
Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials and Chitosan Nanoparticles (CH-NPs)
2.2. Chilling Treatments
2.3. Plant Samples Analysis
2.4. Data Analysis
3. Results
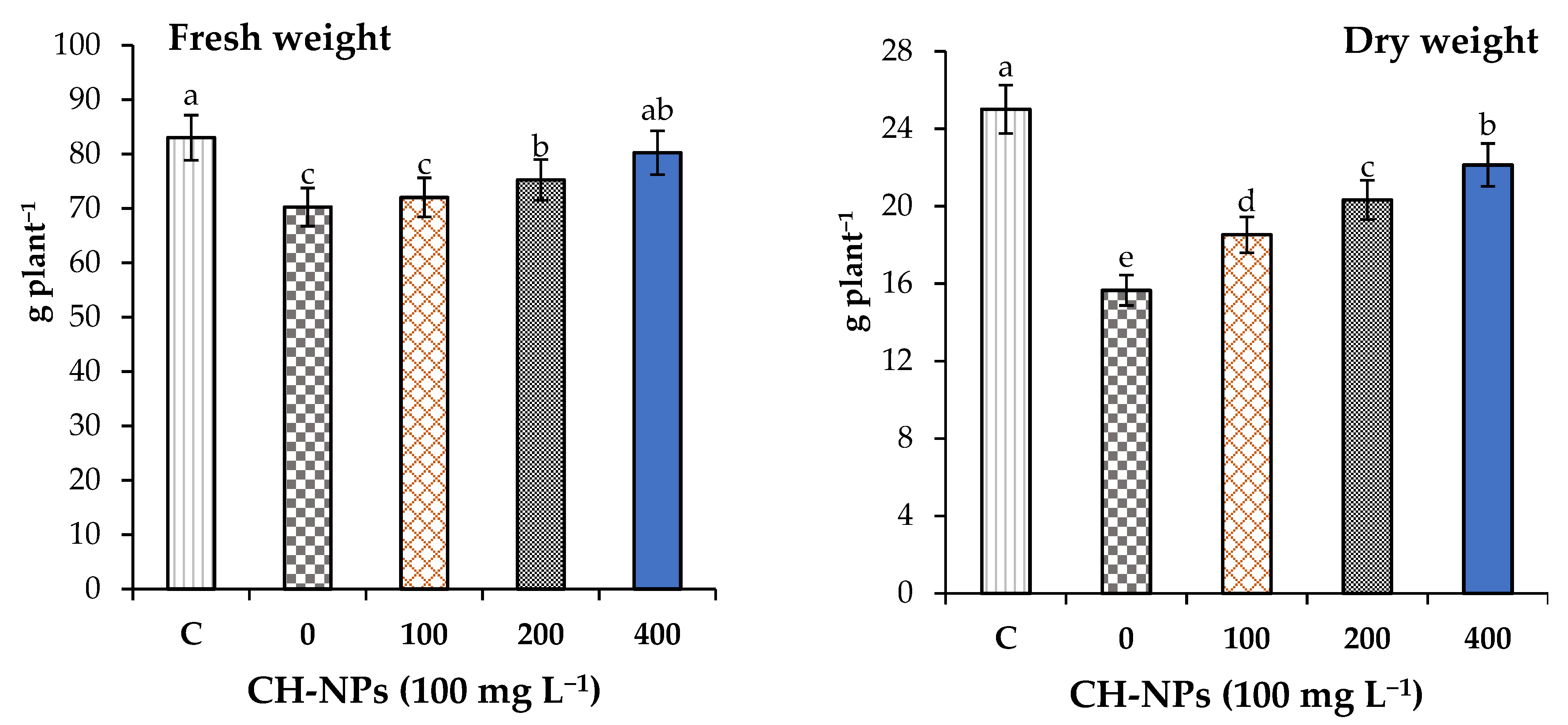
3.1. Effect of CH-NPs on the Growth and Nutrient Content of Banana Plants
3.2. Effect of CH-NPs on the Secondary and Osmo-Metabolic Compounds
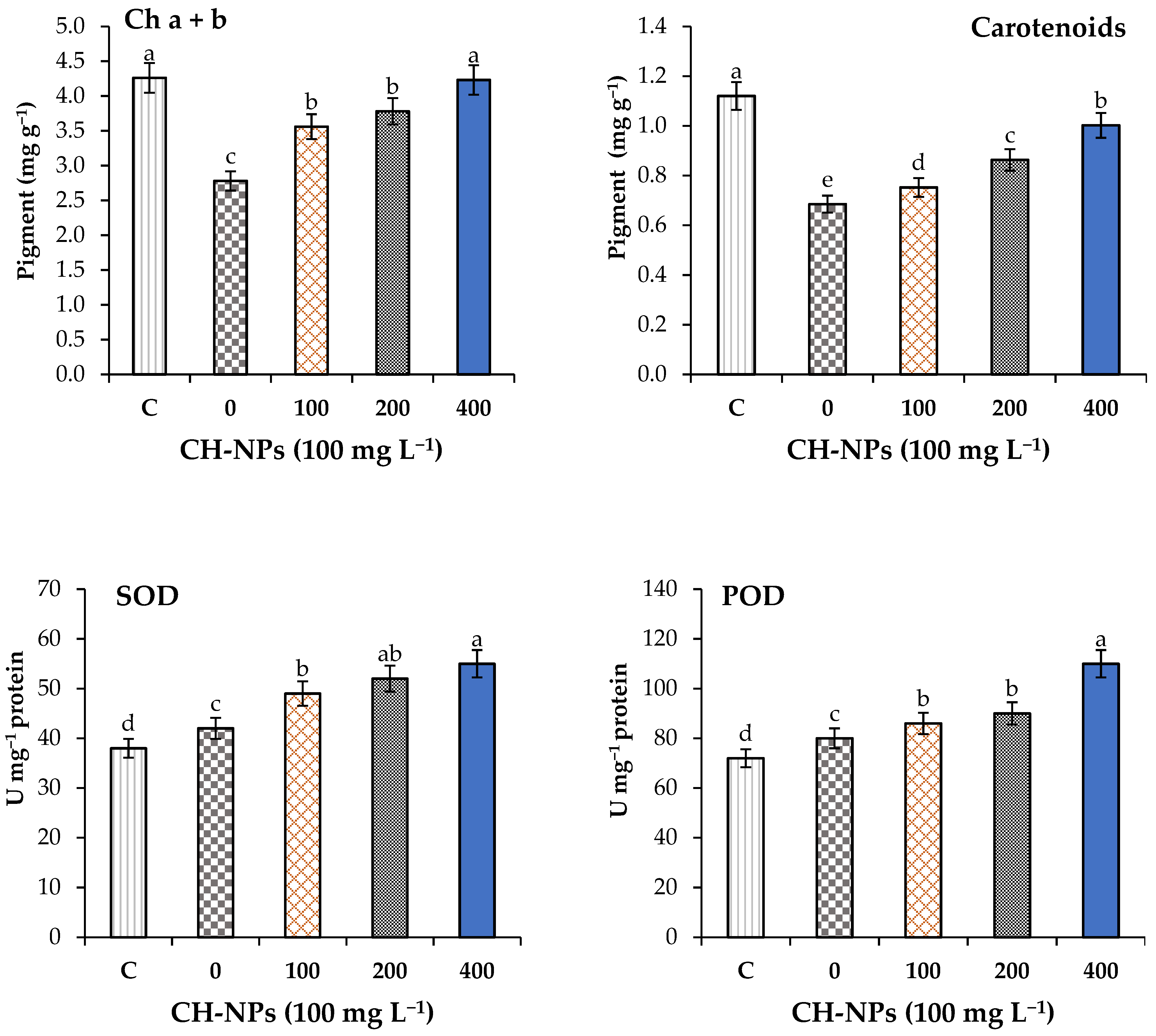
3.3. Effect of CH-NPs on the Photosynthesis Pigments and Antioxidant Enzyme Activities
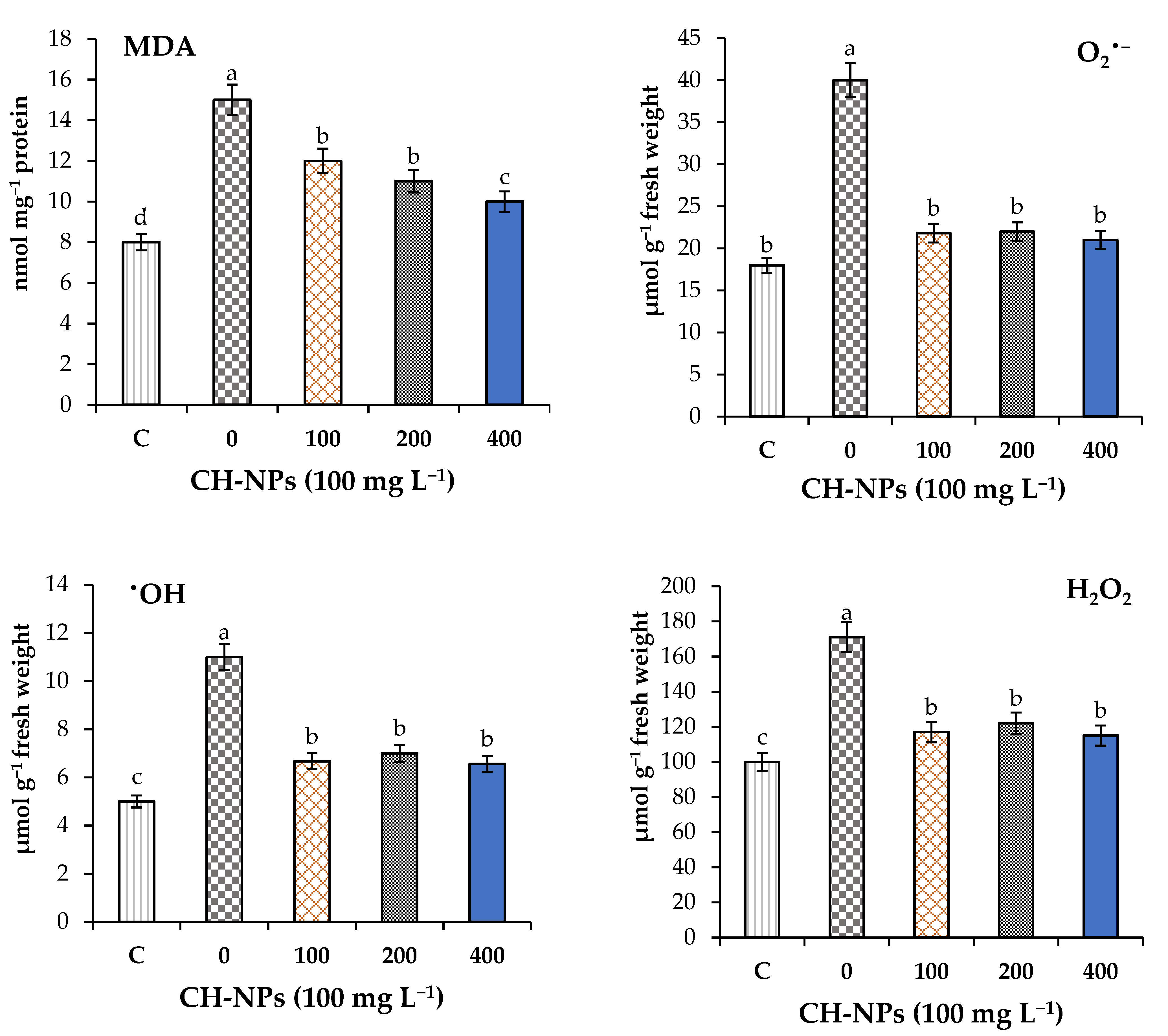
3.4. Effects of CH-NPs on the Stress Markers and Membrane Damage Traits
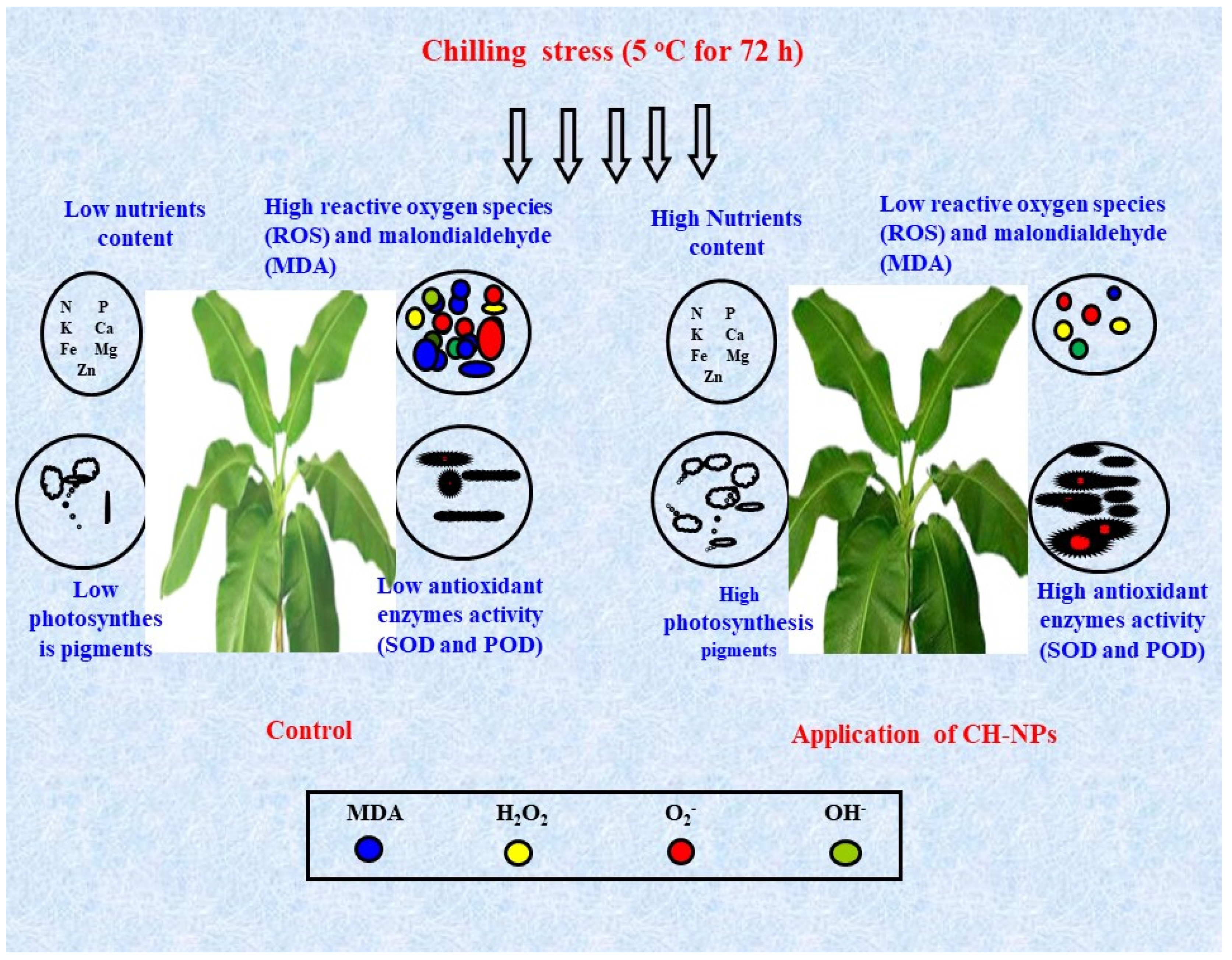
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Robinson, J.C.; Saúco, V.G. Bananas and Plantains; CABI: Wallingford, UK, 2010; Volume 19. [Google Scholar]
- Manchanda, P.; Gill, M.I.S.; Megha, S.; Gosal, S.S. Production of superelite planting material through in vitro culturing in banana. In Biotechnologies of Crop Improvement; Gosal, S., Wani, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 1, pp. 93–129. [Google Scholar]
- Pozza, L.E.; Field, D.J. The science of soil security and food security. Soil Secur. 2020, 1, 100002. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, J.Z.; Chow, W.S.; Sun, L.L.; Chen, J.W.; Chen, Y.J.; Peng, C.L. The influence of low temperature on photosynthesis and antioxidant enzymes in sensitive banana and tolerant plantain (Musa sp.) cultivars. Photosynthetica 2011, 49, 201–208. [Google Scholar] [CrossRef]
