
Lactobacillus rhamnosus TR08 Improves Dyslipidemia in Mice Fed with a High Fat Diet by Regulating the Intestinal Microbiota, Reducing Systemic Inflammatory Response, and Promoting Sphingomholipid Metabolism
 and
and
Abstract
:1. Introduction
2. Results
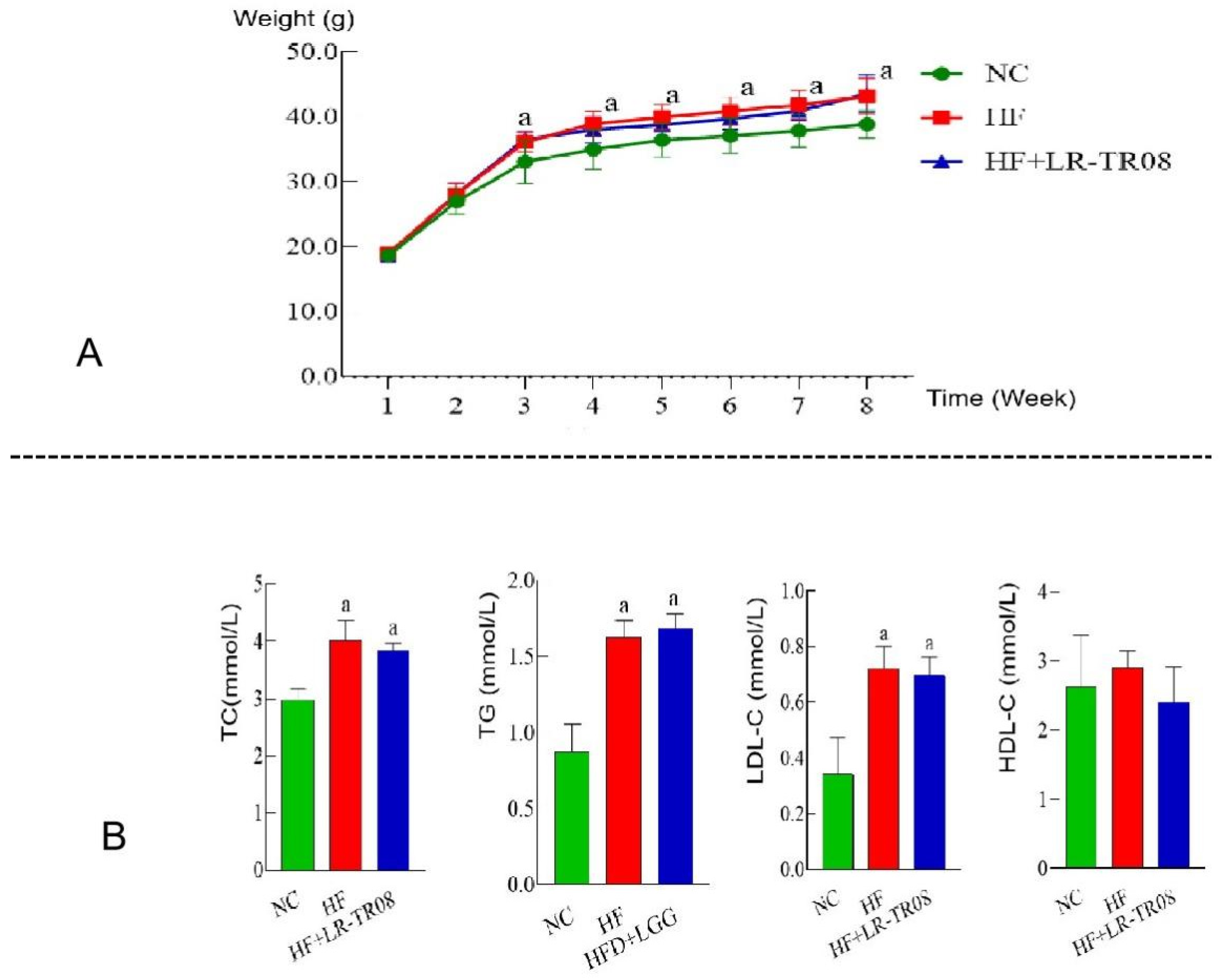
2.1. L. rhamnosus TR08 Decreased Body Weight and Blood Lipids in Hyperlipidemia Mice
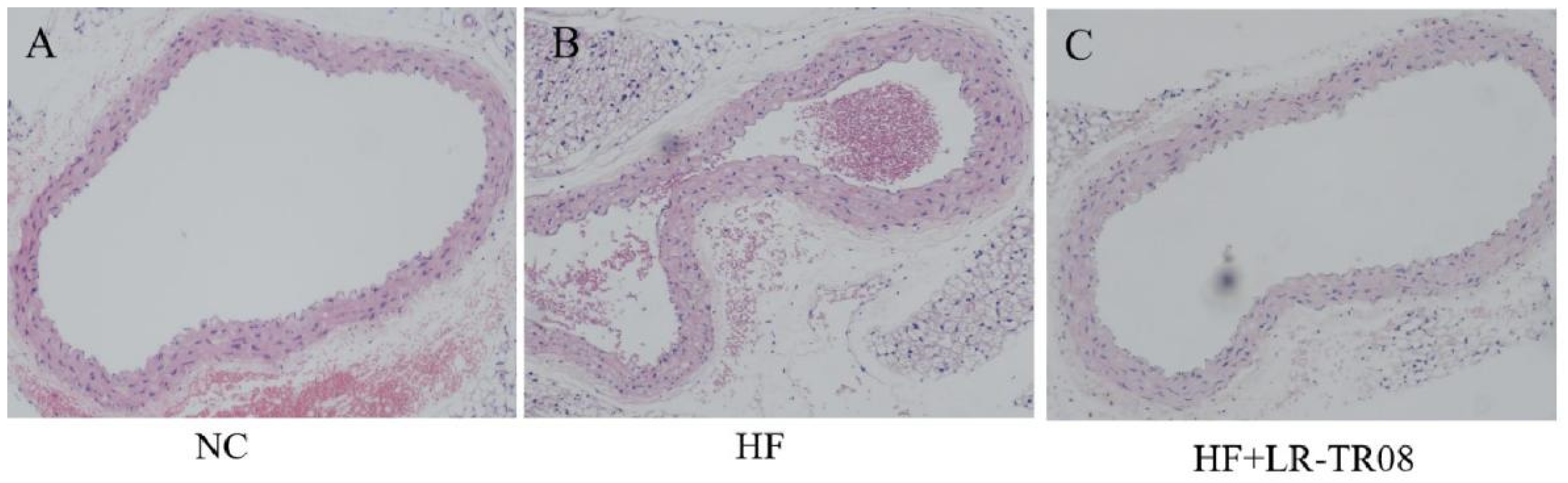
2.2. L. rhamnosus TR08 Attenuated the Inflammation in Blood Vessels Wall of Hyperlipidemia Mice
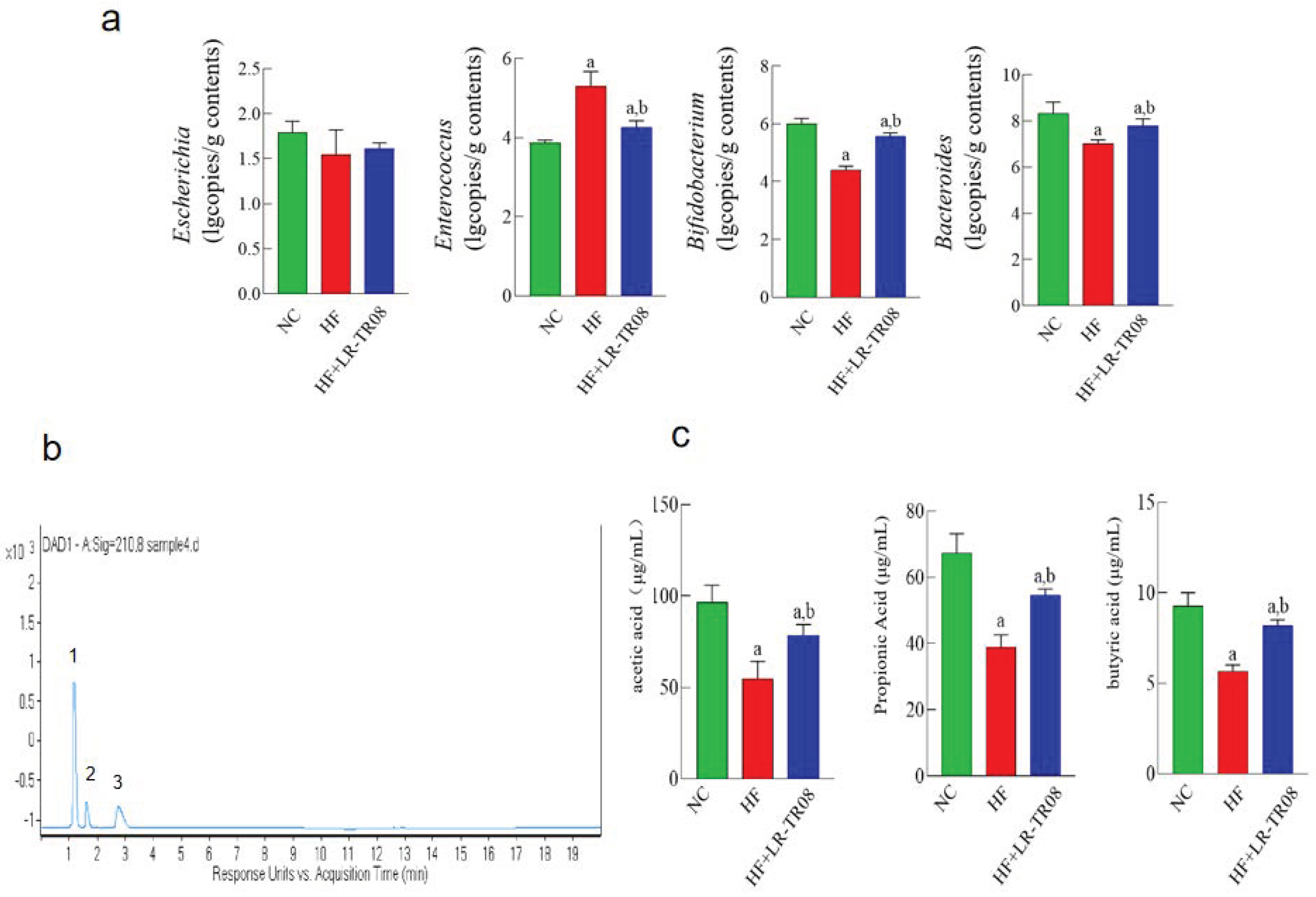
2.3. L. rhamnosus TR08 Influenced the Intestinal Microbiota and SCFAs Production in Hyperlipidemia Mice
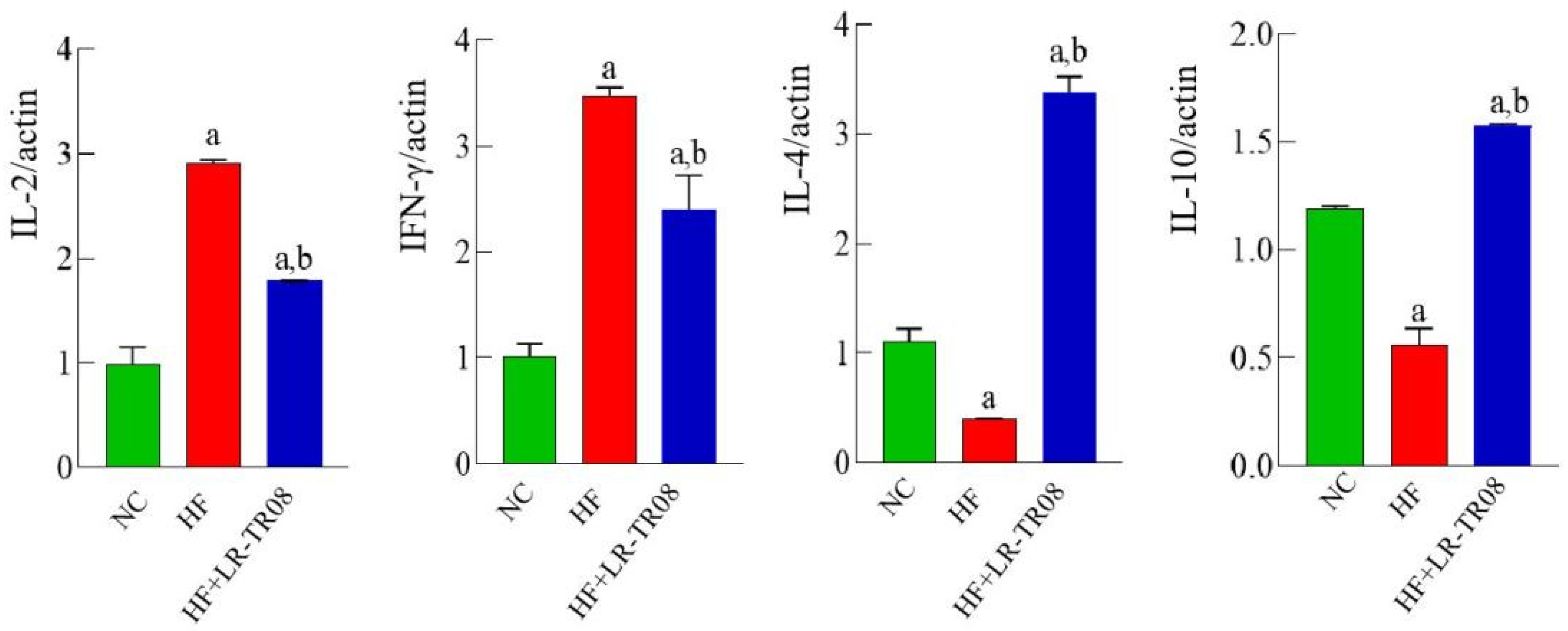
2.4. L. rhamnosus TR08 Reduced the Systemic Inflammatory Response in Hyperlipidemia Mice
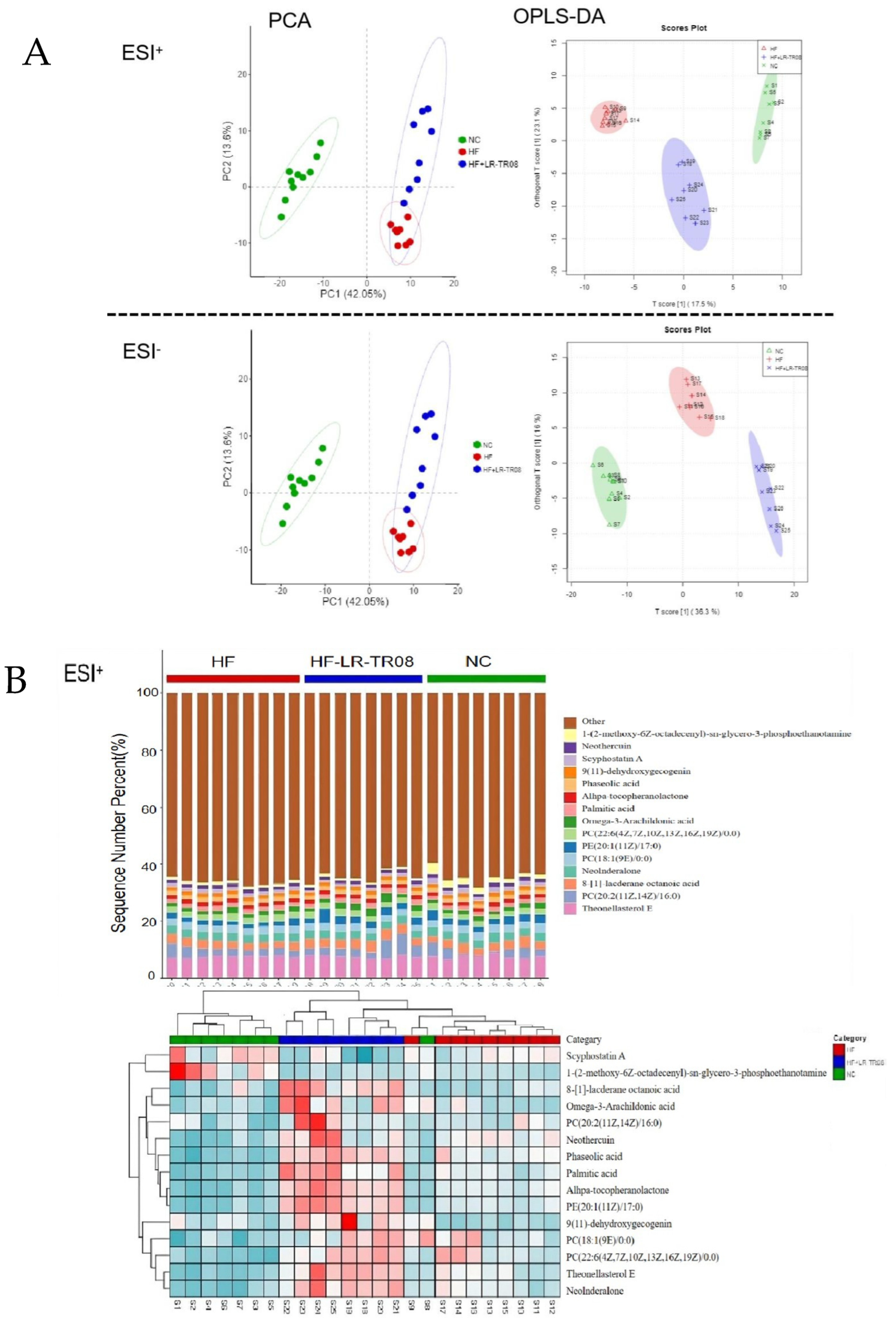
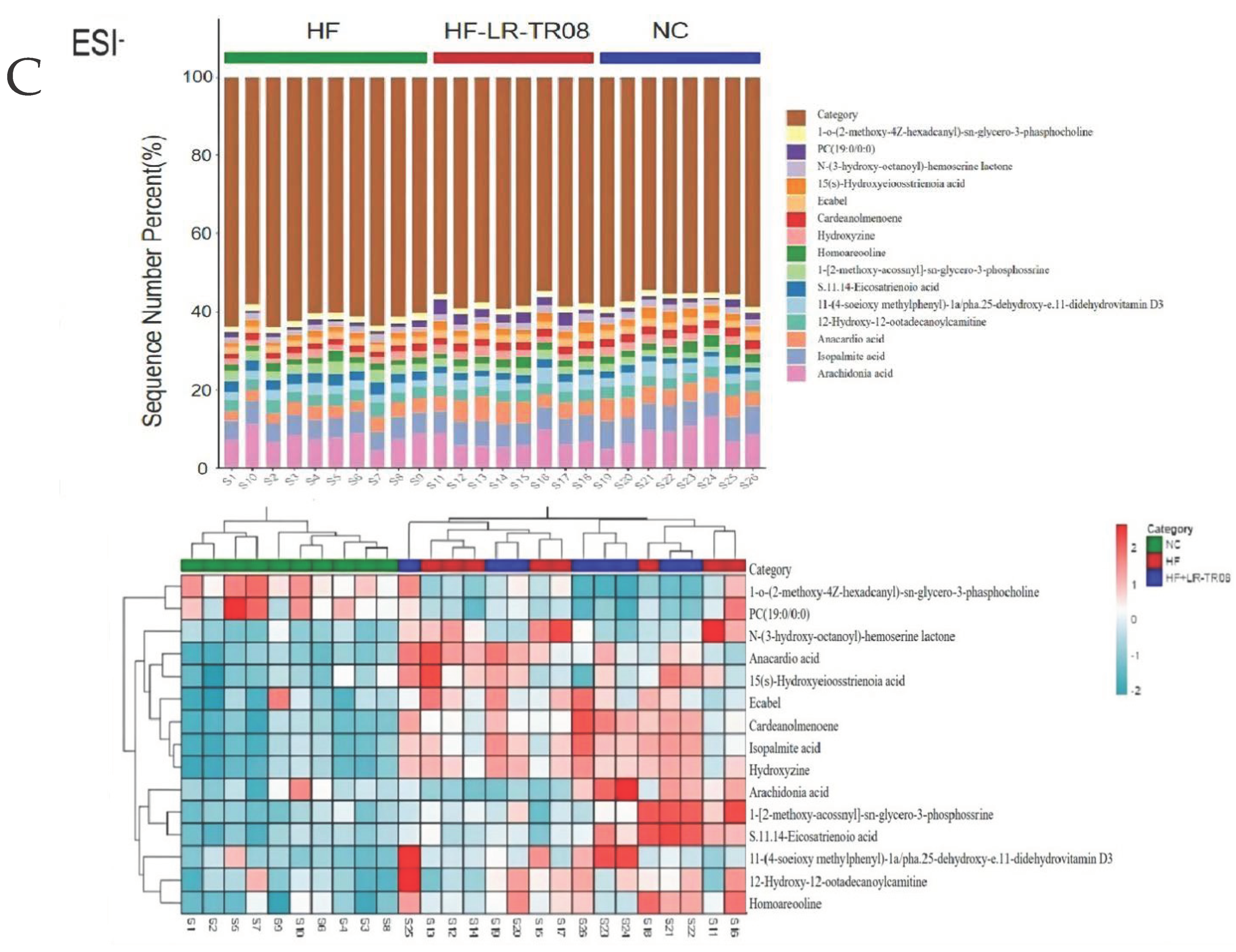
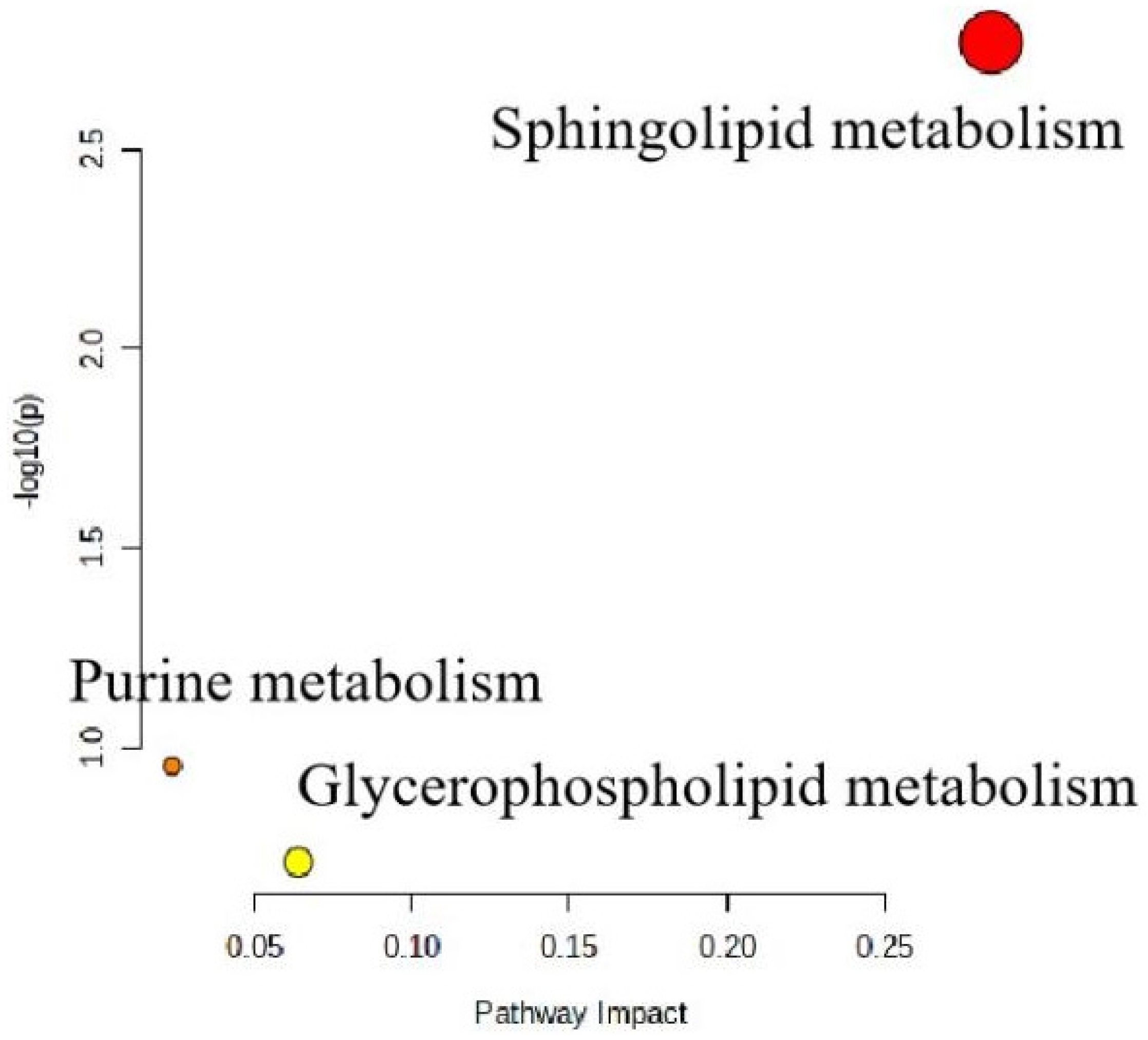
2.5. L. rhamnosus TR08 Regulated Metabolomics in Hyperlipidemia Mice
3. Discussion
4. Materials and Methods
4.1. High Fat Diet Induced Murine Hyperlipidemia
4.2. Histological Analysis
4.3. DNA Extraction and Real Time Polymerase Chain Reaction (qPCR)
4.4. RNA Extraction and qRT–PCR
4.5. Preparation of Serum Sample
4.6. Blood Parameters Analysis
4.7. Liquid Chromatography–Mass Spectrometry (LC–MS/MS)
4.8. Untargeted Metabolomics Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ma, L.Y.; Chen, W.W.; Gao, R.L.; Liu, L.S.; Zhu, M.L.; Wang, Y.J.; Wu, Z.S.; Li, H.J.; Gu, D.F.; Yang, Y.; et al. China cardiovascular diseases report 2018: An updated summary. J. Geriatr. Cardiol. 2020, 17, 1. [Google Scholar]
