Walnut Polyphenol Extract Protects against Fenitrothion-Induced Immunotoxicity in Murine Splenic Lymphocytes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Polyphenols
2.3. Experimental Animals
2.4. Preparation of Splenocytes
2.5. Cell Viability Assay
2.6. Flow Cytometry
2.7. ELISA
2.8. Measurement of GSH-Px, SOD, MDA and •OH Levels
2.9. Western Blotting
2.10. Fractionation of WPE by Column Chromatography
2.11. LC-MS Analyses (HPLC-ESI-IT-TOF-MS)
2.12. Statistical Analysis
3. Results
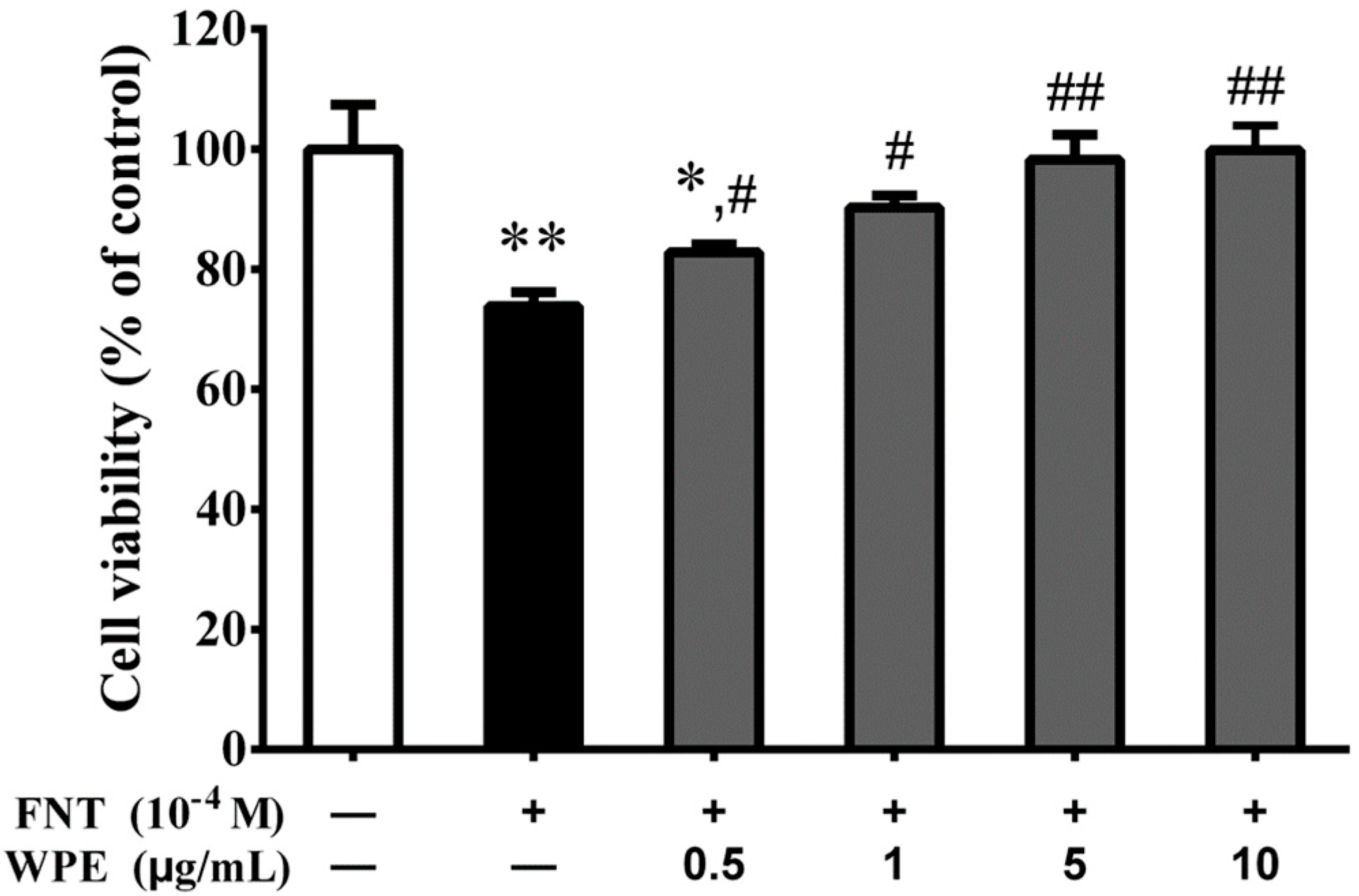
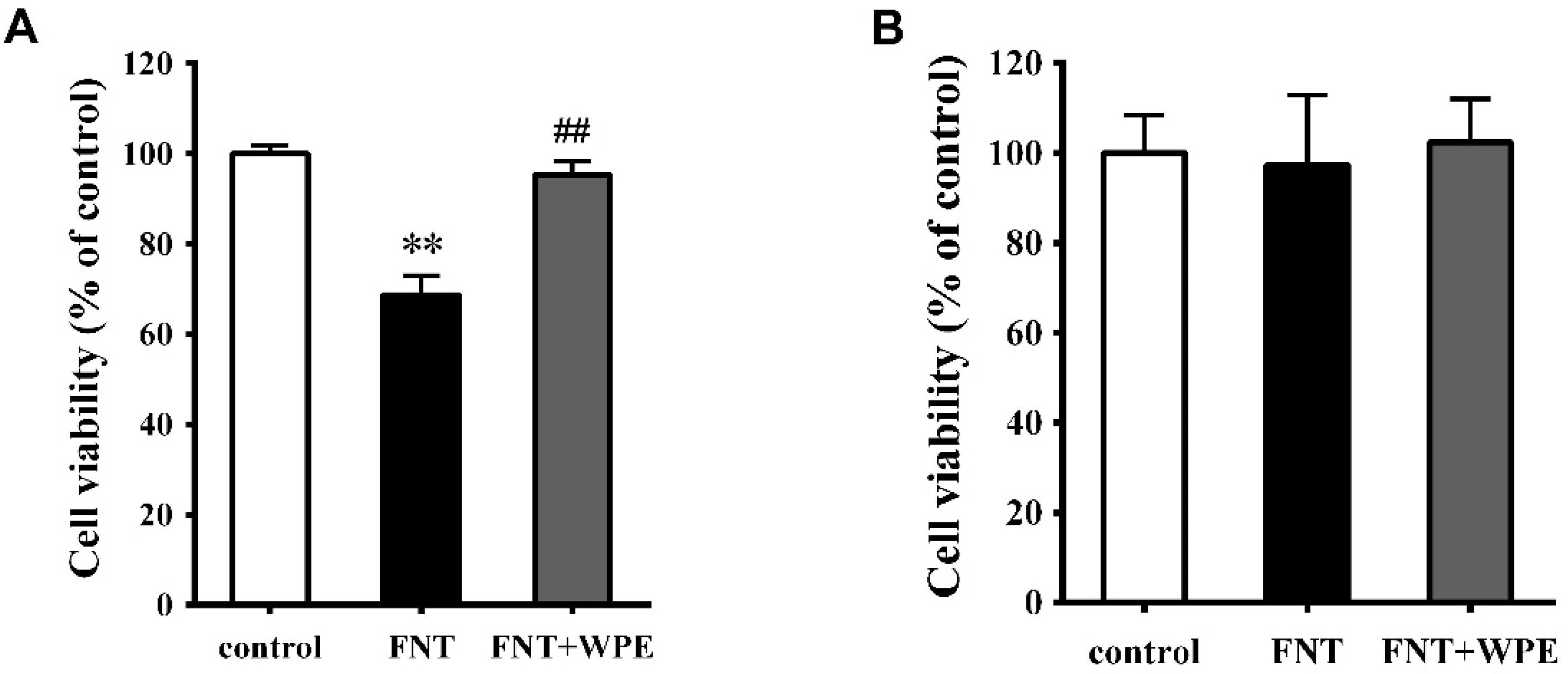
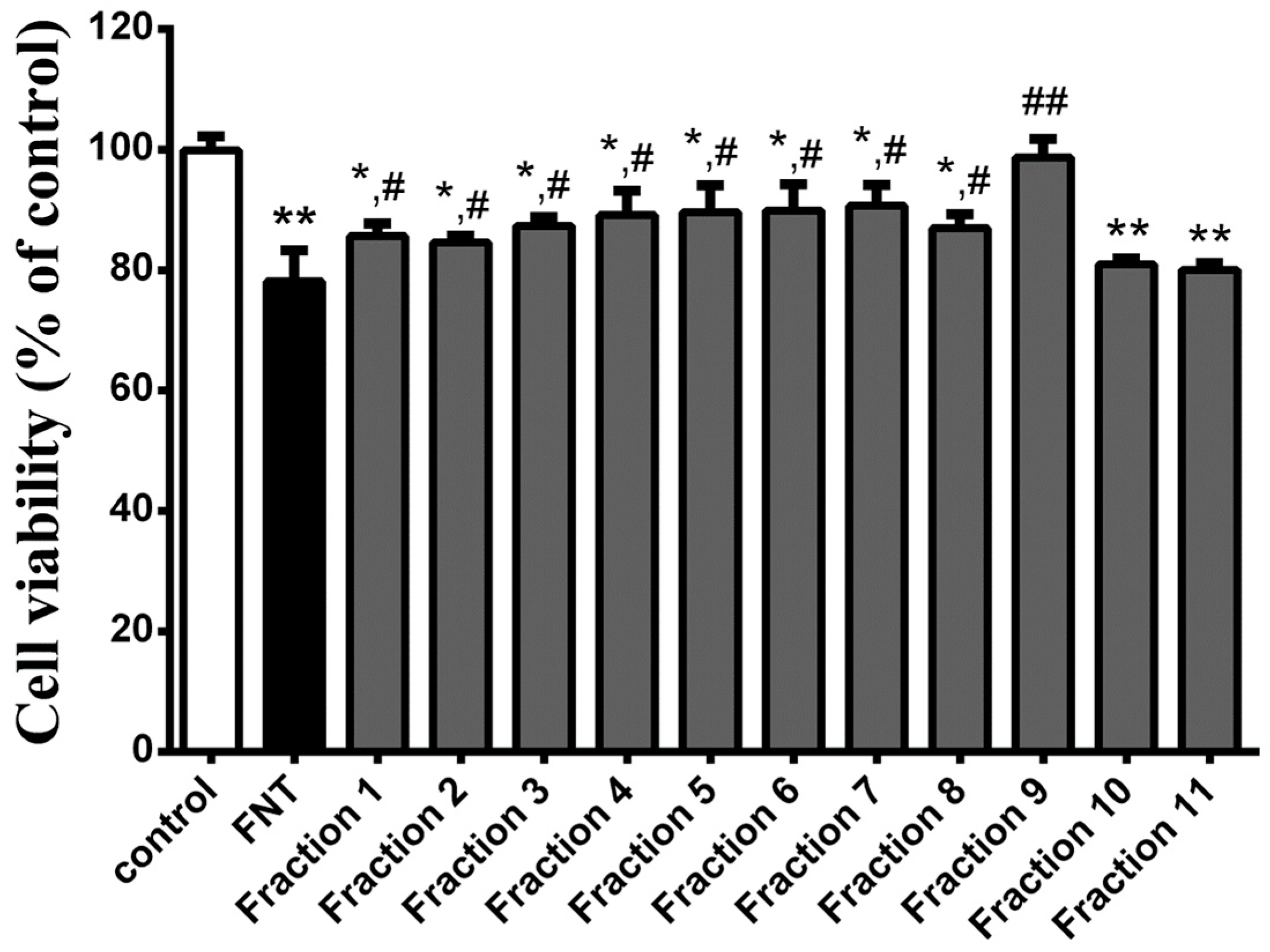
3.1. WPE Protects Splenocytes against FNT Induced Cytotoxicity
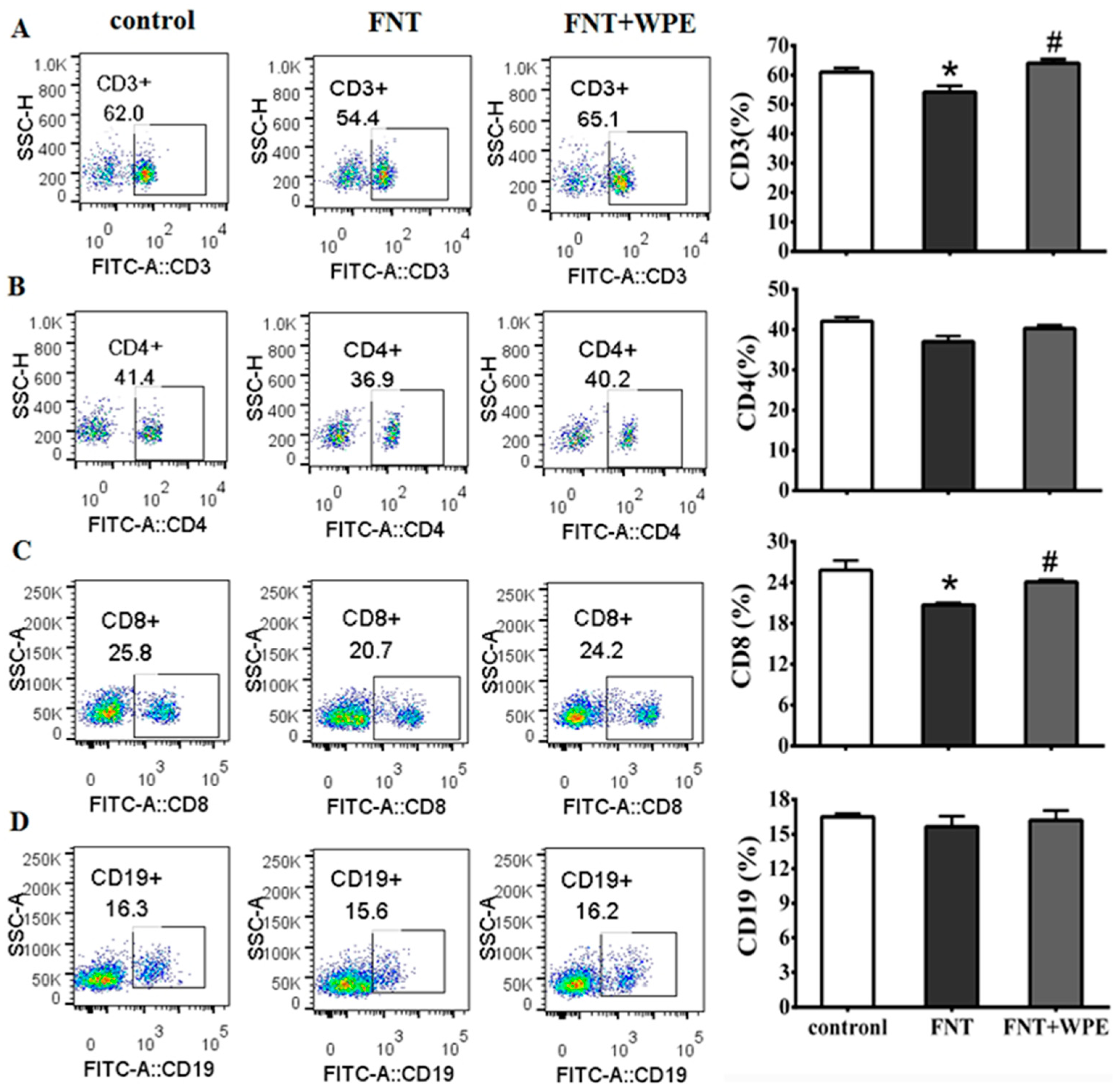
3.2. Effect of WPE on cytotoxicity of FNT in Splenic Lymphocyte Subpopulations
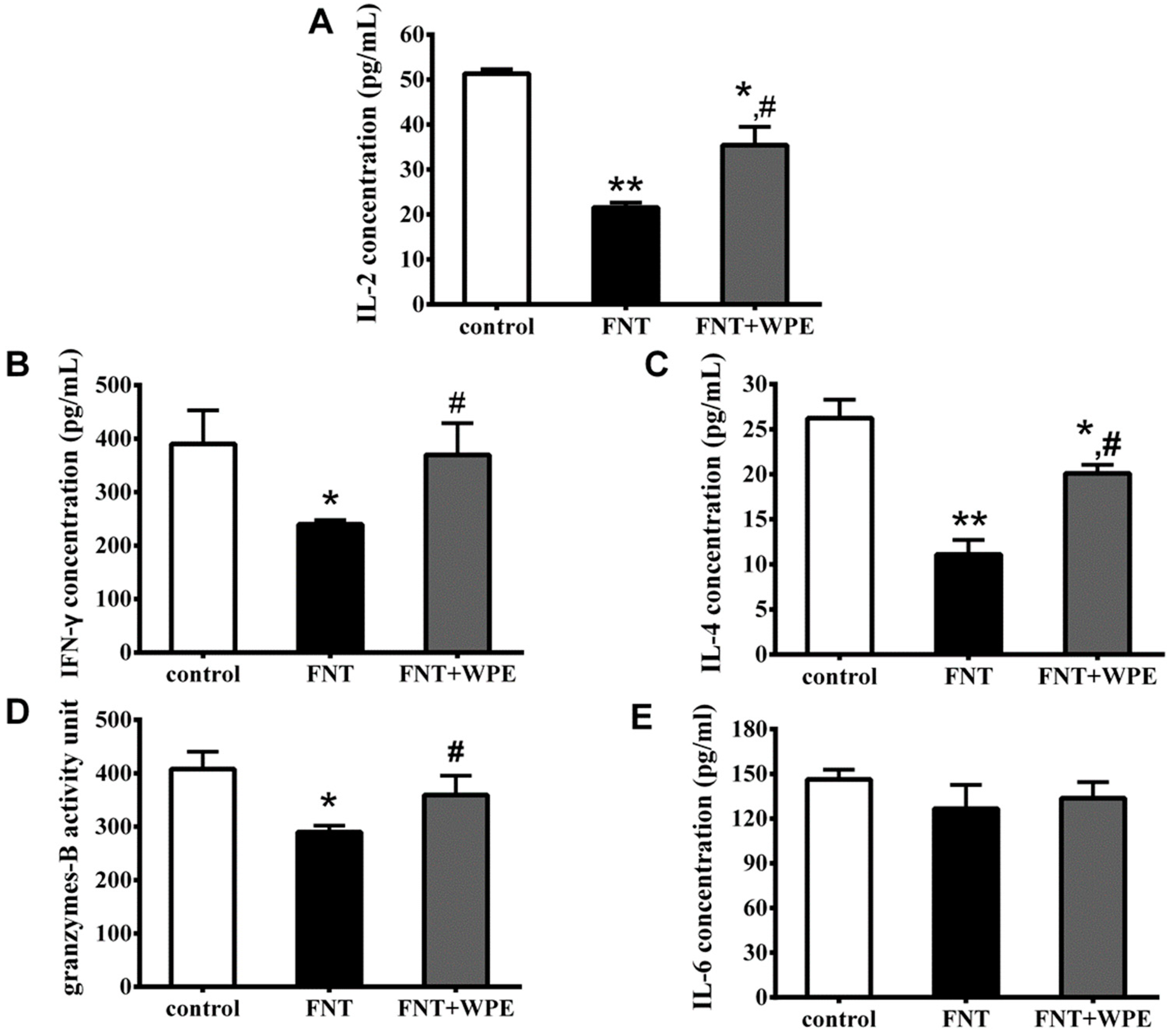
3.3. WPE Protected Against Decreases in Splenic T Cell Cytokine/Granzyme Production
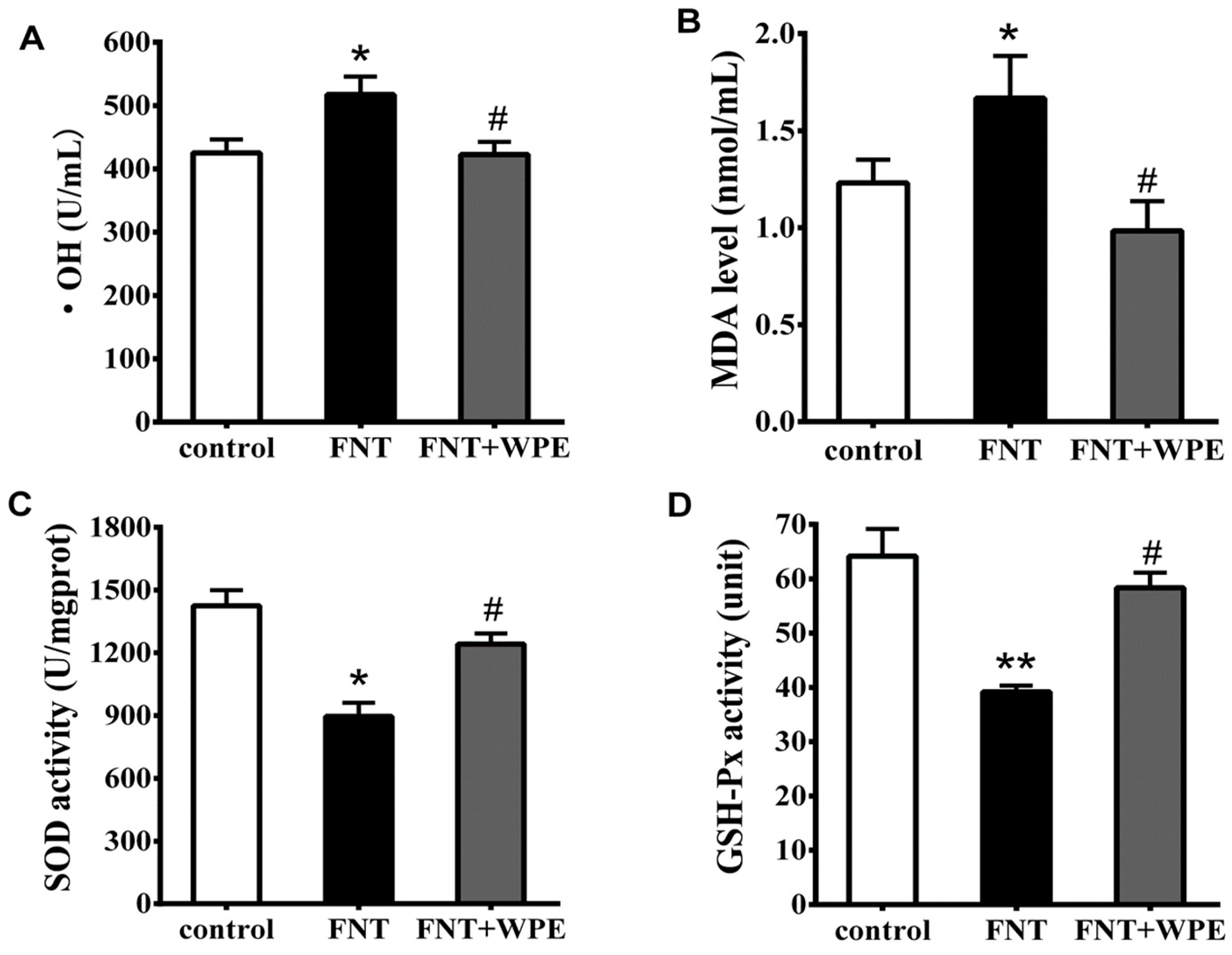
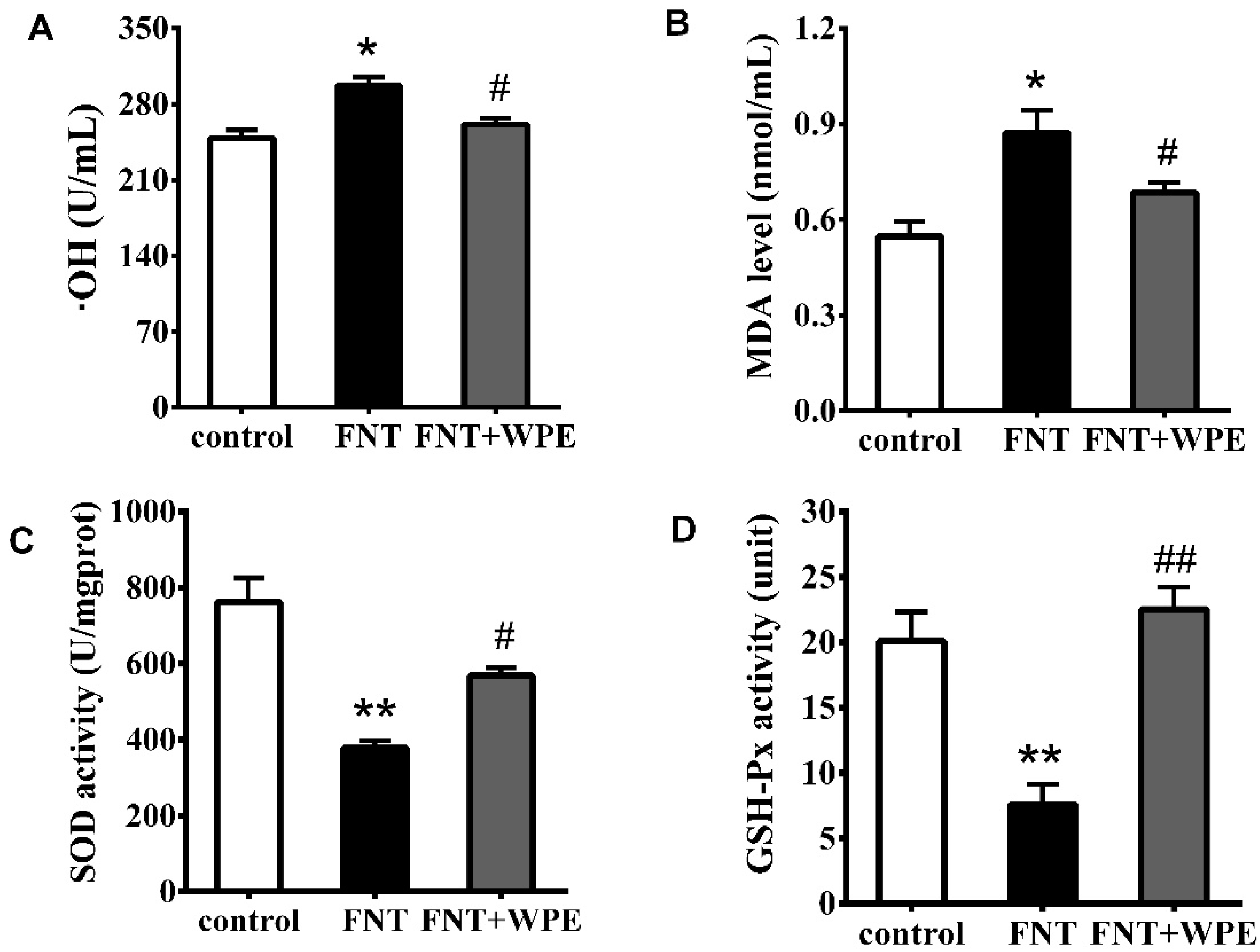
3.4. WPE Attenuates FNT Induced Oxidative Damage in Splenocytes
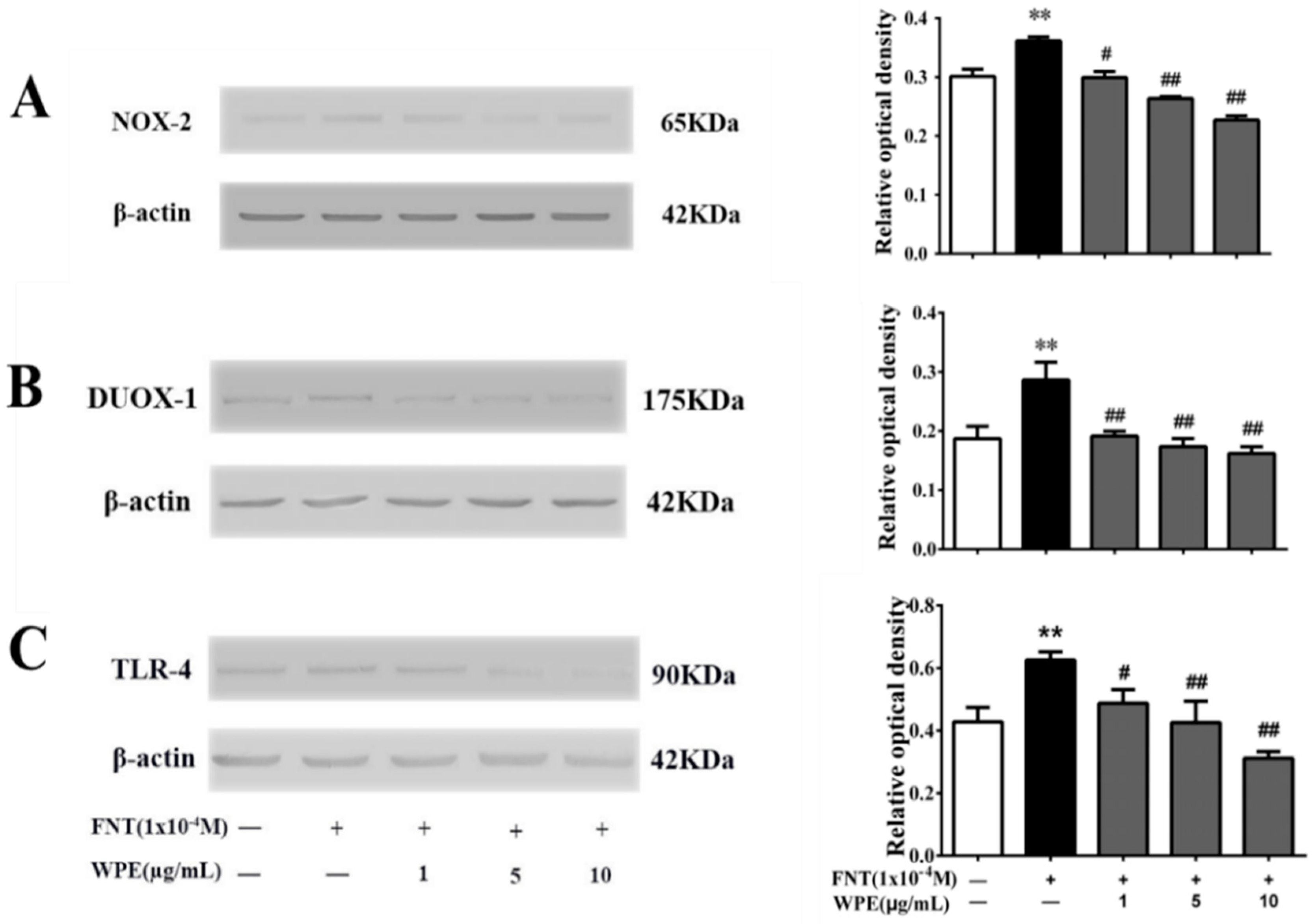
3.5. WPE Inhibits the Expression of NOX-2, DUOX-1 and TLR-4 in Splenic Cells
3.6. Identification of Phenolic Compounds in WPE
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kamath, V.; Joshi, A.K.R.; Rajini, P.S. Dimethoate induced biochemical perturbations in rat pancreas and its attenuation by cashew nut skin extract. Pestic. Biochem. Physiol. 2008, 90, 58–65. [Google Scholar] [CrossRef]
- Ramadan, G.; El-Beih, N.M.; Ahmed, R.S.A. Aged garlic extract ameliorates immunotoxicity, hematotoxicity and impaired burn-healing in malathion- and carbaryl-treated male albino rats: Modulation of Insecticides Toxicity by Age. Environ. Toxicol. 2017, 32, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, J.; Sun, Y.-J.; Yang, L.; Wu, Y.-J. Cadmium and chlorpyrifos inhibit cellular immune response in spleen of rats. Environ. Toxicol. 2017, 32, 1927–1936. [Google Scholar] [CrossRef] [PubMed]
- AL-Gehani, S.A. Effect of sub chronic exposure to Malathion on hematological parameters in the quail. Glob. Adv. Res. J. Environ. Sci. Toxicol. 2013, 2, 077–081. [Google Scholar]
- Sarikaya, R.; Selvı, M.; Erkoç, F. Investigation of acute toxicity of fenitrothion on peppered corydoras (Corydoras paleatus) (Jenyns, 1842). Chemosphere 2004, 56, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Struve, M.F.; Turner, K.J.; Dorman, D.C. Preliminary investigation of changes in the sexually dimorphic nucleus of the rat medial preoptic area following prenatal exposure to fenitrothion. J. Appl. Toxicol. 2007, 27, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Duramad, P.; Tager, I.B.; Leikauf, J.; Eskenazi, B.; Holland, N.T. Expression of Th1/Th2 cytokines in human blood afterin vitro treatment with chlorpyrifos and its metabolites, in combination with endotoxin LPS and allergen Der p1. J. Appl. Toxicol. 2006, 26, 458–465. [Google Scholar] [CrossRef] [PubMed]
