
Andrographolide Induces G2/M Cell Cycle Arrest and Apoptosis in Human Glioblastoma DBTRG-05MG Cell Line via ERK1/2 /c-Myc/p53 Signaling Pathway

Abstract
:1. Introduction
2. Results
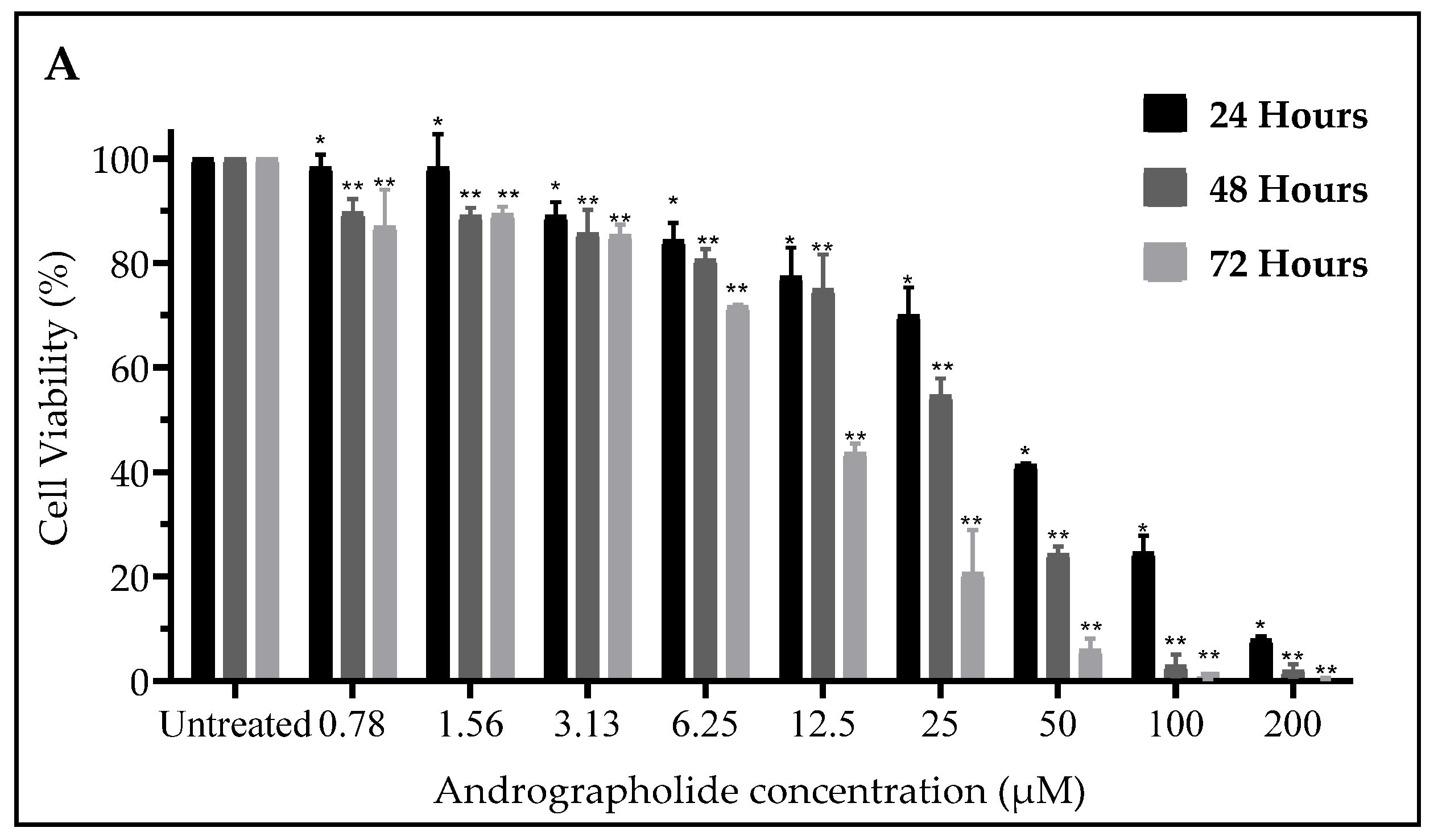
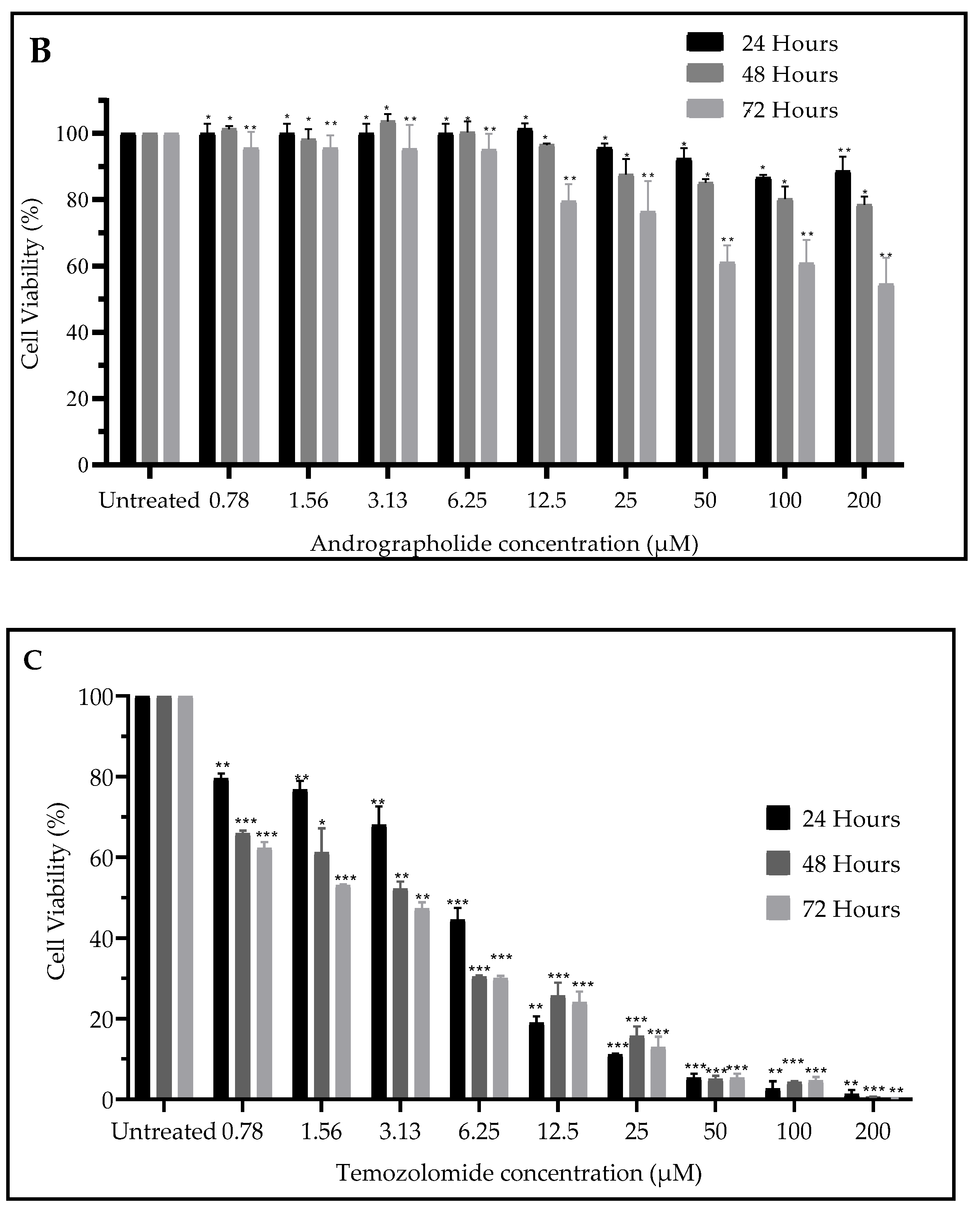
2.1. Andrographolide Suppressed Cell Viability of DBTRG-05MG and SVGp12 Cell Line
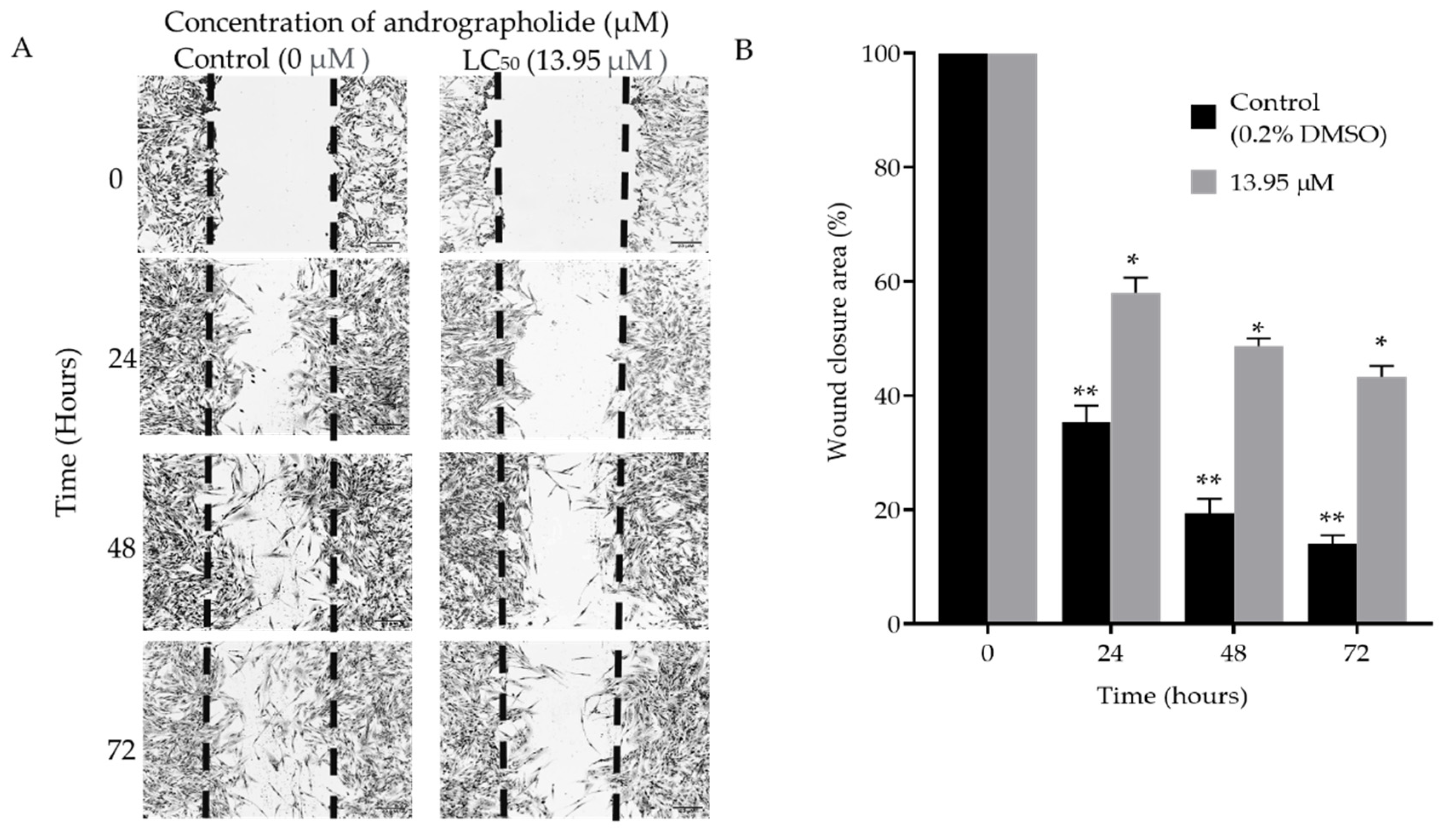
2.2. Scratch
Scratch Assay
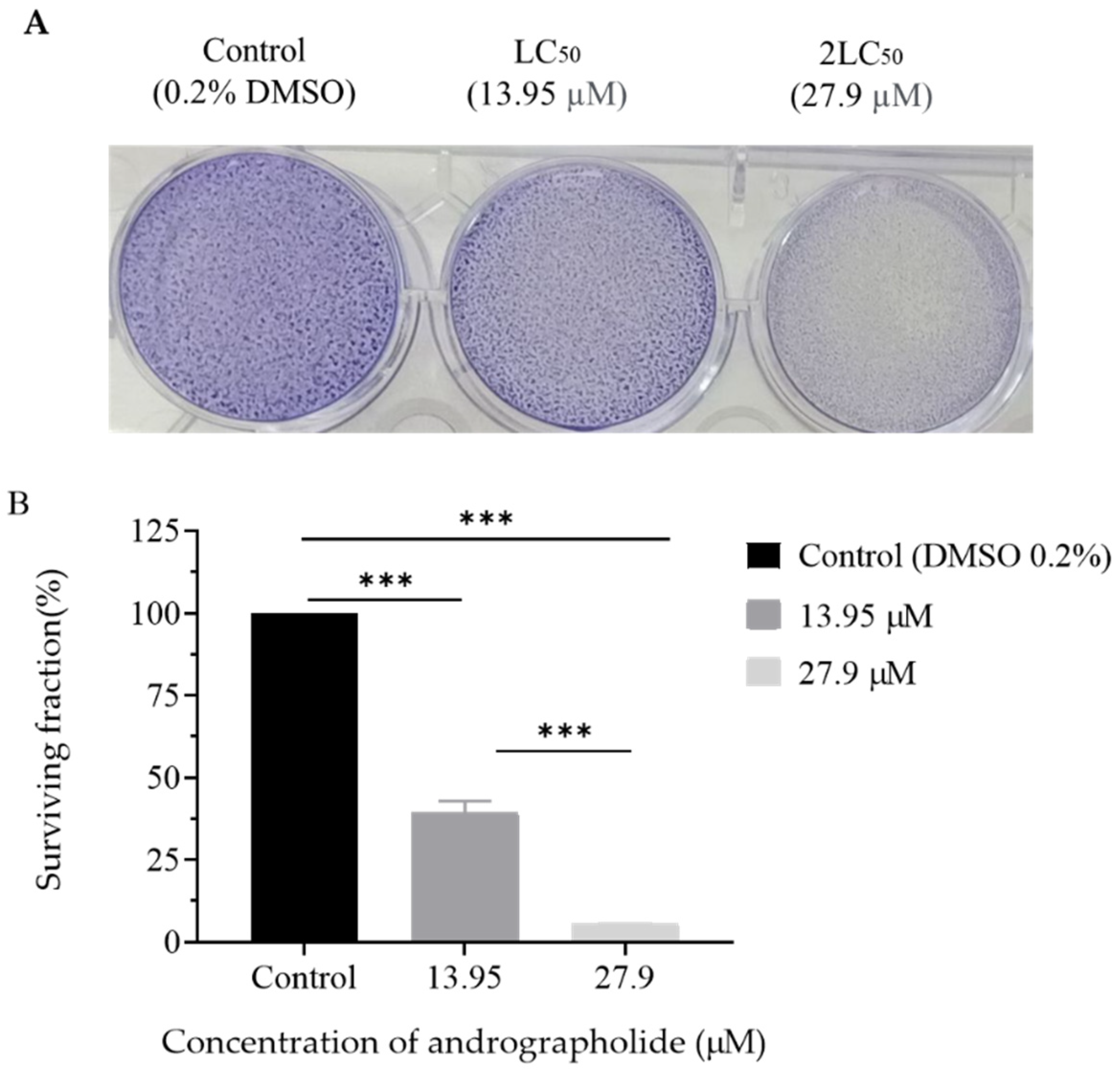
2.3. Clonogenic Assay
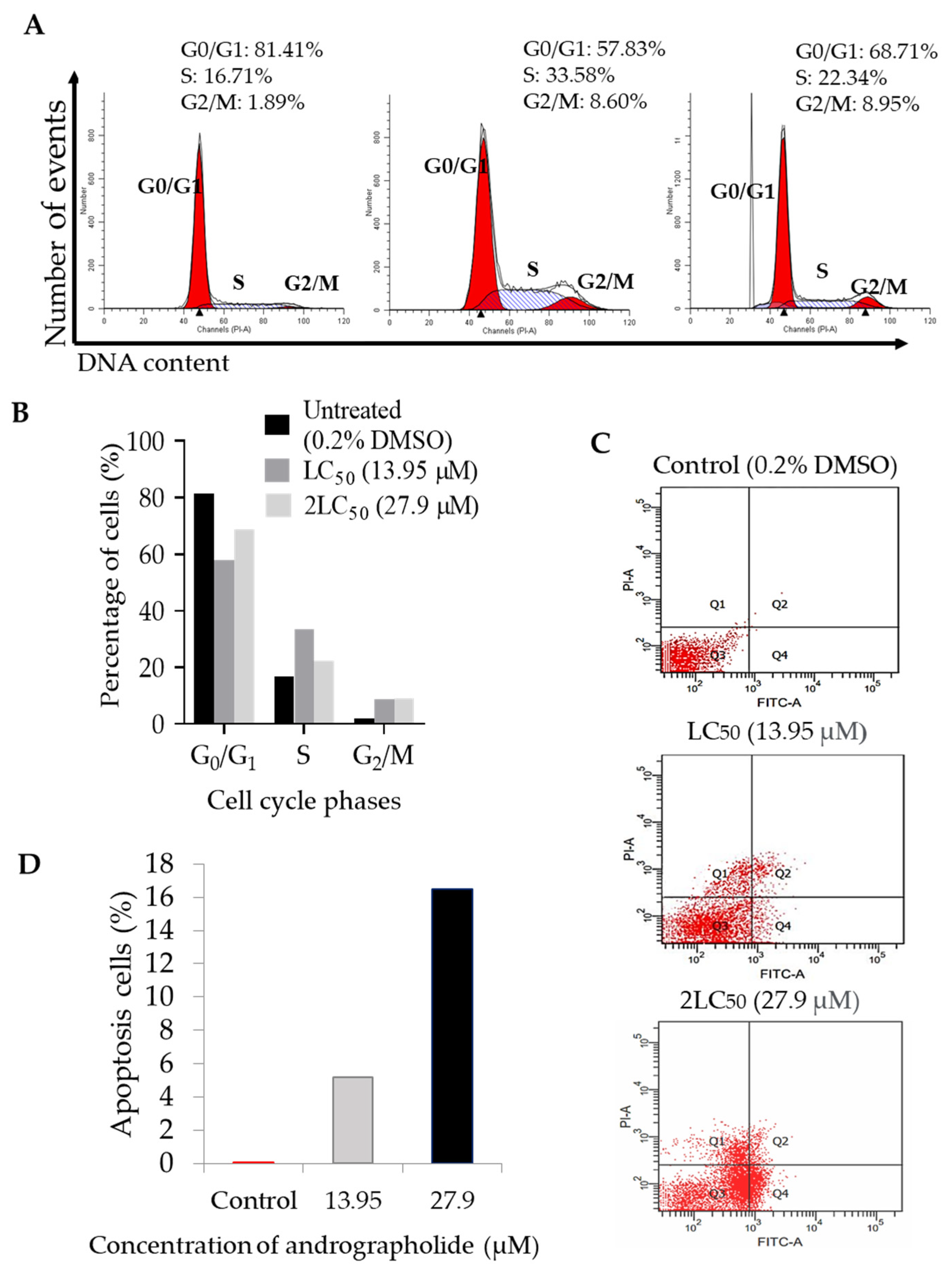
2.4. Andrographolide Induces G2/M Cell Cycle Arrest Followed by Apoptosis in DBTRG-05MG Cancer Cell Lines
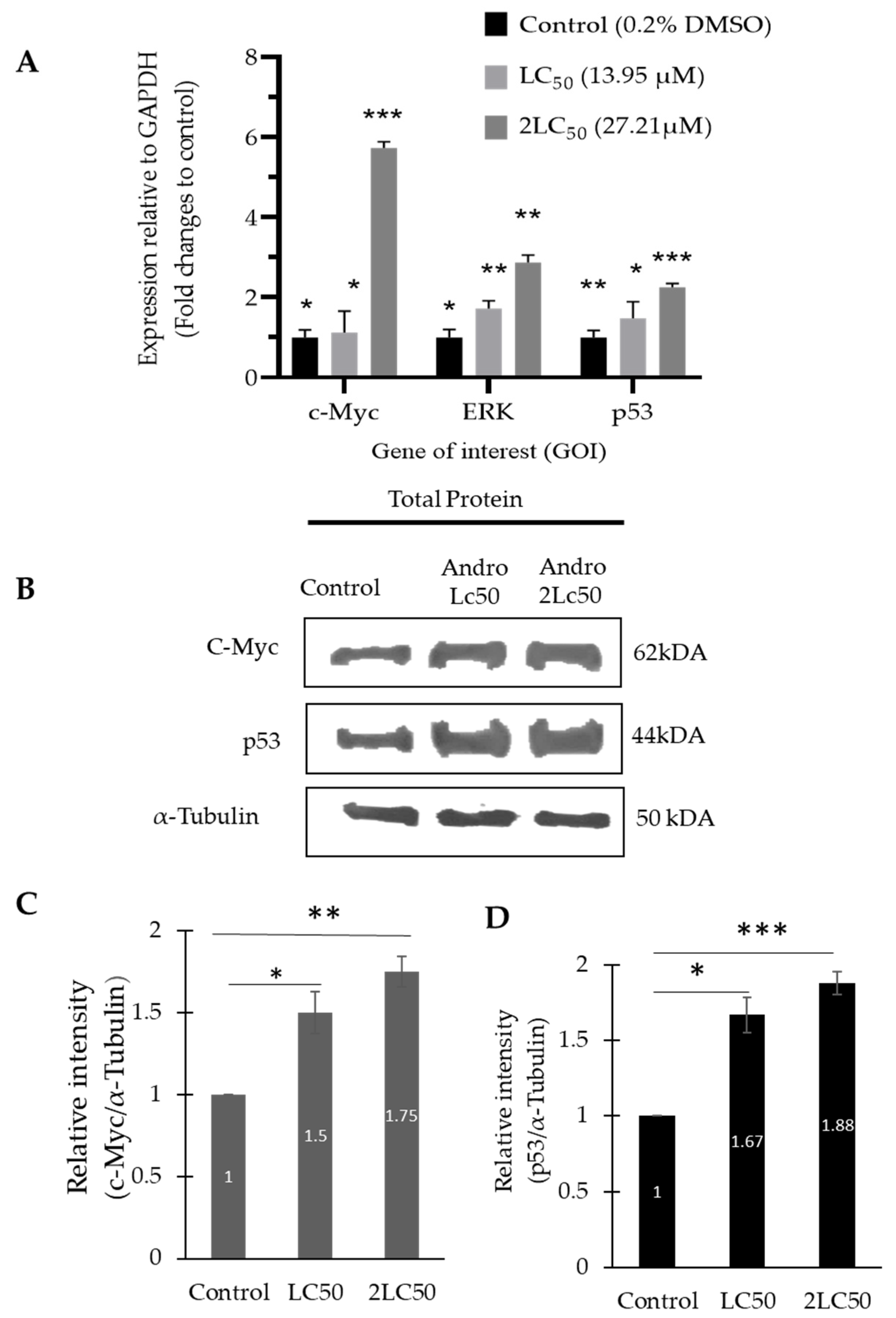
2.5. The mRNA and Protein Expression Level of ERK1/2, p53, and c-Myc Was Increased in DBTRG-05MG Treated with Andrographolide
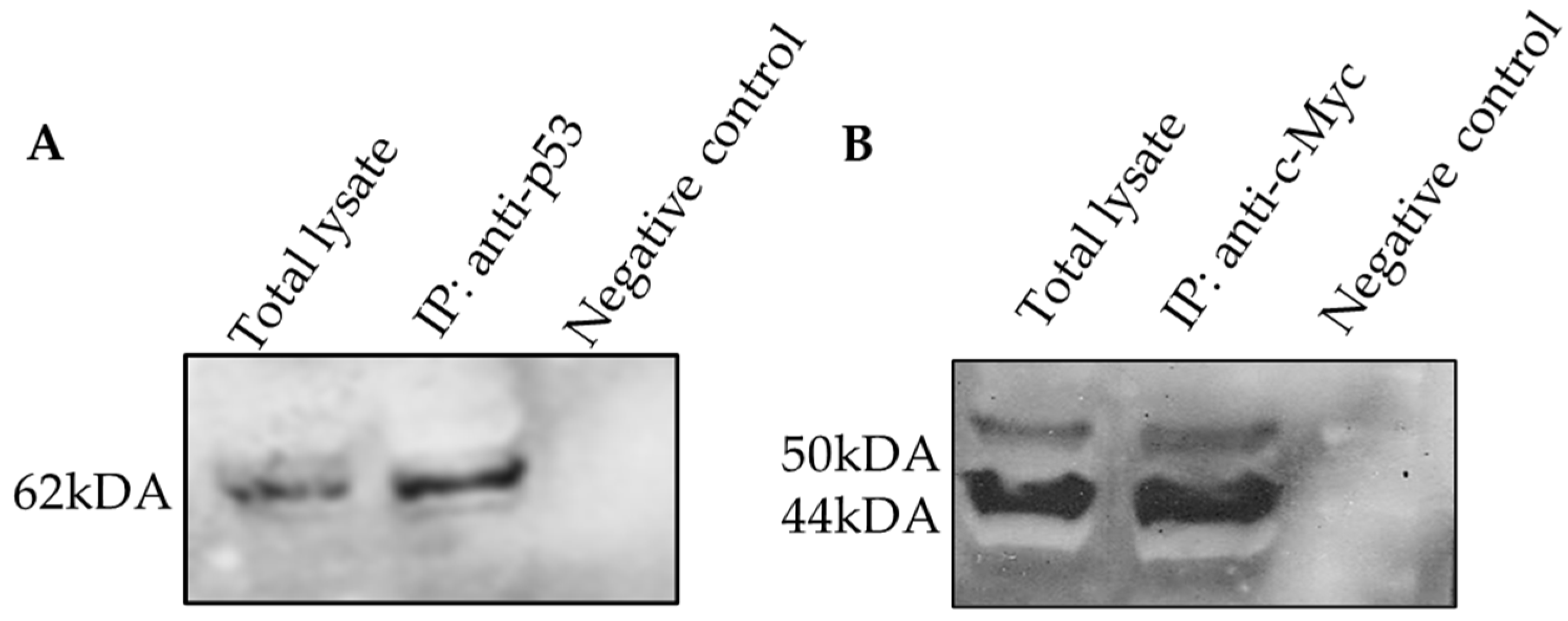
2.6. The Protein–Protein Interaction of c-Myc and p53 Involved in DBTRG-05MG Treated with Andrographolide
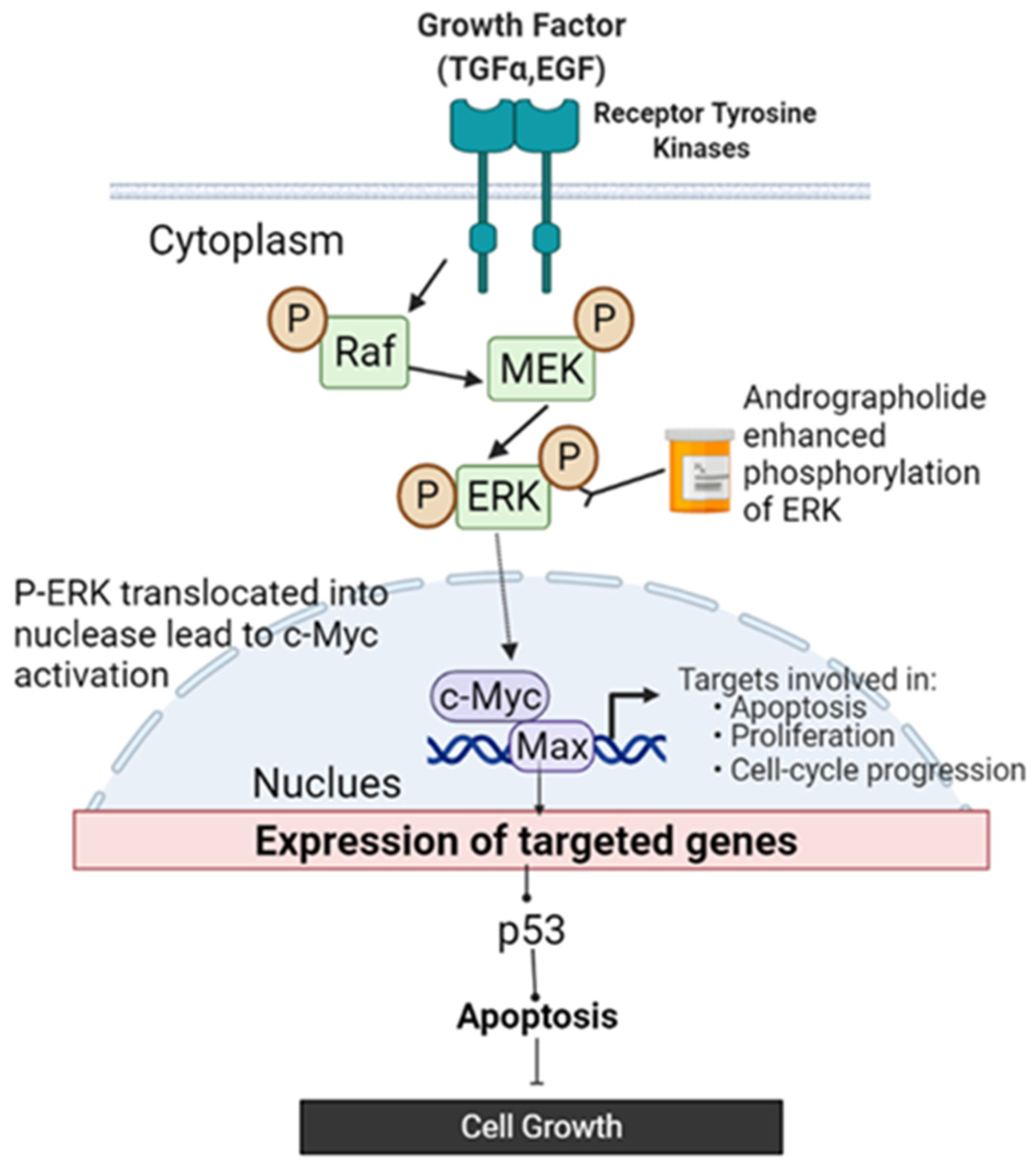
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Chemical Reagents
4.1.2. Human Cell Lines
4.2. Methods
4.2.1. Cytotoxic Activity Assay
4.2.2. Scratch Assay
4.2.3. Clonogenic Assay
4.2.4. Cell Cycle and Apoptosis Analysis
4.2.5. Isolation RNA
4.2.6. Quantitative Real-Time Reverse Transcription–Polymerase Chain Reaction (qRT-PCR)
4.2.7. Preparation of Total Cell Lysate
4.2.8. Co-Immunoprecipitation