- Stewart, J.; McCallin, T.; Martinez, J.; Chacko, S.; Yusuf, S. Hyperlipidemia. Pediatr. Rev. 2020, 41, 393–402. [Google Scholar]
- Pathak, R.K.; Mahajan, R.; Lau, D.H.; Sanders, P. The implications of obesity for cardiac arrhythmia mechanisms and management. Can. J. Cardiol. 2015, 31, 203–210. [Google Scholar]
- Fenk, S.; Fischer, M.; Strack, C.; Schmitz, G.; Loew, T.; Lahmann, C.; Baessler, A. Successful weight reduction improves left ventricular diastolic function and physical performance in severe obesity. Int. Heart J. 2015, 56, 196–202. [Google Scholar]
- Balta, S. Endothelial dysfunction and inflammatory markers of vascular disease. Curr. Vasc. Pharmacol. 2021, 19, 243–249. [Google Scholar]
- Suárez-Rivero, J.M.; Pastor-Maldonado, C.J.; de la Mata, M.; Villanueva-Paz, M.; Povea-Cabello, S.; Álvarez-Córdoba, M.; Villalón-García, I.; Suárez-Carrillo, A.; Talaverón-Rey, M.; Munuera, M.; et al. Atherosclerosis and coenzyme Q10. Int. J. Mol. Sci. 2019, 20, 5195. [Google Scholar]
- Garcia, C.; Blesso, C.N. Antioxidant properties of anthocyanins and their mechanism of action in atherosclerosis. Free Radic. Biol. Med. 2021, 172, 152–166. [Google Scholar]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar]
- Yao, Y.S.; Li, T.D.; Zeng, Z.H. Mechanisms underlying direct actions of hyperlipidemia on myocardium: An updated review. Lipids Health Dis. 2020, 19, 23. [Google Scholar]
- Andreadou, I.; Daiber, A.; Baxter, G.F.; Brizzi, M.F.; Di Lisa, F.; Kaludercic, N.; Lazou, A.; Varga, Z.V.; Zuurbier, C.J.; Schulz, R.; et al. Influence of cardiometabolic comorbidities on myocardial function, infarction, and cardioprotection: Role of cardiac redox signaling. Free Radic. Biol. Med. 2021, 166, 33–52. [Google Scholar]
