
Zebrafish Models for the Safety and Therapeutic Testing of Nanoparticles with a Focus on Macrophages

 , , , and
, , , and
Abstract
:
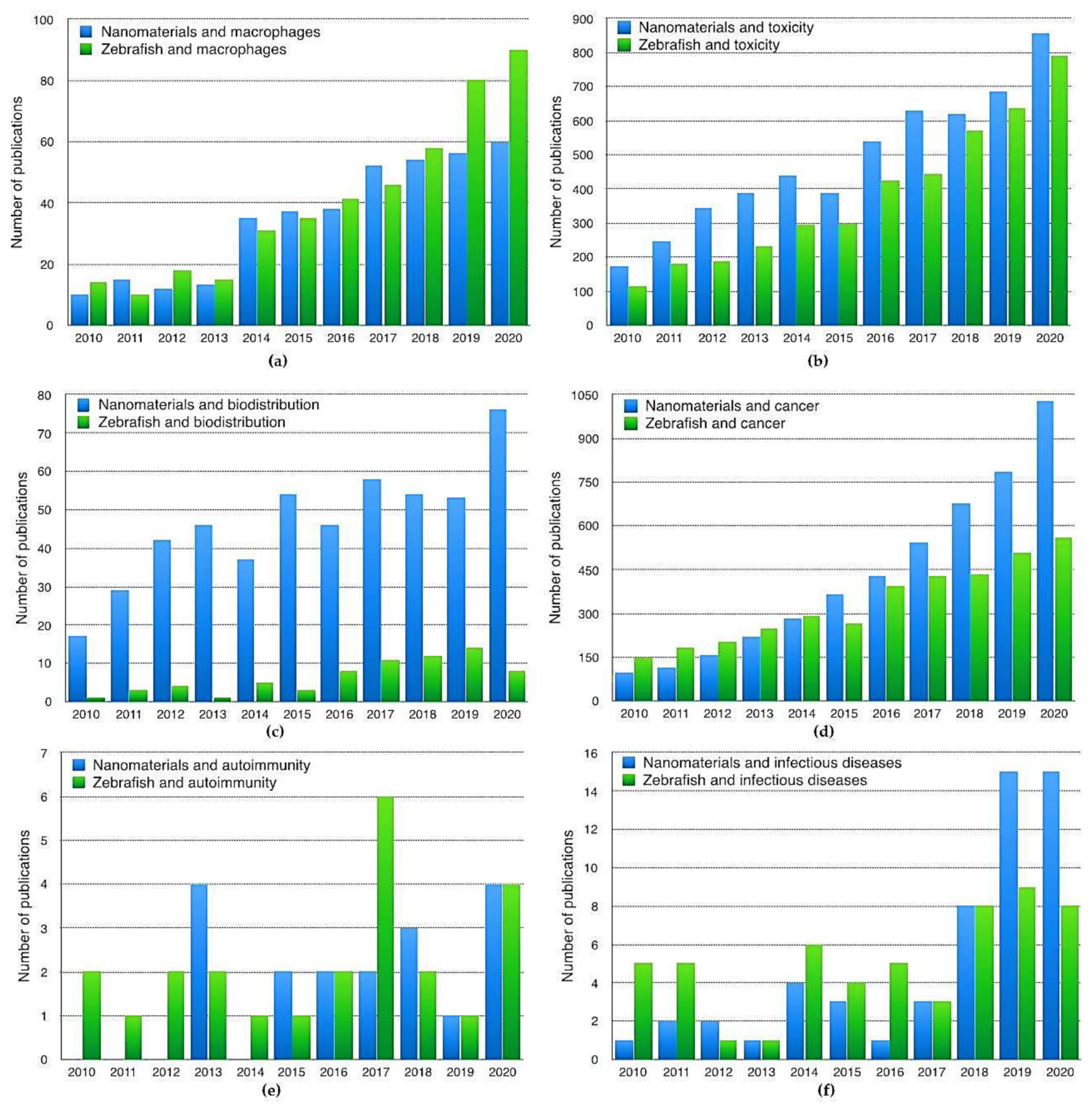
1. Introduction
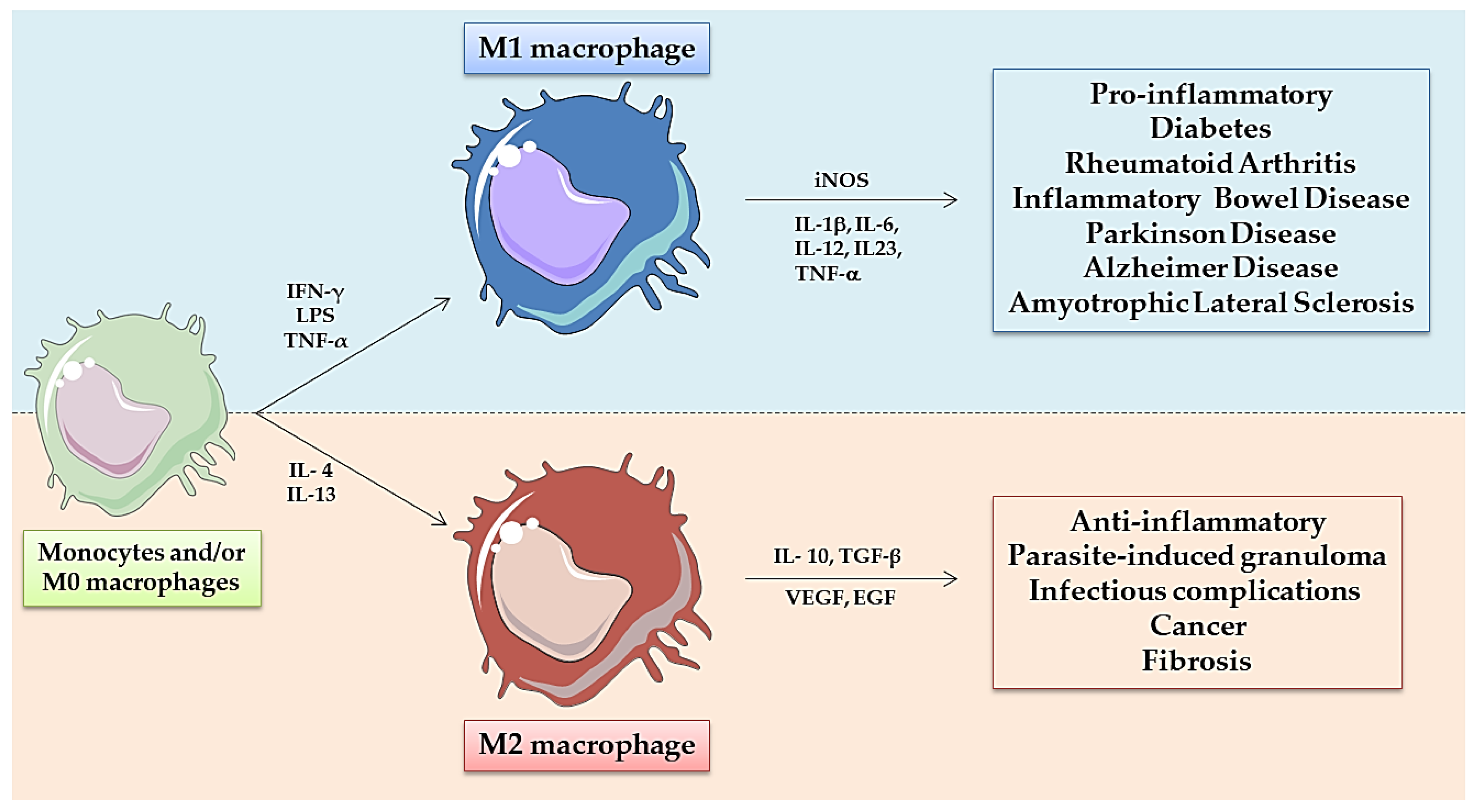
2. Zebrafish Innate Immune System: Role of Macrophages
3. Toxicological and Biodistribution Evaluation of Nanoparticles Using Zebrafish: Focus on Macrophages
3.1. Toxicological Studies
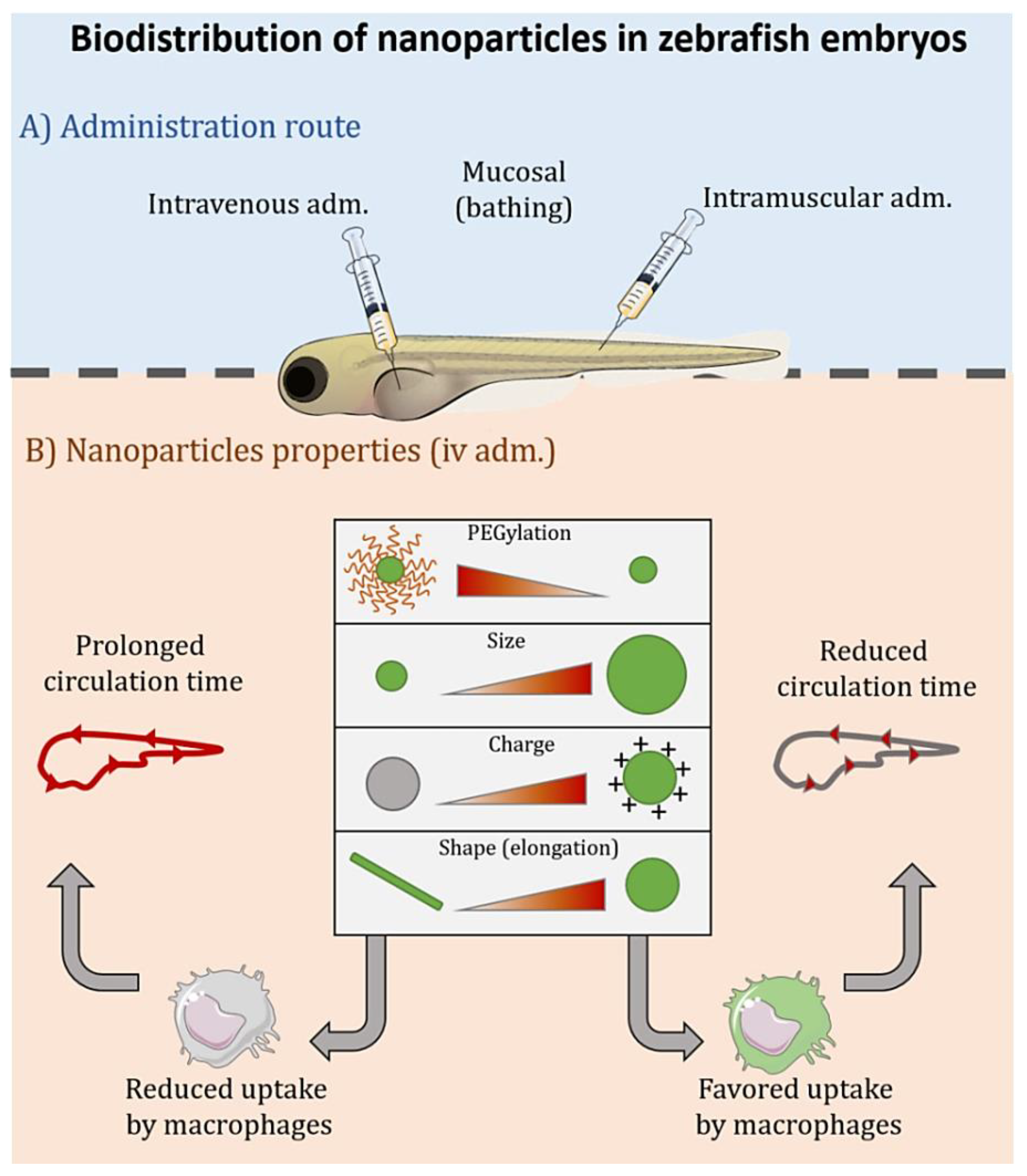
3.2. Biodistribution
4. Preclinical Testing of Nanoparticles Using Zebrafish Models of Disease: Relevance of Macrophages
4.1. Cancer
4.2. Autoimmune Diseases
4.2.1. Inflammatory Bowel Disease
4.2.2. Type I Diabetes Mellitus
4.2.3. Rheumatoid Arthritis
4.2.4. Neuroinflammatory and Neurodegenerative Diseases
4.3. Infectious Diseases
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AgNPs | Silver NPs |
| ALS | Amyotrophic Lateral Sclerosis |
| APCs | Antigen-presenting cells |
| APP | Human amyloid precursor protein |
| ASO-CaP-lipid NPs | SOD1 antisense oligonucleotide-calcium phosphate lipid coated NPs |
| AuNPs | Gold NPs |
| Aβ | β-amyloid |
| BBB | Blood-brain-barrier |
| BmE-PtNPs | Bacopa monnieri platinum NPs |
| CFSE | Carboxyfluorescein succinimidyl ester |
| CHT | Caudal hematopoietic tissue |
| cox-2 | Cyclooxygenase-2 |
| CuNPs | Copper nanoparticles |
| DOX | Doxorubicin |
| dpf | Days post-fertilization |
| dpi | days post-injection |
| EGF | Epithelial growth factor |
| EP/AgNPs | Eysenhardtia polystachya-loaded silver NPs |
| ER | Endoplasmic reticulum |
| FET | Fish Embryo Acute Toxicity |
| GFP | Green fluorescent protein |
| GSH | Glutathione |
| GSH-Px | Glutathione peroxidase |
| hpf | Hours post-fertilization |
| hpi | Hours post-injection |
| HSCs | Hematopoietic stem cells |
| i.m. | Intramuscular |
| i.v. | Intravenously |
| IBD | Inflammatory bowel disease |
| IBs | Inclusion bodies |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| IPNV | Pancreatic necrosis virus |
| LPS | Lipopolysaccharide |
| MDA | Malondialdehyde |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| mRNA | Messenger RNA |
| MVs | Membrane nanovesicles |
| NCs | Nanocapsules |
| NK | Natural killer |
| NPs | Nanoparticles |
| NQs | Quercetin NPs |
| NSAIDs | Nonsteroidal anti-inflammatory drugs |
| PAMAM | Polyamidoamine |
| PD | Parkinson’s disease |
| PDT | Photodynamic therapy |
| PDX | Patient-derived xenografts |
| PEG | Polyethylene glycol |
| PEG-PCL NPs | Poly(ethylene glycol)-co-poly(ε-caprolactone) NPs |
| PEG-PDPA NPs | Poly(ethylene glycol)-block-poly(2-(diisopropyl amino) ethyl methacrylate) NPs |
| PLA NPs | Poly(Lactic Acid) NPs |
| PLGA-NPs | Poly(lactic-co-glycolic acid) NPs |
| pMHC-NPs | Peptide-major histocompatibility complexes NPs |
| PMOsPOR-NH2 | Porous porphyrin-based organosilica NPs |
| PORBSNs | Non-porous porphyrin-based bridged silsesquioxane NPs |
| PS | Polystyrene |
| PtPP-HA | Kiteplatin-pyrophosphate-loaded hydroxyapatite NPs |
| PTZ | Pentylenetetrazole |
| PU-NCs | Puerarin nanocristals |
| RA | Rheumatoid arthritis |
| RFP | Red fluorescent protein |
| ROS | Reactive oxygen species |
| SA-NCs | Schisantherin nanocrystals |
| siHIF | Hypoxia-inducible factor 1a siRNA |
| SiNPs | Silica NPs |
| SLN-Q | Solid lipid NPs of quercetin |
| SOD | Superoxide dismutase |
| STZ | Streptozotocin |
| T1DM | Type 1 diabetes mellitus |
| TAMs | Tumor-associated macrophages |
| Tf-DOX-ReSi-Au | Redox-responsive silica-gold NPs functionalized with transferrin- DOX |
| TGF-β | Transforming growth factor-β |
| TiO2 NPs | Titanium dioxide NPs |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor-α |
| TPE-PDT | Two-photon-excited photodynamic therapy |
| VEGF | Vascular endothelial growth factor |
| VHSV | Viral hemorrhagic septicemia virus |
| VNNV | Viral nervous necrosis virus |
| wpf | Weeks post-fertilization |
| Y-CDs | Amphiphilic yellow-emissive carbon dots |
| ZnO-NPs | Zinc oxide NPs |
| βCas AuNPs | Casein coated-gold NPs |
References
- Pelaz, B.; Alexiou, C.; Alvarez-Puebla, R.A.; Alves, F.; Andrews, A.M.; Ashraf, S.; Balogh, L.P.; Ballerini, L.; Bestetti, A.; Brendel, C.; et al. Diverse Applications of Nanomedicine. ACS Nano 2017, 11, 2313–2381. [Google Scholar] [CrossRef] [Green Version]
- Ramsden, J. Nanotechnology: An Introduction; William Andrew: Norwich, NY, USA, 2016. [Google Scholar]
- Faria, M.; Björnmalm, M.; Thurecht, K.J.; Kent, S.J.; Parton, R.G.; Kavallaris, M.; Johnston, A.P.R.; Gooding, J.J.; Corrie, S.R.; Boyd, B.J.; et al. Minimum information reporting in bio–nano experimental literature. Nat. Nanotechnol. 2018, 13, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B.; Farcal, L.; Hardy, B.; Vázquez-Campos, S.; Hristozov, D.; Marcomini, A.; Lynch, I.; Valsami-Jones, E.; Alenius, H.; Savolainen, K. Advanced tools for the safety assessment of nanomaterials. Nat. Nanotechnol. 2018, 13, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Andón, F.T.; Fadeel, B. Nanotoxicology: Towards safety by design. In Nano-Oncologicals: New Targeting and Delivery Approaches; Alonso, M.J., Garcia-Fuentes, M., Eds.; Springer International Publishing: Cham, Germany, 2014; pp. 391–424. [Google Scholar] [CrossRef]
- Wang, H.; Brown, P.C.; Chow, E.C.Y.; Ewart, L.; Ferguson, S.S.; Fitzpatrick, S.; Freedman, B.S.; Guo, G.L.; Hedrich, W.; Heyward, S.; et al. 3D Cell Culture Models: Drug Pharmacokinetics, Safety Assessment, and Regulatory Consideration. Clin. Transl. Sci. 2021. [Google Scholar] [CrossRef]
- Fazio, M.; Ablain, J.; Chuan, Y.; Langenau, D.M.; Zon, L.I. Zebrafish patient avatars in cancer biology and precision cancer therapy. Nat. Rev. Cancer 2020, 20, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.R.; Zhu, Y.X.; Duan, Q.Y.; Chen, Z.; Wu, F.G. Nanomaterials meet zebrafish: Toxicity evaluation and drug delivery applications. J. Control. Release 2019, 311, 301–318. [Google Scholar] [CrossRef]
- Zon, L.I. Zebrafish: A new model for human disease. Genome Res. 1999, 9, 99–100. [Google Scholar]
- Xie, Y.; Meijer, A.H.; Schaaf, M.J.M. Modeling inflammation in zebrafish for the development of anti-inflammatory drugs. Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef]
- Astin, J.; Keerthisinghe, P.; Du, L.; Sanderson, L.; Crosier, K.; Crosier, P.; Hall, C. Innate immune cells and bacterial infection in zebrafish. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 138, pp. 31–60. [Google Scholar]
- Renshaw, S.A.; Trede, N.S. A model 450 million years in the making: Zebrafish and vertebrate immunity. Dis. Models Mech. 2012, 5, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, S.; Chua, H.; Gong, Z.; Lam, T.; Sin, Y. Development and maturation of the immune system in zebrafish, Danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- de Jong, J.L.; Zon, L.I. Use of the zebrafish system to study primitive and definitive hematopoiesis. Annu. Rev. Genet. 2005, 39, 481–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, E.J.; Zon, L.I. Hematopoietic development in the zebrafish. Int. J. Dev. Biol. 2010, 54, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Herbomel, P.; Thisse, B.; Thisse, C. Ontogeny and behaviour of early macrophages in the zebrafish embryo. Development 1999, 126, 3735–3745. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.V.; Pillay, L.M.; Venero Galanternik, M.; Weinstein, B.M. The zebrafish: A fintastic model for hematopoietic development and disease. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e312. [Google Scholar] [CrossRef] [PubMed]
- Murayama, E.; Kissa, K.; Zapata, A.; Mordelet, E.; Briolat, V.; Lin, H.F.; Handin, R.I.; Herbomel, P. Tracing hematopoietic precursor migration to successive hematopoietic organs during zebrafish development. Immunity 2006, 25, 963–975. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Jia, W.; Li, J.; Li, K.; Zhao, Q. Retinoic acid signaling plays a restrictive role in zebrafish primitive myelopoiesis. PLoS ONE 2012, 7, e30865. [Google Scholar] [CrossRef]
- Chen, A.T.; Zon, L.I. Zebrafish blood stem cells. J. Cell. Biochem. 2009, 108, 35–42. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [Green Version]
- Varol, C.; Mildner, A.; Jung, S. Macrophages: Development and Tissue Specialization. Annu. Rev. Immunol. 2015, 33, 643–675. [Google Scholar] [CrossRef]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of macrophage polarization in autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Bronte, V.; Brandau, S.; Chen, S.H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 2016, 7, 12150. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Erreni, M.; Allavena, P.; Porta, C. Macrophage polarization in pathology. Cell. Mol. Life Sci. CMLS 2015, 72, 4111–4126. [Google Scholar] [CrossRef]