4.2.9. Western Blot
4.2.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current Standards of Care in Glioblastoma Therapy. In Glioblastoma; Codon Publications: Brisbane, Australia, 2017; pp. 197–241. [Google Scholar]
- Prager, G.W.; Braga, S.; Bystricky, B.; Qvortrup, C.; Criscitiello, C.; Esin, E.; Sonke, G.S.; Martínez, G.A.; Frenel, J.S.; Karamouzis, M.; et al. Global Cancer Control: Responding to the Growing Burden, Rising Costs and Inequalities in Access. ESMO Open 2018, 3, e000285. [Google Scholar] [CrossRef] [Green Version]
- Sahu, M.; Rao, S.P. Review Literature: Andrographis Paniculata. Res. J. Pharmacol. Pharmacodyn. 2018, 10, 166. [Google Scholar] [CrossRef]
- Mussard, E.; Cesaro, A.; Lespessailles, E.; Legrain, B.; Berteina-Raboin, S.; Toumi, H. Andrographolide, A Natural Antioxidant: An Update. Antioxidants 2019, 8, 571. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Ma, Y.; Wu, J.; Huang, H.; Wang, X.; Chen, Z.; Chen, J.; He, H.; Huang, C. A Review for the Neuroprotective Effects of Andrographolide in the Central Nervous System. Biomed. Pharmacother. 2019, 117, 109078. [Google Scholar] [CrossRef]
- Yang, R.; Liu, S.; Zhou, J.; Bu, S.; Zhang, J. Andrographolide Attenuates Microglia-Mediated Aβ Neurotoxicity Partially through Inhibiting NF-ΚB and JNK MAPK Signaling Pathway. Immunopharmacol. Immunotoxicol. 2017, 39, 276–284. