
G-Protein Coupled Receptors in Human Sperm: An In Silico Approach to Identify Potential Modulatory Targets



Department of Medical Sciences, Institute of Biomedicine-iBiMED, University of Aveiro, 3810-193 Aveiro, Portugal
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(19), 6503; https://doi.org/10.3390/molecules27196503
Submission received: 26 July 2022
/
Revised: 28 September 2022
/
Accepted: 28 September 2022
/
Published: 1 October 2022
(This article belongs to the Special Issue Molecular Modeling, Synthesis, and Functional Characterization of GPCR (G-Protein Coupled Receptor) Ligands)
Abstract
:G protein-coupled receptors (GPCRs) are involved in several physiological processes, and they represent the largest family of drug targets to date. However, the presence and function of these receptors are poorly described in human spermatozoa. Here, we aimed to identify and characterize the GPCRs present in human spermatozoa and perform an in silico analysis to understand their potential role in sperm functions. The human sperm proteome, including proteomic studies in which the criteria used for protein identification was set as <5% FDR and a minimum of 2 peptides match per protein, was crossed with the list of GPCRs retrieved from GLASS and GPCRdb databases. A total of 71 GPCRs were identified in human spermatozoa, of which 7 had selective expression in male tissues (epididymis, seminal vesicles, and testis), and 9 were associated with male infertility defects in mice. Additionally, ADRA2A, AGTR1, AGTR2, FZD3, and GLP1R were already associated with sperm-specific functions such as sperm capacitation, acrosome reaction, and motility, representing potential targets to modulate and improve sperm function. Finally, the protein-protein interaction network for the human sperm GPCRs revealed that 24 GPCRs interact with 49 proteins involved in crucial processes for sperm formation, maturation, and fertilization. This approach allowed the identification of 8 relevant GPCRs (ADGRE5, ADGRL2, GLP1R, AGTR2, CELSR2, FZD3, CELSR3, and GABBR1) present in human spermatozoa that can be the subject of further investigation to be used even as potential modulatory targets to treat male infertility or to develop new non-hormonal male contraceptives.
1. Introduction
Spermatozoa are highly differentiated haploid cells generated during spermatogenesis, a complex network of mitotic and meiotic divisions that occurs within seminiferous tubules in the testis [1]. Despite being morphologically differentiated when they leave the seminiferous tubules, spermatozoa are functionally immature and unable to fertilize oocytes, requiring two additional maturation steps: epididymal maturation and capacitation [2,3,4]. The first occurs in the epididymis, where spermatozoa acquire progressive motility and their morphological features are tuned [2]. The epididymal biochemical environment induces changes in intracellular concentrations of Ca2+, cAMP, and pH, promoting a shift in protein phosphorylation status, which is fundamental for motility acquisition [2,5,6]. The second maturation round, capacitation, occurs in the female reproductive tract, making spermatozoa capable of attaching to and penetrating the oocyte zona pellucida [3,4]. During this process, spermatozoa acquire hyperactivated motility, membranes become more fluid, and intracellular levels of Ca2+, cAMP, and pH change leading to the activation of protein kinases [7,8,9]. The successful completion of these processes allows spermatozoa to reach and fertilize the oocyte, the main goal of the sperm life journey.
G protein-coupled receptors (GPCRs), the largest family of receptors described, are seven-transmembrane domain proteins that are coupled to different G protein subtypes or arrestins [10,11,12]. These receptors detect molecules, such as hormones, neurotransmitters, chemokines, or odorants [13]. The binding of these ligands induces conformational changes in the transmembrane and intracellular domains of the receptor, allowing its interaction with heterotrimeric G proteins or with arrestins [14,15]. After activation, GPCRs act as guanine nucleotide exchange factors (GEFs) for the α subunits of heterotrimeric G proteins, catalyzing the release of GDP and the binding of GTP for G-protein activation. Activation of different G proteins (e.g., Gi/o, Gs, G12/13, or Gq/11 subtypes) affects several cellular processes through different downstream effectors, such as adenylyl cyclase (AC), RhoGEF, or phospholipase C (PLC) [12,16,17,18,19]. Active GPCRs may also be phosphorylated by GPCR kinases (GRKs), a family of protein kinases that phosphorylate specific serine/threonine residues of GPCRs. GPCR phosphorylation leads to arrestin recruitment and activation [20,21,22,23], leading to receptor desensitization and also mediating a second wave of signaling independent of G proteins [22,23,24,25,26,27].
