Metabolomic Evaluation of Scenedesmus sp. as a Feed Ingredient Revealed Dose-Dependent Effects on Redox Balance, Intermediary and Microbial Metabolism in a Mouse Model

 ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Culture of Scenedesmus Algae
2.2. Chemicals
2.3. Animal Treatment and Sample Collection
2.4. Growth Performance Measurement
2.5. Serum Biochemical Analysis
2.6. Synthesis of Glycol-4-Methyl Pentanoic Acid and Glycol-4-Methyl Hexanoic Acid
2.7. Sample Preparation for LC-MS Analysis
2.8. Chemical Derivatization
2.9. LC-MS Analysis
2.10. Multivariate Data Analysis
2.11. Marker Characterization and Quantification
2.12. Gene Expression Analysis
2.13. Statistical Analysis
3. Results
3.1. Growth Responses to Scenedesmus Feeding
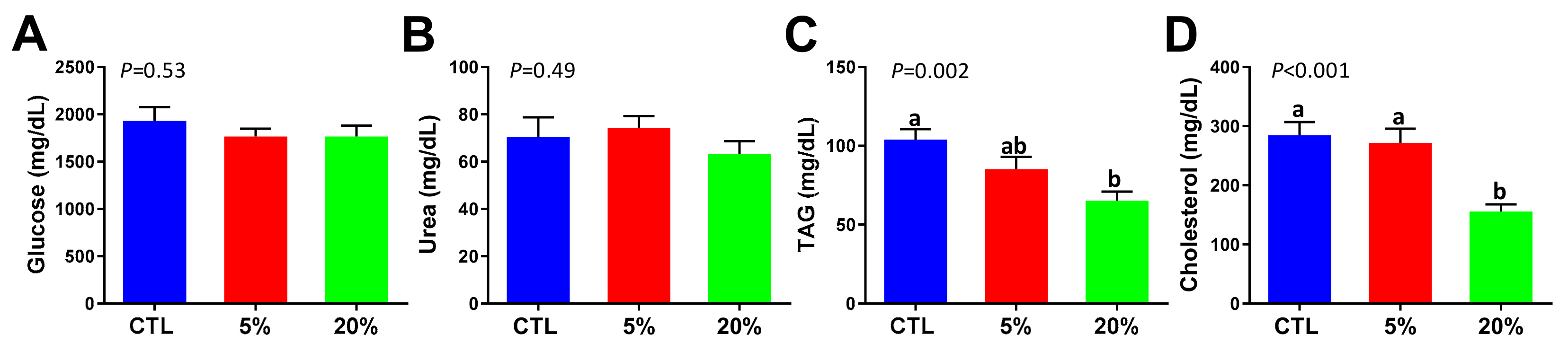
3.2. Effect of Scenedesmus Feeding on Blood Chemistry
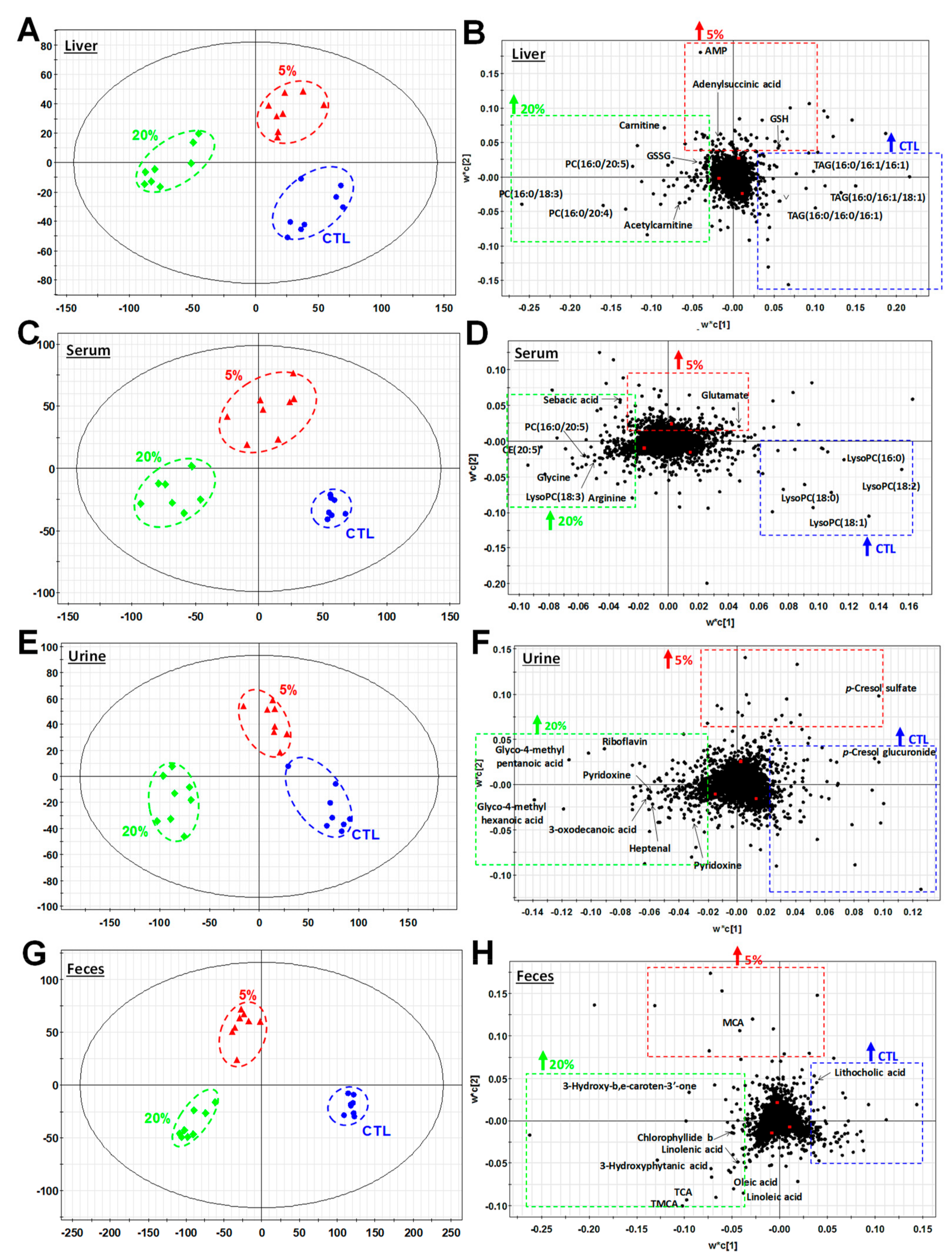
3.3. Metabolomic Investigation of Scenedesmus-Induced Metabolic Events
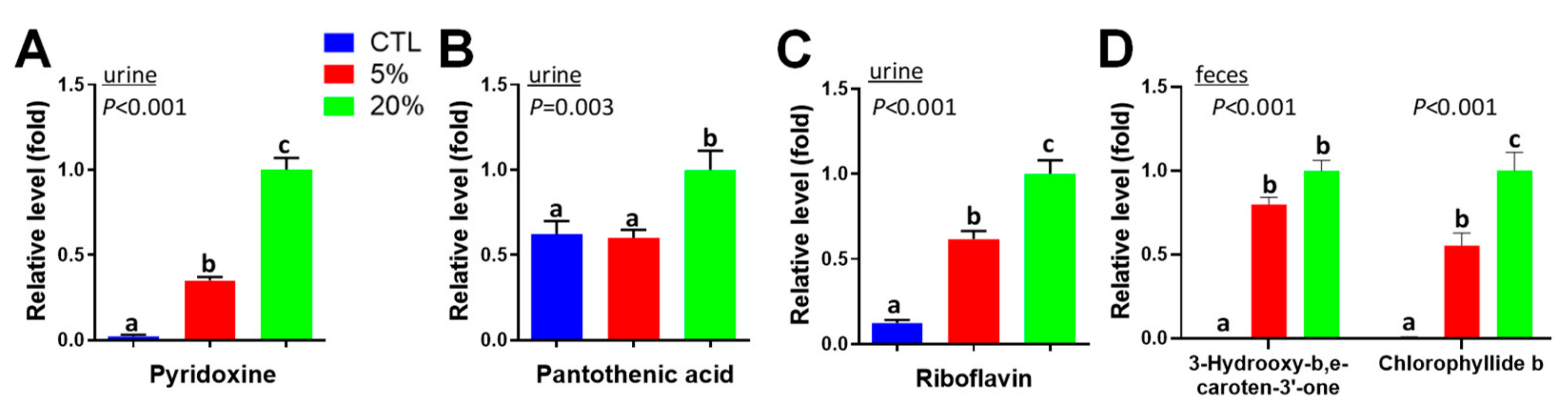
3.4. Scenedesmus Exposure Markers
3.5. Effects of Scenedesmus Feeding on Amino Acid Homeostasis
3.6. Effects of Scenedesmus Feeding on Purine Metabolism
3.7. Effects of Scenedesmus Feeding on the Lipidome
3.8. Effects of Scenedesmus Feeding on Redox Balance
3.9. Effects of Scenedesmus Feeding on Microbial Metabolism
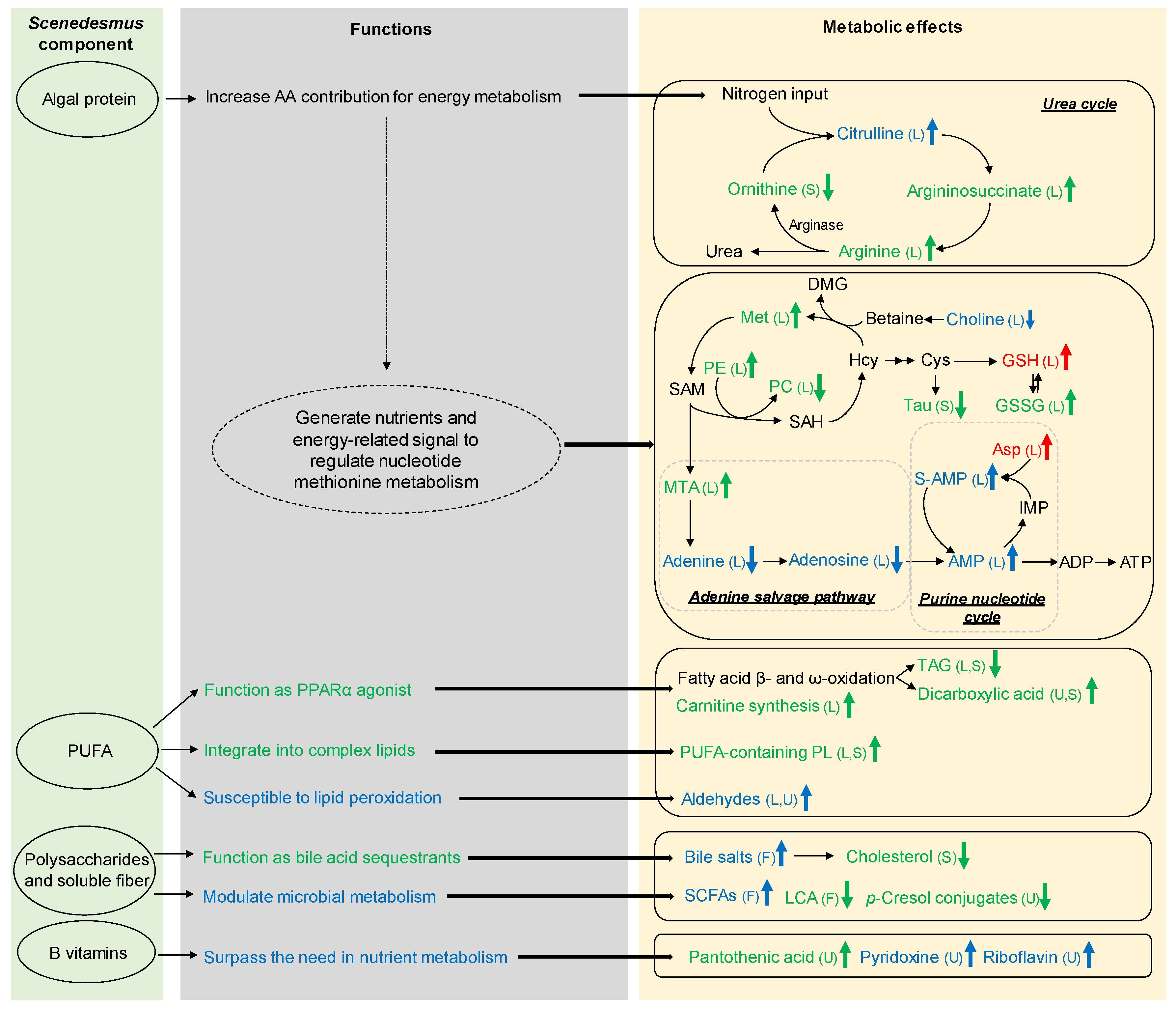
4. Discussion
4.1. Causes and Significance of Scenedesmus-Induced Changes in Vitamins
4.2. Causes and Significance of Scenedesmus-Induced Changes in Nitrogen Metabolism
4.3. Causes and Significance of Scenedesmus-Induced Changes in Purine Nucleotide Metabolism
4.4. Causes and Significance of Scenedesmus-Induced Changes in the Lipidome and Lipid Metabolism
4.5. Causes and Significance of Scenedesmus-Induced Changes in Redox Balance