- Zhang, X.; Mosser, D.M. Macrophage activation by endogenous danger signals. J. Pathol. 2008, 214, 161–178. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allavena, P.; Anfray, C.; Ummarino, A.; Andón, F.T. Therapeutic manipulation of tumor-associated macrophages: Facts and hopes from a clinical and translational perspective. Clin. Cancer Res. 2021, 27, 3291–3297. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Chi, M.; Laplace-Builhe, B.; Travnickova, J.; Luz-Crawford, P.; Tejedor, G.; Phan, Q.T.; Duroux-Richard, I.; Levraud, J.P.; Kissa, K.; Lutfalla, G.; et al. Identification of polarized macrophage subsets in zebrafish. eLife 2015, 4, e07288. [Google Scholar] [CrossRef] [PubMed]
- Ellett, F.; Pase, L.; Hayman, J.W.; Andrianopoulos, A.; Lieschke, G.J. mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood 2011, 117, e49–e56. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Chi, M.; Phan, Q.T.; Gonzalez, C.; Dubremetz, J.F.; Levraud, J.P.; Lutfalla, G. Transient infection of the zebrafish notochord with E. coli induces chronic inflammation. Dis. Models Mech. 2014, 7, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderson, L.E.; Chien, A.T.; Astin, J.W.; Crosier, K.E.; Crosier, P.S.; Hall, C.J. An inducible transgene reports activation of macrophages in live zebrafish larvae. Dev. Comp. Immunol. 2015, 53, 63–69. [Google Scholar] [CrossRef]
- Bernut, A.; Herrmann, J.L.; Kissa, K.; Dubremetz, J.F.; Gaillard, J.L.; Lutfalla, G.; Kremer, L. Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc. Natl. Acad. Sci. USA 2014, 111, E943–E952. [Google Scholar] [CrossRef] [Green Version]
- Johnston, H.J.; Verdon, R.; Gillies, S.; Brown, D.M.; Fernandes, T.F.; Henry, T.B.; Rossi, A.G.; Tran, L.; Tucker, C.; Tyler, C.R.; et al. Adoption of in vitro systems and zebrafish embryos as alternative models for reducing rodent use in assessments of immunological and oxidative stress responses to nanomaterials. Crit. Rev. Toxicol. 2018, 48, 252–271. [Google Scholar] [CrossRef]
- Brannen, K.C.; Chapin, R.E.; Jacobs, A.C.; Green, M.L. Alternative models of developmental and reproductive toxicity in pharmaceutical risk assessment and the 3Rs. ILAR J. 2016, 57, 144–156. [Google Scholar] [CrossRef] [Green Version]
- Sipes, N.S.; Padilla, S.; Knudsen, T.B. Zebrafish: As an integrative model for twenty-first century toxicity testing. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 256–267. [Google Scholar] [CrossRef]
- Cheng, H.; Yan, W.; Wu, Q.; Liu, C.; Gong, X.; Hung, T.C.; Li, G. Parental exposure to microcystin-LR induced thyroid endocrine disruption in zebrafish offspring, a transgenerational toxicity. Environ. Pollut. 2017, 230, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Li, Y.; Zhang, S.; Song, N.; Xu, H.; Dang, Y.; Liu, C.; Giesy, J.P.; Yu, H. Prenatal transfer of decabromodiphenyl ether (BDE-209) results in disruption of the thyroid system and developmental toxicity in zebrafish offspring. Aquat. Toxicol. (Amst. Neth.) 2017, 190, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.A.C.; Carleton, C.R.; Leeke, B.; Hampton, M.B.; Horsfield, J.A. Embryonic oxidative stress results in reproductive impairment for adult zebrafish. Redox Biol. 2015, 6, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, F.; Zhu, L.; Li, H.; Yu, S.; Wang, C.; Qiu, L. Reproductive toxicity of azoxystrobin to adult zebrafish (Danio rerio). Environ. Pollut. 2016, 219, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, S.J.; Bae, M.A.; Kim, J.R.; Cho, K.H. Cadmium exposure exacerbates severe hyperlipidemia and fatty liver changes in zebrafish via impairment of high-density lipoproteins functionality. Toxicol. Vitr. 2018, 47, 249–258. [Google Scholar] [CrossRef]
- Qin, X.; Laroche, F.F.J.; Peerzade, S.; Lam, A.; Sokolov, I.; Feng, H. In Vivo Targeting of Xenografted Human Cancer Cells with Functionalized Fluorescent Silica Nanoparticles in Zebrafish. J. Vis. Exp. 2020, 159, e61187. [Google Scholar] [CrossRef]
- Rothenbücher, T.S.P.; Ledin, J.; Gibbs, D.; Engqvist, H.; Persson, C.; Hulsart-Billström, G. Zebrafish embryo as a replacement model for initial biocompatibility studies of biomaterials and drug delivery systems. Acta Biomater. 2019, 100, 235–243. [Google Scholar] [CrossRef]
- No, O.T. 236: Fish embryo acute toxicity (FET) test. OECD Guidel. Test. Chem. Sect. 2013, 2, 1–22. [Google Scholar]
- OJEU. Directive 2010/63/EU of the European Parliament and of the Council on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Nishimura, Y.; Inoue, A.; Sasagawa, S.; Koiwa, J.; Kawaguchi, K.; Kawase, R.; Maruyama, T.; Kim, S.; Tanaka, T. Using zebrafish in systems toxicology for developmental toxicity testing. Congenit. Anom. 2016, 56, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Cornet, C.; Calzolari, S.; Miñana-Prieto, R.; Dyballa, S.; van Doornmalen, E.; Rutjes, H.; Savy, T.; D’Amico, D.; Terriente, J. ZeGlobalTox: An Innovative Approach to Address Organ Drug Toxicity Using Zebrafish. Int. J. Mol. Sci. 2017, 18, 864. [Google Scholar] [CrossRef] [Green Version]
- Dach, K.; Yaghoobi, B.; Schmuck, M.R.; Carty, D.R.; Morales, K.M.; Lein, P.J. Teratological and Behavioral Screening of the National Toxicology Program 91-Compound Library in Zebrafish (Danio rerio). Toxicol. Sci. 2019, 167, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Marrero-Ponce, Y.; Siverio-Mota, D.; Gálvez-Llompart, M.; Recio, M.C.; Giner, R.M.; García-Domènech, R.; Torrens, F.; Arán, V.J.; Cordero-Maldonado, M.L.; Esguera, C.V.; et al. Discovery of novel anti-inflammatory drug-like compounds by aligning in silico and in vivo screening: The nitroindazolinone chemotype. Eur. J. Med. Chem. 2011, 46, 5736–5753. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Zebrafish: A complete animal model to enumerate the nanoparticle toxicity. J. Nanobiotechnol. 2016, 14, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, C.; Tang, M. Toxicological study of metal and metal oxide nanoparticles in zebrafish. J. Appl. Toxicol. JAT 2020, 40, 37–63. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Xu, P.; Chen, G.; Zeng, G.; Chen, A.; Hu, L.; Huang, Z.; He, K.; Guo, Z.; Liu, W.; et al. Effects of silver nanoparticles with different dosing regimens and exposure media on artificial ecosystem. J. Environ. Sci. (China) 2019, 75, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, X.; Wu, Y.; Hou, J.; Zhang, S.; Zhou, N.; Wang, X. Toxicity responses of different organs of zebrafish (Danio rerio) to silver nanoparticles with different particle sizes and surface coatings. Environ. Pollut. 2019, 246, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Abramenko, N.B.; Demidova, T.B.; Abkhalimov, E.V.; Ershov, B.G.; Krysanov, E.Y.; Kustov, L.M. Ecotoxicity of different-shaped silver nanoparticles: Case of zebrafish embryos. J. Hazard. Mater. 2018, 347, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Rotchell, J.M.; Cheng, J.; Yi, J.; Zhang, Q. Silver nanoparticles affect the neural development of zebrafish embryos. J. Appl. Toxicol. JAT 2015, 35, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Brundo, M.V.; Pecoraro, R.; Marino, F.; Salvaggio, A.; Tibullo, D.; Saccone, S.; Bramanti, V.; Buccheri, M.A.; Impellizzeri, G.; Scuderi, V.; et al. Toxicity Evaluation of New Engineered Nanomaterials in Zebrafish. Front. Physiol. 2016, 7, 130. [Google Scholar] [CrossRef]
- Ghobadian, M.; Nabiuni, M.; Parivar, K.; Fathi, M.; Pazooki, J. Toxic effects of magnesium oxide nanoparticles on early developmental and larval stages of zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2015, 122, 260–267. [Google Scholar] [CrossRef]
- Girigoswami, K.; Viswanathan, M.; Murugesan, R.; Girigoswami, A. Studies on polymer-coated zinc oxide nanoparticles: UV-blocking efficacy and in vivo toxicity. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 56, 501–510. [Google Scholar] [CrossRef]
- Li, X.; Ji, X.; Wang, R.; Zhao, J.; Dang, J.; Gao, Y.; Jin, M. Zebrafish behavioral phenomics employed for characterizing behavioral neurotoxicity caused by silica nanoparticles. Chemosphere 2020, 240, 124937. [Google Scholar] [CrossRef]
- Vranic, S.; Shimada, Y.; Ichihara, S.; Kimata, M.; Wu, W.; Tanaka, T.; Boland, S.; Tran, L.; Ichihara, G. Toxicological Evaluation of SiO2 Nanoparticles by Zebrafish Embryo Toxicity Test. Int. J. Mol. Sci. 2019, 20, 882. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Hu, H.; Li, Q.; Jiang, L.; Zou, Y.; Wang, Y.; Sun, Z. Combined toxicity of silica nanoparticles and methylmercury on cardiovascular system in zebrafish (Danio rerio) embryos. Environ. Toxicol. Pharmacol. 2016, 44, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Ginzburg, A.L.; Truong, L.; Tanguay, R.L.; Hutchison, J.E. Synergistic Toxicity Produced by Mixtures of Biocompatible Gold Nanoparticles and Widely Used Surfactants. ACS Nano 2018, 12, 5312–5322. [Google Scholar] [CrossRef] [PubMed]
