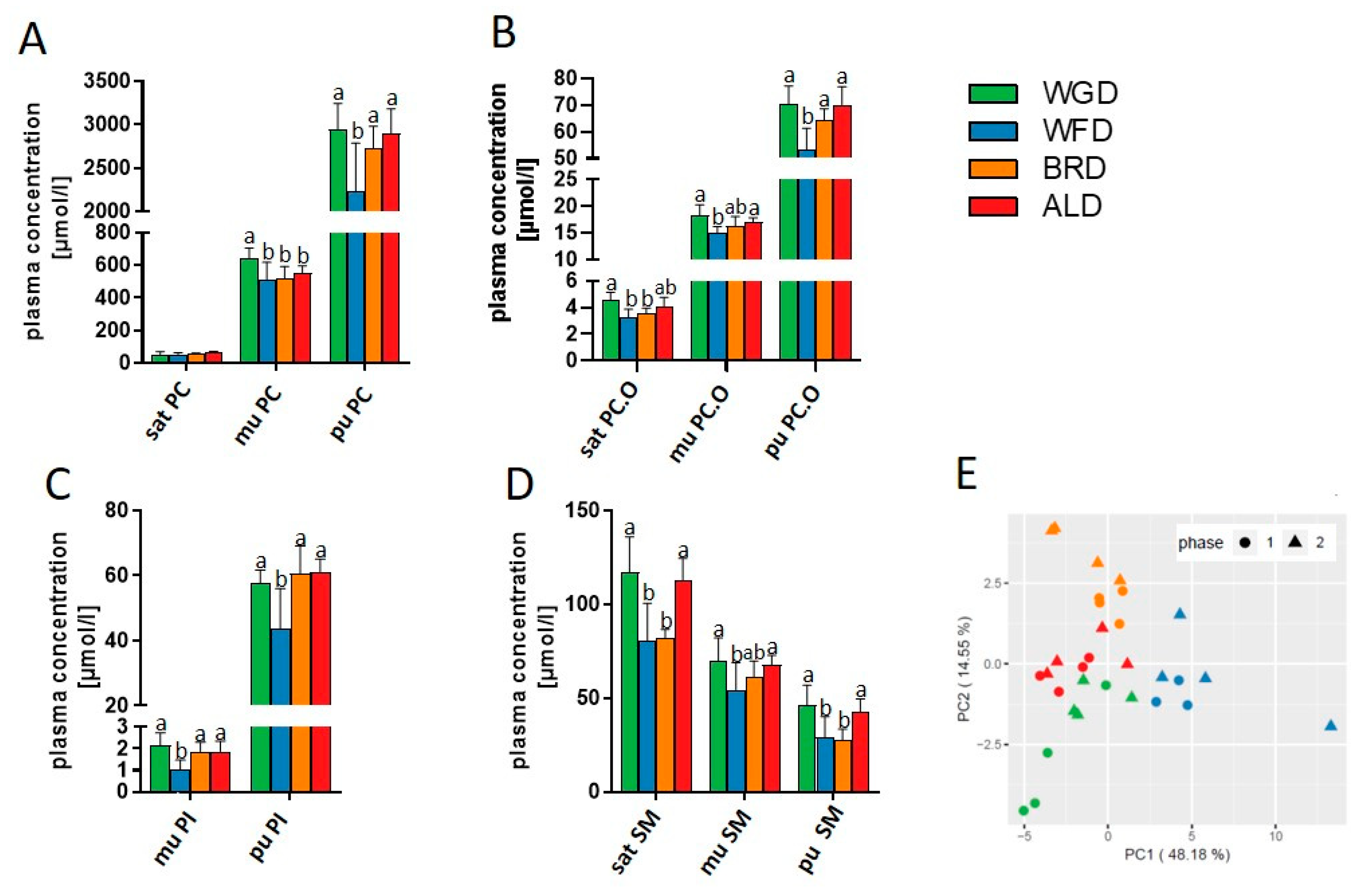
3.2. Lipid Metabolism
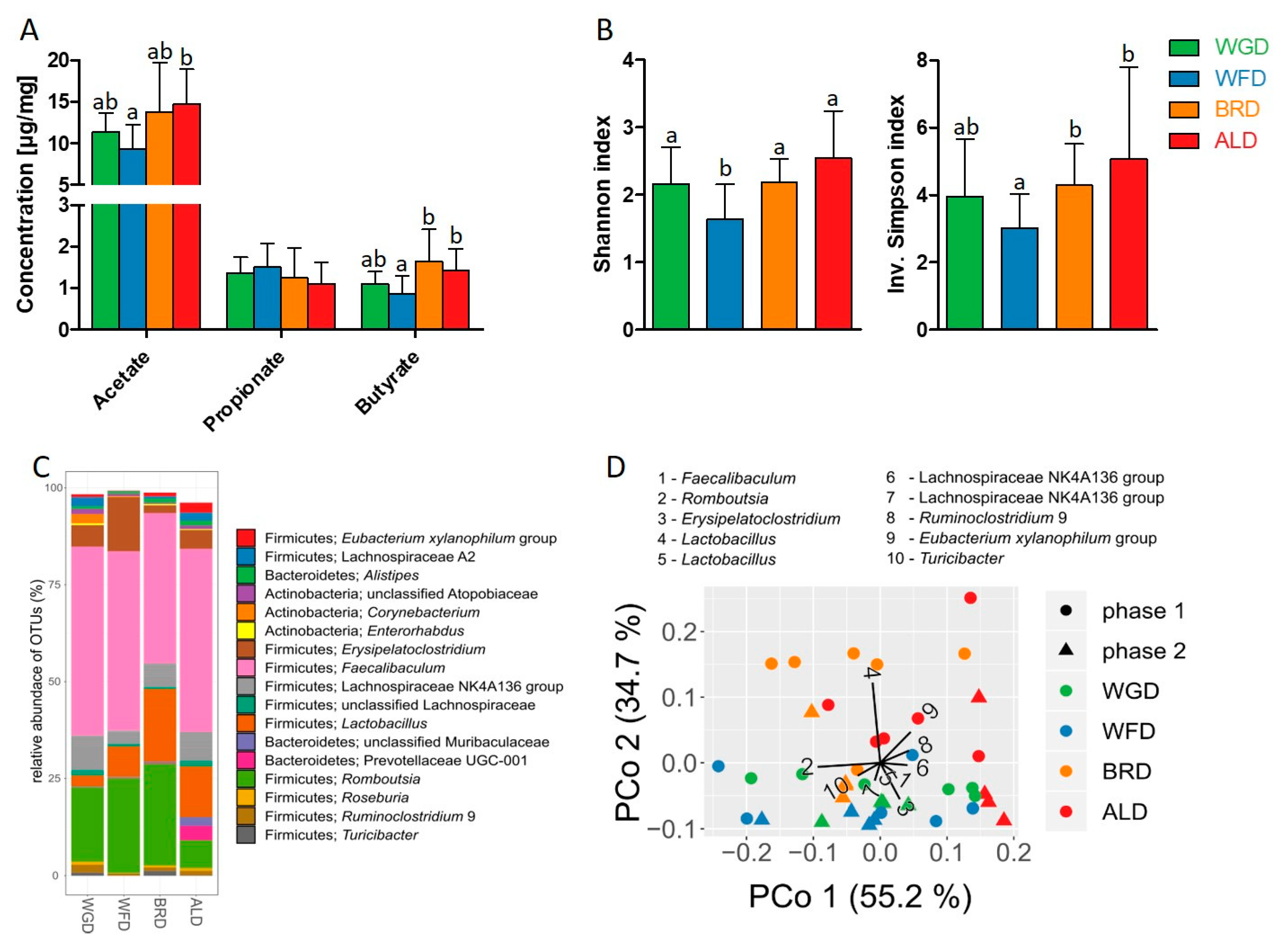
3.3. SCFA Production and Microbiota Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAC-β | Acetyl-coenzyme A carboxylase beta |
| ALD | Aleurone supplemented diet |
| BRD | Bran supplemented diet |
| CPT1α | Carnitine palmitoyltransferase 1 alpha |
| DIO | Diet-induced obesity |
| ELOVL5 | Fatty acid elongase 5 |
| FASN | Fatty acid synthase |
| FID | flame ionization detector |
| HFD | high fat diet |
| HSL | Hormone-sensitive lipase |
| MUFA | Monounsaturated fatty acids |
| NAFLD | Non-alcoholic fatty liver disease |
| PUFA | Polyunsaturated fatty acids |
| SCFA | Short-chain fatty acids |
| SCD1 | Stearoyl-CoA desaturase 1 |
| SFA | Saturated fatty acids |
| SREBF-1 | Sterol regulatory element-binding transcription factor 1 |
| WFD | White wheat flour supplemented diet |
| WGD | Whole grain wheat supplemented diet |
References
- WHO. Fact Sheet: Obesity and Overweight. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 16 May 2019).
