
The Role of miR-217-5p in the Puromycin Aminonucleoside-Induced Morphological Change of Podocytes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Induction of Podocyte Injury
2.2. Comprehensive Small RNA Sequencing
2.3. Real-Time Reverse-Transcription PCR for miRNA and mRNA
2.4. miRNA Overexpression in E11 Podocytes
2.5. Cell Morphology Observation and Fluorescent Immunocytochemistry
2.6. Cell Viability Assay
2.7. Western Blotting
2.8. Reporter Plasmid Construction and Luciferase Assay
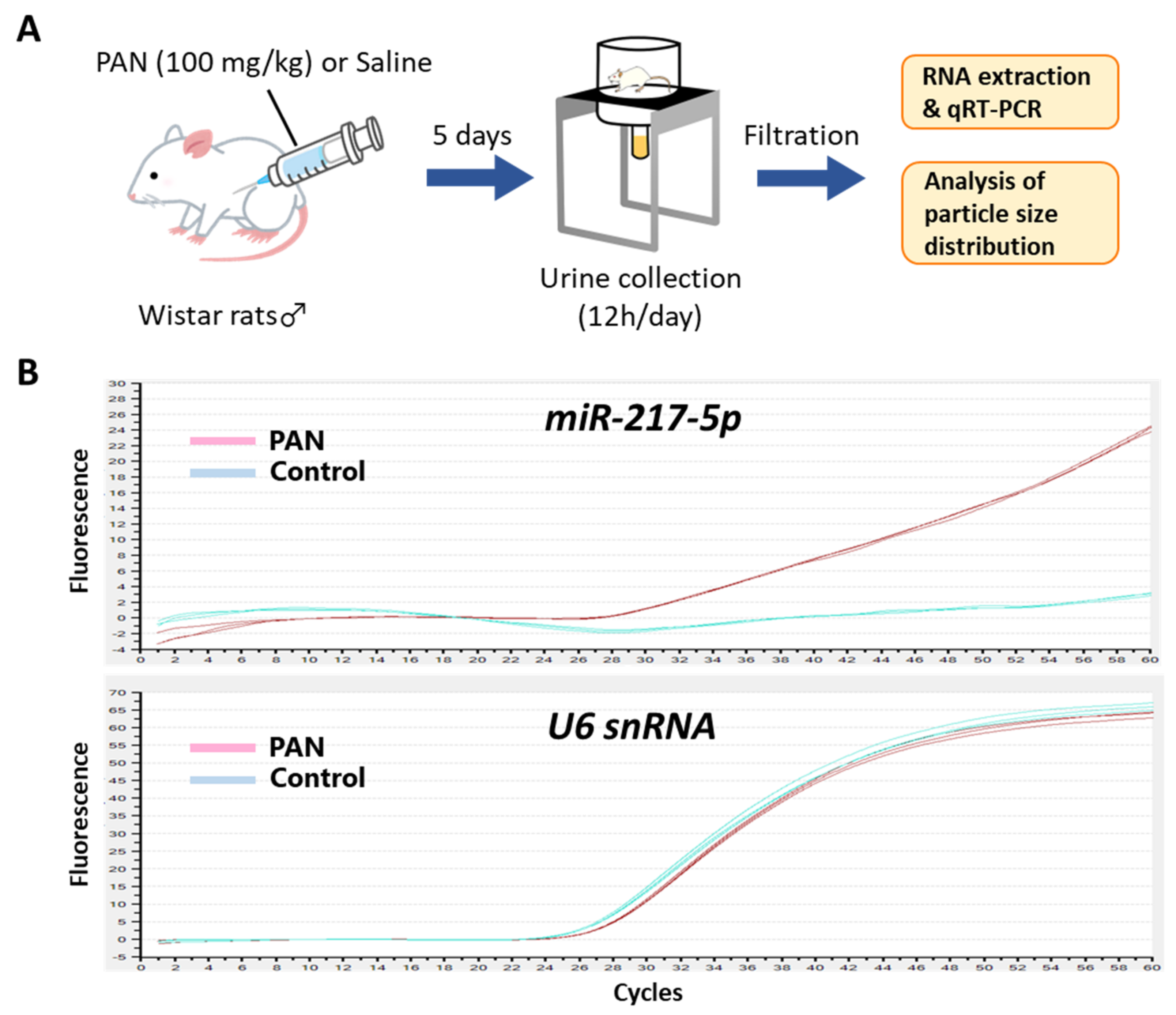