- He, W.-D.; Gao, J.; Dou, T.-X.; Shao, X.-H.; Bi, F.-C.; Sheng, O.; Deng, G.-M.; Li, C.-Y.; Hu, C.-H.; Liu, J.-H.; et al. Early cold-induced peroxidases and aquaporins are associated with high cold tolerance in Dajiao (Musa spp. ‘Dajiao’). Front. Plant Sci. 2018, 9, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mahdy, M.T.; Youssef, M.; Eissa, M.A. Impact of in vitro cold stress on two banana genotypes based on physio-biochemical Evaluation. S. Afr. J. Bot. 2018, 119, 219–225. [Google Scholar] [CrossRef]
- Rodríguez-Zapata, L.C.; Gil, F.L.E.Y.; Cruz-Martínez, S.; Talavera-May, C.R.; Contreras-Marin, F.; Fuentes, G.; Sauri-Duch, E.; Santamaría, J.M. Preharvest foliar applications of glycine-betaine protects banana fruits from chilling injury during the postharvest stage. Chem. Biol. Technol. Agric. 2015, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-L.; Shan, W.; Cai, D.-L.; Chen, J.-Y.; Lu, W.-J.; Su, X.-G.; Kuang, J.-F. Postharvest application of glycine betaine ameliorates chilling injury in cold-stored banana fruit by enhancing antioxidant system. Sci. Hortic. 2021, 287, 110264. [Google Scholar] [CrossRef]
- Lima, J.D.; Rozane, D.E.; Gomes, E.N.; Da Silva, S.H.M.G.; Moraes, W.D.S.; Kluge, R.A. Chilling Prevention on Banana ‘Nanica’ in the Field with Bagging. J. Agric. Sci. 2018, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.R. (Ed.) Handbook of Diseases of Banana, Abacáand Enset; CAB International: Wallingford, UK, 2019. [Google Scholar]
- Levitt, J. Responses of plants to environmental stresses. In Chilling, Partly and High Temperature Stresses, 2nd ed.; Academic: New York, NY, USA, 1980; Volume 1, p. 49. [Google Scholar]