- Milic, S.; Lulic, D.; Stimac, D. Non-alcoholic fatty liver disease and obesity: Biochemical, metabolic and clinical presentations. World J. Gastroenterol. 2014, 20, 9330–9337. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- WHO. Fact Sheet: The Top 10 Causes of Death. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 16 May 2019).
- GBD 2017 Diet Collaborators. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef]
- Public Health England. National Diet and Nutrition Survey Results from Years 1, 2, 3 and 4 (Combined) of the Rolling Programme (2008/2009–2011/2012). 2014. Available online: https://www.gov.uk/government/statistics/national-diet-and-nutrition-survey-results-from-years-1-to-4-combined-of-the-rolling-programme-for-2008-and-2009-to-2011-and-2012 (accessed on 20 May 2019).
- Benisi-Kohansal, S.; Saneei, P.; Salehi-Marzijarani, M.; Larijani, B.; Esmaillzadeh, A. Whole-Grain Intake and Mortality from All Causes, Cardiovascular Disease, and Cancer: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Adv. Nutr. (Bethesda Md.) 2016, 7, 1052–1065. [Google Scholar] [CrossRef] [Green Version]
- Aune, D.; Chan, D.S.; Lau, R.; Vieira, R.; Greenwood, D.C.; Kampman, E.; Norat, T. Dietary fibre, whole grains, and risk of colorectal cancer: Systematic review and dose-response meta-analysis of prospective studies. BMJ (Clin. Res. Ed.) 2011, 343, d6617. [Google Scholar] [CrossRef]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Whole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: Systematic review and dose-response meta-analysis of prospective studies. BMJ (Clin. Res. Ed.) 2016, 353, i2716. [Google Scholar] [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Whole grain and refined grain consumption and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Eur. J. Epidemiol. 2013, 28, 845–858. [Google Scholar] [CrossRef]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Koistinen, V.M.; Karkkainen, O.; Borewicz, K.; Zarei, I.; Jokkala, J.; Micard, V.; Rosa-Sibakov, N.; Auriola, S.; Aura, A.M.; Smidt, H.; et al. Contribution of gut microbiota to metabolism of dietary glycine betaine in mice and in vitro colonic fermentation. Microbiome 2019, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finelli, C.; Tarantino, G. Is there any consensus as to what diet or lifestyle approach is the right one for NAFLD patients? J. Gastrointest. Liver Dis. 2012, 21, 293–302. [Google Scholar]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat aleurone: Separation, composition, health aspects, and potential food use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani, A.I.O.; Beswa, D. Composition and functionality of wheat bran and its application in some cereal food products. Int. J. Food Sci. Technol. 2015, 50, 2509–2518. [Google Scholar] [CrossRef]
- Buri, R.C.; von Reding, W.; Gavin, M.H. Description and Characterization of Wheat Aleurone. Cereal Foods World 2004, 49, 274–282. [Google Scholar]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Weitkunat, K.; Schumann, S.; Nickel, D.; Kappo, K.A.; Petzke, K.J.; Kipp, A.P.; Blaut, M.; Klaus, S. Importance of propionate for the repression of hepatic lipogenesis and improvement of insulin sensitivity in high-fat diet-induced obesity. Mol. Nutr. Food Res. 2016, 60, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Binder, M.; Schifferer, R.; Langmann, T.; Schulz, B.; Schmitz, G. High throughput quantification of cholesterol and cholesteryl ester by electrospray ionization tandem mass spectrometry (ESI-MS/MS). Biochim. Biophys. Acta 2006, 1761, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Lieser, B.; Rathenberg, J.; Drobnik, W.; Schmitz, G. High-throughput quantification of phosphatidylcholine and sphingomyelin by electrospray ionization tandem mass spectrometry coupled with isotope correction algorithm. Biochim. Biophys. Acta 2004, 1686, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Liebisch, G.; Drobnik, W.; Lieser, B.; Schmitz, G. High-throughput quantification of lysophosphatidylcholine by electrospray ionization tandem mass spectrometry. Clin. Chem. 2002, 48, 2217–2224. [Google Scholar]
- Brugger, B.; Erben, G.; Sandhoff, R.; Wieland, F.T.; Lehmann, W.D. Quantitative analysis of biological membrane lipids at the low picomole level by nano-electrospray ionization tandem mass spectrometry. Proc. Natl. Acad. Sci. USA 1997, 94, 2339–2344. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Zemski Berry, K.A.; Murphy, R.C. Electrospray ionization tandem mass spectrometry of glycerophosphoethanolamine plasmalogen phospholipids. J. Am. Soc. Mass Spectrom. 2004, 15, 1499–1508. [Google Scholar] [CrossRef] [Green Version]
- Liebisch, G.; Drobnik, W.; Reil, M.; Trumbach, B.; Arnecke, R.; Olgemoller, B.; Roscher, A.; Schmitz, G. Quantitative measurement of different ceramide species from crude cellular extracts by electrospray ionization tandem mass spectrometry (ESI-MS/MS). J. Lipid Res. 1999, 40, 1539–1546. [Google Scholar]
- Scherer, M.; Schmitz, G.; Liebisch, G. Simultaneous quantification of cardiolipin, bis(monoacylglycero)phosphate and their precursors by hydrophilic interaction LC-MS/MS including correction of isotopic overlap. Anal. Chem. 2010, 82, 8794–8799. [Google Scholar] [CrossRef]
- Liebisch, G.; Vizcaino, J.A.; Kofeler, H.; Trotzmuller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitkunat, K.; Schumann, S.; Petzke, K.J.; Blaut, M.; Loh, G.; Klaus, S. Effects of dietary inulin on bacterial growth, short-chain fatty acid production and hepatic lipid metabolism in gnotobiotic mice. J. Nutr. Biochem. 2015, 26, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Camarinha-Silva, A.; Jauregui, R.; Chaves-Moreno, D.; Oxley, A.P.; Schaumburg, F.; Becker, K.; Wos-Oxley, M.L.; Pieper, D.H. Comparing the anterior nare bacterial community of two discrete human populations using Illumina amplicon sequencing. Environ. Microbiol. 2014, 16, 2939–2952. [Google Scholar] [CrossRef]
- Etchebehere, C.; Tiedje, J. Presence of two different active nirS nitrite reductase genes in a denitrifying Thauera sp. from a high-nitrate-removal-rate reactor. Appl. Environ. Microbiol. 2005, 71, 5642–5645. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Schloss, P.D.; Baxter, N.T.; Jenior, M.L.; Koumpouras, C.C. MiSeq Wet Lab SOP. Available online: https://github.com/SchlossLab/MiSeq_WetLab_SOP/blob/master/MiSeq_WetLab_SOP_v4.md (accessed on 17 May 2018).
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package (Version 2.5-1). Available online: https://CRAN.R-project.org/package=vegan (accessed on 14 April 2018).
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.0.1. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 23 August 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Luepsen, H. Comparison of nonparametric analysis of variance methods: A vote for van der Waerden. Commun. Stat. Simul. Comput. 2018, 47, 2547–2576. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 15 July 2018).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biosciences 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; Lange, K.; Havinga, R.; van Dijk, T.H.; Gerding, A.; van, E.K.; Muller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Phy. Gastrointest. Liver Physiol. 2013, 305, G900–G910. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol. (Hoboken NJ) 2015, 67, 128–139. [Google Scholar] [CrossRef]
