Prevention of Adult Colitis by Oral Ferric Iron in Juvenile Mice Is Associated with the Inhibition of the Tbet Promoter Hypomethylation and Gene Overexpression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
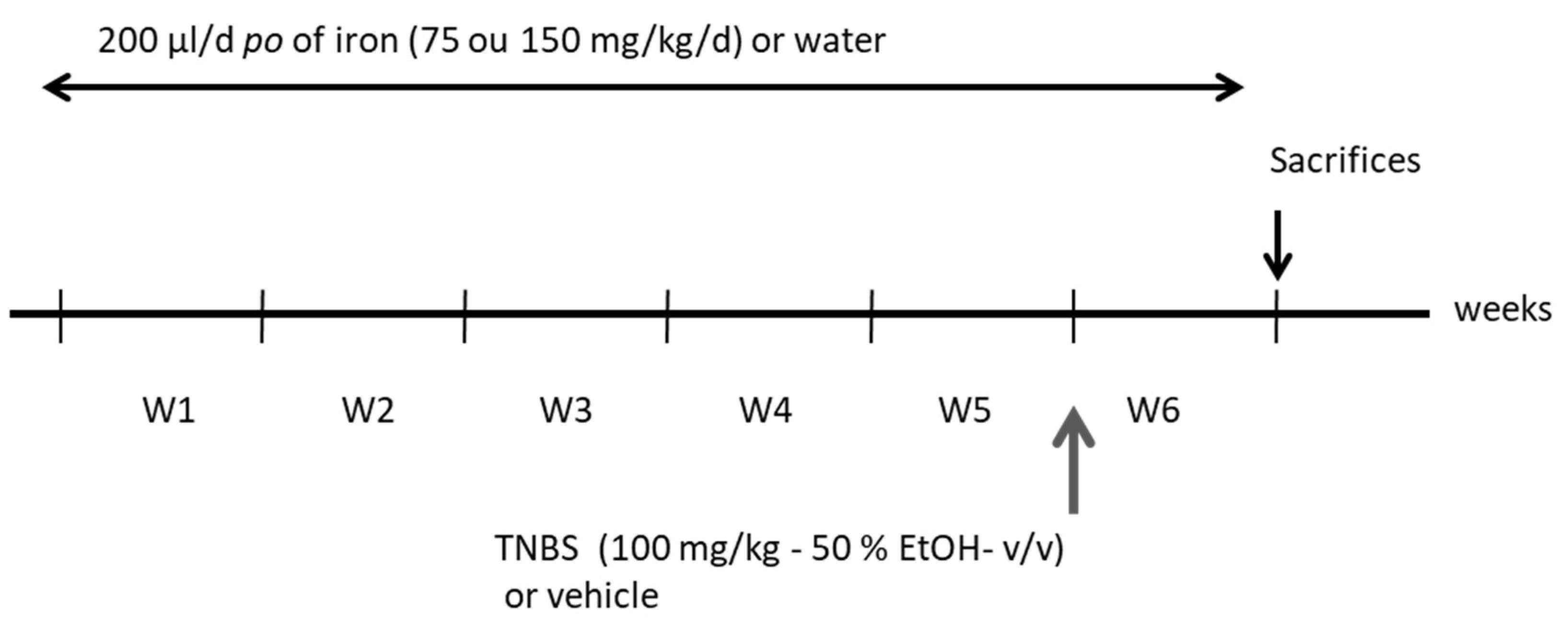
2.3. Experimental Procedure
2.4. Experimental Inflammation
2.5. Assessment of Colitis
2.5.1. Macroscopic Lesions
2.5.2. Myeloperoxidase (MPO) Assay
2.6. Splenic Th Transcription Factors Gene Expression
2.7. Methylation Status of CpG Islands
2.8. Statistical Analysis
3. Results
3.1. Phenotypic Parameters of Colitis after Microencapsulated Ferric Iron Supplementation
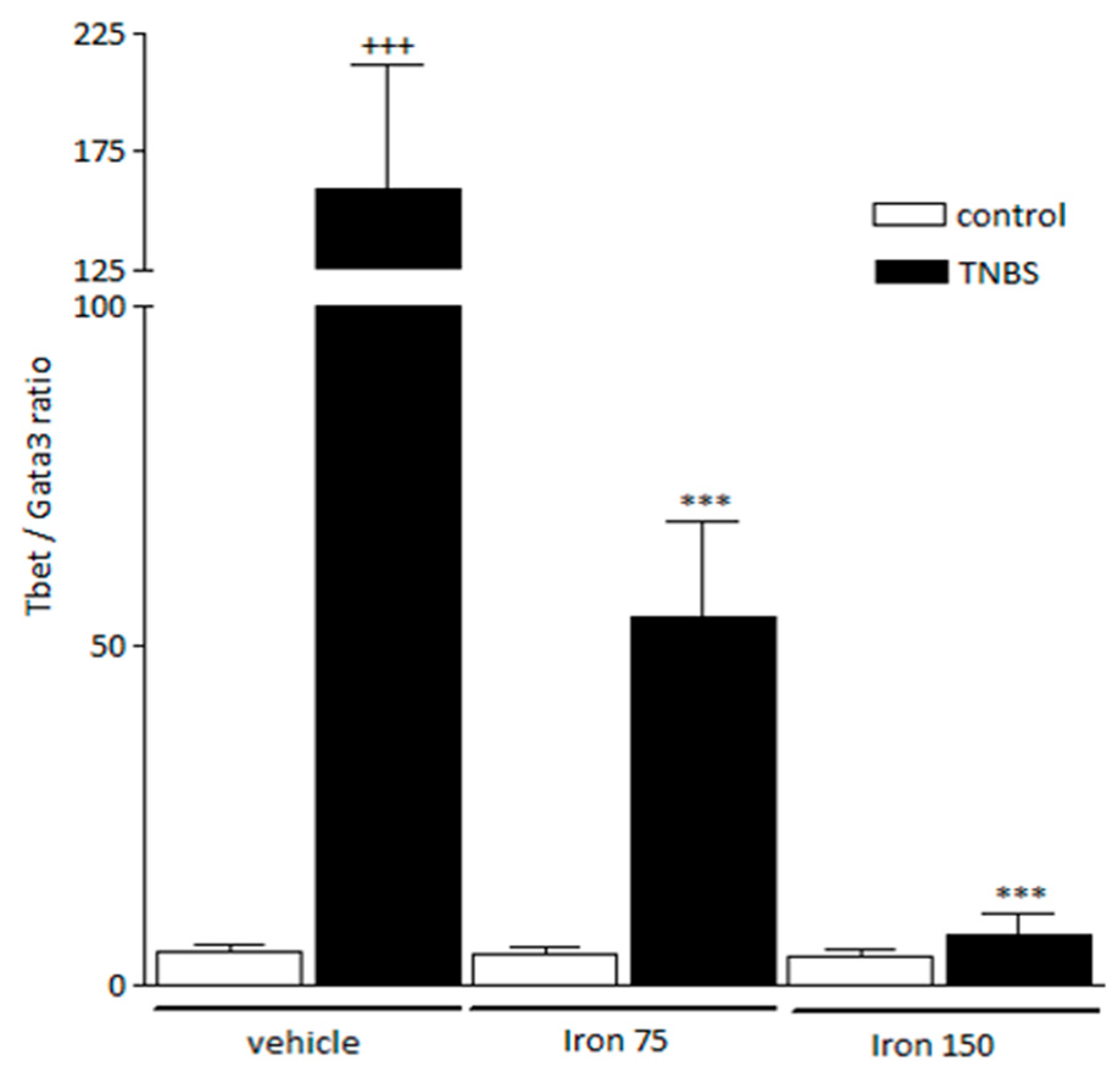
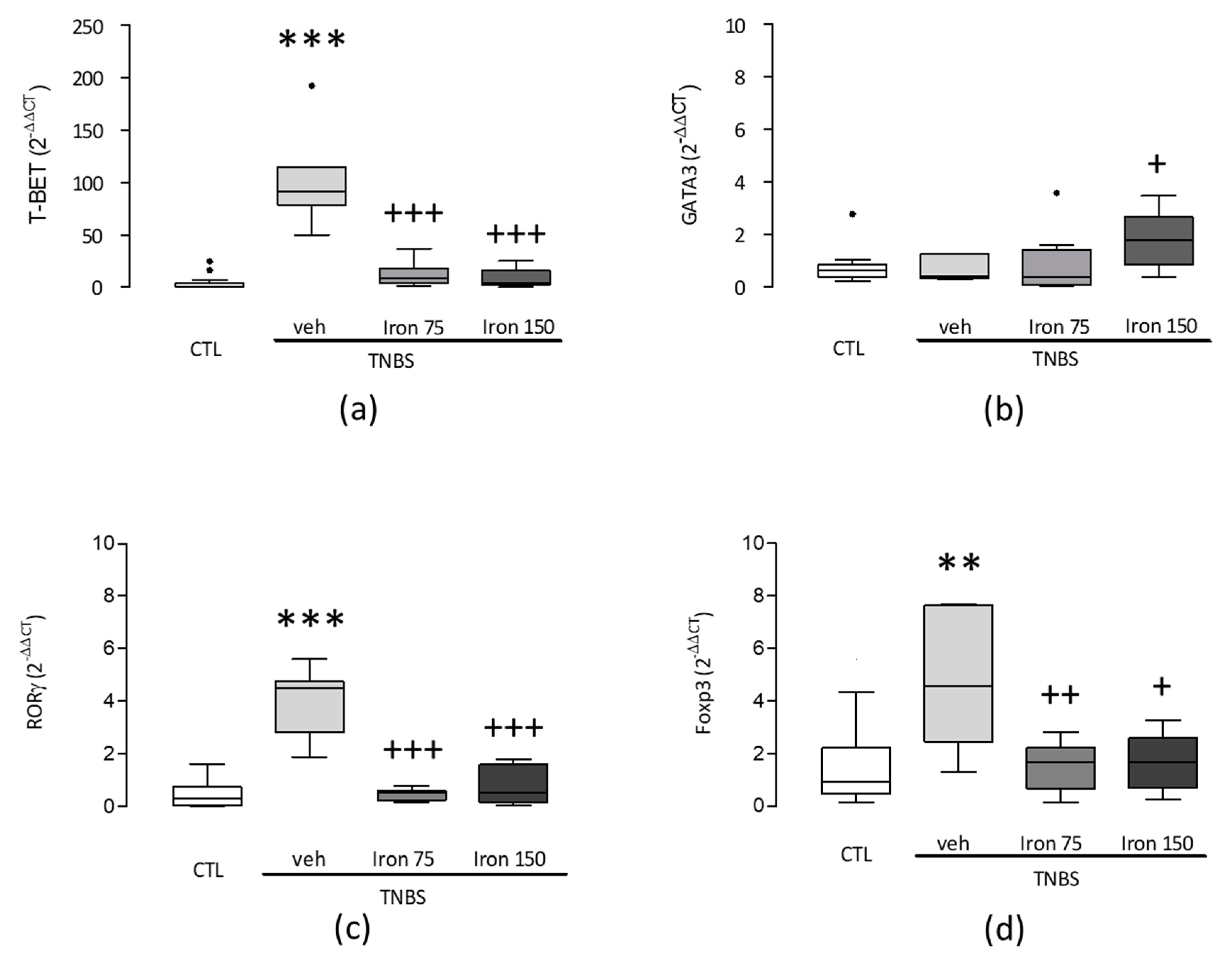
3.2. Beneficial Effect of Iron in the Pathologic Th1 Immune Orientation:
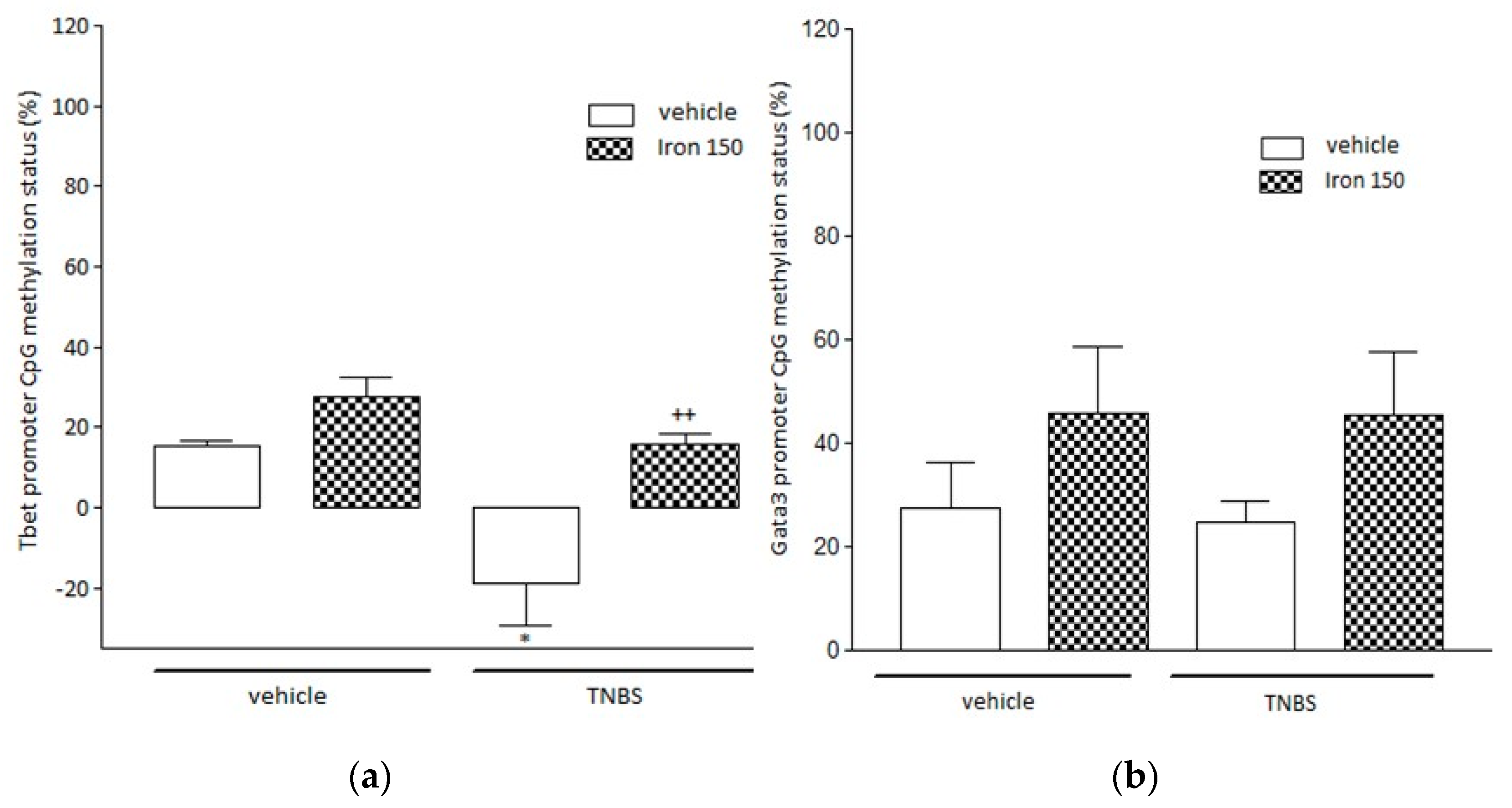
3.3. Evaluation of Locus Specific Methylation in Tbet and Gata3 Gene Promoters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ettreiki, C.; Gadonna-Widehem, P.; Mangin, I.; Coëffier, M.; Delayre-Orthez, C.; Anton, P.M. Juvenile ferric iron prevents microbiota dysbiosis and colitis in adult rodents. World J. Gastroenterol. WJG 2012, 18, 2619–2629. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, P.W.; Vaidya, S.A.; Cheng, G. The Art of War: Innate and adaptive immune responses. Cell. Mol. Life Sci. 2003, 60, 2604–2621. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; Coutts, A.G.P. Early nutrition and the development of immune function in the neonate. Proc. Nutr. Soc. 2000, 59, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C.; Krauss-Etschmann, S.; de Jong, E.C.; Dupont, C.; Frick, J.-S.; Frokiaer, H.; Heinrich, J.; Garn, H.; Koletzko, S.; Lack, G.; et al. Early nutrition and immunity—Progress and perspectives. Br. J. Nutr. 2006, 96, 774–790. [Google Scholar] [PubMed]