- El-Bini Dhouib, I.; Lasram, M.M.; Annabi, A.; Gharbi, N.; El-Fazaa, S. A comparative study on toxicity induced by carbosulfan and malathion in Wistar rat liver and spleen. Pestic. Biochem. Physiol. 2015, 124, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Elhalwagy, M.E.A.; Darwish, N.S.; Zaher, E.M. Prophylactic effect of green tea polyphenols against liver and kidney injury induced by fenitrothion insecticide. Pestic. Biochem. Physiol. 2008, 91, 81–89. [Google Scholar] [CrossRef]
- Moon, C.K.; Yun, Y.P.; Lee, S.H.; Lee, Y.S. Effects of some organophosphate pesticides on the murine immune system following subchronic exposure (I). Arch. Pharm. Res. 1986, 9, 175–181. [Google Scholar] [CrossRef]
- Li, Q.; Kobayashi, M.; Inagaki, H.; Hirata, Y.; Sato, S.; Ishizaki, M.; Okamura, A.; Wang, D.; Nakajima, T.; Kamijima, M.; et al. Effect of oral exposure to fenitrothion and 3-methyl-4-nitrophenol on splenic cell populations and histopathological alterations in spleen in Wistar rats. Hum. Exp. Toxicol. 2011, 30, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yoshimura, T.; Mori, H.; Kawaguchi, M.; Adachi, S.; Nakao, T.; Yamazaki, F. Effects of pesticides on cytokines production by human peripheral blood mononuclear cells—Fenitrothion and glyphosate. Chudoku Kenkyu 2002, 15, 159–165. [Google Scholar] [PubMed]
- Budin, S.B.; Han, C.M.; Jayusman, P.A.; Taib, I.S. Tocotrienol rich fraction prevents fenitrothion induced pancreatic damage by restoring antioxidant status. Pak. J. Biol. Sci. 2012, 15, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, J.X.; Tian, J.L.; Wang, C.; Wang, Y.X.; Wan, Y.F.; Weng, Q.; Xu, M.Y. Selective effects of fenitrothion on murine splenic T-lymphocyte populations and cytokine/granzyme production. J. Environ. Sci. Health Part B 2018, 53, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lin, S.Q.; He, Y.L.; Liu, G.; Wang, Z.Y. Protective Effects of Quercetin on Cadmium-induced Cytotoxicity in Primary Cultures of Rat Proximal Tubular Cells. Biomed. Environ. Sci. 2013, 26, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Lin, J.; Mi, Y.; Zhang, C. Quercetin attenuates cadmium-induced oxidative damage and apoptosis in granulosa cells from chicken ovarian follicles. Reprod. Toxicol. 2011, 31, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Quercetin decreases steroidogenic enzyme activity, NF-κB expression and oxidative stress in cultured Leydig cells exposed to atrazine. Mol. Cell. Biochem. 