
Tetrabromobisphenol Exposure Impairs Bovine Oocyte Maturation by Inducing Mitochondrial Dysfunction
 ,
,
Abstract
:1. Introduction
2. Results
2.1. TBBPA Exposure Disrupts Bovine Oocyte Maturation and Cumulus-Cell Expansion
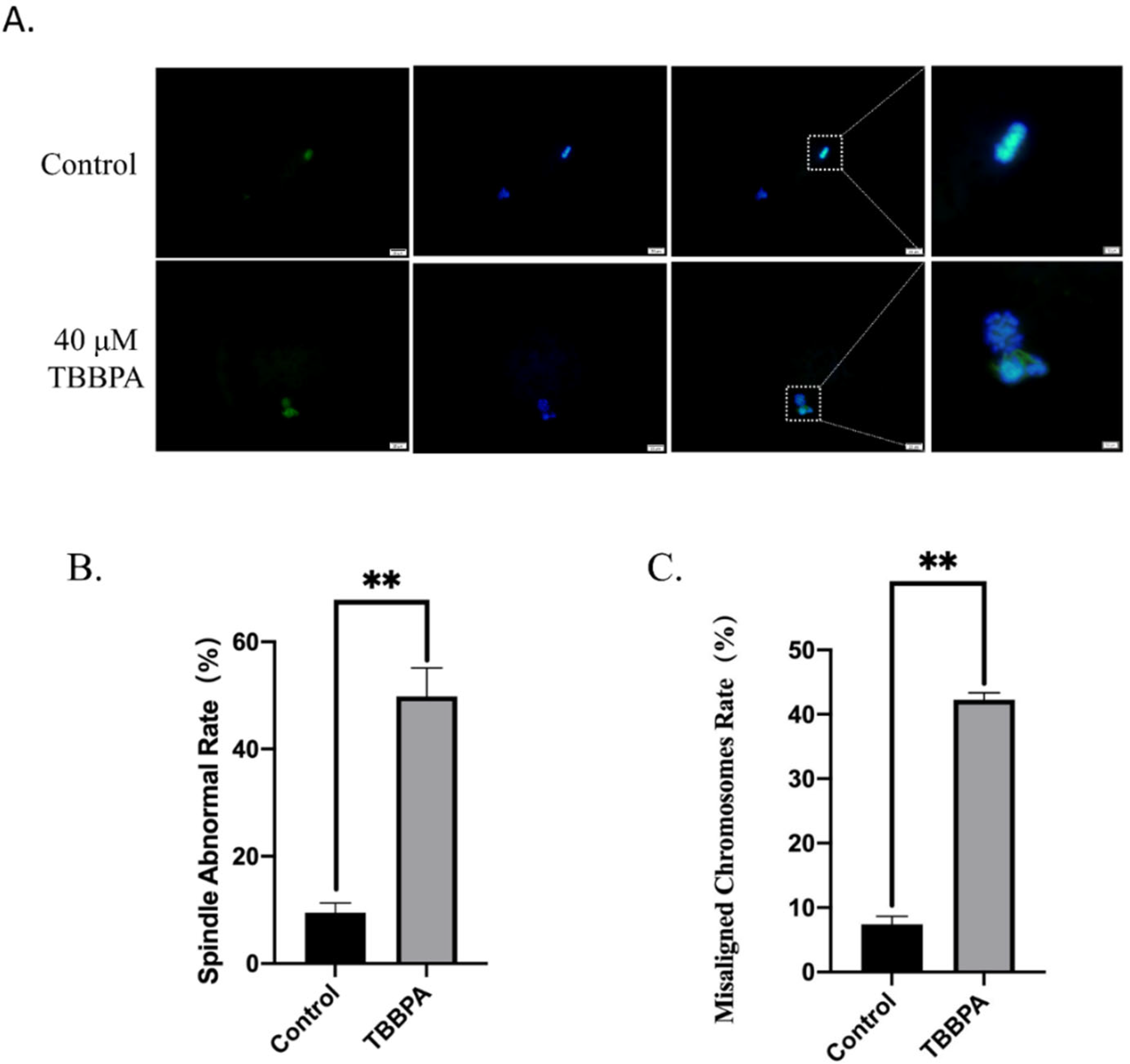
2.2. TBBPA Exposure Destroys Spindle Morphology and Chromosome Alignment during Bovine Oocyte Maturation
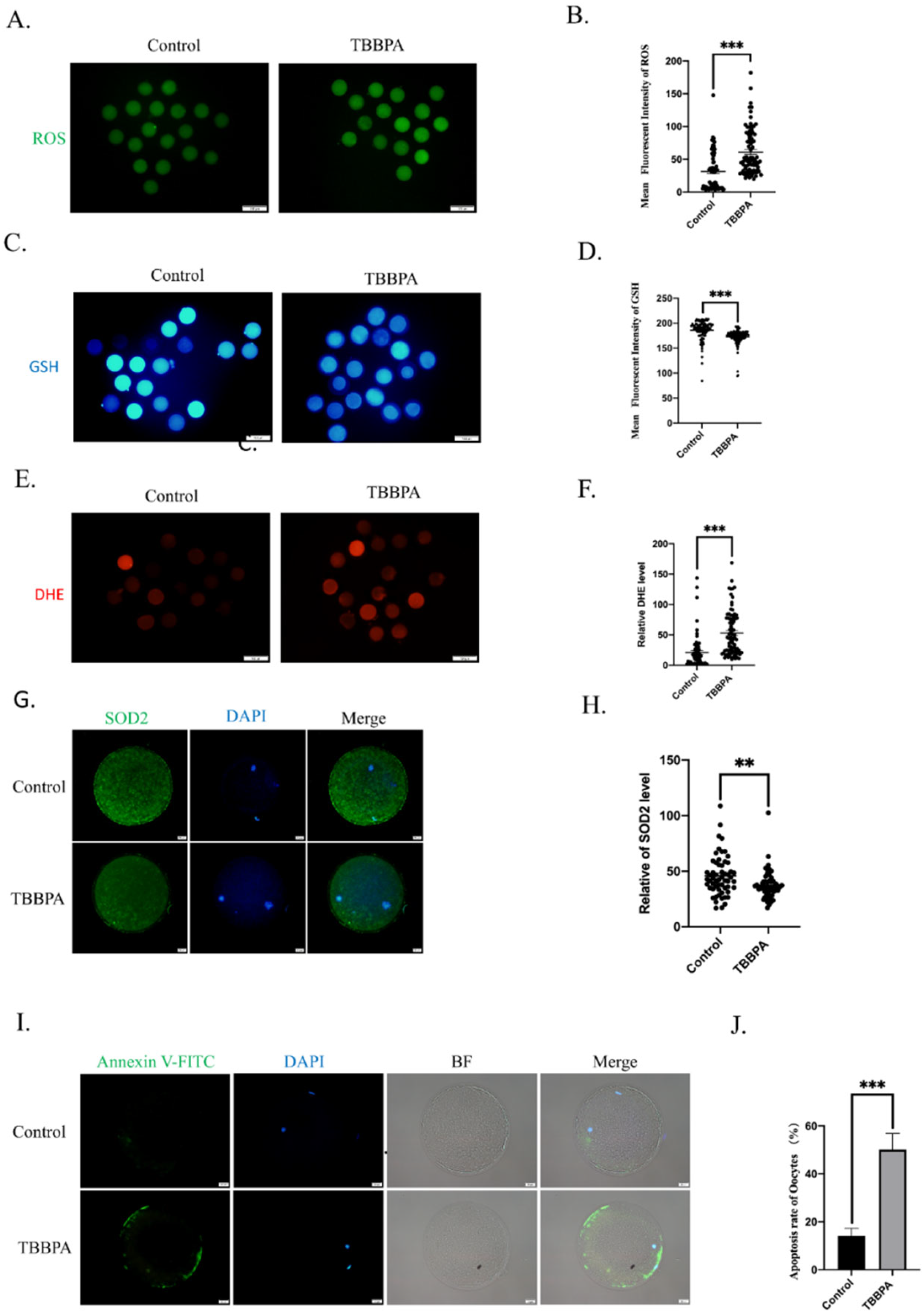
2.3. TBBPA Exposure Causes Oxidative Stress to Induce Early Apoptosis
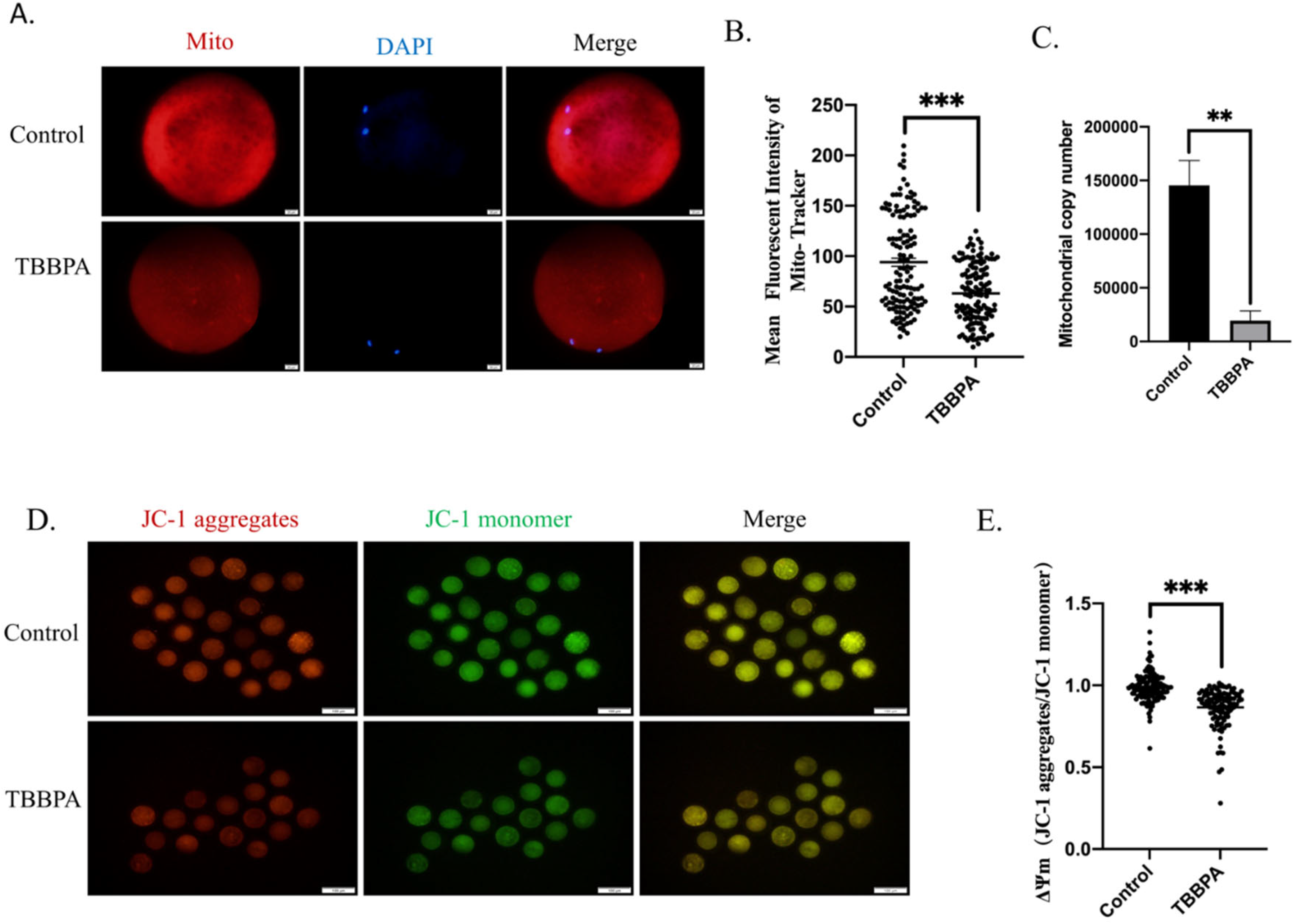
2.4. TBBPA Exposure Induces the Mitochondrial Dysfunction
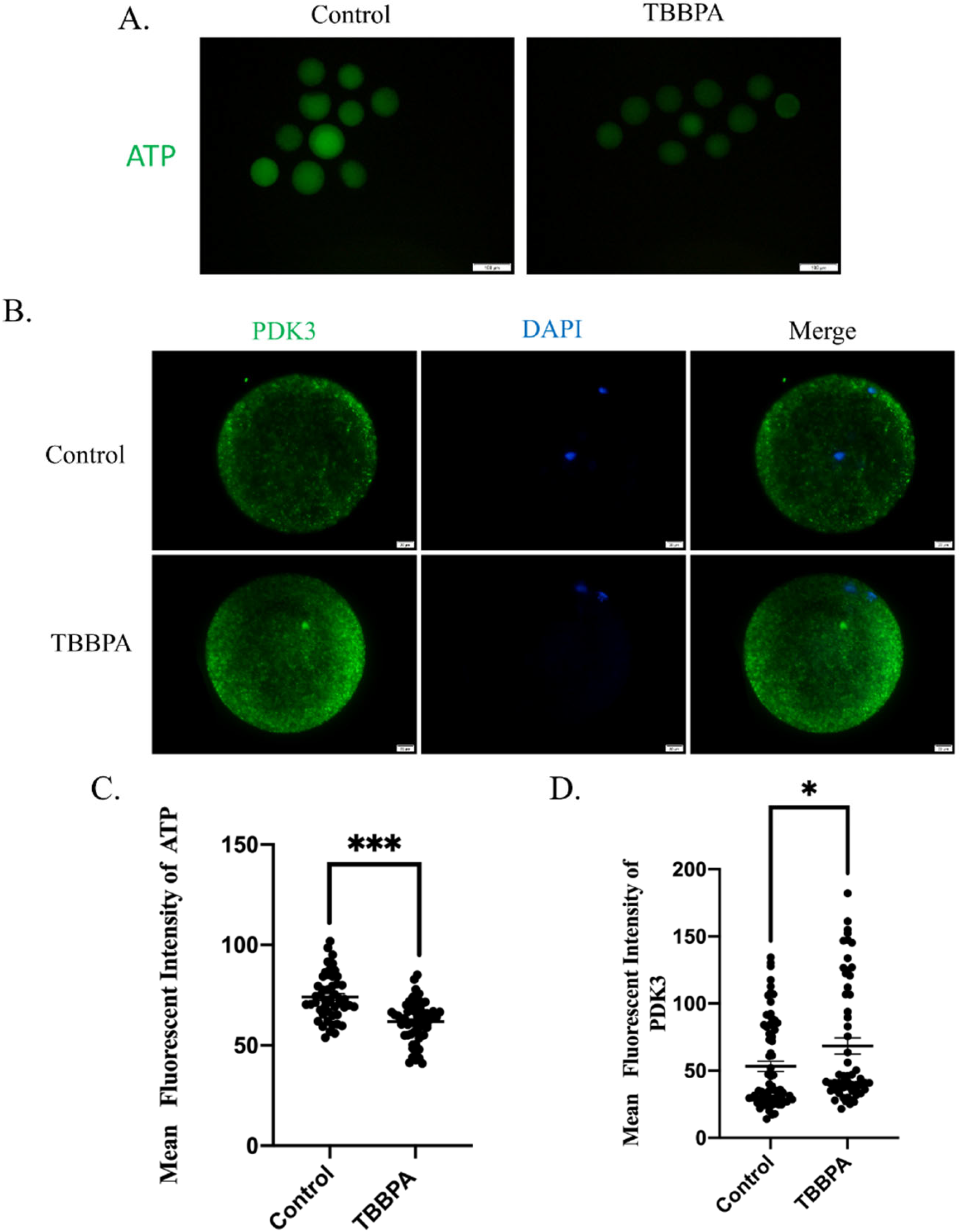
2.5. TBBPA Exposure Causes Mitochondrial Dysfunction by Regulating PDK Activity
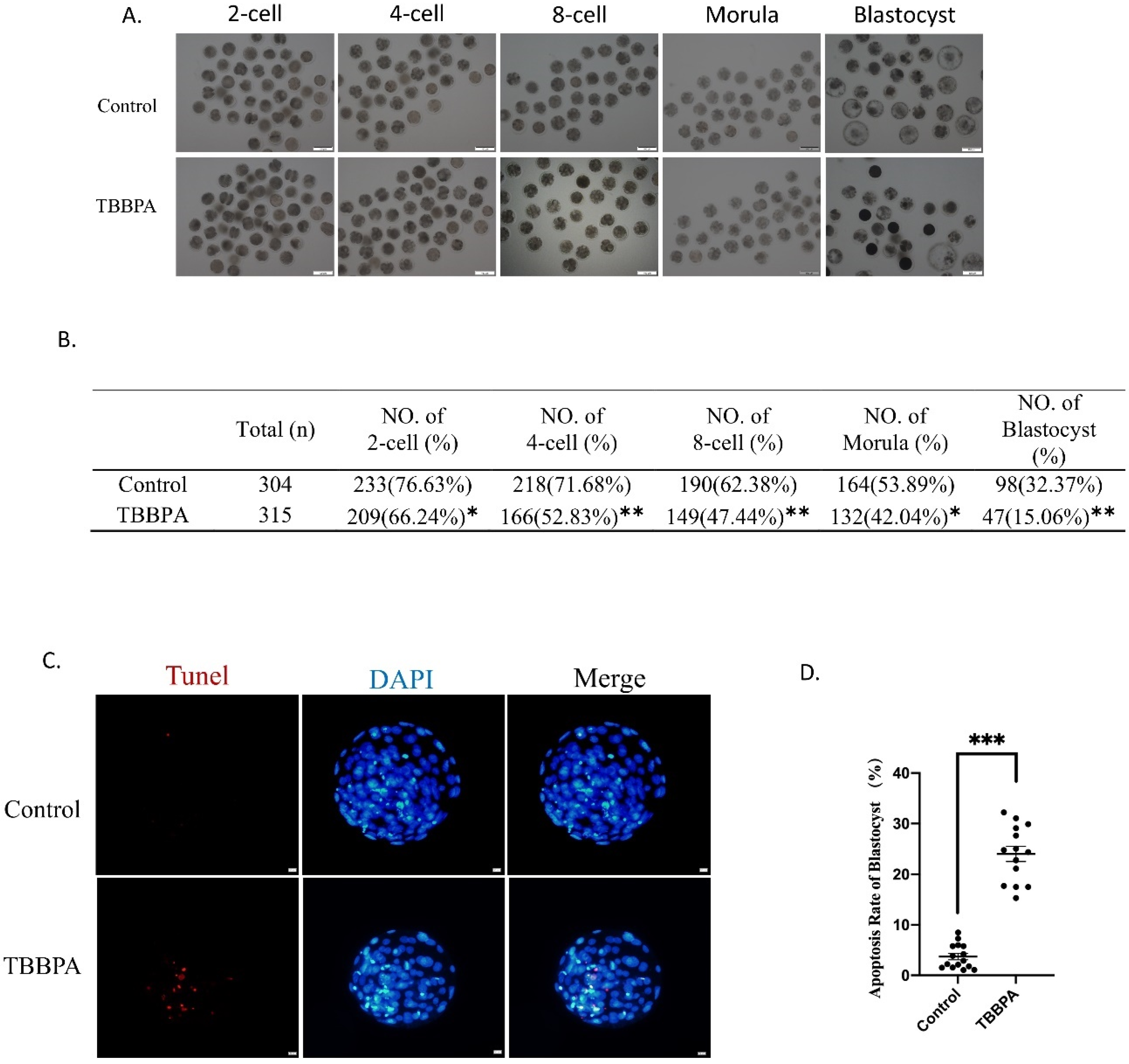
2.6. TBBPA Exposure Reduces Oocyte Competence
3. Discussion
4. Materials and Methods
4.1. Oocyte Collection and In Vitro Maturation (IVM)
4.2. TBBPA Treatment
4.3. Evaluation of Cumulus Expansion
4.4. Measurement of Mitochondrial Membrane Potential (ΔΨm), Reactive Oxygen Species (ROS), Glutathione (GSH), Dihydroergotamine (DHE), Mitochondria, ATP, and Apoptosis
4.5. Immunofluorescence
4.6. Measurement of mtDNA Copy Number
4.7. Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.8. IVF and Embryo Culture
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, K.; Li, J.; Yan, S.; Zhang, W.; Li, Y.; Han, D. A review of status of tetrabromobisphenol A (TBBPA) in China. Chemosphere 2016, 148, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.A.