- Ott, S.J.; Musfeldt, M.; Wenderoth, D.F.; Hampe, J.; Brant, O.; Fölsch, U.R.; Timmis, K.N.; Schreiber, S. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 2004, 53, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Dobbler, P.T.; Procianoy, R.S.; Mai, V.; Silveira, R.C.; Corso, A.L.; Rojas, B.S.; Roesch, L.F.W. Low Microbial Diversity and Abnormal Microbial Succession Is Associated with Necrotizing Enterocolitis in Preterm Infants. Front. Microbiol. 2017, 8, 2243. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Stefan, N.; Häring, H.-U. The Metabolically Benign and Malignant Fatty Liver. Diabetes 2011, 60, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.J.; Sul, H.S. Insulin regulation of fatty acid synthase gene transcription: Roles of USF and SREBP-1c. IUBMB Life 2004, 56, 595–600. [Google Scholar] [CrossRef]
- Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Hasty, A.H.; Osuga, J.; Tamura, Y.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; et al. Sterol regulatory element-binding protein-1 as a key transcription factor for nutritional induction of lipogenic enzyme genes. J. Biol. Chem. 1999, 274, 35832–35839. [Google Scholar] [CrossRef]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Keski-Rahkonen, P.; Kolehmainen, M.; Lappi, J.; Micard, V.; Jokkala, J.; Rosa-Sibakov, N.; Pihlajamaki, J.; Kirjavainen, P.V.; Mykkanen, H.; Poutanen, K.; et al. Decreased plasma serotonin and other metabolite changes in healthy adults after consumption of wholegrain rye: An untargeted metabolomics study. Am. J. Clin. Nutr. 2019, 109, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Kindt, A.; Liebisch, G.; Clavel, T.; Haller, D.; Hormannsperger, G.; Yoon, H.; Kolmeder, D.; Sigruener, A.; Krautbauer, S.; Seeliger, C.; et al. The gut microbiota promotes hepatic fatty acid desaturation and elongation in mice. Nat. Commun. 2018, 9, 3760. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Arora, T.; Bäckhed, F. The gut microbiota and metabolic disease: Current understanding and future perspectives. J. Intern. Med. 2016, 280, 339–349. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, E.D.; Sonnenburg, J.L. Starving our microbial self: The deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab. 2014, 20, 779–786. [Google Scholar] [CrossRef]
- Scharlau, D.; Borowicki, A.; Habermann, N.; Hofmann, T.; Klenow, S.; Miene, C.; Munjal, U.; Stein, K.; Glei, M. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res./Rev. Mutat. Res. 2009, 682, 39–53. [Google Scholar] [CrossRef]
- Ounnas, F.; Salen, P.; Demeilliers, C.; Calani, L.; Scazzina, F.; Hazane-Puch, F.; Laporte, F.; Melegari, C.; Del Rio, D.; de Lorgeril, M. Wheat aleurone fractions and plasma n-3 fatty acids in rats. Int. J. Food Sci. Nutr. 2015, 66, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Toufektsian, M.C.; Salen, P.; Laporte, F.; Tonelli, C.; de, L.M. Dietary flavonoids increase plasma very long-chain (n-3) fatty acids in rats. J. Nutr. 2011, 141, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Seifert, S.; Jaudszus, A.; Bub, A.; Watzl, B. Anthocyanin-Rich Juice Lowers Serum Cholesterol, Leptin, and Resistin and Improves Plasma Fatty Acid Composition in Fischer Rats. PLoS ONE 2013, 8, e66690. [Google Scholar] [CrossRef] [PubMed]