4.6. Causes and Significance of Scenedesmus-Induced Changes in Microbial Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andersen, R.A. Algal Culturing Techniques; Elsevier Science: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Belasco, W. Algae Burgers for a Hungry World? The Rise and Fall of Chlorella Cuisine. Technol. Cult. 1997, 38, 608–634. [Google Scholar] [CrossRef]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Magnusson, M.; Ward, L.; Paul, N.; Brown, L. A green algae mixture of Scenedesmus and Schroederiella attenuates obesity-linked metabolic syndrome in rats. Nutrients 2015, 7, 2771–2787. [Google Scholar] [CrossRef] [PubMed]
- Rolle, I.; Pabst, W. The cholesterol-lowering effect of the unicellular green alga Scenedesmus acutus 276-3a. I. Effect of drum-dried algal material. Nutr. Metab. 1980, 24, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef]
- Trainor, R.F.; Cain, J.R.; Shubert, L.E. Morphology and nutrition of the colonial green algaScenedesmus: 80 years later. Bot. Rev. 1976, 42, 5–25. [Google Scholar] [CrossRef]
- Song, M.; Pei, H.; Hu, W.; Zhang, S.; Ma, G.; Han, L.; Ji, Y. Identification and characterization of a freshwater microalga Scenedesmus SDEC-8 for nutrient removal and biodiesel production. Bioresour. Technol. 2014, 162, 129–135. [Google Scholar] [CrossRef]
- Mandal, S.; Mallick, N. Microalga Scenedesmus obliquus as a potential source for biodiesel production. Appl. Microbiol. Biotechnol. 2009, 84, 281–291. [Google Scholar] [CrossRef]
- Samuels, R.; Mani, U.V.; Iyer, U.M.; Nayak, U.S. Hypocholesterolemic effect of spirulina in patients with hyperlipidemic nephrotic syndrome. J. Med. Food 2002, 5, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, Y.; Min, M.; Hu, B.; Chen, P.; Ruan, R. Local bioprospecting for high-lipid producing microalgal strains to be grown on concentrated municipal wastewater for biofuel production. Bioresour. Technol. 2011, 102, 6909–6919. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Min, M.; Zhou, W.; Li, Y.; Mohr, M.; Cheng, Y.; Lei, H.; Liu, Y.; Lin, X.; Ruan, R.; et al. Influence of exogenous CO2 on biomass and lipid accumulation of microalgae Auxenochlorella protothecoides cultivated in concentrated municipal wastewater. Appl. Biochem. Biotechnol. 2012, 166, 1661–1673. [Google Scholar] [CrossRef] [PubMed]