- Harper, B.; Thomas, D.; Chikkagoudar, S.; Baker, N.; Tang, K.; Heredia-Langner, A.; Lins, R.; Harper, S. Comparative hazard analysis and toxicological modeling of diverse nanomaterials using the embryonic zebrafish (EZ) metric of toxicity. J. Nanoparticle Res. 2015, 17, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodewein, L.; Schmelter, F.; Di Fiore, S.; Hollert, H.; Fischer, R.; Fenske, M. Differences in toxicity of anionic and cationic PAMAM and PPI dendrimers in zebrafish embryos and cancer cell lines. Toxicol. Appl. Pharmacol. 2016, 305, 83–92. [Google Scholar] [CrossRef]
- Jeong, J.; Cho, H.J.; Choi, M.; Lee, W.S.; Chung, B.H.; Lee, J.S. In vivo toxicity assessment of angiogenesis and the live distribution of nano-graphene oxide and its PEGylated derivatives using the developing zebrafish embryo. Carbon 2015, 93, 431–440. [Google Scholar] [CrossRef]
- Teijeiro-Valiño, C.; Yebra-Pimentel, E.; Guerra-Varela, J.; Csaba, N.; Alonso, M.J.; Sánchez, L. Assessment of the permeability and toxicity of polymeric nanocapsules using the zebrafish model. Nanomed. (Lond. Engl.) 2017, 12, 2069–2082. [Google Scholar] [CrossRef]
- Crecente-Campo, J.; Guerra-Varela, J.; Peleteiro, M.; Gutiérrez-Lovera, C.; Fernández-Mariño, I.; Diéguez-Docampo, A.; González-Fernández, Á.; Sánchez, L.; Alonso, M.J. The size and composition of polymeric nanocapsules dictate their interaction with macrophages and biodistribution in zebrafish. J. Control. Release 2019, 308, 98–108. [Google Scholar] [CrossRef]
- Chang, H.; Yhee, J.Y.; Jang, G.H.; You, D.G.; Ryu, J.H.; Choi, Y.; Na, J.H.; Park, J.H.; Lee, K.H.; Choi, K.; et al. Predicting the in vivo accumulation of nanoparticles in tumor based on in vitro macrophage uptake and circulation in zebrafish. J. Control. Release 2016, 244, 205–213. [Google Scholar] [CrossRef]
- Evensen, L.; Johansen, P.L.; Koster, G.; Zhu, K.; Herfindal, L.; Speth, M.; Fenaroli, F.; Hildahl, J.; Bagherifam, S.; Tulotta, C.; et al. Zebrafish as a model system for characterization of nanoparticles against cancer. Nanoscale 2016, 8, 862–877. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cao, F.; Zhao, F.; Yang, Y.; Teng, M.; Wang, C.; Qiu, L. Developmental toxicity, oxidative stress and immunotoxicity induced by three strobilurins (pyraclostrobin, trifloxystrobin and picoxystrobin) in zebrafish embryos. Chemosphere 2018, 207, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yin, J.; Liang, Y.; Yuan, S.; Wang, F.; Song, M.; Wang, H. Oxidative stress and immunotoxicity induced by graphene oxide in zebrafish. Aquat. Toxicol. (Amst. Neth.) 2016, 174, 54–60. [Google Scholar] [CrossRef]
- Andón, F.T.; Fadeel, B. Programmed Cell Death: Molecular Mechanisms and Implications for Safety Assessment of Nanomaterials. Acc. Chem. Res. 2013, 46, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, K.; Andón, F.T.; El-Sayed, R.; Fadeel, B. Mechanisms of carbon nanotube-induced toxicity: Focus on pulmonary inflammation. Adv. Drug Deliv. Rev. 2013, 65, 2087–2097. [Google Scholar] [CrossRef] [PubMed]
- Andón, F.T.; Kapralov, A.A.; Yanamala, N.; Feng, W.; Baygan, A.; Chambers, B.J.; Hultenby, K.; Ye, F.; Toprak, M.S.; Brandner, B.D.; et al. Biodegradation of Single-Walled Carbon Nanotubes by Eosinophil Peroxidase. Small 2013, 9, 2721–2729. [Google Scholar] [CrossRef]
- Truong, L.; Tilton, S.C.; Zaikova, T.; Richman, E.; Waters, K.M.; Hutchison, J.E.; Tanguay, R.L. Surface functionalities of gold nanoparticles impact embryonic gene expression responses. Nanotoxicology 2013, 7, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnaraj, C.; Harper, S.L.; Yun, S.I. In Vivo toxicological assessment of biologically synthesized silver nanoparticles in adult Zebrafish (Danio rerio). J. Hazard. Mater. 2016, 301, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Brun, N.R.; Lenz, M.; Wehrli, B.; Fent, K. Comparative effects of zinc oxide nanoparticles and dissolved zinc on zebrafish embryos and eleuthero-embryos: Importance of zinc ions. Sci. Total Environ. 2014, 476-477, 657–666. [Google Scholar] [CrossRef]
- Vibe, C.B.; Fenaroli, F.; Pires, D.; Wilson, S.R.; Bogoeva, V.; Kalluru, R.; Speth, M.; Anes, E.; Griffiths, G.; Hildahl, J. Thioridazine in PLGA nanoparticles reduces toxicity and improves rifampicin therapy against mycobacterial infection in zebrafish. Nanotoxicology 2016, 10, 680–688. [Google Scholar] [CrossRef]
- Velikova, N.; Mas, N.; Miguel-Romero, L.; Polo, L.; Stolte, E.; Zaccaria, E.; Cao, R.; Taverne, N.; Murguía, J.R.; Martinez-Manez, R.; et al. Broadening the antibacterial spectrum of histidine kinase autophosphorylation inhibitors via the use of ε-poly-L-lysine capped mesoporous silica-based nanoparticles. Nanomedicine 2017, 13, 569–581. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jang, G.H.; Byun, C.H.; Jeun, M.; Searson, P.C.; Lee, K.H. Zebrafish models for functional and toxicological screening of nanoscale drug delivery systems: Promoting preclinical applications. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Shwartz, A.; Goessling, W.; Yin, C. Macrophages in Zebrafish Models of Liver Diseases. Front. Immunol. 2019, 10, 2840. [Google Scholar] [CrossRef]
- Zhang, X.; Song, J.; Klymov, A.; Zhang, Y.; de Boer, L.; Jansen, J.A.; van den Beucken, J.J.; Yang, F.; Zaat, S.A.; Leeuwenburgh, S.C. Monitoring local delivery of vancomycin from gelatin nanospheres in zebrafish larvae. Int. J. Nanomed. 2018, 13, 5377–5394. [Google Scholar] [CrossRef] [Green Version]
- Andón, F.T.; Digifico, E.; Maeda, A.; Erreni, M.; Mantovani, A.; Alonso, M.J.; Allavena, P. Targeting tumor associated macrophages: The new challenge for nanomedicine. Semin. Immunol. 2017, 34, 103–113. [Google Scholar] [CrossRef]
- Duan, X.; Li, Y. Physicochemical characteristics of nanoparticles affect circulation, biodistribution, cellular internalization, and trafficking. Small 2013, 9, 1521–1532. [Google Scholar] [CrossRef]
- Dal, N.K.; Kocere, A.; Wohlmann, J.; Van Herck, S.; Bauer, T.A.; Resseguier, J.; Bagherifam, S.; Hyldmo, H.; Barz, M.; De Geest, B.G.; et al. Zebrafish Embryos Allow Prediction of Nanoparticle Circulation Times in Mice and Facilitate Quantification of Nanoparticle-Cell Interactions. Small 2020, 16, e1906719. [Google Scholar] [CrossRef] [PubMed]
- Campbell, F.; Bos, F.L.; Sieber, S.; Arias-Alpizar, G.; Koch, B.E.; Huwyler, J.; Kros, A.; Bussmann, J. Directing Nanoparticle Biodistribution through Evasion and Exploitation of Stab2-Dependent Nanoparticle Uptake. ACS Nano 2018, 12, 2138–2150. [Google Scholar] [CrossRef] [PubMed]
- Sieber, S.; Grossen, P.; Uhl, P.; Detampel, P.; Mier, W.; Witzigmann, D.; Huwyler, J. Zebrafish as a predictive screening model to assess macrophage clearance of liposomes in vivo. Nanomedicine 2019, 17, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Teijeiro-Valiño, C.; Novoa-Carballal, R.; Borrajo, E.; Vidal, A.; Alonso-Nocelo, M.; de la Fuente Freire, M.; Lopez-Casas, P.P.; Hidalgo, M.; Csaba, N.; Alonso, M.J. A multifunctional drug nanocarrier for efficient anticancer therapy. J. Control. Release 2019, 294, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Crecente-Campo, J.; Lorenzo-Abalde, S.; Mora, A.; Marzoa, J.; Csaba, N.; Blanco, J.; González-Fernández, Á.; Alonso, M.J. Bilayer polymeric nanocapsules: A formulation approach for a thermostable and adjuvanted E. coli antigen vaccine. J. Control. Release 2018, 286, 20–32. [Google Scholar] [CrossRef]
- Vicente, S.; Peleteiro, M.; Gonzalez-Aramundiz, J.V.; Díaz-Freitas, B.; Martínez-Pulgarín, S.; Neissa, J.I.; Escribano, J.M.; Sanchez, A.; González-Fernández, A.; Alonso, M.J. Highly versatile immunostimulating nanocapsules for specific immune potentiation. Nanomed. (Lond. Engl.) 2014, 9, 2273–2289. [Google Scholar] [CrossRef]
- de la Fuente, M.; Raviña, M.; Paolicelli, P.; Sanchez, A.; Seijo, B.; Alonso, M.J. Chitosan-based nanostructures: A delivery platform for ocular therapeutics. Adv. Drug Deliv. Rev. 2010, 62, 100–117. [Google Scholar] [CrossRef]
- Truong, N.P.; Whittaker, M.R.; Mak, C.W.; Davis, T.P. The importance of nanoparticle shape in cancer drug delivery. Expert Opin. Drug Deliv. 2015, 12, 129–142. [Google Scholar] [CrossRef]
- Wibroe, P.P.; Anselmo, A.C.; Nilsson, P.H.; Sarode, A.; Gupta, V.; Urbanics, R.; Szebeni, J.; Hunter, A.C.; Mitragotri, S.; Mollnes, T.E.; et al. Bypassing adverse injection reactions to nanoparticles through shape modification and attachment to erythrocytes. Nat. Nanotechnol. 2017, 12, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Sangabathuni, S.; Murthy, R.V.; Chaudhary, P.M.; Subramani, B.; Toraskar, S.; Kikkeri, R. Mapping the Glyco-Gold Nanoparticles of Different Shapes Toxicity, Biodistribution and Sequestration in Adult Zebrafish. Sci. Rep. 2017, 7, 4239. [Google Scholar] [CrossRef] [Green Version]
