
Ergosterol Isolated from Antrodia camphorata Suppresses LPS-Induced Neuroinflammatory Responses in Microglia Cells and ICR Mice
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Ergosterol Inhibits LPS-Induced Microglial Inflammation
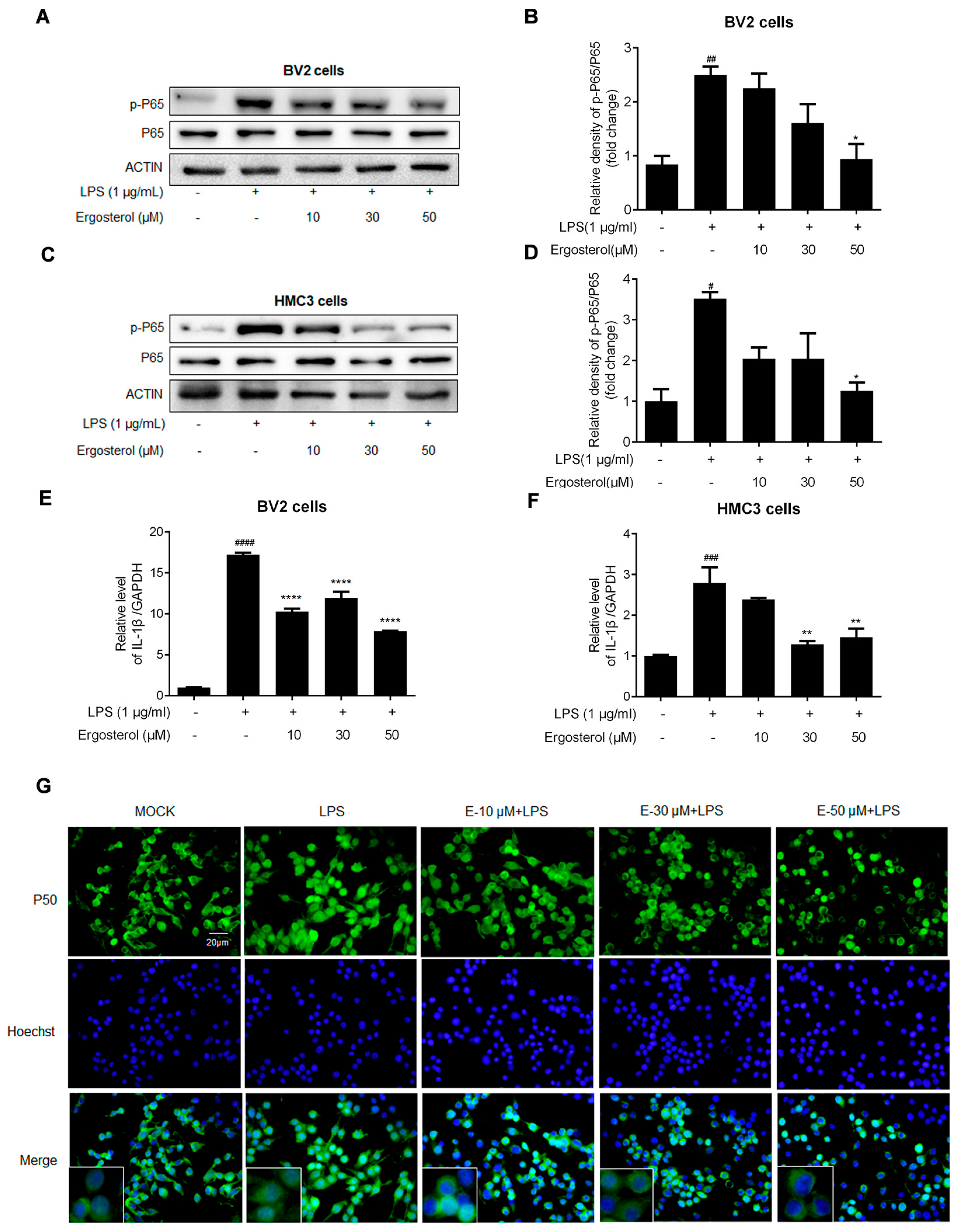
2.2. Ergosterol Suppresses NF-κB Signaling Activated by LPS in BV2 and HMC3 Microglial Cells
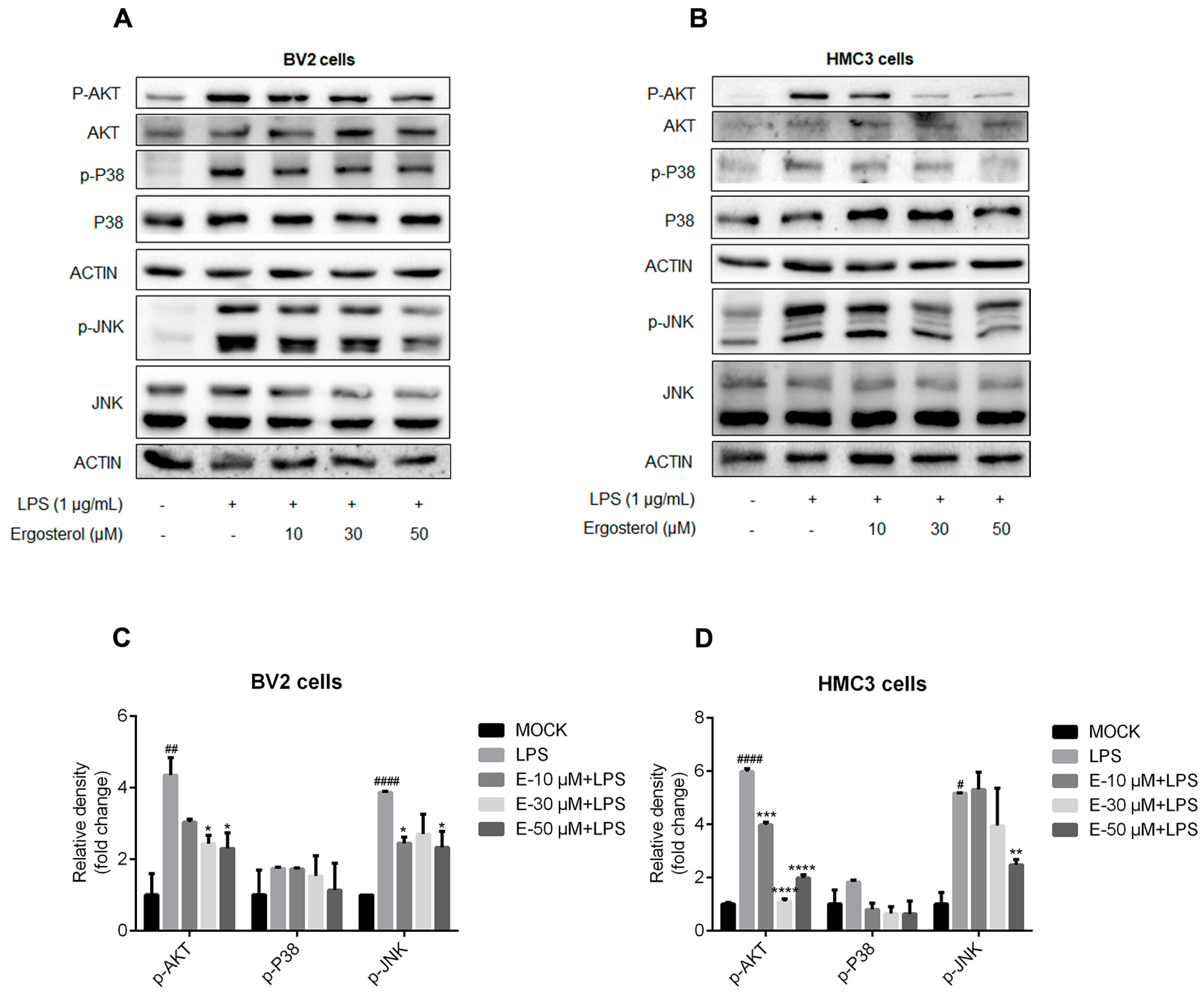
2.3. Ergosterol Inhibits the Activation of MAPK and AKT Signaling Pathways Induced by LPS in Microglial Cells
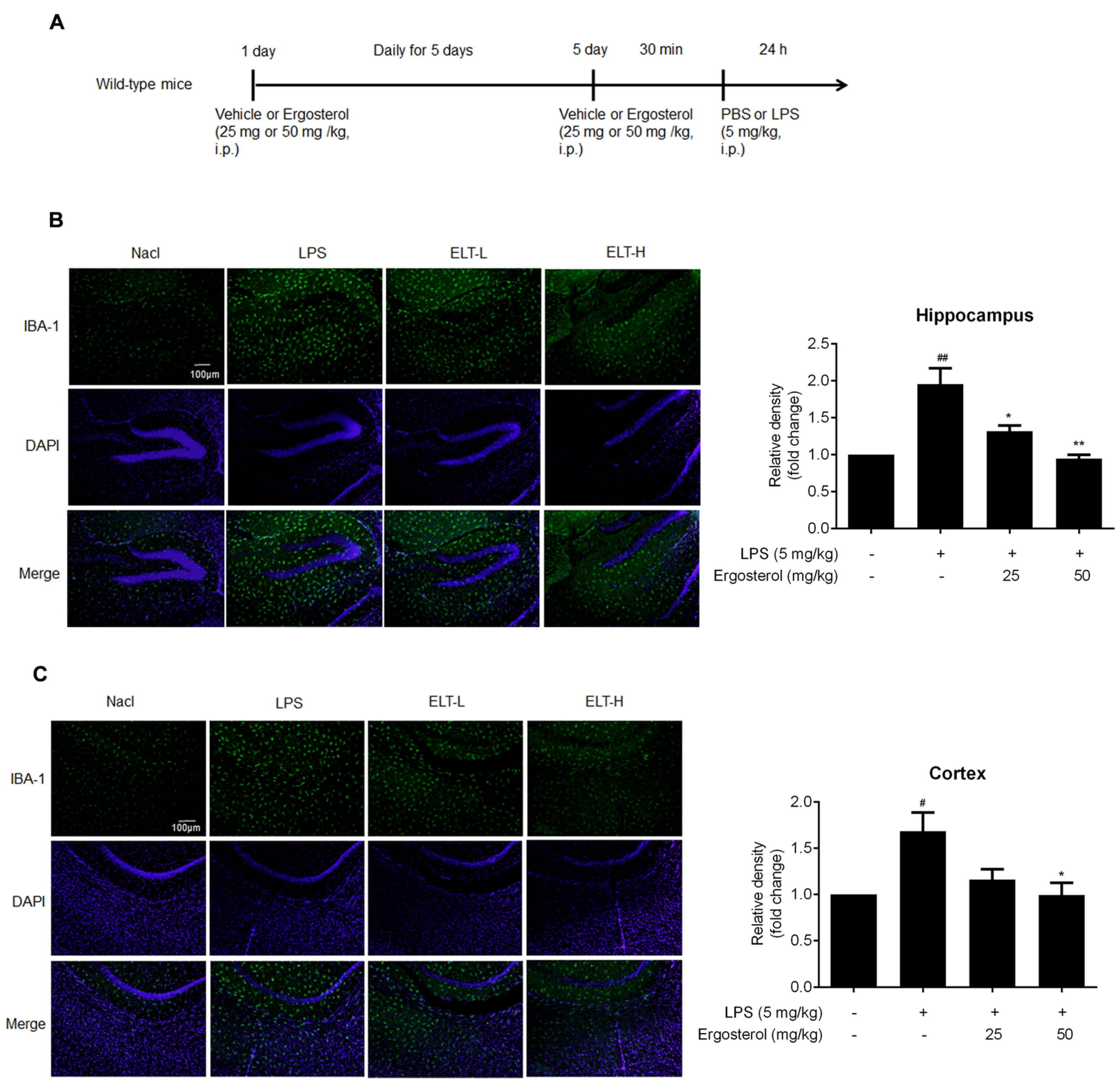
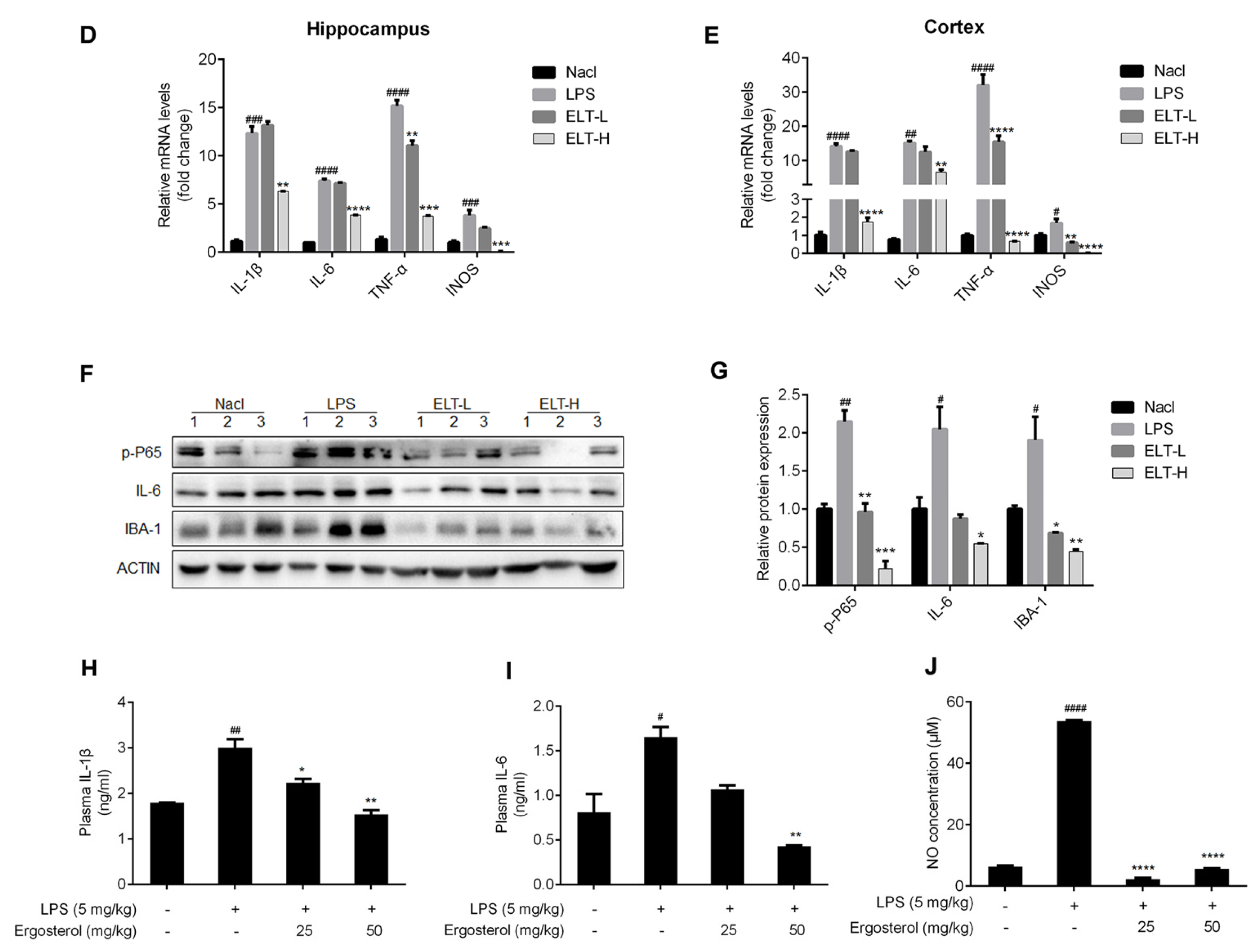
2.4. Ergosterol Suppresses Neuroinflammation in LPS-Treated ICR Mice
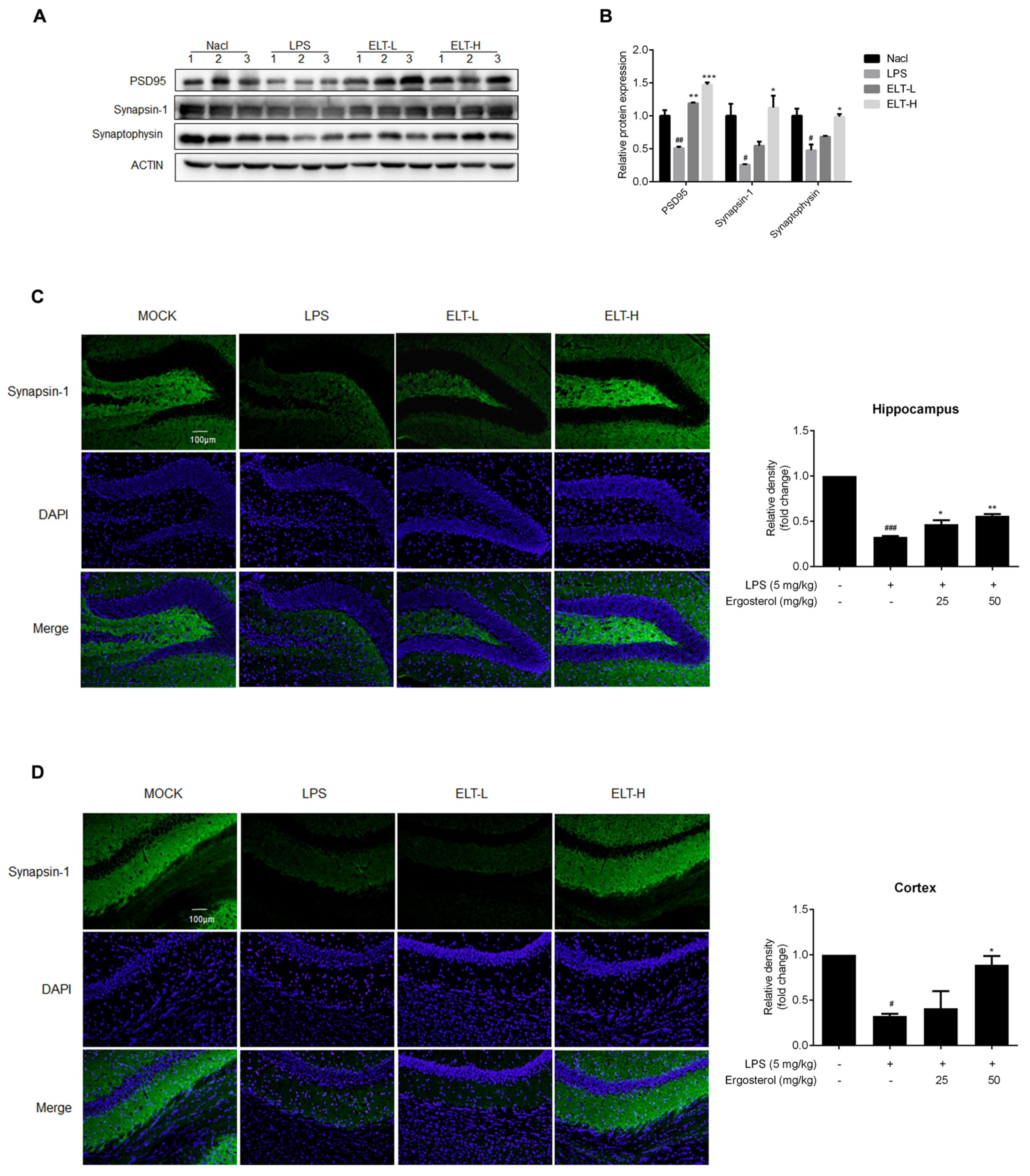
2.5. Ergosterol Reduces Neuron Damage in LPS-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Compounds
4.3. Antibodies and Inhibitors
4.4. Animal and Treatment
4.5. Immunohistochemistry
4.6. Cell Viability
4.7. Quantitative Real-Time PCR Assay
4.8. Enzyme-Linked Immunosorbent Assay
4.9. Western Blot
4.10. Griess Assay (NO Assay)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D. Astrocytic and microglial cells as the modulators of neuroinflammation in Alzheimer’s disease. J. Neuroinflamm. 2022, 19, 206. [Google Scholar] [CrossRef] [PubMed]
- Gillardon, F.; Schmid, R.; Draheim, H. Parkinson’s disease-linked leucine-rich repeat kinase 2(R1441G) mutation increases pro-inflammatory cytokine release from activated primary microglial cells and resultant neurotoxicity. Neuroscience 2012, 208, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Javanmehr, N.; Saleki, K.; Alijanizadeh, P.; Rezaei, N. Microglia dynamics in aging-related neurobehavioral and neuroinflam-matory diseases. J. Neuroinflamm. 