- Bligh, G.E.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, D.; Chen, C. 2-Hydrazinoquinoline as a Derivatization Agent for LC-MS-Based Metabolomic Investigation of Diabetic Ketoacidosis. Metabolites 2013, 3, 993–1010. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Laboratory Animals: 1995, 4th ed.; National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- Maschek, J.A.; Baker, J.B. Baker the Chemistry of Algal Secondary Metabolism. In Algal Chemical Ecology; Amsler, C.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–24. [Google Scholar]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar]
- Croft, T.M.; Warren, M.J.; Smith, A.G. Algae need their vitamins. Eukaryot. Cell. 2006, 5, 1175–1183. [Google Scholar] [CrossRef]
- Pratt, R.; Johnson, E. Production of pantothenic acid and inositol by Chlorella vulgaris and C. pyrenoidosa. J. Pharm. Sci. 1966, 55, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, R. Multivitamins: Review. SA Pharm. Apos’s Assist. 2008, 8, 28–29. [Google Scholar]
- Chacón-Lee, L.T.; González-Mariño, G.E. Microalgae for “Healthy” Foods—Possibilities and Challenges. Compr. Rev. Food Sci. Food Saf. 2010, 9, 655–675. [Google Scholar] [CrossRef]
- Daghigh, F.; Fukuto, J.M.; Ash, D.E. Inhibition of rat liver arginase by an intermediate in NO biosynthesis, NG-hydroxy-L-arginine: Implications for the regulation of nitric oxide biosynthesis by arginase. Biochem. Biophys. Res. Commun. 1994, 202, 174–180. [Google Scholar] [CrossRef]
- Pope, J.A.; Karuppiah, K.; Cardounel, A.J. Role of the PRMT-DDAH-ADMA axis in the regulation of endothelial nitric oxide production. Pharmacol. Res. 2009, 60, 461–465. [Google Scholar] [CrossRef]