-E.; Pawar, G.; Harrad, S. Evaluation of in vitro vs. in vivo methods for assessment of dermal absorption of organic flame retardants: A review. Environ. Int. 2015, 74, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, L.; Zhao, Y.; Sun, Z.; Zhou, X.; Li, J.; Wu, Y. Dietary exposure assessment of Chinese population to tetrabromobisphenol-A, hexabromocyclododecane and decabrominated diphenyl ether: Results of the 5th Chinese Total Diet Study. Environ. Pollut. 2017, 229, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liu, S.; Liu, T.; Yang, C.; Wu, Y.; Tan, H.J.J.; Wei, B.; Ma, X.; Feng, B.; Jiang, Q.; et al. Association of prenatal exposure to bisphenols and birth size in Zhuang ethnic newborns. Chemosphere 2020, 252, 126422. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yu, Z.; Chen, H.; Han, Y.; Xiang, M.; Chen, X.; Ma, R.; Wang, Z. Tetrabromobisphenol A: Disposition, kinetics and toxicity in animals and humans. Environ. Pollut. 2019, 253, 909–917. [Google Scholar] [CrossRef]
- Knudsen, G.A.; Sanders, J.M.; Sadik, A.M.; Birnbaum, L.S. Disposition and kinetics of tetrabromobisphenol A in female Wistar Han rats. Toxicol. Rep. 2014, 1, 214–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Wang, J.; Xiang, Y.; Li, L.; Qie, H.; Ren, M.; Lin, A.; Qi, F. Effects of tetrabromobisphenol A (TBBPA) on the reproductive health of male rodents: A systematic review and meta-analysis. Sci. Total Environ. 2021, 781, 146745. [Google Scholar] [CrossRef]