- Brial, F.; Le Lay, A.; Dumas, M.E.; Gauguier, D. Implication of gut microbiota metabolites in cardiovascular and metabolic diseases. Cell. Mol. Life Sci. 2018, 75, 3977–3990. [Google Scholar]
- Brahe, L.K.; Astrup, A.; Larsen, L.H. Can we prevent obesity-related metabolic diseases by dietary modulation of the gut microbiota? Adv. Nutr. 2016, 7, 90–101. [Google Scholar]
- Jia, X.; Xu, W.; Zhang, L.; Li, X.; Wang, R.; Wu, S. Impact of gut microbiota and microbiota-related metabolites on hyperlipidemia. Front. Cell. Infect. Microbiol. 2021, 11, 634780. [Google Scholar]
- He, K.; Hu, Y.; Ma, H.; Zou, Z.; Xiao, Y.; Yang, Y.; Feng, M.; Li, X.; Ye, X. Rhizoma Coptidis alkaloids alleviate hyperlipidemia in B6 mice by modulating gut microbiota and bile acid pathways. Biochim. Biophys. Acta 2016, 1862, 1696–1709. [Google Scholar]
- Yu, L.; Zhou, X.; Duan, H.; Chen, Y.; Cui, S.; Guo, R.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; et al. Synergistic protective effects of different dietary supplements against type 2 diabetes via regulating gut microbiota. J. Med. Food 2021, 24, 319–330. [Google Scholar]
- Guerrero-Bonmatty, R.; Gil-Fernández, G.; Rodríguez-Velasco, F.J.; Espadaler-Mazo, J. A combination of Lactoplantibacillus plantarum strains CECT7527, CECT7528, and CECT7529 plus monacolin K reduces blood cholesterol: Results from a randomized, double-blind, placebo-controlled study. Nutrients 2021, 13, 1206. [Google Scholar]