- Li, Y.; Zhang, Q.; Ou, L.; Ji, D.; Liu, T.; Lan, R.; Li, X.; Jin, L. Response to the cold stress signaling of the tea plant (Camellia sinensis) Elicited by Chitosan Oligosaccharide. Agronomy 2020, 10, 915. [Google Scholar] [CrossRef]
- Parisi, C.; Vigani, M.; Rodríguez-Cerezo, E. Agricultural nanotechnologies: What are the current possibilities? Nano Today 2015, 10, 124–127. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in Plant protection and fertilization: Current state, foreseen applications, and research priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef]
- Fraceto, L.F.; Grillo, R.; de Medeiros, G.A.; Scognamiglio, V.; Rea, G.; Ebartolucci, C. Nanotechnology in agriculture: Which innovation potential does it have? Front. Environ. Sci. 2016, 4, 20. [Google Scholar] [CrossRef]
- Cheng, G.; Chang, T.-H.; Qin, Q.; Huang, H.; Zhu, Y. Mechanical properties of silicon carbide nanowires: Effect of size-dependent defect density. Nano Lett. 2014, 14, 754–758. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Alkharabsheh, H.; Seleiman, M.; Battaglia, M.; Shami, A.; Jalal, R.; Alhammad, B.; Almutairi, K.; Al-Saif, A. Biochar and its broad impacts in soil quality and fertility, nutrient leaching and crop productivity: A review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-fertilization as an emerging fertilization technique: Why can modern agriculture benefit from its use? Plants 2020, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Al-Dhabaan, F.A.; Mostafa, M.; Almoammar, H.; Abd-Elsalam, K.A. Chitosan-based nanostructures in plant protection applications. In Nanotechnology Applications in Health and Environmental Sciences; Gabler: Wiesbaden, Germany, 2018; pp. 351–384. [Google Scholar]