It is known that GPCRs regulate a variety of physiological and pathological mechanisms, and their role in cardiovascular and neuronal disease, diabetes, and other disorders has been investigated [24,28,29,30]. Furthermore, GPCRs represent an important and attractive group of drug targets, with more than 470 GPCR-targeted drugs currently in the clinical market [31,32,33]. Being key players in cell signaling events, the GPCRs’ presence and function in human spermatozoa are poorly addressed. Indeed, only a couple of articles are focused on this topic [34,35,36,37,38,39,40], and, to the best of our knowledge, no systematic and comprehensive review on this topic exists in the literature. The main objective of the present work was to identify the GPCRs present in human spermatozoa and to perform an in silico analysis to understand their potential role in sperm-related functions. Through this approach, it was possible the identification of relevant sperm GPCRs that can be the subject of further investigation (i) to be used as potential modulatory targets to treat male fertility or (ii) to develop new non-hormonal male contraceptives.
2. Results and Discussion
2.1. GPCRs in Human Sperm
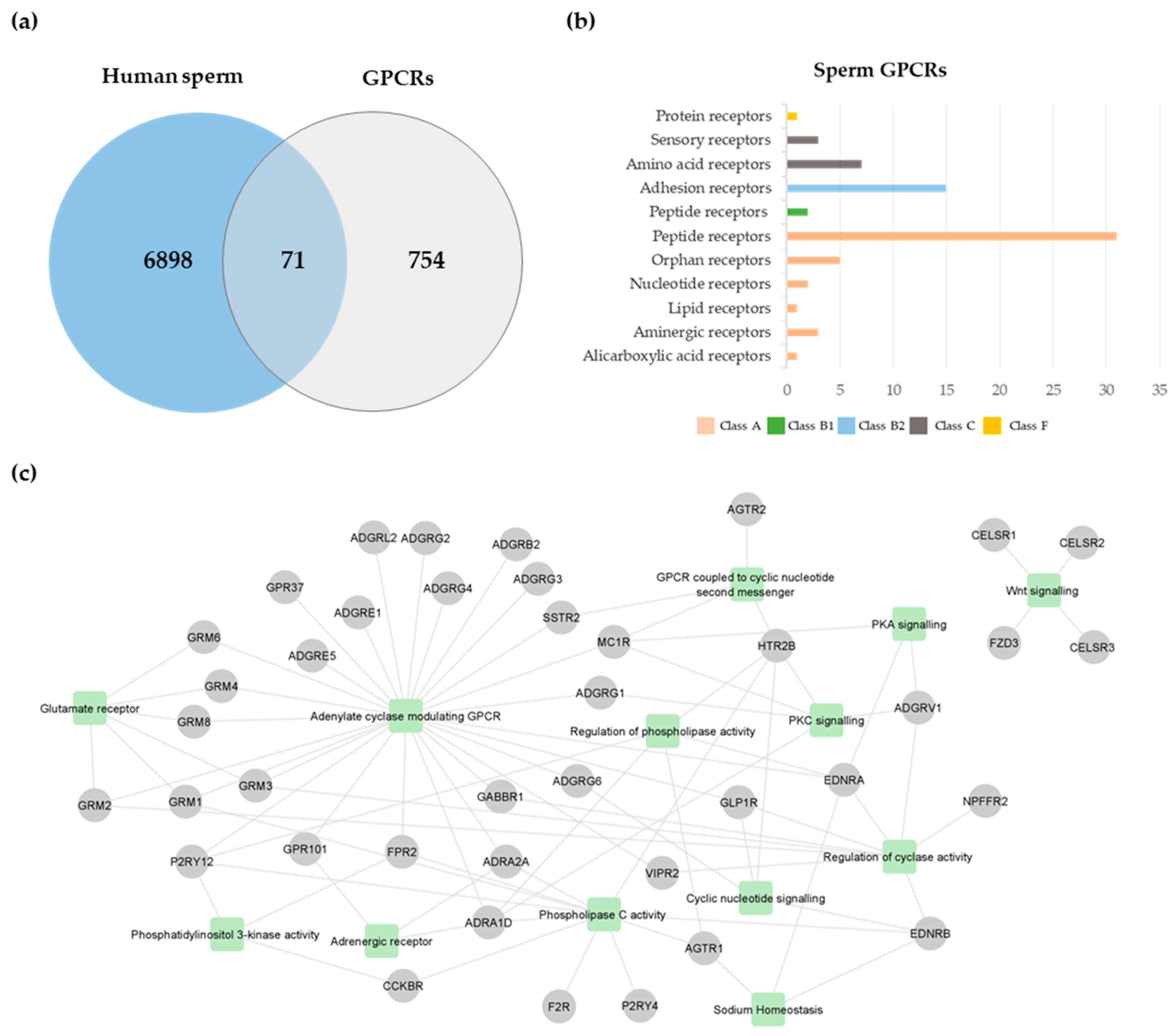
A total of 825 GPCRs were retrieved from GLASS and GPCRdb databases (Table S1). This list was cross-compared with the human sperm proteome previously published [41], revealing the existence of 71 GPCRs in human spermatozoa (Figure 1a; Table 1). From those, only type-1 angiotensin II receptor (AGTR1) [42], AGTR2 [43,44], glucagon-like peptide 1 receptor (GLP1R) [45], and olfactory receptor 4C13 (OR4C13) [40] were previously studied in human spermatozoa. According to the GPCRdb [46], the human sperm GPCRs were members of five classes: class A, rhodopsin (n = 42); class B1, secretin (n = 2); class B2, adhesion (n = 16), class C, glutamate (n = 10); and class F, frizzled (n = 1) (Figure 1b; Table 1). Considering that the human sperm proteome used [41] only included proteomic studies performed using ejaculated human spermatozoa. in which a false discovery rate (FDR) <5% of protein identification was set, and proteins identified with at least two peptides were included, some GPCRs identified by other techniques were excluded. For instance, some GPCRs previously identified through immunocytochemistry and Western blot techniques in human spermatozoa [34,35,37,38,39,40] were not identified through our analysis. To maintain the proposed methodology, these GPCRs were not considered in the subsequent analyses.
The selective tissue expression and male reproductive phenotypes associated with the 71 GPCRs identified were searched in the Human Protein Atlas (HPA) and Mouse Genome Informatics (MGI) databases, respectively. Among those, 7 had selective expression in male tissues, and 9 were associated with male infertility defects in mice (Table 2). ADGRG2 is enriched in the epididymis, CYSLTR2, EDNRA, FZD3, and NPFFR2 are enhanced in the seminal vesicle, CELSR1 is enriched in the seminal vesicle, and GRM2 is enhanced in the testis (Table 2). Regarding male reproductive phenotypes: ADGRV1, ADRA2A, ADGRG1, CELSR1, and VIPR2 were associated with abnormal testis morphology; F2R, GRM1 were associated with abnormal fertility/fecundity; ADGRG2 was associated with absent vas deferens and azoospermia; GRM1, ADGRG1, and HTR2B were associated with reduced fertility; and GRM1 and VIPR2 were associated with male infertility (Table 2). Despite not being described in the MGI database, prosaposin receptor GPR37 null mice presented abnormal testis morphology, altered spermatogenesis, and decreased sperm concentration and motility [47]. Additionally, Agtr1 knock-out mice had abnormal sperm morphology and male infertility, probably due to abnormal spermatogenesis [48].