2.9. Urine Processing
2.10. Bioinformatics
2.11. Statistical Analysis
3. Results
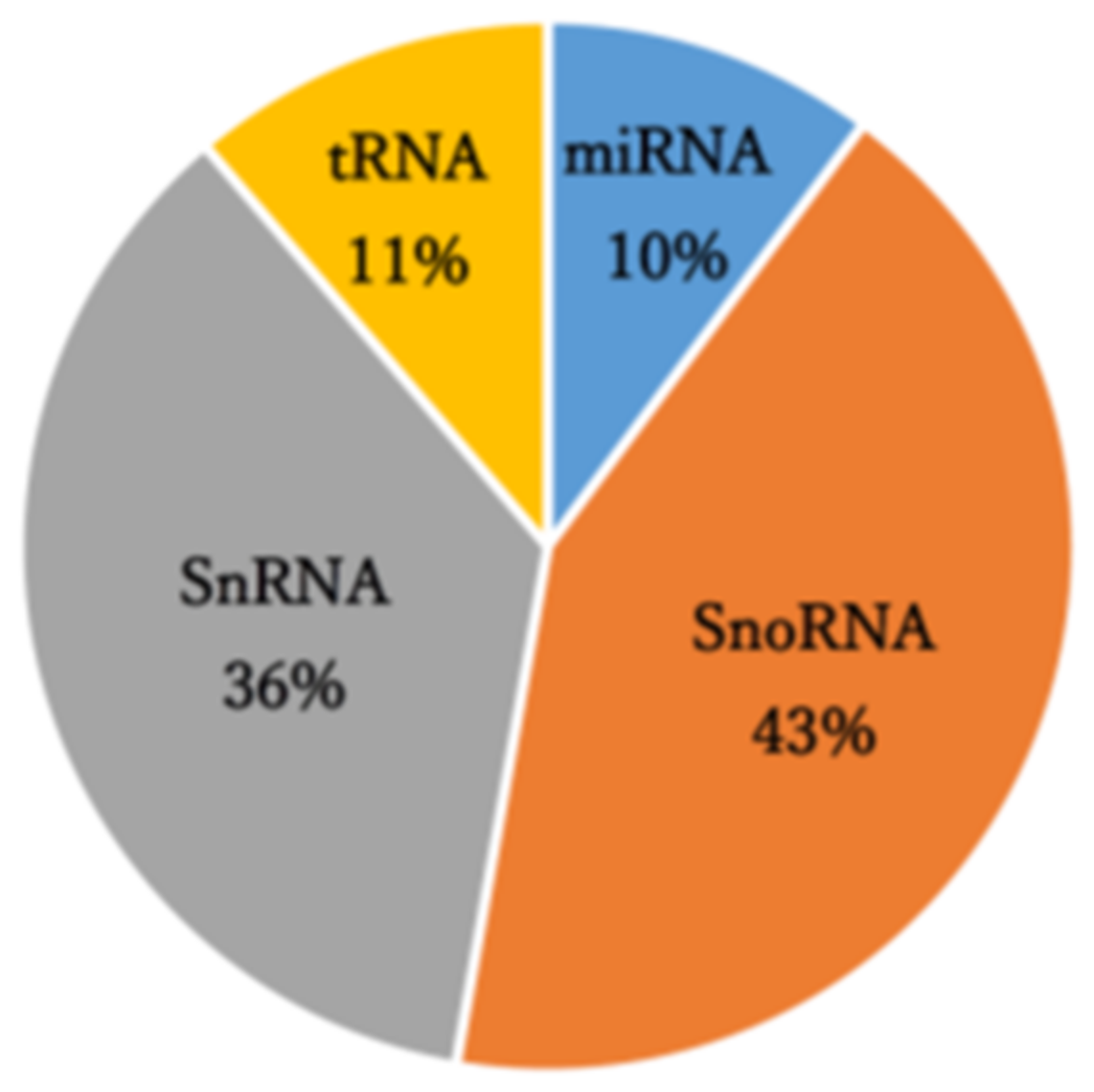
3.1. Identification of miRNAs Dysregulated by PAN-Induced Podocyte Injury
3.2. Validation of PAN-Induced miR-217-5p Expression in Primary Rat Podocytes
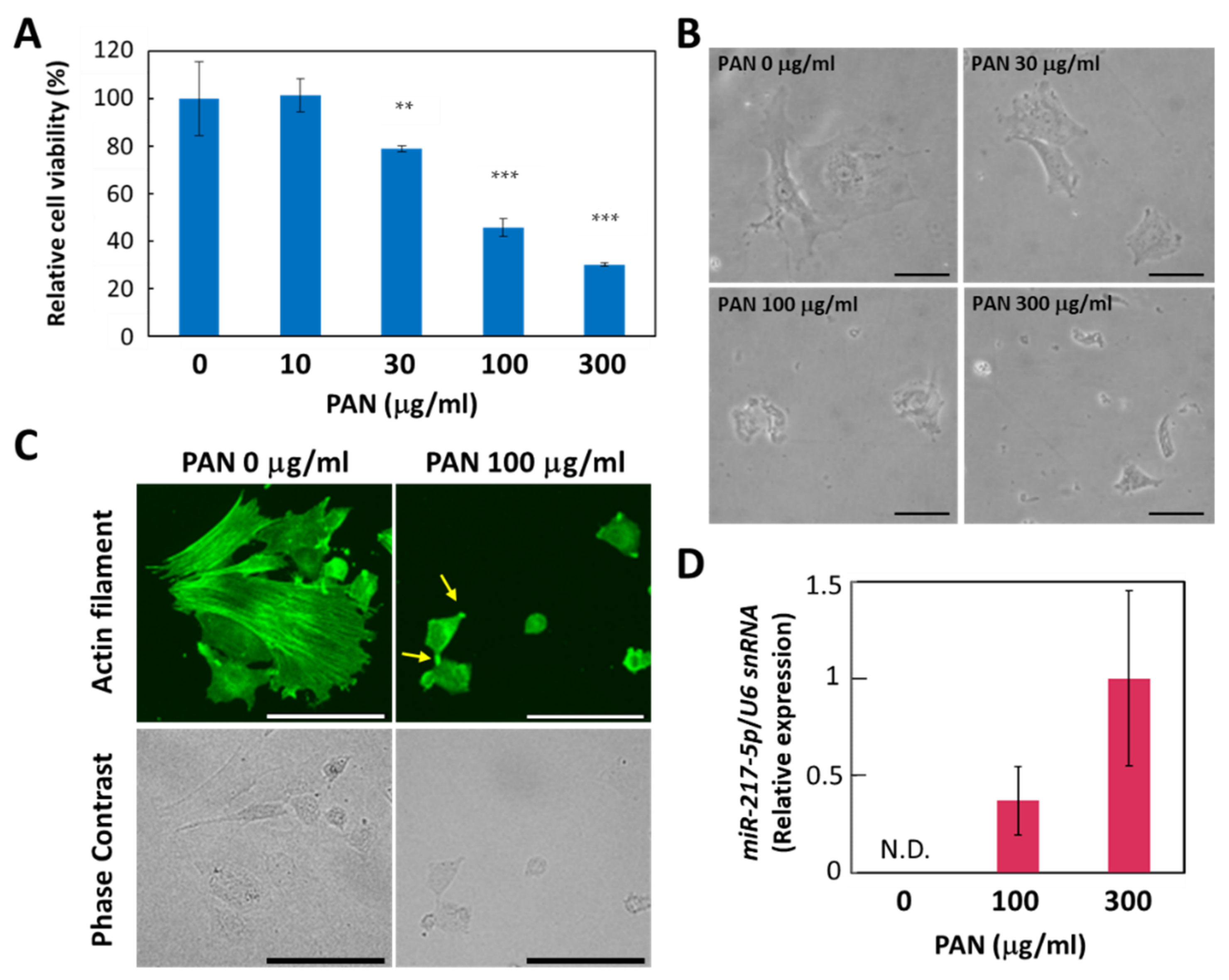
3.3. PAN-Induced Effects on Cell Viability, Cell Morphology, and miR-217-5p Expression in E11 Podocyte Cells
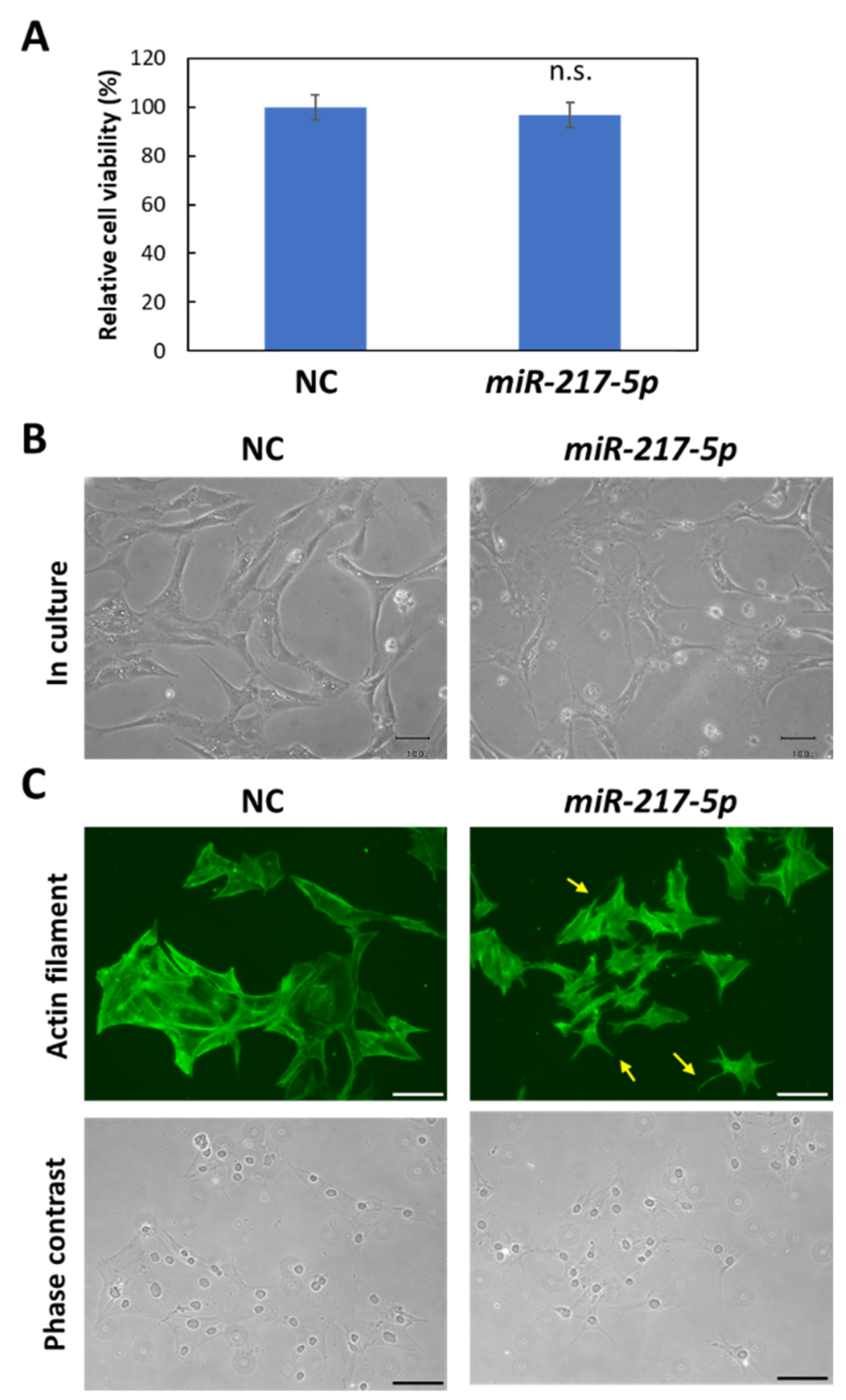