- Liang, S.; Yin, L.; Yu, K.S.; Hofmann, M.-C.; Yu, X. High-Content Analysis Provides Mechanistic Insights into the Testicular Toxicity of Bisphenol A and Selected Analogues in Mouse Spermatogonial Cells. Toxicol. Sci. 2017, 155, 43–60. [Google Scholar] [CrossRef]
- Siracusa, J.S.; Yin, L.; Measel, E.; Liang, S.; Yu, X. Effects of bisphenol A and its analogs on reproductive health: A mini review. Reprod. Toxicol. 2018, 79, 96–123. [Google Scholar] [CrossRef]
- Jia, X.; Yan, R.; Lin, H.; Liu, Z.; Shen, L.; Yang, H.; Wu, H.; Shan, X.; Zhang, H. TBBPA and its alternative TCBPA induced ROS-dependent mitochondria-mediated apoptosis in the liver of Rana nigromaculata. Environ. Pollut. 2022, 297, 118791. [Google Scholar] [CrossRef]
- Lu, L.; Hu, J.; Li, G.; An, T. Low concentration Tetrabromobisphenol A (TBBPA) elevating overall metabolism by inducing activation of the Ras signaling pathway. J. Hazard. Mater. 2021, 416, 125797. [Google Scholar] [CrossRef] [PubMed]
- Babayev, E.; Seli, E. Oocyte mitochondrial function and reproduction. Curr. Opin. Obstet. Gynecol. 2015, 27, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J. Cell. Physiol. 2021, 236, 7966–7983. [Google Scholar] [CrossRef]
- Prasad, S.; Tiwari, M.; Pandey, A.N.; Shrivastav, T.G.; Chaube, S.K. Impact of stress on oocyte quality and reproductive outcome. J. Biomed. Sci. 2016, 23, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Fagbohun, C.F.; Downs, S.M. Maturation of the Mouse Oocyte-Cumulus Cell Complex: Stimulation by Lectins1. Biol. Reprod. 