- Shukla, S.K.; Mishra, A.K.; Arotiba, O.; Mamba, B. Chitosan-based nanomaterials: A state-of-the-art review. Int. J. Biol. Macromol. 2013, 59, 46–58. [Google Scholar] [CrossRef]
- Sen, S.K.; Chouhan, D.; Das, D.; Ghosh, R.; Mandal, P. Improvisation of salinity stress response in mung bean through solid matrix priming with normal and nano-sized chitosan. Int. J. Biol. Macromol. 2020, 145, 108–123. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, C.; Sun, X.; Zang, X.; Zhang, X.; Fang, T.; Xu, N. Comparative transcriptome analysis reveals chitooligosaccharides-induced stress tolerance of Gracilariopsis lemaneiformis under high temperature stress. Aquaculture 2020, 519, 734876. [Google Scholar] [CrossRef]
- Farghly, K.A.; Gomah, H.H.; Ahmed, M.M.; Abdalla, R.M.; Eissa, M.A. Corn wastes and peanut shell as growing media for production of red radish plants in soilless system. Commun. Soil Sci. Plant Anal. 2020, 51, 1799–1810. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4. 3.1–F4. 3.8. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, J.A.; Allen, S.E. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun. Soil Sci. Plant Anal. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Burt, R. Soil Survey Laboratory Methods Manual; Soil Survey Investigations Report No. 42; NRCS: Washington, DC, USA, 2004. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Kubiś, J. Exogenous spermidine differentially alters activities of some scavenging system enzymes, H2O2 and superoxide radical levels in water-stressed cucumber leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef]