- Rinott, E.; Youngster, I.; Meir, A.Y.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Knights, D.; Tuohy, K.; Fava, F.; Scholz, M.U.; et al. Effects of diet-modulated autologous fecal microbiota transplantation on weight regain. Gastroenterology 2021, 160, 158–173. [Google Scholar]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-induced gut microbiota reduces obesity in high-fat diet-fed mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar]
- Silveira Rossi, J.L.; Barbalho, S.M.; Reverete de Araujo, R.; Bechara, M.D.; Sloan, K.P.; Sloan, L.A. Metabolic syndrome and cardiovascular diseases: Going beyond traditional risk factors. Diabetes Metab. Res. Rev. 2022, 38, e3502. [Google Scholar]
- Deng, X.; Ma, J.; Song, M.; Jin, Y.; Ji, C.; Ge, W.; Guo, C. Effects of products designed to modulate the gut microbiota on hyperlipidaemia. Eur. J. Nutr. 2019, 58, 2713–2729. [Google Scholar]
- Miao, J.; Zang, X.; Cui, X.; Zhang, J. Autophagy, hyperlipidemia, and atherosclerosis. Adv. Exp. Med. Biol. 2020, 1207, 237–264. [Google Scholar]
- Handhle, A.; Park, A. Hyperlipidaemia. Medicine 2021, 49, 587–591. [Google Scholar]
- Juste, C.; Gérard, P. Cholesterol-to-coprostanol conversion by the gut microbiota: What we know, suspect, and ignore. Microorganisms 2021, 9, 1881. [Google Scholar]
- Villette, R.; Kc, P.; Beliard, S.; Salas Tapia, M.F.; Rainteau, D.; Guerin, M.; Lesnik, P. Unraveling host-gut microbiota dialogue and its impact on cholesterol levels. Front. Pharmacol. 2020, 11, 278. [Google Scholar]
- Kriaa, A.; Bourgin, M.; Potiron, A.; Mkaouar, H.; Jablaoui, A.; Gérard, P.; Maguin, E.; Rhimi, M. Microbial impact on cholesterol and bile acid metabolism: Current status and future prospects. J. Lipid Res. 2019, 60, 323–332. [Google Scholar]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar]