- Saez Talens, V.; Arias-Alpizar, G.; Makurat, D.M.M.; Davis, J.; Bussmann, J.; Kros, A.; Kieltyka, R.E. Stab2-Mediated Clearance of Supramolecular Polymer Nanoparticles in Zebrafish Embryos. Biomacromolecules 2020, 21, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Rességuier, J.; Delaune, E.; Coolen, A.L.; Levraud, J.P.; Boudinot, P.; Le Guellec, D.; Verrier, B. Specific and Efficient Uptake of Surfactant-Free Poly(Lactic Acid) Nanovaccine Vehicles by Mucosal Dendritic Cells in Adult Zebrafish after Bath Immersion. Front. Immunol. 2017, 8, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Miao, X.; Chen, T.; Yi, X.; Wang, R.; Zhao, H.; Lee, S.M.; Wang, X.; Zheng, Y. Zebrafish as a visual and dynamic model to study the transport of nanosized drug delivery systems across the biological barriers. Colloids Surfaces B Biointerfaces 2017, 156, 227–235. [Google Scholar] [CrossRef] [PubMed]
- van Pomeren, M.; Peijnenburg, W.; Vlieg, R.C.; van Noort, S.J.T.; Vijver, M.G. The biodistribution and immuno-responses of differently shaped non-modified gold particles in zebrafish embryos. Nanotoxicology 2019, 13, 558–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.M.; Recht, L.; Strober, S. The promise of targeting macrophages in cancer therapy. Clin. Cancer Res. 2017, 23, 3241–3250. [Google Scholar] [CrossRef] [Green Version]
- Nicoli, S.; Ribatti, D.; Cotelli, F.; Presta, M. Mammalian tumor xenografts induce neovascularization in zebrafish embryos. Cancer Res. 2007, 67, 2927–2931. [Google Scholar] [CrossRef] [Green Version]
- Nicoli, S.; Presta, M. The zebrafish/tumor xenograft angiogenesis assay. Nat. Protoc. 2007, 2, 2918. [Google Scholar] [CrossRef] [PubMed]
- Patton, E.E.; Widlund, H.R.; Kutok, J.L.; Kopani, K.R.; Amatruda, J.F.; Murphey, R.D.; Berghmans, S.; Mayhall, E.A.; Traver, D.; Fletcher, C.D.; et al. BRAF mutations are sufficient to promote nevi formation and cooperate with p53 in the genesis of melanoma. Curr. Biol. 2005, 15, 249–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michailidou, C.; Jones, M.; Walker, P.; Kamarashev, J.; Kelly, A.; Hurlstone, A.F. Dissecting the roles of Raf- and PI3K-signalling pathways in melanoma formation and progression in a zebrafish model. Dis Model. Mech. 2009, 2, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Davison, J.M.; Rhee, J.; Hruban, R.H.; Maitra, A.; Leach, S.D. Oncogenic KRAS induces progenitor cell expansion and malignant transformation in zebrafish exocrine pancreas. Gastroenterology 2008, 134, 2080–2090. [Google Scholar] [CrossRef] [Green Version]
- Sieber, S.; Grossen, P.; Bussmann, J.; Campbell, F.; Kros, A.; Witzigmann, D.; Huwyler, J. Zebrafish as a preclinical in vivo screening model for nanomedicines. Adv. Drug Deliv. Rev. 2019, 151, 152–168. [Google Scholar] [CrossRef] [PubMed]
- Valle, S.; Alcalá, S.; Martin-Hijano, L.; Cabezas-Sáinz, P.; Navarro, D.; Muñoz, E.R.; Yuste, L.; Tiwary, K.; Walter, K.; Ruiz-Cañas, L.; et al. Exploiting oxidative phosphorylation to promote the stem and immunoevasive properties of pancreatic cancer stem cells. Nat. Commun. 2020, 11, 5265. [Google Scholar] [CrossRef] [PubMed]
- Pudelko, L.; Edwards, S.; Balan, M.; Nyqvist, D.; Al-Saadi, J.; Dittmer, J.; Almlöf, I.; Helleday, T.; Bräutigam, L. An orthotopic glioblastoma animal model suitable for high-throughput screenings. Neuro-oncology 2018, 20, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Sounni, N.E.; Noel, A. Targeting the tumor microenvironment for cancer therapy. Clin. Chem. 2013, 59, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Movahedi, K.; Van Ginderachter, J.A. The Ontogeny and Microenvironmental Regulation of Tumor-Associated Macrophages. Antioxid. Redox Signal. 2016, 25, 775–791. [Google Scholar] [CrossRef] [PubMed]
- Anfray, C.; Ummarino, A.; Andón, F.T.; Allavena, P. Current Strategies to Target Tumor-Associated-Macrophages to Improve Anti-Tumor Immune Responses. Cells 2019, 9, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, N.D.; Weinstein, B.M. In Vivo Imaging of Embryonic Vascular Development Using Transgenic Zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renshaw, S.; Loynes, C.; Trushell, D.; Elworthy, S.; Ingham, P.; Whyte, M. A transgenic zebrafish model of neutrophilic inflammation. Blood 2007, 108, 3976–3978. [Google Scholar] [CrossRef]
- Cabezas-Sáinz, P.; Pensado-López, A.; Sáinz, B., Jr.; Sánchez, L. Modeling Cancer Using Zebrafish Xenografts: Drawbacks for Mimicking the Human Microenvironment. Cells 2020, 9, 1978. [Google Scholar] [CrossRef]
- Baxendale, S.; van Eeden, F.; Wilkinson, R. The power of zebrafish in personalised medicine. In Personalised Medicine; Springer: Berlin/Heidelberg, Germany, 2017; pp. 179–197. [Google Scholar]
- Germain, M.; Caputo, F.; Metcalfe, S.; Tosi, G.; Spring, K.; Åslund, A.K.O.; Pottier, A.; Schiffelers, R.; Ceccaldi, A.; Schmid, R. Delivering the power of nanomedicine to patients today. J. Control. Release 2020, 326, 164–171. [Google Scholar] [CrossRef]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer nanomedicine: Progress, challenges and opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef]
- Alonso, M.; Garcia-Fuentes, M. Nano-Oncologicals: New Targeting and Delivery Approaches; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Goldberg, M.S. Improving cancer immunotherapy through nanotechnology. Nat. Rev. Cancer 2019, 19, 587–602. [Google Scholar] [CrossRef]
- Lächelt, U.; Wagner, E. Nucleic acid therapeutics using polyplexes: A journey of 50 years (and beyond). Chem. Rev. 2015, 115, 11043–11078. [Google Scholar] [CrossRef]
- Hu, G.; Guo, M.; Xu, J.; Wu, F.; Fan, J.; Huang, Q.; Yang, G.; Lv, Z.; Wang, X.; Jin, Y. Nanoparticles Targeting Macrophages as Potential Clinical Therapeutic Agents Against Cancer and Inflammation. Front. Immunol. 2019, 10, 1998. [Google Scholar] [CrossRef]
- Nadar, R.A.; Asokan, N.; Degli Esposti, L.; Curci, A.; Barbanente, A.; Schlatt, L.; Karst, U.; Iafisco, M.; Margiotta, N.; Brand, M.; et al. Preclinical evaluation of platinum-loaded hydroxyapatite nanoparticles in an embryonic zebrafish xenograft model. Nanoscale 2020, 12, 13582–13594. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Lee, C.H.; Lin, M.S.; Chi, C.W.; Chen, Y.J.; Wang, G.S.; Liao, K.W.; Chiu, L.P.; Wu, S.H.; Huang, D.M.; et al. ZnO Nanoparticles Induced Caspase-Dependent Apoptosis in Gingival Squamous Cell Carcinoma through Mitochondrial Dysfunction and p70S6K Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1612. [Google Scholar] [CrossRef] [Green Version]
- Mauriello Jimenez, C.; Aggad, D.; Croissant, J.G.; Tresfield, K.; Laurencin, D.; Berthomieu, D.; Cubedo, N.; Rossel, M.; Alsaiari, S.; Anjum, D.H. Porous Porphyrin-Based Organosilica Nanoparticles for NIR Two-Photon Photodynamic Therapy and Gene Delivery in Zebrafish. Adv. Funct. Mater. 2018, 28, 1800235. [Google Scholar] [CrossRef]
- Dib, S.; Aggad, D.; Mauriello Jimenez, C.; Lakrafi, A.; Hery, G.; Nguyen, C.; Durand, D.; Morère, A.; El Cheikh, K.; Sol, V.; et al. Porphyrin-based bridged silsesquioxane nanoparticles for targeted two-photon photodynamic therapy of zebrafish xenografted with human tumor. Cancer Rep. 2019, 2, e1186. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Chou, H.Y.; Yuh, C.H.; Mekuria, S.L.; Kao, Y.C.; Tsai, H.C. Radiation-Sensitive Dendrimer-Based Drug Delivery System. Adv. Sci. 2018, 5, 1700339. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Guo, W.; Guo, N.; Huangfu, M.; Liu, H.; Lin, M.; Xu, W.; Chen, J.; Wang, T.; Wei, Q.; et al. Targeting tumor hypoxia with stimulus-responsive nanocarriers in overcoming drug resistance and monitoring anticancer efficacy. Acta Biomater. 2018, 71, 351–362. [Google Scholar] [CrossRef]
- Van Driessche, A.; Kocere, A.; Everaert, H.; Nuhn, L.; Van Herck, S.; Griffiths, G.; Fenaroli, F.; De Geest, B.G. pH-Sensitive Hydrazone-Linked Doxorubicin Nanogels via Polymeric-Activated Ester Scaffolds: Synthesis, Assembly, and In Vitro and In Vivo Evaluation in Tumor-Bearing Zebrafish. Chem. Mater. 2018, 30, 8587–8596. [Google Scholar] [CrossRef] [Green Version]