3.4. MiR-217-5p-Induced Effects on Cell Viability and Cell Morphology in E11 Cells
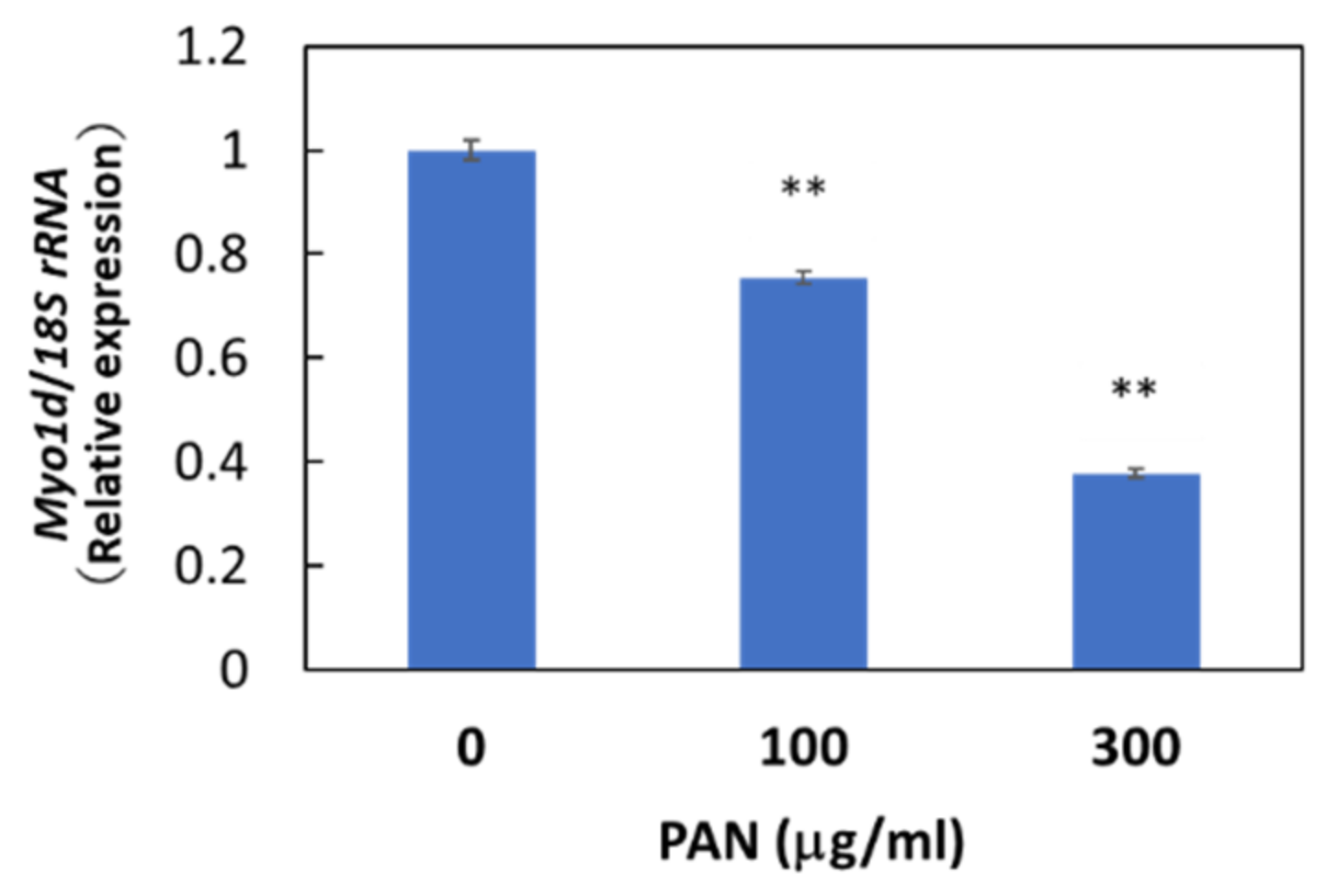
3.5. Integrative Analysis of miRNA and mRNA Expression in PAN-Treated Podocytes
3.6. Gene Ontologies Associated with the Predicted Targets of the PAN-Dysregulated miRNAs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Fold Change | Representative GO (Biological Process) Annotations |
|---|---|---|
| Pard3b | 0.303 | cell adhesion, cell cycle, cell division, etc. |
| Fndc3b | 0.407 | integral component of membrane |