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Islam, M.A.; Shaw, S.; Khan, I.N.; Saravi, S.S.S.; Ahmad, S.; Rehman, S.; Gupta, V.K.; et al. Andrographolide, a Diterpene Lactone from Andrographis Paniculata and Its Therapeutic Promises in Cancer. Cancer Lett. 2018, 420, 129–145. [Google Scholar] [CrossRef]
- Liu, G.; Chu, H. Andrographolide Inhibits Proliferation and Induces Cell Cycle Arrest and Apoptosis in Human Melanoma Cells. Oncol. Lett. 2018, 15, 5301–5305. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Mahfooz, S.; Faisal, M.; Alatar, A.A.; Ansari, I.A. Andrographolide Induces Apoptosis and Cell Cycle Arrest through Inhibition of Aberrant Hedgehog Signaling Pathway in Colon Cancer Cells. Nutr. Cancer 2021, 73, 2428–2446. [Google Scholar] [CrossRef]
- Forestier-Román, I.S.; López-Rivas, A.; Sánchez-Vázquez, M.M.; Rohena-Rivera, K.; Nieves-Burgos, G.; Ortiz-Zuazaga, H.; Torres-Ramos, C.A.; Martínez-Ferrer, M. Andrographolide Induces DNA Damage in Prostate Cancer Cells. Oncotarget 2019, 10, 1085–1101. [Google Scholar] [CrossRef] [Green Version]
- Vetvicka, V.; Vannucci, L. Biological Properties of Andrographolide, an Active Ingredient of Andrographis Paniculata: A Narrative Review. Ann. Transl. Med. 2021, 9, 1186. [Google Scholar] [CrossRef]