- Rao, K.M.; Sresty, T. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. Te deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Abeed, A.H.; Eissa, M.A.; Abdel-Wahab, D.A. Effect of Exogenously applied jasmonic acid and kinetin on drought tolerance of wheat cultivars based on morpho-physiological evaluation. J. Soil Sci. Plant Nutr. 2021, 21, 131–144. [Google Scholar] [CrossRef]
- Eissa, M.A.; Nafady, M.; Ragheb, H.; Attia, K. Effect of soil moisture and forms of phosphorus fertilizers on corn production under sandy calcareous soil. World Appl. Sci. J. 2013, 26, 540–547. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.; Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Moreano, J.P.S.; Xu, X.; Criollo, C.B.A.; Chen, X.; Lin, Y.; Munir, N.; Lai, Z. Genome-wide identification and comprehensive analyses of TCP gene family in banana (Musa L.). Trop. Plant Biol. 2021, 14, 180–202. [Google Scholar] [CrossRef]
- Augspurger, C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Mishra, S.K.; Khan, M.H.; Misra, S.; Dixit, V.K.; Khare, P.; Srivastava, S.; Chauhan, P.S. Characterisation of Pseudomonas spp. and Ochrobactrum sp. isolated from volcanic soil. Antonie Van Leeuwenhoek 2017, 110, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Christie, P.; Zhang, S. Uptake, translocation, and transformation of metal-based nanoparticles in plants: Recent advances and methodological challenges. Environ. Sci. Nano 2019, 6, 41–59. [Google Scholar] [CrossRef]
- Saharan, V.; Pal, A. Current and Future Prospects of Chitosan-Based Nanomaterials in Plant Protection and Growth; Springer: Berlin/Heidelberg, Germany, 2016; pp. 43–48. [Google Scholar]