2013, 373, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Fang, W.; Xiao, W.; Lu, L.; Jia, X. Protective role of oligomeric proanthocyanidin complex against hazardous nodularin-induced oxidative toxicity in Carassius auratus lymphocytes. J. Hazard. Mater. 2014, 274, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Ma, J.Q.; Liu, S.S.; Zheng, G.H.; Feng, Z.J.; Sun, J.M. Proanthocyanidins improves lead-induced cognitive impairments by blocking endoplasmic reticulum stress and nuclear factor-κB-mediated inflammatory pathways in rats. Food Chem. Toxicol. 2014, 72, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Karabag-Coban, F.; Bulduk, I.; Liman, R.; Ince, S.; Cigerci, I.; Hazman, O. Oleuropein alleviates malathion-induced oxidative stress and DNA damage in rats. Toxicol. Environ. Chem. 2016, 98, 101–108. [Google Scholar] [CrossRef]
- Le Sage, F.; Meilhac, O.; Gonthier, M.-P. Anti-inflammatory and antioxidant effects of polyphenols extracted from Antirhea borbonica medicinal plant on adipocytes exposed to Porphyromonas gingivalis and Escherichia coli lipopolysaccharides. Pharmacol. Res. 2017, 119, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.; Angove, M.J.; Tucci, J.; Dennis, C. Walnuts (Juglans regia) Chemical Composition and Research in Human Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Taha, N.A.; Al-Wadaan, M.A. Utility and importance of walnut, Juglans regia Linn: A review. Afr. J. Microbiol. Res. 2011, 5, 5796–5805. [Google Scholar] [CrossRef]
- Yang, L.; Ma, S.; Han, Y.; Wang, Y.; Guo, Y.; Weng, Q.; Xu, M. Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte. Nutrients 2016, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Qamar, W.; Sultana, S. Polyphenols from Juglans regia L. (walnut) kernel modulate cigarette smoke extract induced acute inflammation, oxidative stress and lung injury in Wistar rats. Hum. Exp. Toxicol. 2011, 30, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Shabani, M.; Nazeri, M.; Parsania, S.; Razavinasab, M.; Zangiabadi, N.; Esmaeilpour, K.; Abareghi, F. Walnut consumption protects rats against cisplatin-induced neurotoxicity. NeuroToxicology 2012, 33, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tanaka, J.; Kikuchi, M.; Fukuda, T.; Ito, H.; Hatano, T.; Yoshida, T. Walnut Polyphenols Prevent Liver Damage Induced by Carbon Tetrachloride and d -Galactosamine: Hepatoprotective Hydrolyzable Tannins in the Kernel Pellicles of Walnut. J. Agric. Food Chem. 2008, 56, 4444–4449. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, M.; Su, G.; Cai, M.; Zhou, C.; Huang, J.; Lin, L. The antioxidant activities and the xanthine oxidase inhibition effects of walnut (Juglans regia L.) fruit, stem and leaf. Int. J. Food Sci. Technol. 2015, 50, 233–239. [Google Scholar] [CrossRef]