- Eslami, M.; Bahar, A.; Hemati, M.; Rasouli Nejad, Z.; Mehranfar, F.; Karami, S.; Kobyliak, N.M.; Yousefi, B. Dietary pattern, colonic microbiota and immunometabolism interaction: New frontiers for diabetes mellitus and related disorders. Diabet. Med. 2021, 38, e14415. [Google Scholar]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Zhao, S.; Su, Z.; Ge, C.; Zhang, Y.; Jia, J.; et al. The intestinal microbiome associated with lipid metabolism and obesity in humans and animals. J. Appl. Microbiol. 2022, 133, 2915–2930. [Google Scholar] [CrossRef]
- Kumar, J.; Rani, K.; Datt, C. Molecular link between dietary fibre, gut microbiota and health. Mol. Biol. Rep. 2020, 47, 6229–6237. [Google Scholar]
- Pizzorno, J. Toxins from the gut. Integr. Med. 2014, 13, 8. [Google Scholar]
- Ding, L.; Chang, M.; Guo, Y.; Zhang, L.; Xue, C.; Yanagita, T.; Zhang, T.; Wang, Y. Trimethylamine-N-oxide (TMAO)-induced atherosclerosis is associated with bile acid metabolism. Lipids Health Dis. 2018, 17, 286. [Google Scholar]
- Zhang, Y.; Liu, W.; Wei, Z.; Yin, B.; Man, C.; Jiang, Y. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT 2021, 139, 110590. [Google Scholar]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and improving function of Lactobacilli in the prevention and treatment of cardiovascular-related diseases: A novel perspective from gut microbiota. Front. Nutr. 2021, 8, 299. [Google Scholar]
- Da Silva Pontes, K.S.; Guedes, M.R.; da Cunha, M.R.; de Souza Mattos, S.; Silva, M.I.; Neves, M.F.; Marques, B.C.; Klein, M.R. Effects of probiotics on body adiposity and cardiovascular risk markers in individuals with overweight and obesity: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2021, 40, 4915–4931. [Google Scholar]