| Myo1d | 0.430 | actin filament organization, cellular localization, etc. |
| Akap2 | 0.493 | actin filament organization, protein localization, etc. |
| Peak1 | 0.532 | cell migration, focal adhesion assembly, etc. |
| Srgap1 | 0.551 | negative regulation of cell migration, etc. |
| Fn1 | 0.573 | acute-phase response, acute-phase response, etc. |
| Picalm | 0.604 | amyloid-beta clearance by transcytosis, axonogenesis, etc. |
| Meis1 | 0.610 | Angiogenesis, animal organ morphogenesis, etc. |
| Epha7 | 0.620 | Angiogenesis, animal organ morphogenesis, etc. |
| Crls1 | 0.716 | lens morphogenesis in camera-type eye, etc. |
| Rock1 | 0.733 | regulation of cell population proliferation, etc. |
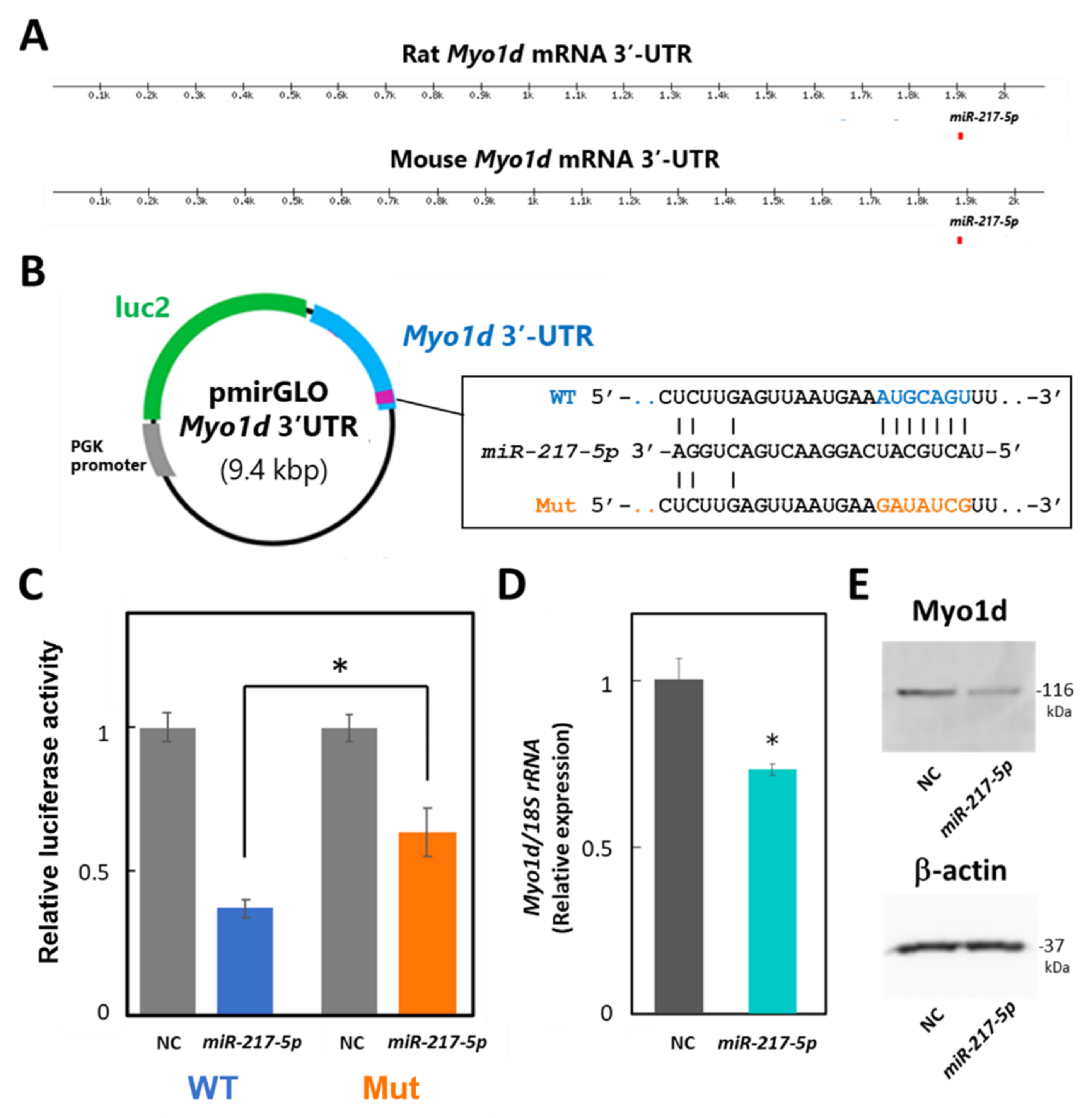
3.7. Target Validation of miR-217-5p
3.8. Detection of miR-217-5p in Urine from PAN-Administrated Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Kidney Day: Chronic Kidney Disease. 2015. Available online: http://www.worldkidneyday.org/faqs/chronic-kidney-disease/ (accessed on 6 June 2022).