- di Giuseppe, R.; de Lorgeril, M.; Salen, P.; Laporte, F.; Di Castelnuovo, A.; Krogh, V.; Siani, A.; Arnout, J.; Cappuccio, F.P.; van Dongen, M.; et al. Alcohol consumption and n-3 polyunsaturated fatty acids in healthy men and women from 3 European populations. Am. J. Clin. Nutr. 2009, 89, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Rosa, N.N.; Pekkinen, J.; Zavala, K.; Fouret, G.; Korkmaz, A.; Feillet-Coudray, C.; Atalay, M.; Hanhineva, K.; Mykkanen, H.; Poutanen, K.; et al. Impact of wheat aleurone structure on metabolic disorders caused by a high-fat diet in mice. J. Agric. Food Chem. 2014, 62, 10101–10109. [Google Scholar] [CrossRef]
- Pol, K.; Christensen, R.; Bartels, E.M.; Raben, A.; Tetens, I.; Kristensen, M. Whole grain and body weight changes in apparently healthy adults: A systematic review and meta-analysis of randomized controlled studies. Am. J. Clin. Nutr. 2013, 98, 872–884. [Google Scholar] [CrossRef]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.H.; Lai, C.S.; Ho, C.T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; De Backer, F.; Cani, P.D.; Bindels, L.B.; Stroobants, A.; Portetelle, D.; Delzenne, N.M. Immunomodulatory properties of two wheat bran fractions-aleurone-enriched and crude fractions—In obese mice fed a high fat diet. Int. Immunopharmacol. 2008, 8, 1423–1432. [Google Scholar] [CrossRef]
- Schutte, S.; Esser, D.; Hoevenaars, F.P.M.; Hooiveld, G.; Priebe, M.G.; Vonk, R.J.; Wopereis, S.; Afman, L.A. A 12-wk whole-grain wheat intervention protects against hepatic fat: The Graandioos study, a randomized trial in overweight subjects. Am. J. Clin. Nutr. 2018, 108, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Nozaki, S.; Makita, M.; Yokozuka, S.; Fukudome, S.I.; Yanagisawa, T.; Aoe, S. Effects of Whole Grain Wheat Bread on Visceral Fat Obesity in Japanese Subjects: A Randomized Double-Blind Study. Plant Foods Hum. Nutr. 2018, 73, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhanel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M.; et al. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef] [PubMed]
- Heyman-Linden, L.; Kotowska, D.; Sand, E.; Bjursell, M.; Plaza, M.; Turner, C.; Holm, C.; Fak, F.; Berger, K. Lingonberries alter the gut microbiota and prevent low-grade inflammation in high-fat diet fed mice. Food Nutr. Res. 2016, 60, 29993. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schurmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Cani, P.D.; Delzenne, N.M.; Amar, J.; Burcelin, R. Role of gut microflora in the development of obesity and insulin resistance following high-fat diet feeding. Pathol. Biol. 2008, 56, 305–309. [Google Scholar] [CrossRef]
- Gose, M.; Krems, C.; Heuer, T.; Hoffmann, I. Trends in food consumption and nutrient intake in Germany between 2006 and 2012: Results of the German National Nutrition Monitoring (NEMONIT). Br. J. Nutr. 2016, 115, 1498–1507. [Google Scholar] [CrossRef]
- Isken, F.; Klaus, S.; Osterhoff, M.; Pfeiffer, A.F.; Weickert, M.O. Effects of long-term soluble vs. insoluble dietary fiber intake on high-fat diet-induced obesity in C57BL/6J mice. J. Nutr. Biochem. 2010, 21, 278–284. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WGD | WFD | BRD | ALD | |
|---|---|---|---|---|
| Casein | 200 | 200 | 200 | 200 |
| Cornstarch | 395 | 395 | 395 | 395 |
| Sucrose | 50 | 50 | 50 | 50 |
| Lard | 190 | 190 | 190 | 190 |
| Whole grain flour | 100 | |||