- Xi, L.; Liu, Y.; Tang, Z.; Sheng, X.; Zhang, H.; Weng, Q.; Xu, M. Expression of leptin receptor in the oviduct of Chinese brown frog (Rana dybowskii). Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2017, 312, R912–R918. [Google Scholar] [CrossRef] [PubMed]
- Regueiro, J.; Sánchez-González, C.; Vallverdú-Queralt, A.; Simal-Gándara, J.; Lamuela-Raventós, R.; Izquierdo-Pulido, M. Comprehensive identification of walnut polyphenols by liquidchromatography coupled to linear ion trap–Orbitrap mass spectrometry. Food Chem. 2014, 158, 229–238. [Google Scholar] [CrossRef]
- Grace, M.H.; Warlick, C.W.; Neff, S.A.; Lila, M.A. Efficient preparative isolation and identification of walnut bioactive components using high-speed counter-current chromatography and LC-ESI-IT-TOF-MS. Food Chem. 2014, 158, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Dangroo, N.A.; Singh, J.; Gupta, N.; Singh, S.; Kaul, A.; Khuroo, M.A.; Sangwan, P.L. T- and B-cell immunosuppressive activity of novel α-santonin analogs with humoral and cellular immune response in Balb/c mice. Med. Chem. Commun. 2017, 8, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Ruwali, P.; Ambwani, T.K.; Gautam, P. In vitro immunomodulatory potential of Artemisia indica Willd. in chicken lymphocytes. Vet. World 2018, 11, 80–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabriso, N.; Massaro, M.; Scoditti, E.; D’Amore, S.; Gnoni, A.; Pellegrino, M.; Storelli, C.; De Caterina, R.; Palasciano, G.; Carluccio, M.A. Extra virgin olive oil rich in polyphenols modulates VEGF-induced angiogenic responses by preventing NADPH oxidase activity and expression. J. Nutr. Biochem. 2016, 28, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, L.; Yue, L.; Wang, B.; Li, X.; Guo, H.; Ma, Y.; Yao, C.; Gao, L.; Deng, J.; et al. Pterostilbene Attenuates Early Brain Injury Following Subarachnoid Hemorrhage via Inhibition of the NLRP3 Inflammasome and Nox2-Related Oxidative Stress. Mol. Neurobiol. 2017, 54, 5928–5940. [Google Scholar] [CrossRef] [PubMed]
- Mangum, L.C.; Borazjani, A.; Stokes, J.V.; Matthews, A.T.; Lee, J.H.; Chambers, J.E.; Ross, M.K. Organochlorine Insecticides Induce NADPH Oxidase-Dependent Reactive Oxygen Species in Human Monocytic Cells via Phospholipase A2/Arachidonic Acid. Chem. Res. Toxicol. 2015, 28, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B.; Hardy, M.; Zielonka, J. A Critical Review of Methodologies to Detect Reactive Oxygen and Nitrogen Species Stimulated by NADPH Oxidase Enzymes: Implications in Pesticide Toxicity. Curr. Pharmacol. Rep. 2016, 2, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.H.; Devadas, S.; Kwon, J.; Pinto, L.A.; Williams, M.S. T cells express a phagocyte-type NADPH oxidase that is activated after T cell receptor stimulation. Nat. Immunol. 