2022, 19, 273. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-S.; An, J.; Li, J.-J.; Han, L.; Xing, Z.-B.; Li, C.-Q. Piperine attenuates lipopolysaccharide (LPS)-induced inflammatory responses in BV2 microglia. Int. Immunopharmacol. 2017, 42, 44–48. [Google Scholar] [CrossRef]
- Joo, S.S.; Yoo, Y.M.; Ahn, B.W.; Nam, S.Y.; Kim, Y.-B.; Hwang, K.W.; Lee, D.I. Prevention of inflammation-mediated neurotoxicity by Rg3 and its role in microglial activation. Biol. Pharm. Bull. 2008, 31, 1392–1396. [Google Scholar] [CrossRef]
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflam. 2017, 14, 1. [Google Scholar] [CrossRef]
- Liu, M.; Chen, F.; Sha, L.; Wang, S.; Tao, L.; Yao, L.; He, M.; Yao, Z.; Liu, H.; Zhu, Z.; et al. (−)-Epigallocatechin-3-gallate ameliorates learning and memory deficits by adjusting the balance of TrkA/p75NTR signaling in APP/PS1 transgenic mice. Mol. Neurobiol. 2013, 49, 1350–1363. [Google Scholar] [CrossRef]
- Kielian, T. Toll-like receptors in central nervous system glial inflammation and homeostasis. J. Neurosci. Res. 2006, 83, 711–730. [Google Scholar] [CrossRef]
- Aksoy, E.; Taboubi, S.; Torres, D.; Delbauve, S.; Hachani, A.; Whitehead, M.A.; Pearce, W.P.; Berenjeno, I.M.; Nock, G.; Filloux, A.; et al. The p110δ isoform of the kinase PI(3)K controls the subcellular compartmentali-zation of TLR4 signaling and protects from endotoxic shock. Nat. Immunol. 2012, 13, 1045–1054. [Google Scholar] [CrossRef]
- Laird, M.H.; Rhee, S.H.; Perkins, D.J.; Medvedev, A.E.; Piao, W.; Fenton, M.J.; Vogel, S.N. TLR4/MyD88/PI3K interactions regulate TLR4 signaling. J. Leukoc. Biol. 2009, 85, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.T.; Bian, C.; Yuan, J.C.; Chu, W.H.; Xiang, X.; Chen, F.; Wang, C.S.; Feng, H.; Lin, J.K. Curcumin attenuates acute inflammatory injury by inhibiting the TLR4/MyD88/NF-κB signaling pathway in experimental traumatic brain injury. J. Neuroinflamm. 