- Tu, W.M.; Huang, X.C.; Chen, Y.L.; Luo, Y.L.; Liau, I.; Hsu, H.Y. Longitudinal and quantitative assessment platform for concurrent analysis of anti-tumor efficacy and cardiotoxicity of nano-formulated medication in vivo. Anal. Chim. Acta 2020, 1095, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Kocere, A.; Resseguier, J.; Wohlmann, J.; Skjeldal, F.M.; Khan, S.; Speth, M.; Dal, N.J.K.; Ng, M.Y.W.; Alonso-Rodriguez, N.; Scarpa, E.; et al. Real-time imaging of polymersome nanoparticles in zebrafish embryos engrafted with melanoma cancer cells: Localization, toxicity and treatment analysis. EBioMedicine 2020, 58, 102902. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.I.; Chen, C.Y.; Liu, W.; Chang, P.C.; Huang, C.W.; Han, K.F.; Lin, I.P.; Lin, M.Y.; Lee, C.H. Sandensolide Induces Oxidative Stress-Mediated Apoptosis in Oral Cancer Cells and in Zebrafish Xenograft Model. Mar. Drugs 2018, 16, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmody, R.J.; Cotter, T.G. Signalling apoptosis: A radical approach. Redox Rep. 2001, 6, 77–90. [Google Scholar] [CrossRef]
- Peng, B.; Almeqdadi, M.; Laroche, F.; Palantavida, S.; Dokukin, M.; Roper, J.; Yilmaz, O.H.; Feng, H.; Sokolov, I. Ultrabright fluorescent cellulose acetate nanoparticles for imaging tumors through systemic and topical applications. Mater. Today 2019, 23, 16–25. [Google Scholar] [CrossRef]
- Costa, B.; Ferreira, S.; Póvoa, V.; Cardoso, M.J.; Vieira, S.; Stroom, J.; Fidalgo, P.; Rio-Tinto, R.; Figueiredo, N.; Parés, O.; et al. Developments in zebrafish avatars as radiotherapy sensitivity reporters-towards personalized medicine. EBioMedicine 2020, 51, 102578. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Emelyanov, A.; Koh, C.H.V.; Spitsbergen, J.M.; Parinov, S.; Gong, Z. An inducible kras(V12) transgenic zebrafish model for liver tumorigenesis and chemical drug screening. Dis. Models Mech. 2012, 5, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Z.; Chen, C.; Zeng, Z.; Yang, H.; Oh, J.; Chen, L.; Lu, S.C. Mechanism and significance of increased glutathione level in human hepatocellular carcinoma and liver regeneration. FASEB J. 2001, 15, 19–21. [Google Scholar] [CrossRef]
- Volk, T.; Jähde, E.; Fortmeyer, H.P.; Glüsenkamp, K.H.; Rajewsky, M.F. pH in human tumour xenografts: Effect of intravenous administration of glucose. Br. J. Cancer 1993, 68, 492–500. [Google Scholar] [CrossRef]
- Ang, C.Y.; Tan, S.Y.; Teh, C.; Lee, J.M.; Wong, M.F.E.; Qu, Q.; Poh, L.Q.; Li, M.; Zhang, Y.; Korzh, V.; et al. Redox and pH Dual Responsive Polymer Based Nanoparticles for In Vivo Drug Delivery. Small 2017, 13, 1602379. [Google Scholar] [CrossRef]
- Palantavida, S.; Guz, N.V.; Woodworth, C.D.; Sokolov, I. Ultrabright fluorescent mesoporous silica nanoparticles for prescreening of cervical cancer. Nanomedicine 2013, 9, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Peerzade, S.; Qin, X.; Laroche, F.J.F.; Palantavida, S.; Dokukin, M.; Peng, B.; Feng, H.; Sokolov, I. Ultrabright fluorescent silica nanoparticles for in vivo targeting of xenografted human tumors and cancer cells in zebrafish. Nanoscale 2019, 11, 22316–22327. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Lamers, G.E.; Beenakker, J.W.M.; Cui, C.; Ghotra, V.P.; Danen, E.H.; Meijer, A.H.; Spaink, H.P.; Snaar-Jagalska, B.E. Neutrophil-mediated experimental metastasis is enhanced by VEGFR inhibition in a zebrafish xenograft model. J. Pathol. 2012, 227, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Roh-Johnson, M.; Shah, A.N.; Stonick, J.A.; Poudel, K.R.; Kargl, J.; Yang, G.H.; di Martino, J.; Hernandez, R.E.; Gast, C.E.; Zarour, L.R.; et al. Macrophage-Dependent Cytoplasmic Transfer during Melanoma Invasion In Vivo. Dev. Cell 2017, 43, 549–562.e6. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.D.; Wyroba, B.; Chen, W.; Lockwood, R.A.; Tran, K.B.; Shepherd, P.R.; Hall, C.J.; Crosier, K.E.; Crosier, P.S.; Astin, J.W. Macrophages enhance Vegfa-driven angiogenesis in an embryonic zebrafish tumour xenograft model. Dis. Models Mech. 2018, 11, dmm035998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seoane, S.; Martinez-Ordoñez, A.; Eiro, N.; Cabezas-Sainz, P.; Garcia-Caballero, L.; Gonzalez, L.O.; Macia, M.; Sanchez, L.; Vizoso, F.; Perez-Fernandez, R. POU1F1 transcription factor promotes breast cancer metastasis via recruitment and polarization of macrophages. J. Pathol. 2019, 249, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Póvoa, V.; Rebelo de Almeida, C.; Maia-Gil, M.; Sobral, D.; Domingues, M.; Martinez-Lopez, M.; de Almeida Fuzeta, M.; Silva, C.; Grosso, A.R.; Fior, R. Innate immune evasion revealed in a colorectal zebrafish xenograft model. Nat. Commun. 2021, 12, 1156. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef]
- Stoletov, K.; Montel, V.; Lester, R.D.; Gonias, S.L.; Klemke, R. High-resolution imaging of the dynamic tumor cell vascular interface in transparent zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 17406–17411. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, S.; Ratnakumar, K.; Langdon, E.M.; Kansler, E.R.; Kim, I.S.; Campbell, N.R.; Perry, E.B.; McMahon, A.J.; Kaufman, C.K.; van Rooijen, E.; et al. A Quantitative System for Studying Metastasis Using Transparent Zebrafish. Cancer Res. 2015, 75, 4272. [Google Scholar] [CrossRef] [Green Version]
- Traver, D.; Winzeler, A.; Stern, H.M.; Mayhall, E.A.; Langenau, D.M.; Kutok, J.L.; Look, A.T.; Zon, L.I. Effects of lethal irradiation in zebrafish and rescue by hematopoietic cell transplantation. Blood 2004, 104, 1298–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Abdelfattah, N.S.; Blackburn, J.S.; Moore, J.C.; Martinez, S.A.; Moore, F.E.; Lobbardi, R.; Tenente, I.M.; Ignatius, M.S.; Berman, J.N. Optimized cell transplantation using adult rag2 mutant zebrafish. Nat. Methods 2014, 11, 821–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.C.; Tang, Q.; Yordán, N.T.; Moore, F.E.; Garcia, E.G.; Lobbardi, R.; Ramakrishnan, A.; Marvin, D.L.; Anselmo, A.; Sadreyev, R.I.; et al. Single-cell imaging of normal and malignant cell engraftment into optically clear prkdc-null SCID zebrafish. J. Exp. Med. 2016, 213, 2575–2589. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Brunson, D.C.; Tang, Q.; Do, D.; Iftimia, N.A.; Moore, J.C.; Hayes, M.N.; Welker, A.M.; Garcia, E.G.; Dubash, T.D.; et al. Visualizing Engrafted Human Cancer and Therapy Responses in Immunodeficient Zebrafish. Cell 2019, 177, 1903–1914.e14. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yang, Q.; Do, D.; Brunson, D.C.; Langenau, D.M. Adult immune compromised zebrafish for xenograft cell transplantation studies. EBioMedicine 2019, 47, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Glasgow, E.; Agarwal, S. Zebrafish Xenografts for Drug Discovery and Personalized Medicine. Trends Cancer 2020, 6, 569–579. [Google Scholar] [CrossRef]
- Yan, C.; Do, D.; Yang, Q.; Brunson, D.C.; Rawls, J.F.; Langenau, D.M. Single-cell imaging of human cancer xenografts using adult immunodeficient zebrafish. Nat. Protoc. 2020, 15, 3105–3128. [Google Scholar] [CrossRef]
- Wawrzyniak, M.; Scharl, M. Genetics and epigenetics of inflammatory bowel disease. Swiss Med. Wkly. 2018, 148, w14671. [Google Scholar] [CrossRef]
- Uhlig, H.H.; Powrie, F. Translating Immunology into Therapeutic Concepts for Inflammatory Bowel Disease. Annu. Rev. Immunol. 2018, 36, 755–781. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Smythies, L.E.; Sellers, M.; Clements, R.H.; Mosteller-Barnum, M.; Meng, G.; Benjamin, W.H.; Orenstein, J.M.; Smith, P.D. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J. Clin. Investig. 2005, 115, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, W.D.; Wang, Y.D. The Relationship Between Gut Microbiota and Inflammatory Diseases: The Role of Macrophages. Front. Microbiol. 2020, 11, 1065. [Google Scholar] [CrossRef]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean meal induces intestinal inflammation in zebrafish larvae. PLoS ONE 2013, 8, e69983. [Google Scholar] [CrossRef] [Green Version]
- Coronado, M.; Solis, C.J.; Hernandez, P.P.; Feijóo, C.G. Soybean Meal-Induced Intestinal Inflammation in Zebrafish Is T Cell-Dependent and Has a Th17 Cytokine Profile. Front. Immunol. 2019, 10, 610. [Google Scholar] [CrossRef]
- Kaya, B.; Doñas, C.; Wuggenig, P.; Diaz, O.E.; Morales, R.A.; Melhem, H.; Hernández, P.P.; Kaymak, T.; Das, S.; Hruz, P.; et al. Lysophosphatidic Acid-Mediated GPR35 Signaling in CX3CR1+ Macrophages Regulates Intestinal Homeostasis. Cell Rep. 2020, 32, 107979. [Google Scholar] [CrossRef] [PubMed]
- Jardine, S.; Anderson, S.; Babcock, S.; Leung, G.; Pan, J.; Dhingani, N.; Warner, N.; Guo, C.; Siddiqui, I.; Kotlarz, D.; et al. Drug Screen Identifies Leflunomide for Treatment of Inflammatory Bowel Disease Caused by TTC7A Deficiency. Gastroenterology 2020, 158, 1000–1015. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Li, H.; Liu, M.; Xie, B.; Wei, W.; Wu, J.; Meng, F.; Wang, H.Y.; Chen, S. A Manganese-Superoxide Dismutase From Thermus thermophilus HB27 Suppresses Inflammatory Responses and Alleviates Experimentally Induced Colitis. Inflamm. Bowel Dis. 2019, 25, 1644–1655. [Google Scholar] [CrossRef]