1990, 42, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Feiteiro, J.; Rocha, S.M.; Mariana, M.; Maia, C.J.; Cairrao, E. Vascular Response of Tetrabromobisphenol a in Rat Aorta: Calcium Channels Inhibition and Potassium Channels Activation. Toxics 2022, 10, 529. [Google Scholar] [CrossRef]
- Kitamura, S.; Jinno, N.; Ohta, S.; Kuroki, H.; Fujimoto, N. Thyroid hormonal activity of the flame retardants tetrabromobisphenol A and tetrachlorobisphenol A. Biochem. Biophys. Res. Commun. 2002, 293, 554–559. [Google Scholar] [CrossRef]
- Fukuda, N.; Ito, Y.; Yamaguchi, M.; Mitumori, K.; Koizumi, M.; Hasegawa, R.; Kamata, E.; Ema, M. Unexpected nephrotoxicity induced by tetrabromobisphenol A in newborn rats. Toxicol. Lett. 2004, 150, 145–155. [Google Scholar] [CrossRef]
- Suh, K.S.; Choi, E.M.; Rhee, S.Y.; Oh, S.; Kim, S.W.; Pak, Y.K.; Choe, W.; Ha, J.; Chon, S. Tetrabromobisphenol A induces cellular damages in pancreatic β-cells in vitro. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2017, 52, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dai, G.-D. Comparative Effects of Brominated Flame Retardants BDE-209, TBBPA, and HBCD on Neurotoxicity in Mice. Chem. Res. Toxicol. 2022, 35, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-H.; Lee, S.; Jeon, H.; Kim, A.H.; Lee, W.; Lee, Y.; Yang, S.; Yun, J.; Jung, Y.