| White flour | 100 | |||
| Bran | 100 | |||
| Aleurone | 100 | |||
| Vitamin mix | 10 | 10 | 10 | 10 |
| Mineral mix | 50 | 50 | 50 | 50 |
| L-Cysteine | 3 | 3 | 3 | 3 |
| Choline chloride | 3 | 3 | 3 | 3 |
| WGD | WFD | BRD | ALD | |
|---|---|---|---|---|
| Food intake [g/day] | 2.9 ± 0.1 | 2.9 ± 0.2 | 3.2 ± 0.4 | 3.0 ± 0.2 |
| Body weight [g] | 35.7 ± 3.3 | 36.0 ± 3.2 | 36.8 ± 4.1 | 36.2 ± 3.2 |
| Liver [g] | 1.6 ± 0.2 | 1.6 ± 0.2 | 1.5 ± 0.2 | 1.5 ± 0.2 |
| EAT [g] | 1.8 ± 0.3 | 1.9 ± 0.4 | 1.9 ± 0.5 | 1.9 ± 0.5 |
| SAT [g] | 1.8 ± 0.6 | 1.9 ± 0.8 | 2.4 ± 0.8 | 2.0 ± 0.5 |
| VAT [g] | 0.5 ± 0.2 | 0.6 ± 0.1 | 0.6 ± 0.2 | 0.5 ± 0.2 |
| Adipocyte density [cell/mm2] | 0.48 ± 0.11 | 0.51 ± 0.11 | 0.47 ± 0.10 | 0.46 ± 0.09 |
| WGD | WFD | BRD | ALD | |
|---|---|---|---|---|
| C14:0 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.02 | 0.10 ± 0.01 |
| C15:0 | 0.05 ± 0.01 | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.01 |
| C16:0 | 23.05 ± 0.49 | 23.25 ± 0.49 | 22.65 ± 0.64 | 22.54 ± 0.76 |
| C16:1n7c | 0.97 ± 0.11 | 0.98 ± 0.12 | 1.00 ± 0.16 | 0.88 ± 0.04 |
| C17:0 | 0.21 ± 0.02 | 0.21 ± 0.02 | 0.23 ± 0.03 | 0.24 ± 0.02 |
| C18:0 | 16.68 ± 0.55 | 16.44 ± 0.55 | 16.63 ± 0.61 | 17.04 ± 0.47 |
| C18:1n9c | 12.33 ± 0.57 a | 12.71 ± 1.42 a | 11.47 ± 0.84 ab | 10.90 ± 1.01 b |
| C18:1n7c | 2.42 ± 0.27 | 2.35 ± 0.34 | 2.42 ± 0.40 | 2.30 ± 0.38 |
| C18:2n6t | 0.09 ± 0.01 ab | 0.08 ± 0.01 a | 0.09 ± 0.02 ab | 0.09 ± 0.01 b |
| C18:2n6c | 7.54 ± 0.70 | 7.44 ± 0.84 | 8.24 ± 0.70 | 7.83 ± 0.94 |
| C18:3n6 | 0.15 ± 0.03 | 0.15 ± 0.02 | 0.15 ± 0.01 | 0.15 ± 0.02 |
| C18:3n3 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.00 |
| C20:0 | 0.19 ± 0.04 | 0.16 ± 0.04 | 0.19 ± 0.02 | 0.20 ± 0.06 |
| C20:1n9 | 0.31 ± 0.02 | 0.30 ± 0.04 | 0.30 ± 0.03 | 0.29 ± 0.03 |
| C20:2n6 | 0.23 ± 0.02 | 0.21 ± 0.03 | 0.24 ± 0.02 | 0.23 ± 0.02 |
| C20:3n6 | 2.47 ± 0.18 | 2.24 ± 0.23 | 2.42 ± 0.23 | 2.29 ± 0.21 |
| C20:4n6 | 21.61 ± 1.14 | 21.42 ± 0.85 | 21.58 ± 0.82 | 22.22 ± 0.43 |
| C20:5n3 | 0.11 ± 0.02 | 0.11 ± 0.03 | 0.13 ± 0.02 | 0.11 ± 0.02 |
| C22:4n6 | 0.38 ± 0.02 a | 0.39 ± 0.03 a | 0.36 ± 0.02 b | 0.38 ± 0.01 ab |
| C22:5n6 | 1.67 ± 0.10 a | 1.57 ± 0.14 ab | 1.36 ± 0.10 c | 1.42 ± 0.15 bc |
| C22:5n3 | 0.45 ± 0.01 | 0.46 ± 0.04 | 0.45 ± 0.02 | 0.47 ± 0.02 |
| C22:6n3 | 8.96 ± 0.72 a | 9.34 ± 0.37 a | 9.87 ± 0.61 b | 10.25 ± 0.51 b |
| ∑ n3 FA | 9.54 ± 0.71 a | 9.95 ± 0.34 a | 10.49 ± 0.62 b | 10.86 ± 0.51 b |
| ∑ n6 FA | 34.14 ± 0.75 | 33.51 ± 1.13 | 34.45 ± 1.01 | 34.61 ± 1.08 |
| n6/n3 FA | 3.59 ± 0.27 a | 3.37 ± 0.15 ab | 3.29 ± 0.17 b | 3.19 ± 0.22 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graf, D.; Weitkunat, K.; Dötsch, A.; Liebisch, G.; Döring, M.; Krüger, R.; Stoll, D.; Vatareck, E.; von Coburg, E.; Loh, G.; et al. Specific Wheat Fractions Influence Hepatic Fat Metabolism in Diet-Induced Obese Mice. Nutrients 2019, 11, 2348. https://doi.org/10.3390/nu11102348
Graf D, Weitkunat K, Dötsch A, Liebisch G, Döring M, Krüger R, Stoll D, Vatareck E, von Coburg E, Loh G, et al. Specific Wheat Fractions Influence Hepatic Fat Metabolism in Diet-Induced Obese Mice. Nutrients. 2019; 11(10):2348. https://doi.org/10.3390/nu11102348
Chicago/Turabian StyleGraf, Daniela, Karolin Weitkunat, Andreas Dötsch, Gerhard Liebisch, Maik Döring, Ralf Krüger, Dominic Stoll, Elisabeth Vatareck, Elena von Coburg, Gunnar Loh, and et al. 2019. "Specific Wheat Fractions Influence Hepatic Fat Metabolism in Diet-Induced Obese Mice" Nutrients 11, no. 10: 2348. https://doi.org/10.3390/nu11102348