- Flanagan, W.F.; Holmes, E.W.; Sabina, R.L.; Swain, J.L. Importance of purine nucleotide cycle to energy production in skeletal muscle. Am. J. Physiol. 1986, 251, C795–C802. [Google Scholar] [CrossRef]
- Avila, M.A.; Garcıa-Trevijano, E.R.; Lu, S.C.; Corrales, F.J.; Mato, J.M. Methylthioadenosine. Int. J. Biochem. Cell Biol. 2004, 36, 2125–2130. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.A.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygiel-Gorniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Montgomery, M.K.; Fiveash, C.E.; Osborne, B.; Cooney, G.J.; Bell-Anderson, K.; Turner, N. PPARα-independent actions of omega-3 PUFAs contribute to their beneficial effects on adiposity and glucose homeostasis. Sci. Rep. 2014, 4, 5538. [Google Scholar] [CrossRef] [PubMed]
- Van Vlies, N.; Ferdinandusse, S.; Turkenburg, M.; Wanders, R.J.; Vaz, F.M. PPARα-activation results in enhanced carnitine biosynthesis and OCTN2-mediated hepatic carnitine accumulation. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1767, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Makowski, L.; Noland, R.C.; Koves, T.R.; Xing, W.; Ilkayeva, O.R.; Muehlbauer, M.J.; Stevens, R.D.; Muoio, D.M. Metabolic profiling of PPARalpha-/- mice reveals defects in carnitine and amino acid homeostasis that are partially reversed by oral carnitine supplementation. FASEB J. 2009, 23, 586–604. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Trehan, I.; Li, X.; Moaddel, R.; Ordiz, M.I.; Maleta, K.M.; Kraemer, K.; Shardell, M.; Ferrucci, L.; Manary, M. Environmental Enteric Dysfunction is Associated with Carnitine Deficiency and Altered Fatty Acid Oxidation. EBioMedicine 2017, 17, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Yahagi, N.; Izumida, Y.; Nishi, M.; Kubota, M.; Teraoka, Y.; Yamamoto, T.; Matsuzaka, T.; Nakagawa, Y.; Iizuka, Y.; et al. Polyunsaturated fatty acids selectively suppress sterol regulatory element-binding protein-1 through proteolytic processing and autoloop regulatory circuit. J. Biol. Chem. 2010, 285, 11681–11691. [Google Scholar] [CrossRef] [PubMed]