-S.; Lee, J. Tetrabromobisphenol A-Induced Apoptosis in Neural Stem Cells Through Oxidative Stress and Mitochondrial Dysfunction. Neurotox. Res. 2020, 38, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Miao, Y.; Gao, Q.; Cui, Z.; Xiong, B. Exposure to perfluorooctane sulfonate in vitro perturbs the quality of porcine oocytes via induction of apoptosis. Environ. Pollut. 2021, 284, 117508. [Google Scholar] [CrossRef]
- Guo, Y.; Cao, Z.; Jiao, X.; Bai, D.; Zhang, Y.; Hua, J.; Liu, W.; Teng, X. Pre-pregnancy exposure to fine particulate matter (PM2.5) increases reactive oxygen species production in oocytes and decrease litter size and weight in mice. Environ. Pollut. 2020, 268, 115858. [Google Scholar] [CrossRef]
- Chen, J.; Cui, Z.; Qiu, Y.; Zhang, X.; Chen, F.; Wang, H.; Xiong, B.; Miao, Y.; Gao, Q. Exposure to Copper Compromises the Maturational Competency of Porcine Oocytes by Impairing Mitochondrial Function. Front. Cell Dev. Biol. 2021, 9, 678665. [Google Scholar] [CrossRef]
- Richani, D.; Dunning, K.R.; Thompson, J.G.; Gilchrist, R.B. Metabolic co-dependence of the oocyte and cumulus cells: Essential role in determining oocyte developmental competence. Hum. Reprod. Updat. 2021, 27, 27–47. [Google Scholar] [CrossRef]
- Qian, Y.; Shi, W.Q.; Ding, J.T.; Sha, J.H.; Fan, B.Q. Predictive Value of the Area of Expanded Cumulus Mass on Development of Porcine Oocytes Matured and Fertilized In Vitro. J. Reprod. Dev. 2003, 49, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Bosco, L.; Chiarelli, R.; Roccheri, M.C.; Matranga, D.; Ruvolo, G. Relationship between apoptosis and survival molecules in human cumulus cells as markers of oocyte competence. Zygote 2017, 25, 583–591. [Google Scholar] [CrossRef]