2004, 5, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.A.; Vlisidou, I.; Blunt, M.D.; Wood, W.; Ward, S.G. Hydrogen Peroxide Triggers a Dual Signaling Axis to Selectively Suppress Activated Human T Lymphocyte Migration. J. Immunol. 2017, 198, 3679–3689. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Zanin-Zhorov, A.; Tal-Lapidot, G.; Cahalon, L.; Cohen-Sfady, M.; Pevsner-Fischer, M.; Lider, O.; Cohen, I.R. Cutting Edge: T Cells Respond to Lipopolysaccharide Innately via TLR4 Signaling. J. Immunol. 2007, 179, 41–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, A.; Ahmad, S.F.; Bakheet, S.A.; Al-Harbi, N.O.; AL-Ayadhi, L.Y.; Attia, S.M.; Zoheir, K.M.A. Toll-like receptor 4 signaling is associated with upregulated NADPH oxidase expression in peripheral T cells of children with autism. Brain Behav. Immun. 2017, 61, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jeong, J.-M.; Kim, S.J.; Seo, W.; Kim, M.-H.; Choi, W.-M.; Yoo, W.; Lee, J.-H.; Shim, Y.-R.; Yi, H.-S.; et al. Pro-inflammatory hepatic macrophages generate ROS through NADPH oxidase 2 via endocytosis of monomeric TLR4–MD2 complex. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, Q.; Chen, Y.; Wang, B.; Pei, D. Identification of major walnut cultivars grown in China based on nut phenotypes and SSR markers. Sci. Hortic. 2014, 168, 240–248. [Google Scholar] [CrossRef]
- Alleva, R.; Manzella, N.; Gaetani, S.; Ciarapica, V.; Bracci, M.; Caboni, M.F.; Pasini, F.; Monaco, F.; Amati, M.; Borghi, B.; et al. Organic honey supplementation reverses pesticide-induced genotoxicity by modulating DNA damage response. Mol. Nutr. Food Res. 2016, 60, 2243–2255. [Google Scholar] [CrossRef] [PubMed]
- Meaklim, J.; Yang, J.; Drummer, O.H.; Killalea, S.; Staikos, V.; Horomidis, S.; Rutherford, D.; Ioannides-Demos, L.L.; Lim, S.; McLean, A.J.; et al. Fenitrothion: Toxicokinetics and toxicologic evaluation in human volunteers. Environ. Health Perspect. 2003, 111, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Li, C.M. Impairment of testicular function in adult male Japanese quail (Coturnix japonica) after a single administration of 3-methyl-4-nitrophenol in diesel exhaust particles. J. Endocrinol. 2006, 189, 555–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nani, A.; Belarbi, M.; Ksouri-Megdiche, W.; Abdoul-Azize, S.; Benammar, C.; Ghiringhelli, F.; Hichami, A.; Khan, N.A. Effects of polyphenols and lipids from Pennisetum glaucum grains on T-cell activation: Modulation of Ca2+ and ERK1/ERK2 signaling. BMC Complement. Altern. Med. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Sanbongi, C.; Suzuki, N.; Sakane, T. Polyphenols in Chocolate, Which Have Antioxidant Activity, Modulate Immune Functions in Humansin Vitro. Cell. Immunol. 