- Sathiyabama, M.; Manikandan, A. Foliar application of chitosan nanoparticle improves yield, mineral content and boost innate immunity in finger millet plants. Carbohydr. Polym. 2021, 258, 117691. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Rajput, V.; Al-Khuraif, A.; Arshad, M.; Minkina, T.; Sushkova, S.; Yu, F. Effect of foliar fertigation of chitosan nanoparticles on cadmium accumulation and toxicity in solanum lycopersicum. Biology 2021, 10, 666. [Google Scholar] [CrossRef] [PubMed]
- Tommasini, L.; Svensson, J.T.; Rodriguez, E.M.; Wahid, A.; Malatrasi, M.; Kato, K.; Wanamaker, S.; Resnik, J.; Close, T.J. Dehydrin gene expression provides an indicator of low temperature and drought stress: Transcriptome-based analysis of Barley (Hordeum vulgare L.). Funct. Integr. Genom. 2008, 8, 387–405. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Duan, Z.; Mao, J.; Li, X.; Dong, F. Effects of root-zone temperature and N, P, and K supplies on nutrient uptake of cucumber (Cucumis sativus L.) seedlings in hydroponics. Soil Sci. Plant Nutr. 2012, 58, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Aziz, T.; Wahid, A.; Lee, D.-J.; Siddique, K. Chilling tolerance in maize: Agronomic and physiological approaches. Crop. Pasture Sci. 2009, 60, 501–516. [Google Scholar] [CrossRef]
- Dzung, N.A.; Khanh, V.T.P.; Dzung, T.T. Research on impact of chitosan oligomers on biophysical characteristics, growth, development and drought resistance of coffee. Carbohydr. Polym. 2011, 84, 751–755. [Google Scholar] [CrossRef]