- Ge, H.; Tang, H.; Liang, Y.; Wu, J.; Yang, Q.; Zeng, L.; Ma, Z. Rhein attenuates inflammation through inhibition of NF-κB and NALP3 inflammasome in vivo and in vitro. Drug Des. Dev. Ther. 2017, 11, 1663–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Zhang, T.; Sun, H.; Liu, J.X. Copper nanoparticles induce zebrafish intestinal defects via endoplasmic reticulum and oxidative stress. Metallomics 2020, 12, 12–22. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Pasquali, L.; Cnop, M. Pancreatic β-cells in type 1 and type 2 diabetes mellitus: Different pathways to failure. Nat. Rev. Endocrinol. 2020, 16, 349–362. [Google Scholar] [CrossRef]
- Cerna, M. Epigenetic Regulation in Etiology of Type 1 Diabetes Mellitus. Int. J. Mol. Sci. 2019, 21, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Luo, Y. Recent advances of polysaccharide-based nanoparticles for oral insulin delivery. Int. J. Biol. Macromol. 2018, 120, 775–782. [Google Scholar] [CrossRef]
- Wong, C.Y.; Al-Salami, H.; Dass, C.R. Potential of insulin nanoparticle formulations for oral delivery and diabetes treatment. J. Control. Release 2017, 264, 247–275. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Bian, L.; Shen, M.; Li, X.; Zhu, J.; Chen, S.; Xiao, L.; Zhang, Q.; Chen, H.; Xu, K.; et al. Multipeptide-coupled nanoparticles induce tolerance in ‘humanised’ HLA-transgenic mice and inhibit diabetogenic CD8+ T cell responses in type 1 diabetes. Diabetologia 2017, 60, 2418–2431. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Neef, T.; Xu, D.; Podojil, J.R.; Getts, D.R.; Shea, L.D.; Miller, S.D. Tolerogenic Ag-PLG nanoparticles induce tregs to suppress activated diabetogenic CD4 and CD8 T cells. J. Autoimmun. 2018, 89, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Delaspre, F.; Beer, R.L.; Rovira, M.; Huang, W.; Wang, G.; Gee, S.; Vitery, M.d.C.; Wheelan, S.J.; Parsons, M.J. Centroacinar Cells Are Progenitors That Contribute to Endocrine Pancreas Regeneration. Diabetes 2015, 64, 3499–3509. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, R.; Rodriguez, I.; Nam, Y.H.; Hong, B.N.; Kang, T.H. Trigonelline promotes auditory function through nerve growth factor signaling on diabetic animal models. Phytomedicine 2017, 36, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Robertson, M.A.; Hesselson, D.; Stainier, D.Y.R.; Anderson, R.M. Glucagon is essential for alpha cell transdifferentiation and beta cell neogenesis. Dev. (Camb. Engl.) 2015, 142, 1407–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Covington, B.A.; Chen, W. In vivo generation and regeneration of β cells in zebrafish. Cell Regen. 2020, 9, 9. [Google Scholar] [CrossRef]
- Lu, J.; Liu, K.-C.; Schulz, N.; Karampelias, C.; Charbord, J.; Hilding, A.; Rautio, L.; Bertolino, P.; Östenson, C.-G.; Brismar, K.; et al. IGFBP1 increases β-cell regeneration by promoting α- to β-cell transdifferentiation. EMBO J. 2016, 35, 2026–2044. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.C.; Leuckx, G.; Sakano, D.; Seymour, P.A.; Mattsson, C.L.; Rautio, L.; Staels, W.; Verdonck, Y.; Serup, P.; Kume, S.; et al. Inhibition of Cdk5 Promotes β-Cell Differentiation From Ductal Progenitors. Diabetes 2018, 67, 58–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Rajpurohit, S.K.; Delaspre, F.; Walker, S.L.; White, D.T.; Ceasrine, A.; Kuruvilla, R.; Li, R.J.; Shim, J.S.; Liu, J.O.; et al. First quantitative high-throughput screen in zebrafish identifies novel pathways for increasing pancreatic β-cell mass. eLife 2015, 4, e08261. [Google Scholar] [CrossRef]
- Garcia Campoy, A.H.; Perez Gutierrez, R.M.; Manriquez-Alvirde, G.; Muñiz Ramirez, A. Protection of silver nanoparticles using Eysenhardtia polystachya in peroxide-induced pancreatic β-Cell damage and their antidiabetic properties in zebrafish. Int. J. Nanomed. 2018, 13, 2601–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Du, S.; Wang, W.; Zhang, F. Therapeutic investigation of quercetin nanomedicine in a zebrafish model of diabetic retinopathy. Biomed. Pharmacother. 2020, 130, 110573. [Google Scholar] [CrossRef]
- Singha, S.; Shao, K.; Yang, Y.; Clemente-Casares, X.; Solé, P.; Clemente, A.; Blanco, J.; Dai, Q.; Song, F.; Liu, S.W.; et al. Peptide–MHC-based nanomedicines for autoimmunity function as T-cell receptor microclustering devices. Nat. Nanotechnol. 2017, 12, 701–710. [Google Scholar] [CrossRef]
- Youssouf, L.; Bhaw-Luximon, A.; Diotel, N.; Catan, A.; Giraud, P.; Gimié, F.; Koshel, D.; Casale, S.; Bénard, S.; Meneyrol, V.; et al. Enhanced effects of curcumin encapsulated in polycaprolactone-grafted oligocarrageenan nanomicelles, a novel nanoparticle drug delivery system. Carbohydr. Polym. 2019, 217, 35–45. [Google Scholar] [CrossRef]
- Tateiwa, D.; Yoshikawa, H.; Kaito, T. Cartilage and Bone Destruction in Arthritis: Pathogenesis and Treatment Strategy: A Literature Review. Cells 2019, 8, 818. [Google Scholar] [CrossRef] [Green Version]
- Anfray, C.; Mainini, F.; Andón, F.T. Chapter 11—Nanoparticles for immunotherapy. In Frontiers of Nanoscience; Parak, W.J., Feliu, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 16, pp. 265–306. [Google Scholar]
- Gambari, L.; Grassi, F.; Roseti, L.; Grigolo, B.; Desando, G. Learning from Monocyte-Macrophage Fusion and Multinucleation: Potential Therapeutic Targets for Osteoporosis and Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 6001. [Google Scholar] [CrossRef]
- Yang, X.; Chang, Y.; Wei, W. Emerging role of targeting macrophages in rheumatoid arthritis: Focus on polarization, metabolism and apoptosis. Cell Prolif. 2020, 53, e12854. [Google Scholar] [CrossRef]
- Li, P.; Zheng, Y.; Chen, X. Drugs for Autoimmune Inflammatory Diseases: From Small Molecule Compounds to Anti-TNF Biologics. Front. Pharmacol. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, L.; Wang, Z.; Liu, P.; Liu, X.; Ding, J.; Zhou, W. Targeted silver nanoparticles for rheumatoid arthritis therapy via macrophage apoptosis and Re-polarization. Biomaterials 2020, 264, 120390. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Su, S.; Wu, Y.; Wu, Y.; Zhang, Z.; Chen, Q. Nanoparticle-facilitated delivery of BAFF-R siRNA for B cell intervention and rheumatoid arthritis therapy. Int. Immunopharmacol. 2020, 88, 106933. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Le, H.D.; Kim, T.N.T.; The, H.P.; Nguyen, T.M.; Cornet, V.; Lambert, J.; Kestemont, P. Anti-Inflammatory and Antioxidant Properties of the Ethanol Extract of Clerodendrum Cyrtophyllum Turcz in Copper Sulfate-Induced Inflammation in Zebrafish. Antioxidants 2020, 9, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Li, H.Y.; Li, X.H.; Lu, J.; Zhang, Q.; Bai, C.G.; Chen, Y. Therapeutic effects of isothiocyanate prodrugs on rheumatoid arthritis. Bioorganic Med. Chem. Lett. 2018, 28, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Xu, M.; Wang, D.; Yang, C.R.; Zeng, Y.; Zhang, Y.J. Anti-inflammatory compounds of “Qin-Jiao”, the roots of Gentiana dahurica (Gentianaceae). J. Ethnopharmacol. 2013, 147, 341–348. [Google Scholar] [CrossRef]
- Dorris, E.R.; Tazzyman, S.J.; Moylett, J.; Ramamoorthi, N.; Hackney, J.; Townsend, M.; Muthana, M.; Lewis, M.J.; Pitzalis, C.; Wilson, A.G. The Autoimmune Susceptibility Gene C5orf30 Regulates Macrophage-Mediated Resolution of Inflammation. J. Immunol. (Baltim. Md.) 2019, 202, 1069–1078. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, Z.; Meng, Q.; Jia, H.; Wang, Y.; Zhang, R. Rapid Response Fluorescence Probe Enabled In Vivo Diagnosis and Assessing Treatment Response of Hypochlorous Acid-Mediated Rheumatoid Arthritis. Adv. Sci. (Weinh. Baden-Wurtt. Ger.) 2018, 5, 1800397. [Google Scholar] [CrossRef]
- Sunke, R.; Bankala, R.; Thirupataiah, B.; Ramarao, E.V.V.S.; Kumar, J.S.; Doss, H.M.; Medishetti, R.; Kulkarni, P.; Kapavarapu, R.K.; Rasool, M.; et al. InCl3 mediated heteroarylation of indoles and their derivatization via CH activation strategy: Discovery of 2-(1H-indol-3-yl)-quinoxaline derivatives as a new class of PDE4B selective inhibitors for arthritis and/or multiple sclerosis. Eur. J. Med. Chem. 2019, 174, 198–215. [Google Scholar] [CrossRef]
- Gao, X.Y.; Li, K.; Jiang, L.L.; He, M.F.; Pu, C.H.; Kang, D.; Xie, J. Developmental toxicity of auranofin in zebrafish embryos. J. Appl. Toxicol. JAT 2017, 37, 602–610. [Google Scholar] [CrossRef]
- Stella, S.L., Jr.; Geathers, J.S.; Weber, S.R.; Grillo, M.A.; Barber, A.J.; Sundstrom, J.M.; Grillo, S.L. Neurodegeneration, Neuroprotection and Regeneration in the Zebrafish Retina. Cells 2021, 10, 633. [Google Scholar] [CrossRef]