- Whaley-Connell, A.; Nistala, R.; Chaudhary, K. The importance of early identification of chronic kidney disease. Mo. Med. 2011, 108, 25–28. [Google Scholar] [PubMed]
- Darshi, M.; Van Espen, B.; Sharma, K. Metabolomics in Diabetic Kidney Disease: Unraveling the Biochemistry of a Silent Killer. Am. J. Nephrol. 2016, 44, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.R.; Aeddula, N.R. Chronic Renal Failure. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Zhong, J.; Yang, H.C.; Fogo, A.B. A perspective on chronic kidney disease progression. Am. J. Physiol. Renal. Physiol. 2017, 312, F375–F384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turin, T.C.; Tonelli, M.; Manns, B.J.; Ravani, P.; Ahmed, S.B.; Hemmelgarn, B.R. Chronic kidney disease and life expectancy. Nephrol. Dial. Transplant. 2012, 27, 3182–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J.K.; Li, P.K. Chronic kidney disease epidemic: How do we deal with it? Nephrology (Carlton) 2018, 23 (Suppl. S4), 116–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whalen, H.; Clancy, M.; Jardine, A. Future challenges in renal transplantation. Minerva Chir. 2012, 67, 15–30. [Google Scholar] [PubMed]
- Neipp, M.; Jackobs, S.; Klempnauer, J. Renal transplantation today. Langenbecks Arch. Surg. 2009, 394, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Coates, P.T.; Wong, G. Current controversies in nephrology—how to crossmatch for transplantation? Kidney Int. 2020, 97, 662–663. [Google Scholar] [CrossRef]
- Sandip, K. Challenges & Controversies in Kidney Transplantation, 1st ed.; JAYPEE Digital: New Delhi, India, 2015; pp. 1–172. [Google Scholar]
- Shrestha, B.M. Strategies for reducing the renal transplant waiting list: A review. Exp. Clin. Transplant. 2009, 7, 173–179. [Google Scholar]
- Voora, S.; Adey, D.B. Management of Kidney Transplant Recipients by General Nephrologists: Core Curriculum 2019. Am. J. Kidney. Dis. 2019, 73, 866–879. [Google Scholar] [CrossRef]
- Murray, I.; Paolini, M.A. Histology, Kidney and Glomerulus. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Liapis, H.; Romagnani, P.; Anders, H.J. New insights into the pathology of podocyte loss: Mitotic catastrophe. Am. J. Pathol. 2013, 183, 1364–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, P. A Review of Podocyte Biology. Am. J. Nephrol. 2018, 47 (Suppl. S1), 3–13. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M. Podocyte injury and its consequences. Kidney Int. 2016, 89, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Wakiyama, M.; Takimoto, K.; Ohara, O.; Yokoyama, S. Let-7 microRNAmediated mRNA deadenylation and translational repression in a mammalian cell-free system. Genes Dev. 2007, 21, 1857–1862. [Google Scholar] [CrossRef] [Green Version]
- Meister, G.; Tuschl, T. Mechanisms of gene silencing by double-stranded RNA. Nature 2004, 431, 343–349. [Google Scholar] [CrossRef]
- Shi, S.; Yu, L.; Chiu, C.; Sun, Y.; Chen, J.; Khitrov, G.; Merkenschlager, M.; Holzman, L.B.; Zhang, W.; Mundel, P.; et al. Podocyte-selective deletion of dicer induces proteinuria and glomerulosclerosis. J. Am. Soc. Nephrol. 2008, 19, 2159–2169. [Google Scholar] [CrossRef] [Green Version]
- Harvey, S.J.; Jarad, G.; Cunningham, J.; Goldberg, S.; Schermer, B.; Harfe, B.D.; McManus, M.T.; Benzing, T.; Miner, J.H. Podocyte-specific deletion of dicer alters cytoskeletal dynamics and causes glomerular disease. J. Am. Soc. Nephrol. 2008, 19, 2150–2158. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Ng, K.H.; Rosen, S.; Dostal, A.; Gregory, R.I.; Kreidberg, J.A. Podocyte-specific loss of functional microRNAs leads to rapid glomerular and tubular injury. J. Am. Soc. Nephrol. 2008, 19, 2069–2075. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Kaneko, S.; Yanai, K.; Aomatsu, A.; Hirai, K.; Ookawara, S.; Ishibashi, K.; Morishita, Y. MicroRNAs in Podocyte Injury in Diabetic Nephropathy. Front. Genet. 2020, 11, 993. [Google Scholar] [CrossRef]
- Li, J.Y.; Yong, T.Y.; Michael, M.Z.; Gleadle, J.M. Review: The role of microRNAs in kidney disease. Nephrology (Carlton) 2010, 15, 599–608. [Google Scholar] [CrossRef]