- Paparo, L.; Di Costanzo, M.; Di Scala, C.; Cosenza, L.; Leone, L.; Nocerino, R.; Canani, R.B. The influence of early life nutrition on epigenetic regulatory mechanisms of the immune system. Nutrients 2014, 6, 4706–4719. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. Heterogeneity and plasticity of T helper cells. Cell Res. 2010, 20, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Krausgruber, T.; Schiering, C.; Adelmann, K.; Harrison, O.J.; Chomka, A.; Pearson, C.; Ahern, P.P.; Shale, M.; Oukka, M.; Powrie, F. T-bet is a key modulator of IL-23-driven pathogenic CD4+ T cell responses in the intestine. Nat. Commun. 2016, 7, 1–12. [Google Scholar]
- Reiner, S.L. Epigenetic control in the immune response. Hum. Mol. Genet. 2005, 14, 41–46. [Google Scholar] [CrossRef]
- Wilson, C.B.; Rowell, E.; Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat. Rev. Immunol. 2009, 9, 91–105. [Google Scholar] [CrossRef]
- Cedar, H.; Bergman, Y. Epigenetic mechanisms Epigenetics of haematopoietic cell development. Nat. Rev. Immunol. 2011, 11, 478–488. [Google Scholar] [CrossRef]
- Hamm, C.A.; Stevens, J.W.; Xie, H.; Vanin, E.F.; Morcuende, J.A.; Abdulkawy, H.; Seftor, E.A.; Sredni, S.T.; Bischof, J.M.; Wang, D.; et al. Microenvironment alters epigenetic and gene expression profiles in Swarm rat chondrosarcoma tumors. BMC Cancer 2010, 10, 471. [Google Scholar] [CrossRef] [PubMed]
- Feil, R.; Fraga, M.F. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.; Aiello, A.E.; Wildman, D.E.; Koenen, K.C.; Pawelec, G.; de los Santos, R.; Goldmann, E.; Galea, S. Epigenetic and immune function profiles associated with posttraumatic stress disorder. Proc. Natl. Acad. Sci. USA 2010, 107, 9470–9475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olden, K.; Lin, Y.S.; Gruber, D.; Sonawane, B. Epigenome: Biosensor