2014, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhabinskii, V.N.; Drasar, P.; Khripach, V.A. Structure and biological activity of ergostane-type steroids from fungi. Molecules 2022, 27, 2103. [Google Scholar] [CrossRef] [PubMed]
- Loria-Kohen, V.; Lourenço-Nogueira, T.; Espinosa-Salinas, I.; Marín, F.R.; Soler-Rivas, C.; de Molina, A.R. Nutritional and functional properties of edible mushrooms: A food with promising health claims. J. Pharm. Nutr. Sci. 2014, 4, 187–198. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Xu, L.T.; Li, A.X.; Wang, S.M. Effects of ergosterol, isolated from scleroderma polyrhizum pers., on lipopolysaccha-ride-induced inflammatory responses in acute lung injury. Inflammation 2015, 38, 1979–1985. [Google Scholar] [CrossRef]
- Nallathamby, N.; Guan-Serm, L.; Vidyadaran, S.; Malek, S.N.A.; Raman, J.; Sabaratnam, V. Ergosterol of Cordyceps militaris attenuates LPS induced inflammation in BV2 microglia cells. Nat. Prod. Commun. 2015, 10, 885–886. [Google Scholar] [CrossRef]
- Kuo, C.F.; Hsieh, C.H.; Lin, W.Y. Proteomic response of LAP-activated RAW 264.7 macrophages to the anti-inflammatory prop-erty of fungal ergosterol. Food Chem. 2011, 126, 207–212. [Google Scholar] [CrossRef]
- Yang, J.; Wang, T.; Jin, X.; Wang, G.; Zhao, F.; Jin, Y. Roles of crosstalk between astrocytes and microglia in triggering neuroin-flammation and brain edema formation in 1,2-dichloroethane-intoxicated mice. Cells 2021, 10, 2647. [Google Scholar] [CrossRef]