- Jesch, D.E.; Carr, T.P. Food Ingredients That Inhibit Cholesterol Absorption. Prev. Nutr. Food Sci. 2017, 22, 67–80. [Google Scholar]
- Zhu, X.; Song, J.; Mei-Heng, M.A.R.; Edwards, L.J.; Zeisel, S.H. Phosphatidylethanolamine N-methyltransferase (PEMT) knockout mice have hepatic steatosis and abnormal hepatic choline metabolite concentrations despite ingesting a recommended dietary intake of choline. Biochem. J. 2003, 370, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Vance, D.E. Expression of phosphatidylethanolamine N-methyltransferase-2 is markedly enhanced in long term choline-deficient rats. J. Biol. Chem. 1996, 271, 2839–2843. [Google Scholar] [CrossRef] [PubMed]
- Ikawa, M.; Borowski, P.T.; Chakravarti, A. Choline and inositol distribution in algae and fungi. Appl. Microbiol. 1968, 16, 620–623. [Google Scholar] [PubMed]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, H.J.; Wang, X.C.; Wu, S.G.; Wang, J.; Xu, L.; Qi, G.H. Dietary choline and phospholipid supplementation enhanced docosahexaenoic acid enrichment in egg yolk of laying hens fed a 2% Schizochytrium powder-added diet. Poult. Sci. 2017, 96, 2786–2794. [Google Scholar] [CrossRef] [PubMed]
- Forman, J.H.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Ann. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Piche, A.L.; Draper, H.H.; Cole, P.D. Malondialdehyde excretion by subjects consuming cod liver oil vs a concentrate of n-3 fatty acids. Lipids 1988, 23, 370–371. [Google Scholar] [CrossRef] [PubMed]
- De Raposo, F.M.; de Morais, A.M.; de Morais, R.M. Emergent Sources of Prebiotics: Seaweeds and Microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Thompson, L.; Spiller, R.C. Impact of polyunsaturated fatty acids on human colonic bacterial metabolism: An in vitro and in vivo study. Br. J. Nutr. 2007, 74, 733–741. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Harris, S.C.; Bhowmik, S.; Kang, D.J.; Hylemon, P.B. Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes 2016, 7, 22–39. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Engle, A.; Simi, B.; Goldman, M. Effect of dietary fiber on colonic bacterial enzymes and bile acids in relation to colon cancer. Gastroenterology 1992, 102, 1475–1482. [Google Scholar] [CrossRef]
- Salmean, Y.A.; Segal, M.S.; Palii, S.P.; Dahl, W.J. Fiber supplementation lowers plasma p-cresol in chronic kidney disease patients. J. Ren. Nutr. 2015, 25, 316–320. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Markers of CTL Diet | Markers of 5% Diet | Markers of 20% Diet | |
|---|---|---|---|
| Liver | Adenione 1, adenine 1, choline 1, nicotinamide 1 | Glutathione (GSH) 1, dephospho-CoA 2, adenylosuccinate (S-AMP) 1, AMP 1 | Oxidized glutathione (GSSG) 1, dimethylarginine 1, carnitine 1, acetylcarnitine 1, 3-dehydroxycarnitine 3, glutarylcarnitine 3, glycerylphosphoethanolamine 3, 5′-methylthioadenosine (MTA) 1, malondialdehyde (MDA) 3, heptanal 1, hexanal 1 |
| Serum | Palmitoleic acid 3 | Glutamate 1, suberic acid 1 | Glycine 1, arginine 1, sebacic acid 1, capric acid 3 |
| Urine | p-Cresol sulfate 2, p-cresol glucuronide 2, glycine 1, phenylalanine 1, pyroglutamate 1,indolelactic-acid 3, 3-oxo-4-pentenoic acid 3 | Riboflavin 1, pyridoxine 1, pantothenic acid 1, glycol-4-methyl pentanoic acid 1, glycol-4-methyl hexanoic acid 1, heptenal 1, sebacic acid 1, capric acid 3 | |
| Feces | Indole-3-carboxylic acid 1, coprocholic acid 3, hydrocinnamic acid 3, eicosatrienoic acid 3, 3-hydroxy-hexadecanoic acid 3, 7-ketodeoxycholic acid 3 | Muricholic acid (MCA) 1, deoxycholic acid (DCA) 1, lithocholic acid (LCA) 1, N-acetylhistamine 3 | α-Linolenic acid 1, linoleic acid 1, oleic acid 1, tauromuricholic acid (TMCA) 1, taurochenodeoxycholic acid (TCDCA) 1, taurocholic acid (TCA) 1, acetic acid1, butyric acid 1, propionic acid 1, valeric acid 1, 3-hydroxy-b,e-caroten-3′-one 3, chlorophyllide b 3 |
| Serum (μM) | Liver (μg/g Tissue) | |||||||