- Horikawa, A.; Yoneda, T.; Yaoita, E.; Yamaguchi, K.; Shigenobu, S.; Kuramochi, M.; Yamate, J.; Inui, T.; Ishibashi, O. A novel splicing variant of small nucleolar RNA host gene 4 is a podocyte-selective non-coding RNA upregulated in response to puromycin aminonucleoside-induced podocyte injury. J. Biochem. 2019, 165, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Katsuya, K.; Yaoita, E.; Yoshida, Y.; Yamamoto, Y.; Yamamoto, T. An improved method for primary culture of rat podocytes. Kidney Int. 2006, 69, 2101–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, T.; Yamada, E.; Okada, S.; Shimoda, Y.; Tagaya, Y.; Hashimoto, K.; Satoh, T.; Mori, M.; Okada, J.; Pessin, J.E.; et al. Nucleobindin-2 is a positive regulator for insulin-stimulated glucose transporter 4 translocation in fenofibrate treated E11 podocytes. Endocr. J. 2014, 61, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, O.; Akagi, I.; Ogawa, Y.; Inui, T. MiR-141-3p is upregulated in esophageal squamous cell carcinoma and targets pleckstrin homology domain leucine-rich repeat protein phosphatase-2, a negative regulator of the PI3K/AKT pathway. Biochem. Biophys. Res. Commun. 2018, 501, 507–513. [Google Scholar] [CrossRef]
- Pomaznoy, M.; Ha, B.; Peters, B. GOnet: A tool for interactive Gene Ontology analysis. BMC Bioinform. 2018, 19, 470. [Google Scholar] [CrossRef]
- Thilo, F.; Liu, Y.; Loddenkemper, C.; Schuelein, R.; Schmidt, A.; Yan, Z.; Zhu, Z.; Zakrzewicz, A.; Gollasch, M.; Tepel, M. VEGF regulates TRPC6 channels in podocytes. Nephrol. Dial. Transplant. 2012, 27, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Nosaka, K.; Takahashi, T.; Nishi, T.; Imaki, H.; Suzuki, T.; Suzuki, K.; Kurokawa, K.; Endou, H. An adenosine deaminase inhibitor prevents puromycin aminonucleoside nephrotoxicity. Free Radic. Biol. Med. 1997, 22, 597–605. [Google Scholar] [CrossRef]
- Xia, L.; Zhou, M.; Kalhorn, T.F.; Ho, H.T.; Wang, J. Podocyte-specific expression of organic cation transporter PMAT: Implication in puromycin aminonucleoside nephrotoxicity. Am. J. Physiol. Renal. Physiol. 2009, 296, F1307–F1313. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [Green Version]
- Noris, M.; Remuzzi, G. Non-muscle myosins and the podocyte. Clin. Kidney J. 2012, 5, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solanki, A.K.; Srivastava, P.; Rahman, B.; Lipschutz, J.H.; Nihalani, D.; Arif, E. The Use of High-throughput transcriptomics to identify pathways with therapeutic significance in podocytes. Int. J. Mol. Sci. 2019, 21, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okabe, M.; Motojima, M.; Miyazaki, Y.; Pastan, I.; Yokoo, T.; Matsusaka, T. Global polysome analysis of normal and injured podocytes. Am. J. Physiol. Renal. Physiol. 2019, 316, F241–F252. [Google Scholar] [CrossRef]
- Erdos, Z.; Barnum, J.E.; Wang, E.; DeMaula, C.; Dey, P.M.; Forest, T.; Bailey, W.J.; Glaab, W.E. Evaluation of the Relative Performance of Pancreas-Specific MicroRNAs in Rat Plasma as Biomarkers of Pancreas Injury. Toxicol. Sci. 2020, 173, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Rouse, R. Co-sequencing and novel delayed anti-correlation identify function for pancreatic enriched microRNA biomarkers in a rat model of acute pancreatic injury. BMC Genom. 2018, 19, 297. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, W.; Thibault, S.; Brown, T.P.; Bobrowski, W.; Gukasyan, H.J.; Evering, W.; Hu, W.; John-Baptiste, A.; Vitsky, A. Evaluation of miR-216a and miR-217 as Potential Biomarkers of Acute Exocrine Pancreatic Toxicity in Rats. Toxicol. Pathol. 2017, 45, 321–334. [Google Scholar] [CrossRef]
- Rouse, R.; Rosenzweig, B.; Shea, K.; Knapton, A.; Stewart, S.; Xu, L.; Chockalingam, A.; Zadrozny, L.; Thompson, K. MicroRNA biomarkers of pancreatic injury in a canine model. Exp. Toxicol. Pathol. 2017, 69, 33–43. [Google Scholar] [CrossRef]
- Calvano, J.; Edwards, G.; Hixson, C.; Burr, H.; Mangipudy, R.; Tirmenstein, M. Serum microRNAs-217 and -375 as biomarkers of acute pancreatic injury in rats. Toxicology 2016, 368–369, 1–9. [Google Scholar] [CrossRef]