- Sillapachaiyaporn, C.; Chuchawankul, S.; Nilkhet, S.; Moungkote, N.; Sarachana, T.; Ung, A.T.; Baek, S.J.; Tencomnao, T. Ergosterol isolated from cloud ear mushroom (Auricularia polytricha) attenuates bisphenol A-induced BV2 microglial cell inflamma-tion. Food Res. Int. 2022, 157, 111433. [Google Scholar] [CrossRef]
- Sun, X.; Feng, X.; Zheng, D.; Li, A.; Li, C.; Li, S.; Zhao, Z. Ergosterol attenuates cigarette smoke extract-induced COPD by modulat-ing inflammation, oxidative stress and apoptosis in vitro and in vivo. Clin. Sci. 2019, 133, 1523–1536. [Google Scholar] [CrossRef]
- Disabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The devil is in the details. J. Neurochem. 2016, 139 (Suppl. 2), 136–153. [Google Scholar] [CrossRef]
- Lindhout, I.A.; Murray, T.E.; Richards, C.M.; Klegeris, A. Potential neurotoxic activity of diverse molecules released by microglia. Neurochem. Int. 2021, 148, 105117. [Google Scholar] [CrossRef] [PubMed]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol. Med. 2019, 11, e10248. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Neumann, H. Role of microglia in neuronal degeneration and regeneration. Semin. Immunopathol. 2009, 31, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, F.; Liu, F.; Li, D.; Yang, T. NRF2/HO-1 activation via ERK pathway involved in the anti-neuroinflammatory effect of Astragaloside IV in LPS induced microglial cells. Neurosci. Lett. 2018, 666, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Xu, Z.; Zhang, W.; Wilson, B.; Hong, J.-S.; Flood, P.M. Sinomenine, a natural dextrorotatory morphinan analog, is anti-inflammatory and neuroprotective through inhibition of microglial NADPH oxidase. J. Neuroinflamm. 2007, 4, 23. [Google Scholar] [CrossRef]