of cumulative exposure to chemical and nonchemical stressors related to environmental justice. Am. J. Public Health 2014, 104, 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Morera, J.L.; Calvanese, V.; Rodríguez-Rodero, S.; Menéndez-Torre, E.; Fraga, M.F. Epigenetic regulation of the immune system in health and disease. Tissue Antigens 2010, 76, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Isidoro-García, M.; Dávila-González, I.; Pascual De Pedro, M.; Sanz-Lozano, C.; Lorente-Toledano, F. Interactions between genes and the environment. Epigenetics in allergy. Allergol. Immunopathol. Madr. 2007, 35, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, B. Primer: Epigenetics of autoimmunity. Nat. Clin. Pract. Rheumatol. 2007, 3, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Wren, J.D.; Garner, H.R. Data-mining analysis suggests an epigenetic pathogenesis for type 2 diabetes. J. Biomed. Biotechnol. 2005, 2005, 104–112. [Google Scholar] [CrossRef]
- Waterland, R.A.; Dolinoy, D.C.; Lin, J.R.; Smith, C.A.; Shi, X.; Tahiliani, K.G. Maternal methyl supplements increase offspring DNA methylation at Axin fused. Genesis 2006, 44, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Waterland, R.A. Assessing the Effects of High Methionine Intake on DNA Methylation. J. Nutr. 2006, 136, 1706–1710. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, L.; Boukouvala, S.; Sim, E. Characterisation of CpG methylation in the upstream control region of mouse Nat2: Evidence for a gene-environment interaction in a polymorphic gene implicated in folate metabolism. Gene 2010, 452, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.C.; Houseman, E.A.; Poage, G.M.; Godleski, J.J.; Bueno, R.; Sugarbaker, D.J.; Wiencke, J.K.; Nelson, H.H.; Marsit, C.J.; Kelsey, K.T. Integrated profiling reveals a global correlation between epigenetic and genetic alterations in mesothelioma. Cancer Res. 2010, 70, 5686–5694. [Google Scholar] [CrossRef] [PubMed]