ADGRG2 has been pointed out as a potential target for the development of a non-testicular, non-steroid male contraceptive [49]. ADGRG2 is an adhesion G-protein coupled receptor highly expressed in efferent ducts and proximal epididymis, being a key player in the signaling pathways that promote fluid reabsorption in the efferent ductus and sperm maturation [50,51,52]. In men, X-linked ADGRG2 deleterious mutations have been associated with congenital bilateral absence of the vas deferens [53], one of the causes underlying obstructive azoospermia. Those men presented normal spermatogenesis and were able to conceive a child through assisted reproduction techniques [53]. Nevertheless, the ADGRG2 role in sperm physiology is unknown. A proteomic study revealed increased ADGRG2 levels in spermatozoa from asthenozoospermic men [54], which may suggest a role of this receptor in sperm motility. On the other hand, ADGRG1, ADGRV1, ADRA2A, and VIPR2 were associated with histological testicular defects and impaired spermatogenesis, which make them unattractive targets for the development of drugs for male contraception since they can negatively and permanently impact the testicular environment and thus, spermatogenesis and sperm maturation. HTR2B, GRM1, and F2R were already associated with reduced or abnormal fertility; however, how they lead to this phenotype is still unknown. Hence, more studies focusing on the role and function of these GPCRs in human sperm physiology and fertilization should be performed.
2.2. Biological Processes Associated with Sperm GPCRs
An enrichment analysis was performed to identify significant biological processes associated with the 71 sperm GPCRs. Ninety-one gene ontology (GO) terms related to biological processes were retrieved (Table S2). As expected, the most significant terms were “G protein-coupled receptor signaling pathway” (GO:0007186, p-value = 0.0) and “G protein-coupled receptor activity” (GO:0004930, p-value = 0.0). Since spermatozoa are a highly specific cell type, the GO term list was filtered to highlight relevant processes for sperm physiology. A total of 30 GO terms were retrieved (Table S2), being grouped into 13 generic classes: adenylate cyclase modulating GPCR (GO:0007188; GO:0007193; GO:0007189), glutamate receptor (GO:0007196; GO:0001640; GO:0007215), adrenergic receptor (GO:0071880; GO:0071875), GPCR coupled to cyclic nucleotide second messenger (GO:0007187), cyclic nucleotide signaling (GO:0019935), phospholipase C activity (GO:0007200; GO:0051482), regulation of phospholipase activity (GO:0010518; GO:1900274; GO:0010863; GO:0010517), regulation of cyclase activity (GO:0045761; GO:0007194; GO:0031280; GO:0031279; GO:0007190), phosphatidylinositol 3-kinase activity (GO:0035004; GO:0043551), PKA signaling (GO:0010737), PKC signaling (GO:0070528; GO:0090037; GO:0090036), sodium homeostasis (GO:0055078), and Wnt signaling (GO:0060071; GO:0035567) (Figure 1c; Table S2). Forty sperm GPCRs were associated with those categories (Figure 1c), which may indicate that those GPCRs have a role in the retrieved biological processes.
Although this analysis did not reveal any association between GPCRs and sperm-specific processes, ADRA2A, AGTR1, AGTR2, FZD3, and GLP1R have been implicated in sperm-specific functions. The ADRA2A stimulates the cAMP production and accelerates mouse sperm capacitation [55]. AGTR1 also stimulates mammalian sperm capacitation, acrosome reaction, and motility [42,56,57,58,59,60]. In men, increased AGTR2 expression levels were positively correlated with progressive motility, suggesting that sperm motility may be regulated by this GPCR [43,44]. In porcine spermatozoa, FZD3 was reported to have a regulatory role in in vitro capacitation and acrosome exocytosis [61]. GLP1R stimulation by exenatide (a GLP1R agonist) increased human sperm motility and glucose metabolism [45]. Additionally, high fat-induced obese mice treated with exenatide presented improved sperm quality regarding motility, mitochondrial metabolism, and DNA integrity parameters [62]. Nevertheless, since exenatide treatment may directly impact sperm quality, the mice’s weight reduction may also contribute to sperm quality improvement. Considering their roles in sperm-specific functions, AGTR1, AGTR2, FZD3, and GLP1R are potential targets for further investigation and to modulate sperm function.