- Yang, H.L.; Korivi, M.; Chen, C.H.; Peng, W.J.; Chen, C.S.; Li, M.L.; Hsu, L.S.; Liao, J.W.; Hseu, Y.C. Antrodia camphorata attenuates ciga-rette smoke-induced ROS production, DNA damage, apoptosis, and inflammation in vascular smooth muscle cells, and ath-erosclerosis in ApoE-deficient mice. Environ. Toxicol. 2017, 32, 2070–2084. [Google Scholar] [CrossRef]
- Huang, G.J.; Deng, J.S.; Chen, C.C.; Huang, C.J.; Sung, P.J.; Huang, S.S.; Kuo, Y.H. Methanol extract of Antrodia camphorata protects against lipopolysaccharide-induced acute lung injury by suppressing NF-κB and MAPK pathways in mice. J. Agric. Food Chem. 2014, 62, 5321–5329. [Google Scholar] [CrossRef]
- Kuang, Y.; Li, B.; Wang, Z.; Qiao, X.; Ye, M. Terpenoids from the medicinal mushroom Antrodia camphorata: Chemistry and me-dicinal potential. Nat. Prod. Rep. 2021, 38, 83–102. [Google Scholar] [CrossRef]
- Chao, T.Y.; Hsieh, C.C.; Hsu, S.M.; Wan, C.H.; Lian, G.T.; Tseng, Y.H.; Kuo, Y.H.; Hsieh, S.C. Ergostatrien-3β-ol (EK100) from Antrodia camphorata attenuates oxidative stress, inflammation, and liver injury in vitro and in vivo. Prev. Nutr. Food Sci. 2021, 26, 58–66. [Google Scholar] [CrossRef]
- -Miklossy, J. Chronic inflammation and amyloidogenesis in Alzheimer’s disease role of spirochetes. JAD 2008, 13, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Remick, D.; Newcomb, D.E.; Bolgos, G.L.; Call, D.R. Comparison of the mortality and inflammatory response of two models of sepsis: Lipopolysaccharide vs. Cecal ligation and puncture. Shock 2000, 13, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Ji, H.M.; Kim, D.W.; Choi, S.-M.; Kim, S.; Yang, E.J. Effects of Hominis placenta on LPS-induced cell toxicity in BV2 microglial cells. J. Ethnopharmacol. 