- Severson, A.F.; von Dassow, G.; Bowerman, B. Oocyte Meiotic Spindle Assembly and Function. Curr. Top Dev. Biol. 2016, 116, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Bennabi, I.; Terret, M.-E.; Verlhac, M.-H. Meiotic spindle assembly and chromosome segregation in oocytes. J. Cell Biol. 2016, 215, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.T.; Lane, S.; Liu, W.; Shan, T.; Yang, X.; Liang, S.; Zhang, P.; Liu, Y.; Liu, X.; Kuang, S. Molecular causes of aneuploidy in mammalian eggs. Development 2013, 140, 3719–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, F.D.D.A.; Adona, P.R.; Guemra, S.; Damião, B.C.M. Oxidative homeostasis in oocyte competence for in vitro embryo development. Anim. Sci. J. 2019, 90, 1343–1349. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C. The importance of having high glutathione (GSH) level after bovine in vitro maturation on embryo development: Effect of β-mercaptoethanol, cysteine and cystine. Theriogenology 2000, 53, 761–771. [Google Scholar] [CrossRef]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2022, 36, e22942. [Google Scholar] [CrossRef]
- Otera, H.; Mihara, K. Mitochondrial Dynamics: Functional Link with Apoptosis. Int. J. Cell Biol. 2012, 2012, 821676. [Google Scholar] [CrossRef] [Green Version]