1997, 177, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Chabalgoity, J.A.; Baz, A.; Rial, A.; Grille, S. The relevance of cytokines for development of protective immunity and rational design of vaccines. Cytokine Growth Factor Rev. 2007, 18, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, H.; Zhang, M.; Yue, L.-T.; Wang, C.-C.; Zhang, P.; Liu, Y.; Duan, R.-S. Curcumin ameliorates experimental autoimmune myasthenia gravis by diverse immune cells. Neurosci. Lett. 2016, 626, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Maiti, M.; Chattopadhyay, K.; Verma, M.; Chattopadhyay, B. Curcumin protects against nicotine-induced stress during protein malnutrition in female rat through immunomodulation with cellular amelioration. Mol. Biol. Rep. 2015, 42, 1623–1637. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Shin, M.S.; Kang, Y.; Park, K.; Maeda, T.; Nishioka, H.; Fujii, H.; Kang, I. Oligonol, a lychee fruit-derived low-molecular form of polyphenol mixture, suppresses inflammatory cytokine production from human monocytes. Hum. Immunol. 2016, 77, 512–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magrone, T.; Fontana, S.; Laforgia, F.; Dragone, T.; Jirillo, E.; Passantino, L. Administration of a Polyphenol-Enriched Feed to Farmed Sea Bass (Dicentrarchus labrax L.) Modulates Intestinal and Spleen Immune Responses. Oxid. Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Zerin, T.; Song, H.-Y. Antioxidant Action of Ellagic Acid Ameliorates Paraquat-Induced A549 Cytotoxicity. Biol. Pharm. Bull. 2013, 36, 609–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londhe, J.S.; Devasagayam, T.P.A.; Foo, L.Y.; Shastry, P.; Ghaskadbi, S.S. Geraniin and amariin, ellagitannins from Phyllanthus amarus, protect liver cells against ethanol induced cytotoxicity. Fitoterapia 2012, 83, 1562–1568. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Singh, D.; Patel, S.; Singh, M.R. Role of enzymatic free radical scavengers in management of oxidative stress in autoimmune disorders. Int. J. Biol. Macromol. 2017, 101, 502–517. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, J.; Pang, Q.; Song, S.; Miao, R.; Chen, W.; Zhou, Y.; Liu, C. The Protective Role of Curcumin in Zymosan-Induced Multiple Organ Dysfunction Syndrome in Mice. SHOCK 2016, 45, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Chen, C.; Zhao, S.; Ge, F.; Liu, D.; Song, H. Effects of Walnut Polyphenol on Learning and Memory Functions in Hypercholesterolemia Mice. J. Food Nutr. Res. 2014, 2, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Ghosh, S.; Mukherjee, S.; Gupta, P.; Bhattacharya, S.; Adhikary, A.; Chattopadhyay, S. Pomegranate protects against arsenic-induced p53-dependent ROS-mediated inflammation and apoptosis in liver cells. J. Nutr. Biochem. 2016, 38, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Leto, T.L.; Morand, S.; Hurt, D.; Ueyama, T. Targeting and Regulation of Reactive Oxygen Species Generation by Nox Family NADPH Oxidases. Antioxid. Redox Signal. 2009, 11, 2607–2619. [Google Scholar] [CrossRef] [PubMed]