2013, 147, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Matamoros-Recio, A.; Franco-Gonzalez, J.F.; Perez-Regidor, L.; Billod, J.M.; Guzman-Caldentey, J.; Martin-Santamaria, S. Full-atom model of the agonist LPS-bound Toll-like receptor 4 dimer in a membrane environment. Chemistry 2021, 27, 15406–15425. [Google Scholar] [CrossRef]
- Streit, W.J.; Xue, Q.-S. Life and death of microglia. J. Neuroimmune Pharmacol. 2009, 4, 371–379. [Google Scholar] [CrossRef]
- An, J.; Chen, B.; Kang, X.; Zhang, R.; Guo, Y.; Zhao, J.; Yang, H. Neuroprotective effects of natural compounds on LPS-induced in-flammatory responses in microglia. Am. J. Transl. Res. 2020, 12, 2353–2378. [Google Scholar]
- Doherty, G.H. Nitric oxide in neurodegeneration: Potential benefits of non-steroidal anti-inflammatories. Neurosci. Bull. 2011, 27, 366–382. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-S.; Yang, X.; Yeung, S.-C.; Chiu, K.; Lau, C.-F.; Tsang, A.W.-T.; Mak, J.C.-W.; Chang, R.C.-C. Cigarette smoking accelerated brain aging and induced pre-alzheimer-like neuropathology in rats. PLoS ONE 2012, 7, e36752. [Google Scholar] [CrossRef]
- Hedegaard, C.; Kjaer-Sorensen, K.; Madsen, L.B.; Henriksen, C.; Momeni, J.; Bendixen, C.; Oxvig, C.; Larsen, K. Porcine synapsin 1:SYN1 gene analysis and functional characterization of the promoter. FEBS Open Bio. 2013, 3, 411–420. [Google Scholar] [CrossRef]
- Meyer, D.; Bonhoeffer, T.; Scheuss, V. Balance and stability of synaptic structures during synaptic plasticity. Neuron 2014, 82, 430–443. [Google Scholar] [CrossRef]