2.3. Sperm GPCRs Interactions
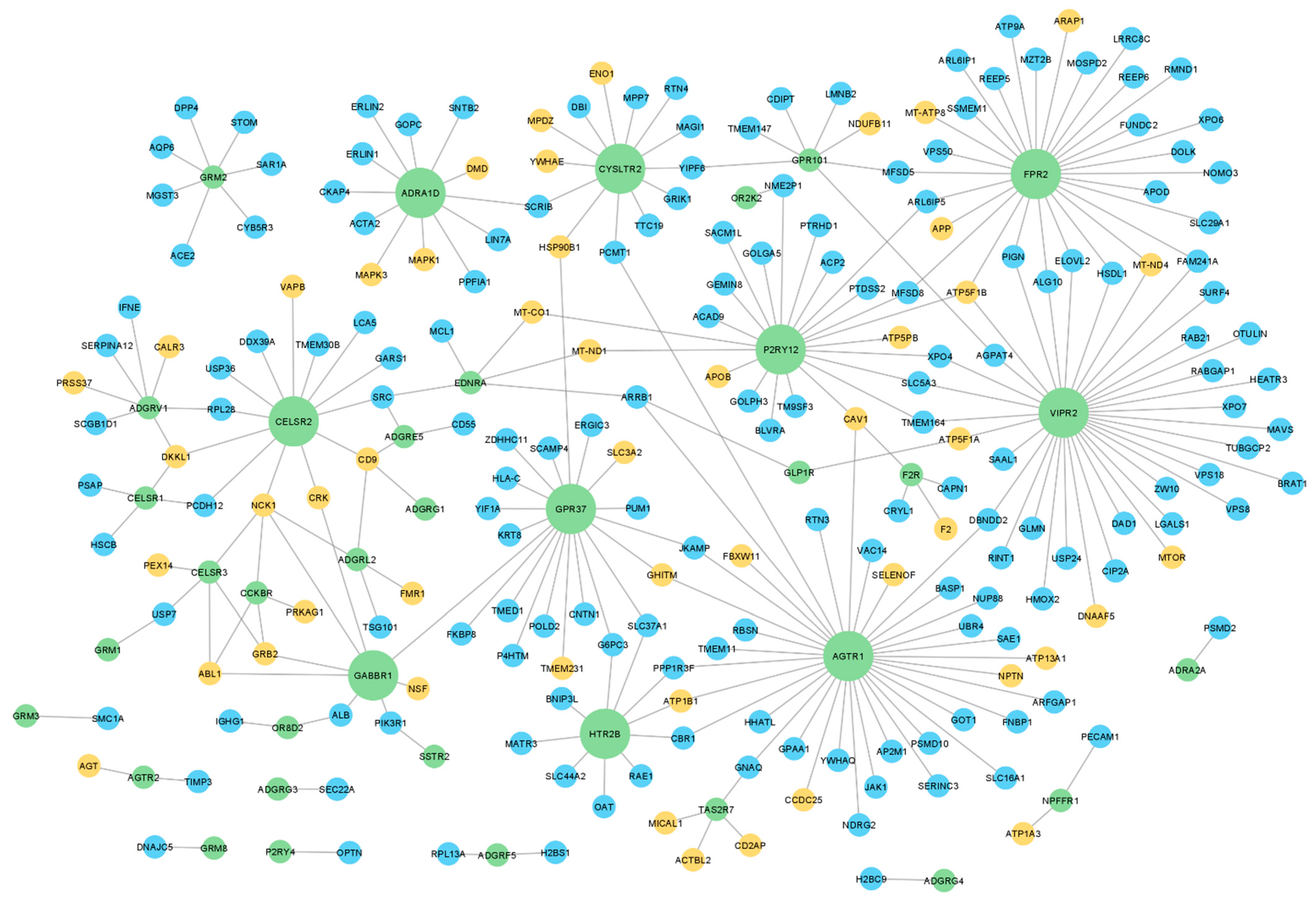
To investigate whether GPCRs present in human sperm interact with relevant proteins for sperm physiology, a protein-protein interaction (PPI) network for the human sperm GPCRs was retrieved using the IMEx data (through the Cytoscape). Only 47 sperm GPCRs had described interactions and were included in this network, for which 212 sperm interactors were retrieved, with a total of 270 interactions among sperm GPCRs and those interactors (Figure 2, Table S3). Regarding the node degree (i.e., number of connected edges to a node in the network), the top 10 GPCRs with a higher degree were highlighted: VIPR2, AGTR1, N-formyl peptide receptor 2 (FPR2), P2Y purinoceptor 12 (P2RY12), GPR37, CELSR2, CYSLTR2, alpha-1D adrenergic receptor (ADRA1D), HTR2B, and gamma-aminobutyric acid type B receptor subunit 1 (GABBR1) (bigger green nodes in Figure 2). This result highlighted that the same GPCRs may be involved in different signaling pathways. A good example of that is the AGTR1, which was associated with capacitation, acrosome reaction, and motility in mammalian sperm, all distinct sperm-related processes [42,56,57,58,59,60].