- Al-Huqail, A.A. Changes in antioxidant status, water relations and physiological indices of maize seedlings under drought stress conditions. J. Biol. Sci. 2019, 19, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Eissa, M.A.; Abeed, A.H. Growth and biochemical changes in quail bush (Atriplex lentiformis (Torr.) S.Wats) under Cd stress. Environ. Sci. Pollut. Res. 2019, 26, 628–635. [Google Scholar] [CrossRef]
- Abou-Zaid, E.A.; Eissa, M.A. Thompson seedless grapevines growth and quality as affected by glutamic acid, vitamin b, and algae. J. Soil Sci. Plant Nutr. 2019, 19, 725–733. [Google Scholar] [CrossRef]
- Ali, A.M.; Awad, M.Y.; Hegab, S.A.; Gawad, A.M.A.E.; Eissa, M.A. Effect of potassium solubilizing bacteria (Bacillus cereus) on growth and yield of potato. J. Plant Nutr. 2021, 44, 411–420. [Google Scholar] [CrossRef]
- Al-Sayed, H.; Hegab, S.A.; Youssef, M.; Khalafalla, M.; Almaroai, Y.A.; Ding, Z.; Eissa, M.A. Evaluation of quality and growth of roselle (Hibiscus sabdariffa L.) as affected by bio-fertilizers. J. Plant Nutr. 2021, 43, 1025–1035. [Google Scholar] [CrossRef]
- Ding, Z.; Zhou, Z.; Lin, X.; Zhao, F.; Wang, B.; Lin, F.; Ge, Y.; Eissa, M.A. Biochar impacts on NH3-volatilization kinetics and growth of sweet basil (Ocimum basilicum L.) under saline conditions. Ind. Crop. Prod. 2020, 157, 11290. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CH-NPs mg L−1 | N | P | K | Ca | Fe | Mn | Zn | Cu |
|---|---|---|---|---|---|---|---|---|
| (g kg−1) | (g kg−1) | (g kg−1) | (g kg−1) | (mg kg−1) | (mg kg−1) | (mg kg−1) | (mg kg−1) | |
| C | 42 ± 4 a | 2.8 ± 0.2 a | 30 ± 2 a | 23 ± 2 a | 215 ± 12 a | 152 ± 5 a | 133 ± 6 a | 57 ± 3 a |
| 0 | 32 ± 2 d | 2.5 ± 0.1 b | 18 ± 1 d | 15 ± 1 c | 200 ± 4 c | 150 ± 7 a | 128 ± 5 a | 56 ± 3 a |