- Cheng, P.C.; Hsu, C.-Y.; Chen, C.-C.; Lee, K.-M. In vivo immunomodulatory effects of Antrodia camphorata polysaccharides in a T1/T2 doubly transgenic mouse model for inhibiting infection of Schistosoma mansoni. Toxicol. Appl. Pharmacol. 2008, 227, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Kathy, K.; Auyeung Quan-Bin Han Joshua, K.K. Astragalus membranaceus: A review of its protection against inflamma-tion and gastrointestinal cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar]
- Wang, R.; Wu, X.; Liu, L.; An, Y. Activity directed investigation on anti-inflammatory fractions and compounds from flowers of Trollius chinensis. Pak. J. Pharm. Sci. 2014, 27, 285–288. [Google Scholar]
- Kwon, H.C.; Zee, S.D.; Cho, S.Y.; Choi, S.U.; Lee, K.R. Cytotoxic ergosterols from Paecilomyces sp. J300. Arch. Pharm. Res. 2002, 25, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Morrison, H. Quantifying microglia morphology from photomicrographs of immunohistochemistry prepared tissue using imagej. J. Vis. Exp. 2018, 136, 57648. [Google Scholar]
- Gallo-Oller, G.; Ordoñez, R.; Dotor, J. A new background subtraction method for Western blot densitometry band quantifica-tion through image analysis software. J. Immunol. Methods 2018, 457, 1–5. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Manufacturer | Catalog No. |
|---|---|---|
| P50 | Proteintech | 14220-1-AP |
| P65 | Cell Signaling | 8242S |
| p-P65 | Cell Signaling | 3033S |
| AKT | Cell Signaling | 9272S |
| p-AKT | Cell Signaling | 4060S |
| P38 | Cell Signaling | 8690S |
| p-P38 | Cell Signaling | 4511S |
| JNK | Cell Signaling | 9252S |
| p-JNK | Cell Signaling | 9255S |
| INOS | Proteintech | 22226-1-AP |
| Beta-actin | Proteintech | 81115-1-RR |
| COX-2 | Absin | abs131986 |
| IL-6 | Abcam | ab290735 |
| IBA-1 | Abcam | ab178846 |