- Roger, A.J.; Muñoz-Gómez, S.A.; Kamikawa, R. The Origin and Diversification of Mitochondria. Curr. Biol. 2017, 27, R1177–R1192. [Google Scholar] [CrossRef] [Green Version]
- Hatefi, Y. ATP synthesis in mitochondria. Eur. J. Biochem. 1993, 218, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yu, Q. Role of mTOR Signaling in Female Reproduction. Front. Endocrinol. 2019, 10, 692. [Google Scholar] [CrossRef]
- Malott, K.F.; Luderer, U. Toxicant effects on mammalian oocyte mitochondria. Biol. Reprod. 2021, 104, 784–793. [Google Scholar] [CrossRef]
- Fagbohun, C.F.; Downs, S.M. Requirement for glucose in ligand-stimulated meiotic maturation of cumulus cell-enclosed mouse oocytes. Reproduction 1992, 96, 681–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korotchkina, L.G.; Patel, M.S. Probing the Mechanism of Inactivation of Human Pyruvate Dehydrogenase by Phosphorylation of Three Sites. J. Biol. Chem. 2001, 276, 5731–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Zhang, L.; Han, L.; Ge, J.; Ma, R.; Zhang, X.; Moley, K.; Schedl, T.; Wang, Q. Differing roles of pyruvate dehydrogenase kinases during mouse oocyte maturation. J. Cell Sci. 2015, 128, 2319–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, C.C.; Bhandari, S.; Moorey, S.E. Concurrent Measurement of Mitochondrial DNA Copy Number and ATP Concentration in Single Bovine Oocytes. Methods Protoc. 2021, 4, 88. [Google Scholar] [CrossRef] [PubMed]