- Lee, J.G.; Cho, H.J.; Jeong, Y.M.; Lee, J.S. Genetic Approaches Using Zebrafish to Study the Microbiota-Gut-Brain Axis in Neurological Disorders. Cells 2021, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Mehta, A.; Tong, Z.; Esser, L.; Voelcker, N.H. Development of Polymeric Nanoparticles for Blood-Brain Barrier Transfer-Strategies and Challenges. Adv. Sci. (Weinh. Baden-Wurtt. Ger.) 2021, 8, 2003937. [Google Scholar] [CrossRef]
- Zorkina, Y.; Abramova, O.; Ushakova, V.; Morozova, A.; Zubkov, E.; Valikhov, M.; Melnikov, P.; Majouga, A.; Chekhonin, V. Nano Carrier Drug Delivery Systems for the Treatment of Neuropsychiatric Disorders: Advantages and Limitations. Molecules 2020, 25, 5294. [Google Scholar] [CrossRef]
- Nellore, J.; Pauline, C.; Amarnath, K. Bacopa monnieri Phytochemicals Mediated Synthesis of Platinum Nanoparticles and Its Neurorescue Effect on 1-Methyl 4-Phenyl 1,2,3,6 Tetrahydropyridine-Induced Experimental Parkinsonism in Zebrafish. J. Neurodegener. Dis. 2013, 2013, 972391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Li, C.; Li, Y.; Yi, X.; Lee, S.M.; Zheng, Y. Oral Delivery of a Nanocrystal Formulation of Schisantherin A with Improved Bioavailability and Brain Delivery for the Treatment of Parkinson’s Disease. Mol. Pharm. 2016, 13, 3864–3875. [Google Scholar] [CrossRef]
- Hu, Q.; Guo, F.; Zhao, F.; Fu, Z. Effects of titanium dioxide nanoparticles exposure on parkinsonism in zebrafish larvae and PC12. Chemosphere 2017, 173, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, B.; Li, X.L.; Li, Y.X.; Sun, M.Z.; Chen, D.Y.; Zhao, X.; Feng, X.Z. SiO2 nanoparticles change colour preference and cause Parkinson’s-like behaviour in zebrafish. Sci. Rep. 2014, 4, 3810. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Liu, W.; Xiong, S.; Li, D.; Fang, S.; Wu, Z.; Wang, Q.; Chen, X. Nanoparticles Mediating the Sustained Puerarin Release Facilitate Improved Brain Delivery to Treat Parkinson’s Disease. ACS Appl. Mater. Interfaces 2019, 11, 45276–45289. [Google Scholar] [CrossRef]
- Xiong, S.; Liu, W.; Li, D.; Chen, X.; Liu, F.; Yuan, D.; Pan, H.; Wang, Q.; Fang, S.; Chen, T. Oral Delivery of Puerarin Nanocrystals To Improve Brain Accumulation and Anti-Parkinsonian Efficacy. Mol. Pharm. 2019, 16, 1444–1455. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiong, S.; Liu, P.; Liu, W.; Wang, Q.; Liu, Y.; Tan, H.; Chen, X.; Shi, X.; Wang, Q.; et al. Polymeric Nanoparticles-Based Brain Delivery with Improved Therapeutic Efficacy of Ginkgolide B in Parkinson’s Disease. Int. J. Nanomed. 2020, 15, 10453–10467. [Google Scholar] [CrossRef] [PubMed]
- Javed, I.; Peng, G.; Xing, Y.; Yu, T.; Zhao, M.; Kakinen, A.; Faridi, A.; Parish, C.L.; Ding, F.; Davis, T.P.; et al. Inhibition of amyloid beta toxicity in zebrafish with a chaperone-gold nanoparticle dual strategy. Nat. Commun. 2019, 10, 3780. [Google Scholar] [CrossRef] [PubMed]
- Rishitha, N.; Muthuraman, A. Therapeutic evaluation of solid lipid nanoparticle of quercetin in pentylenetetrazole induced cognitive impairment of zebrafish. Life Sci. 2018, 199, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liyanage, P.Y.; Devadoss, D.; Rios Guevara, L.R.; Cheng, L.; Graham, R.M.; Chand, H.S.; Al-Youbi, A.O.; Bashammakh, A.S.; El-Shahawi, M.S.; et al. Nontoxic amphiphilic carbon dots as promising drug nanocarriers across the blood-brain barrier and inhibitors of β-amyloid. Nanoscale 2019, 11, 22387–22397. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Watson, C.; Morsch, M.; Cole, N.J.; Chung, R.S.; Saunders, D.N.; Yerbury, J.J.; Vine, K.L. Improving the Delivery of SOD1 Antisense Oligonucleotides to Motor Neurons Using Calcium Phosphate-Lipid Nanoparticles. Front. Neurosci. 2017, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Igartúa, D.E.; Martinez, C.S.; Temprana, C.F.; Alonso, S.D.V.; Prieto, M.J. PAMAM dendrimers as a carbamazepine delivery system for neurodegenerative diseases: A biophysical and nanotoxicological characterization. Int. J. Pharm. 2018, 544, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igartúa, D.E.; Martinez, C.S.; Del, V.A.S.; Prieto, M.J. Combined Therapy for Alzheimer’s Disease: Tacrine and PAMAM Dendrimers Co-Administration Reduces the Side Effects of the Drug without Modifying its Activity. AAPS PharmSciTech 2020, 21, 110. [Google Scholar] [CrossRef] [PubMed]
- Thwaite, R.; Ji, J.; Torrealba, D.; Coll, J.; Sabés, M.; Villaverde, A.; Roher, N. Protein Nanoparticles Made of Recombinant Viral Antigens: A Promising Biomaterial for Oral Delivery of Fish Prophylactics. Front. Immunol. 2018, 9, 1652. [Google Scholar] [CrossRef]
- Sullivan, C.; Kim, C.H. Zebrafish as a model for infectious disease and immune function. Fish. Shellfish. Immunol. 2008, 25, 341–350. [Google Scholar] [CrossRef]
- Levraud, J.P.; Palha, N.; Langevin, C.; Boudinot, P. Through the looking glass: Witnessing host-virus interplay in zebrafish. Trends Microbiol. 2014, 22, 490–497. [Google Scholar] [CrossRef]
- Torraca, V.; Masud, S.; Spaink, H.P.; Meijer, A.H. Macrophage-pathogen interactions in infectious diseases: New therapeutic insights from the zebrafish host model. Dis. Model. Mech. 2014, 7, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Palha, N.; Guivel-Benhassine, F.; Briolat, V.; Lutfalla, G.; Sourisseau, M.; Ellett, F.; Wang, C.H.; Lieschke, G.J.; Herbomel, P.; Schwartz, O.; et al. Real-time whole-body visualization of Chikungunya Virus infection and host interferon response in zebrafish. PLoS Pathog. 2013, 9, e1003619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesureur, J.; Feliciano, J.R.; Wagner, N.; Gomes, M.C.; Zhang, L.; Blanco-Gonzalez, M.; van der Vaart, M.; O’Callaghan, D.; Meijer, A.H.; Vergunst, A.C. Macrophages, but not neutrophils, are critical for proliferation of Burkholderia cenocepacia and ensuing host-damaging inflammation. PLoS Pathog. 2017, 13, e1006437. [Google Scholar] [CrossRef] [PubMed]
- Bouz, G.; Al Hasawi, N. The zebrafish model of tuberculosis-no lungs needed. Crit. Rev. Microbiol. 2018, 44, 779–792. [Google Scholar] [CrossRef]
- Clay, H.; Davis, J.M.; Beery, D.; Huttenlocher, A.; Lyons, S.E.; Ramakrishnan, L. Dichotomous role of the macrophage in early Mycobacterium marinum infection of the zebrafish. Cell Host Microbe 2007, 2, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.M.; Ramakrishnan, L. The role of the granuloma in expansion and dissemination of early tuberculous infection. Cell 2009, 136, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.C.; Hsu, P.C.; Jen, C.F.; Chen, I.H.; Wang, C.H.; Chan, H.C.; Tsai, P.W.; Tung, K.C.; Wang, C.H.; Lan, C.Y.; et al. Zebrafish as a model host for Candida albicans infection. Infect. Immun. 2010, 78, 2512–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos, J.S.; Ripoll-Gomez, J.; Alfaro, J.M.; Sastre, I.; Valdivieso, F. Zebrafish as a new model for herpes simplex virus type 1 infection. Zebrafish 2008, 5, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Phennicie, R.T.; Sullivan, M.J.; Singer, J.T.; Yoder, J.A.; Kim, C.H. Specific resistance to Pseudomonas aeruginosa infection in zebrafish is mediated by the cystic fibrosis transmembrane conductance regulator. Infect. Immun. 2010, 78, 4542–4550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rounioja, S.; Saralahti, A.; Rantala, L.; Parikka, M.; Henriques-Normark, B.; Silvennoinen, O.; Rämet, M. Defense of zebrafish embryos against Streptococcus pneumoniae infection is dependent on the phagocytic activity of leukocytes. Dev. Comp. Immunol. 2012, 36, 342–348. [Google Scholar] [CrossRef]
- Schmidt, J.G.; Korbut, R.; Ohtani, M.; Jørgensen, L.V.G. Zebrafish (Danio rerio) as a model to visualize infection dynamics of Vibrio anguillarum following intraperitoneal injection and bath exposure. Fish Shellfish. Immunol. 2017, 67, 692–697. [Google Scholar] [CrossRef]
- Lin, B.; Chen, S.; Cao, Z.; Lin, Y.; Mo, D.; Zhang, H.; Gu, J.; Dong, M.; Liu, Z.; Xu, A. Acute phase response in zebrafish upon Aeromonas salmonicida and Staphylococcus aureus infection: Striking similarities and obvious differences with mammals. Mol. Immunol. 2007, 44, 295–301. [Google Scholar] [CrossRef]
- Korbut, R.; Mehrdana, F.; Kania, P.W.; Larsen, M.H.; Frees, D.; Dalsgaard, I.; Jørgensen, L. Antigen Uptake during Different Life Stages of Zebrafish (Danio rerio) Using a GFP-Tagged Yersinia ruckeri. PLoS ONE 2016, 11, e0158968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohi, O.; Parikka, M.; Rämet, M. The zebrafish as a model for paediatric diseases. Acta Paediatr. (Oslo Nor.) 2013, 102, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.V.G. Zebrafish as a Model for Fish Diseases in Aquaculture. Pathogens 2020, 9, 609. [Google Scholar] [CrossRef]