- Tunapong, W.; Apaijai, N.; Yasom, S.; Tanajak, P.; Wanchai, K.; Chunchai, T.; Kerdphoo, S.; Eaimworawuthikul, S.; Thiennimitr, P.; Pongchaidecha, A.; et al. Chronic treatment with prebiotics, probiotics and synbiotics attenuated cardiac dysfunction by improving cardiac mitochondrial dysfunction in male obese insulin-resistant rats. Eur. J. Nutr. 2018, 57, 2091–2104. [Google Scholar]
- Jang, H.R.; Park, H.J.; Kang, D.; Chung, H.; Nam, M.H.; Lee, Y.; Park, J.H.; Lee, H.Y. A protective mechanism of probiotic Lactobacillus against hepatic steatosis via reducing host intestinal fatty acid absorption. Exp. Mol. Med. 2019, 51, 1–4. [Google Scholar]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar]
- Den Besten, G.; Lange, K.; Havinga, R.; van Dijk, T.H.; Gerding, A.; van Eunen, K.; Müller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G900–G910. [Google Scholar]
- Ziętek, M.; Celewicz, Z.; Kikut, J.; Szczuko, M. Implications of SCFAs on the parameters of the lipid and hepatic profile in pregnant women. Nutrients 2021, 13, 1749. [Google Scholar]
- Wu, G.; Jiang, Z.; Pu, Y.; Chen, S.; Wang, T.; Wang, Y.; Xu, X.; Wang, S.; Jin, M.; Yao, Y.; et al. Serum short-chain fatty acids and its correlation with motor and non-motor symptoms in Parkinson’s disease patients. BMC Neurol. 2022, 22, 13. [Google Scholar]
- Duan, C.; Tang, X.; Wang, W.; Qian, W.; Fu, X.; Deng, X.; Zhou, S.; Han, C.; Hou, X. Lactobacillus rhamnosus attenuates intestinal inflammation induced by Fusobacterium nucleatum infection by restoring the autophagic flux. Int. J. Mol. Med. 2021, 47, 125–136. [Google Scholar]