|---|---|---|---|---|---|---|---|---|
| CTL | 5% | 20% | P-Value | CTL | 5% | 20% | P-Value | |
| Ala | 209.35 ± 33.23 | 236.3 ± 86.33 | 235.01 ± 44.69 | 0.599 | 210.22 ± 34.46 | 222.46 ± 35.82 | 186.01 ± 39.50 | 0.154 |
| Arg * | 189.76 ± 47.01 ab | 169.36 ± 66.64 a | 251.15 ± 43.27 b | 0.037 | 4.42 ± 0.16 a | 4.58 ± 0.09 a | 4.94 ± 0.21 b | <0.001 |
| Asn | 59.06 ± 14.85 | 63.24 ± 37.03 | 66.88 ± 32.66 | 0.872 | 23.02 ± 5.69 | 23.60 ± 5.22 | 22.25 ± 4.77 | 0.875 |
| Asp | 16.18 ± 2.60 ab | 19.06 ± 4.15 b | 14.06 ± 1.83 a | 0.012 | 58.09 ± 9.40 a | 73.69 ± 19.44 b | 55.07 ± 16.09 ab | 0.051 |
| Cit | 63.85 ± 15.44 ab | 59.09 ± 27.04 a | 87.03 ± 18.62 b | 0.032 | 5.56 ± 3.09 a | 4.02 ± 0.96 b | 9.31 ± 2.13b | 0.000 |
| Gln | 624.04 ± 87.01 | 612.60 ± 115.57 | 655.91 ± 69.84 | 0.632 | 1025.03 ± 226.17 | 1078.73 ± 388.07 | 976.34 ± 247.78 | 0.789 |
| Glu | 88.73 ± 12.19 | 101.84 ± 23.28 | 76.75 ± 16.16 | 0.065 | 354.49 ± 95.59 | 437.33 ± 121.75 | 310.89 ± 130.99 | 0.114 |
| Gly | 302.98 ± 47.69 a | 325.65 ± 55.44 ab | 383.40 ± 77.11 b | 0.044 | 276.28 ± 61.80 | 266.79 ± 58.33 | 233.65 ± 37.91 | 0.272 |
| His * | 97.93 ± 17.58 | 104.71 ± 33.91 | 97.46 ± 28.02 | 0.840 | 102.56 ± 15.82 | 101.49 ± 35.32 | 93.93 ± 17.81 | 0.322 |
| Ile/Leu * | 120.65 ± 24.04 | 139.01 ± 43.68 | 151.53 ± 46.01 | 0.306 | 45.04 ± 11.34 | 50.70 ± 26.84 | 46.06 ± 10.59 | 0.601 |
| Lys * | 542.26 ± 90.24 a | 405.56 ± 63.32 b | 439.35 ± 72.62 b | 0.006 | 100.18 ± 27.85 | 98.27 ± 38.28 | 94.53 ± 21.71 | 0.930 |
| Met * | 25.54 ± 7.03 a | 35.83 ± 17.73 ab | 41.96 ± 11.76 b | 0.050 | 14.10 ± 3.35 | 13.71 ± 2.74 | 13.99 ± 3.95 | 0.972 |
| Orn | 143.29 ± 31.78 a | 155.24 ± 46.54 ab | 166.18 ± 43.12 b | 0.545 | 43.31 ± 15.75 | 36.58 ± 11.81 | 38.26 ± 11.44 | 0.574 |
| Phe * | 89.85 ± 20.86 | 99.04 ± 36.21 | 108.95 ± 22.25 | 0.393 | 30.97 ± 6.57 | 37.04 ± 16.30 | 34.94 ± 7.93 | 0.857 |
| Pro | 125.51 ± 34.56 | 154.58 ± 82.08 | 164.88 ± 42.78 | 0.376 | 50.92 ± 23.48 | 52.92 ± 19.28 | 50.95 ± 16.47 | 0.974 |