| Synapsin-1 | Cell Signaling | 5297S |
| Synaptophysin | Cell Signaling | 25056S |
| PSD95 | Cell Signaling | 3450S |
| Primer | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| mIL-1β | GCTGCTTCCAAACCTTTGAC | AGCTTCTCCACAGCCACAAT |
| mGAPDH | AGAACATCATCCCTGCATCC | CACATTGGGGGTAGGAACAC |
| mTNF-α | CCGATGGGTTGTACCTTGTC | CCGATGGGTTGTACCTTGTC |
| mIL-18 | ACGTGTTCCAGGACACAACA | GGCGCATGTGTGCTAATCAT |
| mIL-6 | CCACTTCACAAGTCGGAGGC | GGAGAGCATTGGAAATTGGGGT |
| mINOS | CCGGCAAACCCAAGGTCTAC | GCATTTCGCTGTCTCCCCAA |
| hIL-1β | ATGATGGCTTATTACAGTGGCAA | GTCGGAGATTCGTAGCTGGA |
| hGAPDH | CTGCACCACCAACTGCTT | TTCTGGGTGGCAGTGATG |
| hTNF-α | TATGGCTCAGGGTCCAACTC | GGAAAGCCCATTTGAGTCCT |
| hIL-6 | ACTCACCTCTTCAGAACGAATTG | CCATCTTTGGAAGGTTCAGGTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, P.; Li, W.; Guo, J.; Peng, Q.; Ye, X.; Hu, S.; Liu, Y.; Liu, W.; Chen, H.; Qiao, J.; et al. Ergosterol Isolated from Antrodia camphorata Suppresses LPS-Induced Neuroinflammatory Responses in Microglia Cells and ICR Mice. Molecules 2023, 28, 2406. https://doi.org/10.3390/molecules28052406
Sun P, Li W, Guo J, Peng Q, Ye X, Hu S, Liu Y, Liu W, Chen H, Qiao J, et al. Ergosterol Isolated from Antrodia camphorata Suppresses LPS-Induced Neuroinflammatory Responses in Microglia Cells and ICR Mice. Molecules. 2023; 28(5):2406. https://doi.org/10.3390/molecules28052406
Chicago/Turabian StyleSun, Ping, Weiling Li, Jiazheng Guo, Qian Peng, Xiansheng Ye, Song Hu, Yuchen Liu, Wei Liu, Haifeng Chen, Jialu Qiao, and et al. 2023. "Ergosterol Isolated from Antrodia camphorata Suppresses LPS-Induced Neuroinflammatory Responses in Microglia Cells and ICR Mice" Molecules 28, no. 5: 2406. https://doi.org/10.3390/molecules28052406