- Strakova, J.; Williams, K.T.; Gupta, S.; Schalinske, K.L.; Kruger, W.D.; Rozen, R.; Jiracek, J.; Li, L.; Garrow, T.A. Dietary intake of S-(α-carboxybutyl)-dl-homocysteine induces hyperhomocysteinemia in rats. Nutr. Res. 2010, 30, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; MacNaughton, W.K.; Morris, G.P.; Beck, P.L. Inhibition of leukotriene synthesis markedly accelerates healing in a rat model of inflammatory bowel disease. Gastroenterology 1989, 96, 29–36. [Google Scholar] [CrossRef]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Elson, C.O.; Sartor, R.B.; Tennyson, G.S.; Riddell, R.H., II. Experimental Models of Inflammatory Bowel Disease Acetic Acid-Induced Colitis. Gastroenterology 1995, 109, 1344–1367. [Google Scholar] [CrossRef]
- Rodriguez, R.M.; Lopez-Larrea, C.; Suarez-Alvarez, B. Epigenetic dynamics during CD4+ T cells lineage commitment. Int. J. Biochem. Cell Biol. 2015, 67, 75–85. [Google Scholar] [CrossRef] [PubMed]
- An, M.M.; Fan, K.X.; Zhang, J.D.; Li, H.J.; Song, S.C.; Liu, B.G.; Gao, P.H.; Zhou, Q.; Jiang, Y.Y. Lymphtoxin β receptor-Ig ameliorates TNBS-induced colitis via blocking LIGHT/HVEM signaling. Pharmacol. Res. 2005, 52, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Dubuquoy, L.; Barnouin, R.; Annicotte, J.-S.; Magnier, B.; Notti, M.; Corazza, N.; Antal, M.C.; Metzger, D.; Desreumaux, P.; et al. LRH-1-mediated glucocorticoid synthesis in enterocytes protects against inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2007, 104, 13098–13103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Chakir, H.; Wang, H.; Lefebvre, D.E.; Webb, J.; Scott, F.W. T-bet/GATA-3 ratio as a measure of the Th1/Th2 cytokine profile in mixed cell populations: Predominant role of GATA-3. J. Immunol. Methods 2003, 278, 157–169. [Google Scholar] [CrossRef]