- Nisimoto, Y.; Diebold, B.A.; Cosentino-Gomes, D.; Lambeth, J.D. Nox4: A Hydrogen Peroxide-Generating Oxygen Sensor. Biochemistry 2014, 53, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Dal-Ros, S.; Zoll, J.; Lang, A.-L.; Auger, C.; Keller, N.; Bronner, C.; Geny, B.; Schini-Kerth, V.B. Chronic intake of red wine polyphenols by young rats prevents aging-induced endothelial dysfunction and decline in physical performance: Role of NADPH oxidase. Biochem. Biophys. Res. Commun. 2011, 404, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.I.; Griendling, K.K. Nox proteins in signal transduction. Free Radic. Biol. Med. 2009, 47, 1239–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampfrath, T.; Maiseyeu, A.; Ying, Z.; Shah, Z.; Deiuliis, J.A.; Xu, X.; Kherada, N.; Brook, R.D.; Reddy, K.M.; Padture, N.P.; et al. Chronic Fine Particulate Matter Exposure Induces Systemic Vascular Dysfunction via NADPH Oxidase and TLR4 Pathways. Circ. Res. 2011, 108, 716–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Z.; Yan, C.; Deng, Q.; Gao, D.; Niu, X. Curcumin inhibits LPS-induced inflammation in rat vascular smooth muscle cells in vitro via ROS-relative TLR4-MAPK/NF-κB pathways. Acta Pharmacol. Sin. 2013, 34, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR (min) | Measured [M-H]− (m/z) | Predicted [M-H]− (m/z) | MS/MS 2 Fragments (m/z) | Identification | Molecular Formula | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 5.02 | 783.0773 | 783.0681 | 481.06, 300.99, 275.02 | Pedunculagin/casuariin isomer (bis-HHDP-glucose) | C34H24O22 | [30,31] |
| 2 | 7.14 | 951.0762 | 951.0740 | 907.08, 783.07, 481.06, 300.99, 275.02 | PraecoxinA/ platycariin isomer (trigalloyl-HHDP-glucose) | C41H28O27 | [30] |
| 3 | 8.31,10.14, 11.12, 11.89, 12.75 | 785.0845 | 785.0840 | 633.07, 483.08, 300.99, 275.02 | Tellimagrandin I isomer (digalloyl-HHDP-glucose) | C34H26O22 | [30,31] |
| 4 | 8.96 | 633.0763 | 633.0720 | 463.05, 300.99 275.02 | Strictinin/isostrictinin isomers (galloyl-HHDP-glucose) | C27H22O18 | [30,31] |
| 5 | 9.45, 12.29, 14.16 | 935.0813 | 935.0786 | 783.07, 633.07, 300.99, 275.02 | Tellimagrandin I isomer (digalloyl-HHDP-glucose) | C34H26O22 | [30] |
| 6 | 9.79 | 301.0417 | 301.0347 | 151.04 | Quercetin | C15H10O7 | [30,31] |
| 7 | 10.52, 11.60, 12.96, 16.24 | 933.0655 | 933.0630 | 631.06, 481.06, 450.99, 300.99 | Glansrin C isomer | C41H26O26 | [30,31] |