- Plaza-Diaz, J.; Gomez-Llorente, C.; Fontana, L.; Gil, A. Modulation of immunity and inflammatory gene expression in the gut, in inflammatory diseases of the gut and in the liver by probiotics. World J. Gastroenterol. 2014, 20, 15632–15649. [Google Scholar]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Moreno Guerrero, S.S.; Ramírez Pacheco, A.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus strains stimulate the inflammatory response and activate human macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar]
- Fu, X.; Zong, T.; Yang, P.; Li, L.; Wang, S.; Wang, Z.; Li, M.; Li, X.; Zou, Y.; Zhang, Y.; et al. Nicotine: Regulatory roles and mechanisms in atherosclerosis progression. Food Chem. Toxicol. 2021, 151, 112154. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolite | Library ID | Formula | M/Z | RT/min | Trend |
|---|---|---|---|---|---|---|
| 1 | 7,10,13,16–Docosatetraynoic acid | LMFA 01030682 | C22H28O2 | 342.24 | 27.11 | ↓ |
| 2 | PC(P-15:0/0:0) | LMGP0 1070003 | C23H48NO6P | 466.33 | 26.64 | ↑ |
| 3 | 4b–Hydroxycholesterol | HMDB 13643 | C27H46O2 | 447.35 | 41.88 | ↑ |
| 4 | 10–Octadecenoic acid | LMFA 01031090 | C18H34O2 | 327.25 | 36.00 | ↑ |
| 5 | 5–Dodecenoic acid | HMDB 00529 | C12H22O2 | 593.48 | 43.07 | ↑ |
| 6 | 1–(2–methoxy–hexadecanyl)–sn–glycero–3–phosphoserine | LMGP 03060014 | C23H48NO9P | 534.28 | 26.37 | ↑ |
| 7 | Leukotriene C5 | HMDB 12993 | C30H45N3O9S | 624.29 | 29.36 | ↓ |
| 8 | PA(22:2(13Z,16Z)/16:0) | LMGP 10010761 | C41H77O8P | 751.52 | 30.24 | ↑ |