- Pivonello, C.; Muscogiuri, G.; Nardone, A.; Garifalos, F.; Provvisiero, D.P.; Verde, N.; De Angelis, C.; Conforti, A.; Piscopo, M.; Auriemma, R.S.; et al. Bisphenol A: An emerging threat to female fertility. Reprod. Biol. Endocrinol. 2020, 18, 22. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Primer Sequence (5’–3’) |
|---|---|---|
| 18S | Forward | GACTCATTGGCCCTGTAATTGGAATGAGTC |
| Reverse | GCTGCTGGCACCAGACTTG | |
| Tnfaip6 | Forward | TATGGGAAGAGGCTCACGGATGG |
| Reverse | GGTAGACGCCTGCTGCTTGTTC | |
| Ptx3 | Forward | TGGTCGCTGATGCTGTGATTTCC |
| Reverse | GCCACCGAGTCACCATTTACCC | |
| Bcl-2 | Forward | TCGTGGCCTTCTTTGAGTTCG |
| Reverse | GCCTGTGGGCTTCACTTATGG | |
| Bax | Forward | CGGAGATGAATTGGACAGTAAC |
| Reverse | AGCACTCCAGCCACAAAGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Min, C.-G.; Zhang, K.-Y.; Zhan, C.-L.; Wang, Y.-C.; Hou, S.-K.; Ma, X.; Lu, W.-F. Tetrabromobisphenol Exposure Impairs Bovine Oocyte Maturation by Inducing Mitochondrial Dysfunction. Molecules 2022, 27, 8111. https://doi.org/10.3390/molecules27228111
Guo J, Min C-G, Zhang K-Y, Zhan C-L, Wang Y-C, Hou S-K, Ma X, Lu W-F. Tetrabromobisphenol Exposure Impairs Bovine Oocyte Maturation by Inducing Mitochondrial Dysfunction. Molecules. 2022; 27(22):8111. https://doi.org/10.3390/molecules27228111
Chicago/Turabian StyleGuo, Jing, Chang-Guo Min, Kai-Yan Zhang, Cheng-Lin Zhan, Yu-Chan Wang, Sheng-Kui Hou, Xin Ma, and Wen-Fa Lu. 2022. "Tetrabromobisphenol Exposure Impairs Bovine Oocyte Maturation by Inducing Mitochondrial Dysfunction" Molecules 27, no. 22: 8111. https://doi.org/10.3390/molecules27228111