- Daniel, C.; Sartory, N.A.; Zahn, N.; Radeke, H.H.; Stein, J.M. Immune Modulatory Treatment of Trinitrobenzene Sulfonic Acid Colitis with Calcitriol Is Associated with a Change of a T Helper (Th) 1/Th17 to a Th2 and Regulatory T Cell Profile. J. Pharmacol. Exp. Ther. 2008, 324, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Eastaff-Leung, N.; Mabarrack, N.; Barbour, A.; Cummins, A.; Barry, S. Foxp3+ regulatory T cells, Th17 effector cells, and cytokine environment in inflammatory bowel disease. J. Clin. Immunol. 2010, 30, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Zhang, J.; Chen, X.; Xie, Y.F.; Pang, Y.H.; Liu, X.J. Increased CD4+CD45RA(-)FoxP3(low) cells alter the balance between Treg and Th17 cells in colitis mice. World J. Gastroenterol. 2016, 22, 9356–9367. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Konkel, J.E. TGF-β and ’adaptive ’Foxp3+ regulatory T cells. J. Mol. Cell Biol. 2010, 2, 30–36. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Martin, R.J.; Rino, J.G.; Breed, R.; Torres, R.M.; Chu, H.W. IL-23-dependent IL-17 production is essential in neutrophil recruitment and activity in mouse lung defense against respiratory Mycoplasma pneumoniae infection. Microbes Infect. 2007, 9, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Bair, S.; Spaulding, E.; Parkkinen, J.; Shulman, H.M.; Lesnikov, V.; Beauchamp, M.; Kowdley, K.V.; Deeg, H.J. Brief report Transplantation of allogeneic T cells alters iron homeostasis in NOD/SCID mice. Blood 2009, 113, 1841–1844. [Google Scholar] [CrossRef]
- Wang, C.; Collins, M.; Kuchroo, V.K. Effector T cell differentiation: Are master regulators of effector T cells still the masters? Curr. Opin. Immunol. 2015, 37, 6–10. [Google Scholar] [CrossRef]