| 9 | PA(P–20:0/17:2(9Z,12Z)) | LMGP 10030066 | C40H75O7P | 721.51 | 32.94 | ↑ |
| 10 | PC(P–18:1(11Z)/16:0) | LMGP 01030134 | C42H82NO7P | 744.59 | 41.95 | ↑ |
| 11 | Cer(d18:0/13:0) | LMSP 02010018 | C31H63NO3 | 520.47 | 26.81 | ↓ |
| Ingredients | Content |
|---|---|
| Cane sugar | 20.0% |
| Lard oil | 15.0% |
| Casein | 10.0% |
| Cholesterol | 1.2% |
| Calcium bicarbonate | 0.6% |
| Land plaster | 0.4% |
| Gunk | 0.4% |
| Sodium cholate | 0.2% |
| Basal feed | 52.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Ding, L.; Ma, G.; Zhang, Y.; Sun, Y.; Li, Z.; Tao, X.; Ali, A.; Wang, D.; Wu, L. Lactobacillus rhamnosus TR08 Improves Dyslipidemia in Mice Fed with a High Fat Diet by Regulating the Intestinal Microbiota, Reducing Systemic Inflammatory Response, and Promoting Sphingomholipid Metabolism. Molecules 2022, 27, 7357. https://doi.org/10.3390/molecules27217357
Feng X, Ding L, Ma G, Zhang Y, Sun Y, Li Z, Tao X, Ali A, Wang D, Wu L. Lactobacillus rhamnosus TR08 Improves Dyslipidemia in Mice Fed with a High Fat Diet by Regulating the Intestinal Microbiota, Reducing Systemic Inflammatory Response, and Promoting Sphingomholipid Metabolism. Molecules. 2022; 27(21):7357. https://doi.org/10.3390/molecules27217357
Chicago/Turabian StyleFeng, Xiaohong, Longkun Ding, Guifang Ma, Ying Zhang, Yefu Sun, Zhengzhang Li, Xiaojun Tao, Asmaa Ali, Dongxu Wang, and Liang Wu. 2022. "Lactobacillus rhamnosus TR08 Improves Dyslipidemia in Mice Fed with a High Fat Diet by Regulating the Intestinal Microbiota, Reducing Systemic Inflammatory Response, and Promoting Sphingomholipid Metabolism" Molecules 27, no. 21: 7357. https://doi.org/10.3390/molecules27217357