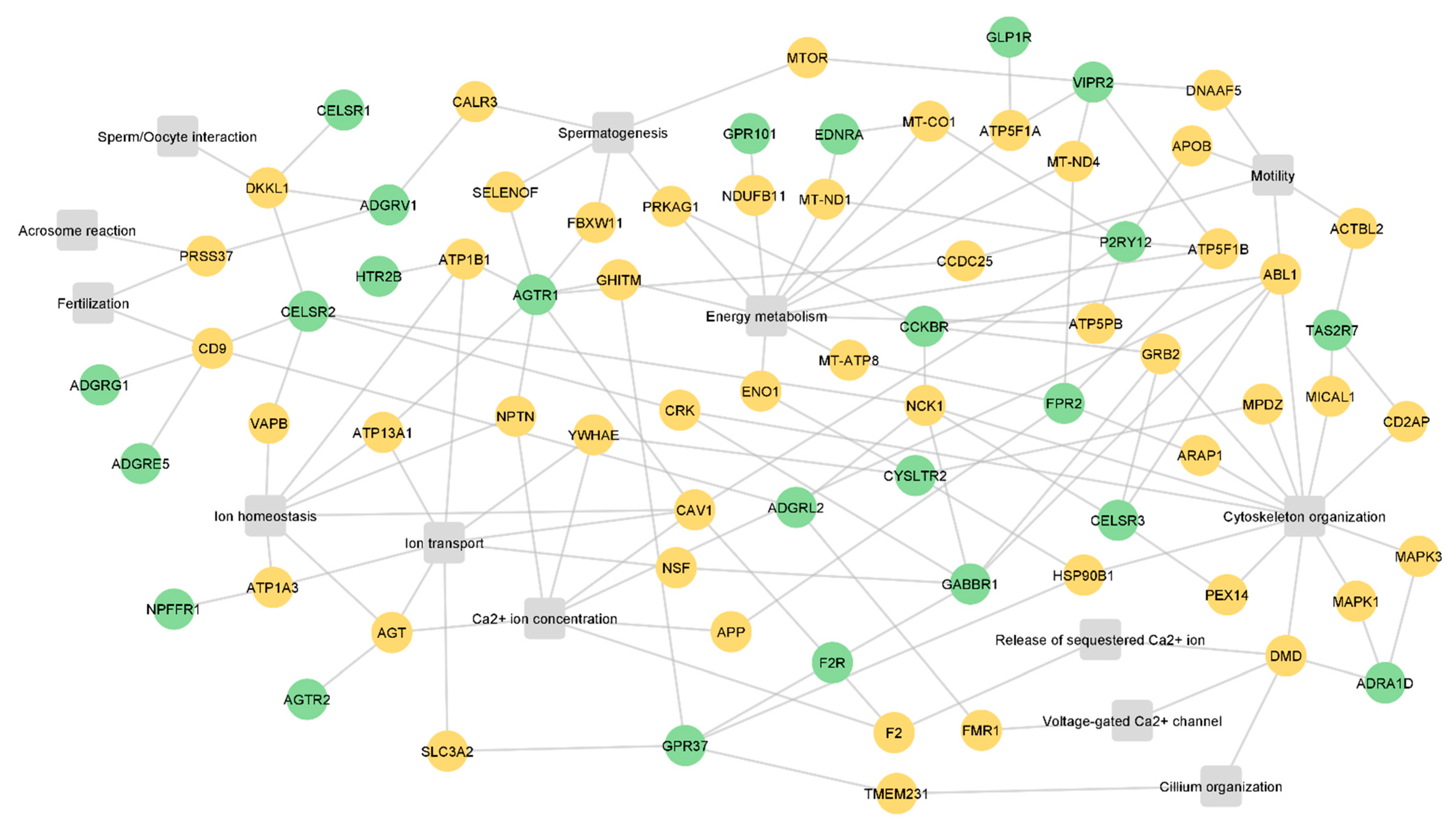
Next, to infer the potential role of the GPCRs in sperm physiology, the interactors were searched in the UniProt database to identify relevant biological processes to sperm physiology. Of the 212 identified GPCR interactors, 49 were associated with the following processes: spermatogenesis, cilium organization, cytoskeleton organization, Ca2+ ion concentration, the release of sequestered Ca2+ ion, voltage-gated Ca2+ channels, ion homeostasis, ion transport, energy metabolism, motility, sperm/oocyte interactions, acrosome reactions, and fertilization (Figure 3, Table S3). The identified processes are associated with sperm formation and maturation, as well as with fertilization. Only 24 sperm GPCRs were associated with these 49 interactors (Figure 3). ADGRC2, ADGRG1, ADGRE5, ADGRV1, AGTR1, gastrin/cholecystokinin type B receptor (CCKBR), CELSR1, CELSR2, CELSR3, GABBR1, P2RY12, taste receptor type 2 member 7 (TAS2R7), and VIPR2 interact with proteins related to sperm-specific processes (motility, sperm/oocyte interactions, acrosome reaction, and fertilization) (Figure 3).
The use of small molecules that target GPCRs involved in sperm motility regulation (for example, P2RY12 and TAS2R7) may be a useful strategy for developing non-hormonal male contraceptives targeting only post-testicular sperm maturation, stopping sperm. Additionally, the use of natural or synthetic drugs that activate or inhibit GPCRs associated with low sperm motility has the potential to be used as an additive in sperm cryopreservation and preparation media to improve the motility of samples used for artificial insemination and in vitro fertilization. On the other