| 8 | 10.67 | 907.0862 | 907.0837 | 783.07, 764.05, 481.06, 300.99, 275.02 | Heterophylliin E isomer | C40H28O25 | [30] |
| 9 | 10.86 | 469.0040 | 469.0049 | 425.01, 300.99, 166.99 | Valoneic acid dilactone/Flavogallonic acid dilactone isomer | C21H10O13 | [30] |
| 10 | 11.27 | 463.0512 | 463.0517 | 300.99 | Ellagic acid hexoside isomer | C20H16O13 | [30,31] |
| 11 | 11.47, 12.13 | 635.0882 | 635.0877 | 483.08, 465.07, 423.06, 313.06, 169.01 | Trigalloyl-glucose isomer | C27H24O18 | [30] |
| 12 | 12.55 | 1103.0873 | 1103.0850 | 1059.09, 935.07, 757.09, 633.07, 300.99 | Rugosin C/platycaryanin A/glansrin A isomer | C48H32O31 | [30] |
| 13 | 13.14, 14.02, 14.32 | 787.0996 | 787.0996 | 635.08, 465.07, 169.01 | Tetragalloyl-glucose | C34H28O22 | [30] |
| 14 | 13.39, 15.11 | 937.0956 | 937.0947 | 785.08, 635.08, 483.07, 300.99 | Tellimagrandin II/pterocaryanin C isomer | C41H30O26 | [30] |
| 15 | 13.42 | 433.0399 | 433.0405 | 300.99 | Ellagic acid pentoside isomer | C19H14O12 | [30,31] |
| 16 | 14.49 | 300.9989 | 300.9989 | 283.99, 257.01, 229.01, 185.02 | Ellagic acid | C14H6O8 | [30,31] |
| 17 | 15.49, 18.90 | 1085.0771 | 1085.0740 | 783.07, 633.07, 450.99, 300.99 | Eucalbanin A/cornusiin B isomer | C48H30O30 | [30] |
| 18 | 15.72 | 939.1120 | 939.1100 | 787.09, 769.09, 617.08 | 1,2,3,4,6-Pentagalloyl-glucose | C41H32O26 | [30] |
| 19 | 17.03 | 433.1127 | 433.0772 | 300.03, 301.33 | Quercetin pentoside isomer | C20H18O11 | [30,31] |
| No. | tR (min) | Measured [M-H]− (m/z) | Predicted [M-H]− (m/z) | MS/MS Fragments (m/z) | Identification | Molecular Formula | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 9.79 | 301.0417 | 301.0347 | 150.00 | Quercetin | C15H10O7 | [30,31] |
| 2 | 11.27 | 463.0512 | 463.0517 | 300.99 | Ellagic acid hexoside isomer | C20H16O13 | [30,31] |
| 3 | 13.42 | 433.0396 | 433.0405 | 300.99 | Ellagic acid pentoside isomer | C19H14O12 | [30,31] |
| 4 | 14.49 | 300.9990 | 300.9990 | 283.99, 257.01, 229.01 | Ellagic acid | C14H6O8 | [30,31] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Wan, Y.; Wang, Y.; Zhao, Y.; Zhang, Y.; Zhang, A.; Weng, Q.; Xu, M. Walnut Polyphenol Extract Protects against Fenitrothion-Induced Immunotoxicity in Murine Splenic Lymphocytes. Nutrients 2018, 10, 1838. https://doi.org/10.3390/nu10121838
Liu H, Wan Y, Wang Y, Zhao Y, Zhang Y, Zhang A, Weng Q, Xu M. Walnut Polyphenol Extract Protects against Fenitrothion-Induced Immunotoxicity in Murine Splenic Lymphocytes. Nutrients. 2018; 10(12):1838. https://doi.org/10.3390/nu10121838
Chicago/Turabian StyleLiu, Hong, Yifang Wan, Yuxin Wang, Yue Zhao, Yue Zhang, Ao Zhang, Qiang Weng, and Meiyu Xu. 2018. "Walnut Polyphenol Extract Protects against Fenitrothion-Induced Immunotoxicity in Murine Splenic Lymphocytes" Nutrients 10, no. 12: 1838. https://doi.org/10.3390/nu10121838