- Loréal, O.; Bardou-Jacquet, E.; Jouanolle, A.M.; Gandon, Y.; Deugnier, Y.; Brissot, P.; Ropert, M. Métabolisme du fer et outils diagnostiques pour le clinicien. Rev. Med. Interne 2012, 33, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Constante, M.; Fragoso, G.; Lupien-Meilleur, J.; Calvé, A.; Santos, M.M. Iron Supplements Modulate Colon Microbiota Composition and Potentiate the Protective Effects of Probiotics in Dextran Sodium Sulfate-induced Colitis. Inflamm. Bowel Dis. 2017, 23, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Delgoffe, G.M.; Meyer, C.F.; Chan, W.; Powell, J.D. Anergic T Cells Are Metabolically Anergic. J. Immunol. 2009, 183, 6095–6101. [Google Scholar] [CrossRef] [PubMed]
- Buffinton, G.D.; Doe, W.F. Depleted mucosal antioxidant defences in inflammatory bowel disease. Free Radic. Biol. Med. 1995, 9, 911–918. [Google Scholar] [CrossRef]
- Werner, T.; Wagner, S.J.; Martínez, I.; Walter, J.; Chang, J.S.; Clavel, T.; Kisling, S.; Schuemann, K.; Haller, D. Depletion of luminal iron alters the gut microbiota and prevents Crohn’s disease-like ileitis. Gut 2011, 60, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Mahalhal, A.; Williams, J.M.; Johnson, S.; Ellaby, N.; Duckworth, C.A.; Burkitt, M.D.; Liu, X.; Hold, G.L.; Campbell, B.J.; Mark Pritchard, D.; et al. Oral iron exacerbates colitis and influences the intestinal microbiome. PLoS ONE 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Cyr, A.R.; Domann, F.E. The Redox Basis of Epigenetic Modifications: From Mechanisms to Functional Consequences. Antioxid. Redox Signal. 2010, 15, 551–589. [Google Scholar] [CrossRef] [PubMed]