- Oksanen, K.E.; Halfpenny, N.J.; Sherwood, E.; Harjula, S.K.; Hammarén, M.M.; Ahava, M.J.; Pajula, E.T.; Lahtinen, M.J.; Parikka, M.; Rämet, M. An adult zebrafish model for preclinical tuberculosis vaccine development. Vaccine 2013, 31, 5202–5209. [Google Scholar] [CrossRef]
- Torrealba, D.; Parra, D.; Seras-Franzoso, J.; Vallejos-Vidal, E.; Yero, D.; Gibert, I.; Villaverde, A.; Garcia-Fruitós, E.; Roher, N. Nanostructured recombinant cytokines: A highly stable alternative to short-lived prophylactics. Biomaterials 2016, 107, 102–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavaliauskis, A.; Arnemo, M.; Speth, M.; Lagos, L.; Rishovd, A.L.; Estepa, A.; Griffiths, G.; Gjøen, T. Protective effect of a recombinant VHSV-G vaccine using poly(I:C) loaded nanoparticles as an adjuvant in zebrafish (Danio rerio) infection model. Dev. Comp. Immunol. 2016, 61, 248–257. [Google Scholar] [CrossRef]
- Tandberg, J.; Lagos, L.; Ropstad, E.; Smistad, G.; Hiorth, M.; Winther-Larsen, H.C. The Use of Chitosan-Coated Membrane Vesicles for Immunization against Salmonid Rickettsial Septicemia in an Adult Zebrafish Model. Zebrafish 2018, 15, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Díez-Martínez, R.; García-Fernández, E.; Manzano, M.; Martínez, Á.; Domenech, M.; Vallet-Regí, M.; García, P. Auranofin-loaded nanoparticles as a new therapeutic tool to fight streptococcal infections. Sci. Rep. 2016, 6, 19525. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanosystem | Drug | Mechanism | Tumor Type/ Cell Line | Zebrafish Stage | Injection Site | NPs Delivery | Remarkable Results | Reference |
|---|---|---|---|---|---|---|---|---|
| PtPP-HA | Kiteplatin pyrophosphate | Apoptosis through DNA platination | Breast cancer/ MDA-MB-231-GFP | 48 hpf | Duct of Cuvier | Co-injection with cells | Decrease in breast cancer cells survival | [129] |
| Zinc oxide NPs | - | Apoptosis and ROS induction | Gingival squamous cell carcinoma/ Ca9-22-DiL | 48 hpf | Yolk sac | Immersion/ 48 hpf | Dose-dependent antitumoral activity | [130] |
| PMOsPOR-NH2/TPE-PDT | - | Porphyrin photosensitivity and ROS production | Breast Cancer/ MDA-MB-231-GFP | 30 hpf | Duct of Cuvier | Pre-treatment of cancer cells | Complete extinction of cancer cells | [131] |
| PORBSNs/ TPE-PDT | - | Porphyrin photosensitivity and ROS production | Breast cancer/ MDA-MB-231-GFP | 24-30 hpf | Perivitelline space | Intravenously/ 4 dpi | Decrease of the tumor area | [132] |
| PAMAM-GC/DOX/γ-radiation | DOX | GC radiosensitivity increases DOX release | Uterine cervical carcinoma/ HeLa- CSFE | 48 hpf | Yolk sac | Immersion/ 1 dpi | Synergistic antitumoral effect for the combination of GC/DOX and radiotherapy | [133] |
| PAMAM-DOX-siHIF | DOX/siHIF | NPs responsiveness to hypoxia and increased drug release | Breast cancer/ MCF-7-CM-DiL | 48 hpf | Perivitelline space | Intracardiac injection/1 dpi | Feasibility of the cooperative strategy for in vivo applications | [134] |
| NanogelDOX | DOX | Hydrazone sensitivity to pH accelerate drug release | Melanoma/ B6-RFP or GPF | 48 hpf | Neural tube | Intravenously/ 1 dpi | Selective accumulation of the NPs in the tumor and reduction in tumor growth | [135] |
| Tf-DOX-ReSi-Au | DOX | Enhanced tumor targeting by interaction between Tf and Tf receptor | Colorectal cancer/ HCT116-GFP | 48 hpf | Yolk sac | Retro-orbital injection/1 dpi | Antitumoral activity without DOX-related cardiotoxic effects | [136] |
| PEG liposomes | - | - | Melanoma/ Melmet 5-dsRed Kidney/HEK293-mCherry | 48 hpf | Duct of Cuvier | Intravenously/ 2 dpi | NPs accumulation in human tumor structure, low macrophage uptake and high survival rate | [72] |
| PEG-PDPA-DOX | DOX | Polymersomes release the drug only at low pH | Melanoma/ B6-RFP or GPF | 72 hpf | Neural tube | Intravenously/ 1 dpi | Selective accumulation of NPs in the tumor area, increased cancer cell apoptosis and reduced proliferation | [137] |
| Disease | Disease Induction | Zebrafish Stage | Nanosystem | NPs Delivery | Remarkable Results | Reference |
|---|---|---|---|---|---|---|
| Inflammatory Bowel Disease | Copper NPs- induced intestinal developmental defects | From 0 hpf | Copper NPs | Immersion | CuNPs cause intestinal developmental defects via inducing ER stress and ROS generation, which corresponds with elevated serum copper levels in IBD patients | [174] |
| Type 1 diabetes mellitus | Glucose-induced diabetic zebrafish | Adult | EP/AgNPs | Immersion | Hyperglycemia amelioration | [189] |
| Type 1 diabetes mellitus | STZ- induced diabetic retinopathy | Adult | Quercetin NPs | Intraperitoneal injection | Reduction of glycemia and improvement of morphological, behavioral and biochemical parameters linked to retinopathy | [190] |
| Type 1 diabetes mellitus | - | From 4 hpf | pMHC-NPs | Immersion | Neither off-target toxicity, nor morphological abnormalities | [191] |
| Parkinson’s Disease | MPTP-induced parkinsonism | Adult | BmE-PtNPs | Intraperitoneal injection | Significant reversion of toxic effects of MPTP by increasing the levels of dopamine, GSH, GPx, catalase, SOD and complex I, and reducing levels of MDA | [211] |
| Parkinson’s Disease | MPTP-induced parkinsonism | From 72 hpf | Schisantherin nanocrystals | Immersion | Reversed dopaminergic neuronal loss and locomotion deficiency by the activation of the Akt/Gsk3β pathway | [212] |
| Parkinson’s Disease | - | From 0 hpf | Titanium dioxide NPs | Immersion | Loss of dopaminergic neurons, ROS generation and cell death in hypothalamus. Increased Lewy bodies-related markers. | [213] |
| Parkinson’s Disease | - | Adult | Silica NPs | Immersion | Changes in dopaminergic neurons with disturbed light/dark preference, dampened exploratory behavior, inhibited memory capability and PD-like behavior | [62,214] |
| Parkinson’s Disease | - | From 6 hpf | Puerarin Nanocristals | Immersion | Promising anti-Parkinsonism NCs. Unaltered morphology, hatching, survival rate, body length and heart rate | [215,216] |
| Parkinson’s Disease | - | From 120 hpf | Ginkgolide B-PEG-PCL NPs | Immersion | Correct bioavailability and cerebral accumulation in zebrafish models | [217] |
| Alzheimer’s Disease | Aβ- induced toxicity | Adult | Casein coated-gold NPs | Retro-orbital injection | Inhibition of Aβ toxicity and recover of the mobility and cognitive function | [218] |
| Alzheimer’s Disease | PTZ- induced cognitive impairment | Adult | Solid lipid NPs of Quercetin | Intraperitoneal injection | Attenuation of PTZ-induced neurocognitive impairments and amelioration of biochemical changes | [219] |
| Amyotrophic Lateral Sclerosis | - | From 96 hpf | ASO- CaP-lipid NPs | Brain, spinal cord, intravenous and retro-orbital injection | Successful delivery and preferential accumulation in brain, bloodstream and spinal cord | [221] |
| Disease Induction | Zebrafish Stage | Nanosystem | NPs Delivery | Immune Cells Behavior | Remarkable Results | Reference |
|---|---|---|---|---|---|---|
| Pseudomonas aeruginosa infection | Adult | Nanostructured cytokines (IBTNFα, IBCCL4) | Intraperitoneal injection | Interaction of IBs with immune cells | Prophylactic potential in vivo | [243] |
| - | Adult | IPNV, VHSV and VNNV-encapsulated IBs | Oral intubation | - | Successful NPs uptake in gut cells after oral administration | [224] |
| VHSV infection | Adult | Viral glycoprotein G encapsulated in chitosan-poly(I:C) NPs | Intraperitoneal injection | Upregulation of antiviral cytokines | Significant protection against VHSV through induction of anti-viral state | [244] |
| Piscirickettsia salmonis infection | Adult | Chitosan-coated MVs from P. salmonis | Intraperitoneal injection | Upregulation of immune related genes | Successful immunisation. Potential use of chitosan-coated MVs for vaccination | [245] |
| Streptococcus pneumonia infection | From 48 hpf | Auranofin-PLGA-NPs | Immersion | - | Auranofin-NPs capability of decreasing the bacterial population compared to free drug | [246] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pensado-López, A.; Fernández-Rey, J.; Reimunde, P.; Crecente-Campo, J.; Sánchez, L.; Torres Andón, F. Zebrafish Models for the Safety and Therapeutic Testing of Nanoparticles with a Focus on Macrophages. Nanomaterials 2021, 11, 1784. https://doi.org/10.3390/nano11071784
Pensado-López A, Fernández-Rey J, Reimunde P, Crecente-Campo J, Sánchez L, Torres Andón F. Zebrafish Models for the Safety and Therapeutic Testing of Nanoparticles with a Focus on Macrophages. Nanomaterials. 2021; 11(7):1784. https://doi.org/10.3390/nano11071784
Chicago/Turabian StylePensado-López, Alba, Juan Fernández-Rey, Pedro Reimunde, José Crecente-Campo, Laura Sánchez, and Fernando Torres Andón. 2021. "Zebrafish Models for the Safety and Therapeutic Testing of Nanoparticles with a Focus on Macrophages" Nanomaterials 11, no. 7: 1784. https://doi.org/10.3390/nano11071784