- Yang, T.; Yao, S.; Zhang, X.; Guo, Y. Andrographolide Inhibits Growth of Human T-Cell Acute Lymphoblastic Leukemia Jurkat Cells by Downregulation of PI3K/AKT and Upregulation of P38 MAPK Pathways. Drug Des. Devel. Ther. 2016, 10, 1389–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Zarabi, A.; Hushmandi, K.; Moghadam, E.R.; Hashemi, F.; Daneshi, S.; Hashemi, F.; Tavakol, S.; Mohammadinejad, R.; Najafi, M.; et al. C-Myc Signaling Pathway in Treatment and Prevention of Brain Tumors. Curr. Cancer Drug Targets 2020, 21, 2–20. [Google Scholar] [CrossRef]
- Feng, J.; Ma, J.; Liu, S.; Wang, J.; Chen, Y. A Noncoding RNA LINC00504 Interacts with C-Myc to Regulate Tumor Metabolism in Colon Cancer. J. Cell. Biochem. 2019, 120, 14725–14734. [Google Scholar] [CrossRef]
- Ji, W.; Zhang, W.; Wang, X.; Shi, Y.; Yang, F.; Xie, H.; Zhou, W.; Wang, S.; Guan, X. C-Myc Regulates the Sensitivity of Breast Cancer Cells to Palbociclib via c-Myc/MiR-29b-3p/CDK6 Axis. Cell Death Dis. 2020, 11, 760. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, Y.; Pan, Y.; Ding, X.; Rhea, P.; Ding, J.; Hawke, D.H.; Felsher, D.; Narla, G.; Lu, Z.; et al. Mistletoe Extract Fraxini Inhibits the Proliferation of Liver Cancer by Down-Regulating c-Myc Expression. Sci. Rep. 2019, 9, 6428. [Google Scholar] [CrossRef] [Green Version]
- Sur, S.; Nakanishi, H.; Steele, R.; Ray, R.B. Depletion of PCAT-1 in Head and Neck Cancer Cells Inhibits Tumor Growth and Induces Apoptosis by Modulating c-Myc-AKT1-P38 MAPK Signalling Pathways. BMC Cancer 2019, 19, 354. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, G.J. Emerging Roles of Myc in Stem Cell Biology and Novel Tumor Therapies. J. Exp. Clin. Cancer Res. 2018, 37, 173. [Google Scholar] [CrossRef] [Green Version]
- Farooqi, A.A.; Attar, R.; Uteuliyev, Y.S.; Alaaeddine, N.; de Sousa, D.P.; Xu, B.; Cho, W.C. The Prowess of Andrographolide as a Natural Weapon in the War against Cancer. Cancers 2020, 12, 2159. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of Its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar]
- Zhang, H.; Li, S.; Si, Y.; Xu, H. Andrographolide and Its Derivatives: Current Achievements and Future Perspectives. Eur. J. Med. Chem. 2021, 224, 113710. [Google Scholar] [CrossRef]
- Chiu, S.P.; Batsaikhan, B.; Huang, H.M.; Wang, J.Y. Application of Electric Cell-Substrate Impedance Sensing to Investigate the Cytotoxic Effects of Andrographolide on U-87 MG Glioblastoma Cell Migration and Apoptosis. Sensors 2019, 19, 2275. [Google Scholar] [CrossRef]
- Yang, S.L.; Kuo, F.H.; Chen, P.N.; Hsieh, Y.H.; Yu, N.Y.; Yang, W.E.; Hsieh, M.J.; Yang, S.F. Andrographolide Suppresses the Migratory Ability of Human Glioblastoma Multiforme Cells by Targeting ERK1/2-Mediated Matrix Metalloproteinase-2 Expression. Oncotarget 2017, 8, 105860–105872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Chen, S.