- Gasche, C. Iron, anaemia, and inflammatory bowel diseases. Gut 2004, 53, 1190–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mencacci, A.; Cenci, E.; Boelaert, J.R.; Bucci, P.; Mosci, P.; d’Ostiani, C.F.; Bistoni, F.; Romani, L. Iron Overload Alters Innate and T Helper Cell Responses to Candida albicans in Mice. J. Infect. Dis. 2009, 175, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, W.Y.; Zhao, M.F.; Cao, X.L.; Jiang, Y.Y.; Jin, X.; Xu, P.; Yuan, T.T.; Zhang, Y.C.; Chai, X.; et al. Reactive oxygen species mediated T lymphocyte abnormalities in an iron-overloaded mouse model and iron-overloaded patients with myelodysplastic syndromes. Ann. Hematol. 2017, 96, 1085–1095. [Google Scholar] [CrossRef]
- Markota, A.; Metzger, R.; Heiseke, A.F.; Jandl, L.; Dursun, E.; Reindl, W.; Haller, D.; Id, A.B.K. Comparison of iron-reduced and iron- supplemented semisynthetic diets in T cell transfer colitis. PLoS ONE 2019, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Wolf, P.G.; Carbonero, F.; Zhong, W.; Reid, T.; Gaskins, H.R.; McIntosh, M.K. Intestinal and Systemic Inflammatory Responses Are Positively Associated with Sulfidogenic Bacteria Abundance in High-Fat–Fed Male C57BL/6J Mice. J. Nutr. 2014, 144, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/− mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Yuan, K.T.; Yu, L.; Meng, Q.H.; Chung, P.C.K.; Yang, D.H. Bifidobacterium infantis attenuates colitis by regulating T cell subset responses. World J. Gastroenterol. 2014, 20, 18316–18329. [Google Scholar] [CrossRef] [PubMed]
- Paganini, D.; Uyoga, M.A.; Zimmermann, M.B. Iron fortification of foods for infants and children in low-income countries: Effects on the gut microbiome, gut inflammation, and diarrhea. Nutrients 2016, 8, 494. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | 5′ → 3′ Sequence | Accession Number |
|---|---|---|
| Tbet | For-TGC CTA CCA GAA CGC AGA GA | NM_019507 |