| Ser | 198.01 ± 30.60 | 210.75 ± 69.02 | 209.04 ± 52.15 | 0.873 | 41.01 ± 7.28 | 41.97 ± 17.28 | 44.76 ± 13.58 | 0.845 |
| Tau | 774.51 ± 99.48 a | 698.40 ± 114.27 a | 564.45 ± 61.28 b | 0.001 | 2844.17 ± 336.96 a | 2682.88 ± 451.51 b | 2358.50 ± 274.23 b | 0.036 |
| Thr * | 182.54 ± 39.22 a | 127.68 ± 50.76 b | 136.21 ± 25.90 ab | 0.026 | 44.70 ± 11.74 | 45.95 ± 11.41 | 41.53 ± 11.64 | 0.738 |
| Try * | 136.90 ± 19.45 | 142.41 ± 20.01 | 121.20 ± 37.15 | 0.776 | 18.08 ± 2.33 | 18.56 ± 4.01 | 17.10 ± 2.45 | 0.626 |
| Tyr | 121.85 ± 24.15 | 140.49 ± 44.45 | 153.90 ± 46.08 | 0.599 | 59.20 ± 15.25 | 65.52 ± 36.55 | 58.84 ± 14.24 | 0.827 |
| Val * | 242.94 ± 46.19 | 276.93 ± 112.46 | 270.08 ± 78.47 | 0.695 | 68.63 ± 18.98 | 77.47 ± 31.43 | 60.94 ± 14.85 | 0.369 |
| Total | 4376.13 ± 460.59 | 4298.71 ± 895.25 | 4451.39 ± 580.96 | 0.902 | 5245.86 ± 483.405 | 5404.06 ± 1108.41 | 4610.51 ± 713.365 | 0.142 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Zhou, W.; Chen, P.; Urriola, P.E.; Shurson, G.C.; Ruan, R.; Chen, C. Metabolomic Evaluation of Scenedesmus sp. as a Feed Ingredient Revealed Dose-Dependent Effects on Redox Balance, Intermediary and Microbial Metabolism in a Mouse Model. Nutrients 2019, 11, 1971. https://doi.org/10.3390/nu11091971
Ma Y, Zhou W, Chen P, Urriola PE, Shurson GC, Ruan R, Chen C. Metabolomic Evaluation of Scenedesmus sp. as a Feed Ingredient Revealed Dose-Dependent Effects on Redox Balance, Intermediary and Microbial Metabolism in a Mouse Model. Nutrients. 2019; 11(9):1971. https://doi.org/10.3390/nu11091971
Chicago/Turabian StyleMa, Yiwei, Wenguang Zhou, Paul Chen, Pedro E. Urriola, Gerald C. Shurson, Roger Ruan, and Chi Chen. 2019. "Metabolomic Evaluation of Scenedesmus sp. as a Feed Ingredient Revealed Dose-Dependent Effects on Redox Balance, Intermediary and Microbial Metabolism in a Mouse Model" Nutrients 11, no. 9: 1971. https://doi.org/10.3390/nu11091971