-R.; Chai, L.; Zhao, J.; Wang, Y.; Wang, Y. Overview of Pharmacological Activities of Andrographis Paniculata and Its Major Compound Andrographolide Overview of Pharmacological Activities of Andrographis Paniculata and Its Major Compound Andrographolide. Crit. Rev. Food Sci. Nutr. 2018, 59, S17–S29. [Google Scholar] [CrossRef]
- Yang, S.H.; Wang, S.M.; Syu, J.P.; Chen, Y.; Wang, S.D.; Peng, Y.S.; Kuo, M.F.; Kung, H.N. Andrographolide Induces Apoptosis of C6 Glioma Cells via the ERK-P53-Caspase 7-PARP Pathway. Biomed Res. Int. 2014, 2014, 312847. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK Signalling Pathway and Tumorigenesis (Review). Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, C.; Zhang, L.; Heng, Y.; Xu, T.; Zhang, Y.; Chen, X.; Hoffman, R.M.; Jia, L. Andrographolide Induces Noxa-Dependent Apoptosis by Transactivating ATF4 in Human Lung Adenocarcinoma Cells. Front. Pharmacol. 2021, 12, 924. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lovén, J.; Rahl, P.B.; Paranal, R.M.; Burge, C.B.; Bradner, J.E.; Lee, T.I.; Young, R.A. Transcriptional Amplification in Tumor Cells with Elevated C-Myc. Cell 2012, 151, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, S.E.; Rahimi, S.; Zarandi, B.; Chegeni, R.; Safa, M. MYC: A Multipurpose Oncogene with Prognostic and Therapeutic Implications in Blood Malignancies. J. Hematol. Oncol. 2020, 14, 121. [Google Scholar] [CrossRef]