| 100 | 36 ± 3 c | 2.8 ± 0.2 a | 23 ± 2 c | 19 ± 1 b | 208 ± 5 b | 154 ± 6 a | 134 ± 7 a | 57 ± 4 a |
| 200 | 38 ± 2 b | 2.7 ± 0.3 a | 24 ± 3 c | 20 ± 3 b | 207 ± 6 b | 152 ± 5 a | 125 ± 4 a | 56 ± 3 a |
| 400 | 38 ± 3 b | 2.8 ± 0.3 a | 26 ± 3 b | 21 ± 2 b | 210 ± 8 a | 155 ± 4 a | 135 ± 6 a | 58 ± 2 a |
| CH-NPs (mg L−1) | Phenolic Compounds | Soluble Carbohydrates | Proline | Amino Acids |
|---|---|---|---|---|
| (mg g−1) | ||||
| C | 2.02 ± 0.8 d | 22.52 ± 1.23 c | 3.00 ± 0.07 e | 22 ± 2 e |
| 0 | 2.45 ± 0.05 c | 26.57 ± 1.53 b | 3.12 ± 0.09 d | 26 ± 2 d |
| 100 | 3.46 ± 0.06 b | 34.68 ± 2.93 a | 3.89 ± 0.13 c | 37 ± 3 c |
| 200 | 4.15 ± 0.09 a | 35.23 ± 2.48 a | 4.27 ± 0.10 b | 42 ± 3 b |
| 400 | 4.39 ± 0.04 a | 38.12 ± 3.59 a | 4.69 ± 0.12 a | 55 ± 4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, A.; Li, J.; AL-Huqail, A.A.; AL-Harbi, M.S.; Ali, E.F.; Wang, J.; Ding, Z.; Rekaby, S.A.; Ghoneim, A.M.; Eissa, M.A. Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants. Nanomaterials 2021, 11, 2670. https://doi.org/10.3390/nano11102670
Wang A, Li J, AL-Huqail AA, AL-Harbi MS, Ali EF, Wang J, Ding Z, Rekaby SA, Ghoneim AM, Eissa MA. Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants. Nanomaterials. 2021; 11(10):2670. https://doi.org/10.3390/nano11102670
Chicago/Turabian StyleWang, Anbang, Jingyang Li, Arwa Abdulkreem AL-Huqail, Mohammad S. AL-Harbi, Esmat F. Ali, Jiashui Wang, Zheli Ding, Saudi A. Rekaby, Adel M. Ghoneim, and Mamdouh A. Eissa. 2021. "Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants" Nanomaterials 11, no. 10: 2670. https://doi.org/10.3390/nano11102670