- Goodwin, D.; Rosenzweig, B.; Zhang, J.; Xu, L.; Stewart, S.; Thompson, K.; Rouse, R. Evaluation of miR-216a and miR-217 as potential biomarkers of acute pancreatic injury in rats and mice. Biomarkers 2014, 19, 517–529. [Google Scholar] [CrossRef]
- Rachagani, S.; Macha, M.A.; Menning, M.S.; Dey, P.; Pai, P.; Smith, L.M.; Mo, Y.Y.; Batra, S.K. Changes in microRNA (miRNA) expression during pancreatic cancer development and progression in a genetically engineered KrasG12D; Pdx1-Cre mouse (KC) model. Oncotarget 2015, 6, 40295–40309. [Google Scholar] [CrossRef] [Green Version]
- Egeli, U.; Tezcan, G.; Cecener, G.; Tunca, B.; Demirdogen Sevinc, E.; Kaya, E.; Ak, S.; Dundar, H.Z.; Sarkut, P.; Ugras, N.; et al. Pancreas. miR-216b Targets FGFR1 and Confers Sensitivity to Radiotherapy in Pancreatic Ductal Adenocarcinoma Patients Without EGFR or KRAS Mutation. Pancreas 2016, 45, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Safaralizadeh, R.; Ajami, N.; Nemati, M.; Hosseinpourfeizi, M.; Azimzadeh Isfanjani, A.; Moaddab, S.Y. Disregulation of miR-216a and miR-217 in Gastric Cancer and Their Clinical Significance. J. Gastrointest. Cancer 2019, 50, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Z.P.; Zhang, R.Q.; Zhang, H.M. Repression of miR-217 protects against high glucose-induced podocyte injury and insulin resistance by restoring PTEN-mediated autophagy pathway. Biochem. Biophys. Res. Commun. 2017, 483, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.W.; Pan, M.; Ye, H.Y.; Zheng, Y.; Chen, Y.; Huang, W.W.; Xu, X.Y.; Zheng, S.B. Down-regulation of the long non-coding RNA XIST ameliorates podocyte apoptosis in membranous nephropathy via the miR-217-TLR4 pathway. Exp. Physiol. 2019, 104, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, N. Mechanism of the process formation; podocytes vs. neurons. Microsc. Res. Tech. 2002, 57, 217–223. [Google Scholar] [CrossRef]
- Tapia, R.; Guan, F.; Gershin, I.; Teichman, J.; Villegas, G.; Tufro, A. Semaphorin3a disrupts podocyte foot processes causing acute proteinuria. Kidney Int. 2008, 73, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Li, Q.; Pisarek-Horowitz, A.; Rasouly, H.M.; Wang, X.; Bonegio, R.G.; Wang, H.; McLaughlin, M.; Mangos, S.; Kalluri, R.; et al. Inhibitory effects of Robo2 on nephrin: A crosstalk between positive and negative signals regulating podocyte structure. Cell Rep. 2012, 2, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Arif, E.; Wagner, M.C.; Johnstone, D.B.; Wong, H.N.; George, B.; Pruthi, P.A.; Lazzara, M.J.; Nihalani, D. Motor protein Myo1c is a podocyte protein that facilitates the transport of slit diaphragm protein Neph1 to the podocyte membrane. Mol. Cell Biol. 2011, 31, 2134–2150. [Google Scholar] [CrossRef] [Green Version]
- Benesh, A.E.; Fleming, J.T.; Chiang, C.; Carter, B.D.; Tyska, M.J. Expression and localization of myosin-1d in the developing nervous system. Brain Res. 2012, 1440, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Szeto, C.C. Urinary podocyte markers in kidney diseases. Clin. Chim. Acta 2021, 523, 315–324. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| miRNA | Fold Change | Regulation |
|---|---|---|
| rno-miR-217-5p | 5.733 | Upregulated |
| rno-miR-216a-5p | 4.641 | Upregulated |
| rno-miR-338-3p | 2.295 | Upregulated |
| rno-miR-3583-5p | 2,125 | Upregulated |
| rno-miR-3572 | 0.3526 | Downregulated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishibashi, O.; Hayashi, M.; Horikawa, A.; Owada, H.; Miyamoto, R.; Mizukami, N.; Inui, T. The Role of miR-217-5p in the Puromycin Aminonucleoside-Induced Morphological Change of Podocytes. Non-Coding RNA 2022, 8, 43. https://doi.org/10.3390/ncrna8030043
Ishibashi O, Hayashi M, Horikawa A, Owada H, Miyamoto R, Mizukami N, Inui T. The Role of miR-217-5p in the Puromycin Aminonucleoside-Induced Morphological Change of Podocytes. Non-Coding RNA. 2022; 8(3):43. https://doi.org/10.3390/ncrna8030043
Chicago/Turabian StyleIshibashi, Osamu, Mika Hayashi, Aya Horikawa, Hitoshi Owada, Ryotaro Miyamoto, Naoya Mizukami, and Takashi Inui. 2022. "The Role of miR-217-5p in the Puromycin Aminonucleoside-Induced Morphological Change of Podocytes" Non-Coding RNA 8, no. 3: 43. https://doi.org/10.3390/ncrna8030043