- Jung, Y.; Kraikivski, P.; Shafiekhani, S.; Terhune, S.S.; Dash, R.K. Crosstalk between Plk1, P53, Cell Cycle, and G2/M DNA Damage Checkpoint Regulation in Cancer: Computational Modeling and Analysis. NPJ Syst. Biol. Appl. 2021, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Liu, Z.; Cheng, C.; Wang, H.; Deng, X.; Liu, J.; Liu, C.; Li, Y.; Fang, W. VPS33B Interacts with NESG1 to Modulate EGFR/PI3K/AKT/c-Myc/P53/MiR-133a-3p Signaling and Induce 5-Fluorouracil Sensitivity in Nasopharyngeal Carcinoma. Cell Death Dis. 2019, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Z.; Wang, H.; Tang, Z.; Liu, Y.; Liang, Z.; Deng, X.; Zhao, M.; Fu, Q.; Li, L.; et al. VPS33B Negatively Modulated by Nicotine Functions as a Tumor Suppressor in Colorectal Cancer. Int. J. Cancer 2020, 146, 496–509. [Google Scholar] [CrossRef]
- Reinhardt, S.; Groen, L.; Morten, K.; Bourdon, B.C.; Avery-Kiejda, J.-C.; Cytoplasmic, K.A.; Steffens Reinhardt, L.; Groen, K.; Morten, B.C.; Bourdon, J.-C.; et al. Cytoplasmic P53β Isoforms Are Associated with Worse Disease-Free Survival in Breast Cancer. Int. J. Mol. Sci. 2022, 23, 6670. [Google Scholar] [CrossRef]
- Rojas, E.A.; Corchete, L.A.; De Ramón, C.; Krzeminski, P.; Quwaider, D.; García-Sanz, R.; Martínez-López, J.; Oriol, A.; Rosiñol, L.; Bladé, J.; et al. Expression of P53 Protein Isoforms Predicts Survival in Patients with Multiple Myeloma. Am. J. Hematol. 2022, 97, 700–710. [Google Scholar] [CrossRef]