| Rev-CGG AAT CCT TTG GCA AAG G | ||
| Gata3 | For-GAC CCC TTC TAC TTG CGT TTT TC | NM_008091 |
| Rev-ACA TTT TGC TTT CTG CCT TCA AA | ||
| RORɣ | For-GCT CTG CCC CCA GTG ACA | NM_011281 |
| Rev-TGC AAC CTC AAG GAA GAG ATT G | ||
| Foxp3 | For-CCT CTA GCA GTC CAC TTC ACC AA | NM_001199347 |
| Rev-TCA ATA CCT CTC TGC CAC TTT CG | ||
| Gapdh | For-CTG CCA AGT ATG ATG ACA TCA AGA | NM_008084 |
| Rev-GCC CAG GAT GCC CTT TAG T |
| Weight Variations After Inflammation (%) | Macroscopic Damage Scores (AU) | MPO Activity (U/mg Protein) | |
|---|---|---|---|
| Control | 5.03 ± 2.09 | 0 ± 0 | 1.24 ± 0.24 |
| Vehicle TNBS | –52.47 ± 20.37 ** | 5.75 ± 0.75 *** | 2.85 ± 0.29 *** |
| Iron 75 + TNBS | 0.03 ± 0.01 | 1.41 ± 0.72 +++ | 1.93 ± 0.15 + |
| Iron 150 + TNBS | –4.16 ± 0.5 | 0.20 ± 0.20 +++ | 1.84 ± 0.17 ++ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ettreiki, C.; Chango, A.; Barbezier, N.; Coeffier, M.; Anton, P.M.; Delayre-Orthez, C. Prevention of Adult Colitis by Oral Ferric Iron in Juvenile Mice Is Associated with the Inhibition of the Tbet Promoter Hypomethylation and Gene Overexpression. Nutrients 2019, 11, 1758. https://doi.org/10.3390/nu11081758
Ettreiki C, Chango A, Barbezier N, Coeffier M, Anton PM, Delayre-Orthez C. Prevention of Adult Colitis by Oral Ferric Iron in Juvenile Mice Is Associated with the Inhibition of the Tbet Promoter Hypomethylation and Gene Overexpression. Nutrients. 2019; 11(8):1758. https://doi.org/10.3390/nu11081758
Chicago/Turabian StyleEttreiki, Chourouk, Abalo Chango, Nicolas Barbezier, Moise Coeffier, Pauline M Anton, and Carine Delayre-Orthez. 2019. "Prevention of Adult Colitis by Oral Ferric Iron in Juvenile Mice Is Associated with the Inhibition of the Tbet Promoter Hypomethylation and Gene Overexpression" Nutrients 11, no. 8: 1758. https://doi.org/10.3390/nu11081758