- Takahashi, R.; Giannini, C.; Sarkaria, J.N.; Schroeder, M.; Rogers, J.; Mastroeni, D.; Scrable, H. P53 Isoform Profiling in Glioblastoma and Injured Brain. Oncogene 2013, 32, 3165–3174. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Cell Line | Treatment (h)/LC50 Values (µM) | ||
| 24 | 48 | 72 | ||
| DBTRG-05MG | 42.82 | 27.21 | 13.93 | |
| SVGp12 | >200(n.d) 1 | >200(n.d) 1 | >200(n.d) 1 | |
| B | Cell Line | Treatment (h)/LC50 Values (µM) | ||
| 24 | 48 | 72 | ||
| DBTRG-05MG | 5.80 | 4.61 | 4.02 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othman, N.S.; Mohd Azman, D.K. Andrographolide Induces G2/M Cell Cycle Arrest and Apoptosis in Human Glioblastoma DBTRG-05MG Cell Line via ERK1/2 /c-Myc/p53 Signaling Pathway. Molecules 2022, 27, 6686. https://doi.org/10.3390/molecules27196686
Othman NS, Mohd Azman DK. Andrographolide Induces G2/M Cell Cycle Arrest and Apoptosis in Human Glioblastoma DBTRG-05MG Cell Line via ERK1/2 /c-Myc/p53 Signaling Pathway. Molecules. 2022; 27(19):6686. https://doi.org/10.3390/molecules27196686
Chicago/Turabian StyleOthman, Nurul Syamimi, and Daruliza Kernain Mohd Azman. 2022. "Andrographolide Induces G2/M Cell Cycle Arrest and Apoptosis in Human Glioblastoma DBTRG-05MG Cell Line via ERK1/2 /c-Myc/p53 Signaling Pathway" Molecules 27, no. 19: 6686. https://doi.org/10.3390/molecules27196686