hand, the modulation of ADGRV1, AGTR1, CCKBR, and VIPR2, which interact with proteins related to spermatogenesis, is considerably more difficult since (i) the drugs used need to cross the blood-testis-barrier to reach the germ cells niche; (ii) the spermatogenic process may be irreversibly impacted; (iii) other signaling pathways may be activated or inhibited, resulting in unexpected phenotypes. Both Figure 2 and Figure 3 showed that some interactors are common to different GPCRs, which suggests two hypotheses: (i) different GPCRs can modulate the same biological process in the sperm (for example, CELSR1, ADGRV1, and CELSR2); (ii) the common interactors may be involved in different signaling pathways that are dependent on the activated GPCRs (for example, ABL1 and PRKAG1). Considering this, caution should be taken when selecting the target to modulate to avoid disturbing other important signaling pathways. Therefore, the selection of the best GPCR to modulate, both for infertility treatment or for contraception, should take into consideration its involvement in other physiological processes in sperm and even in other cell types. The selection of a sperm-specific GPCR can overcome this limitation; however, at least in this study, none of the GPCRs identified are exclusively present in sperm.
3. Materials and Methods
3.1. Collection of GPCRs in Human Spermatozoa
To collect the human GPCRs, the GPCR-Ligand Association (GLASS; Ann Arbor; USA) and GPCRdb (Copenhagen, Denmark) databases were used (downloaded on 7 March 2022) [46,63]. To avoid redundancy, all GPCRs were mapped in the UniProt database (Cambridge, UK) (downloaded on 7 March 2022), resulting in the final list of human GPCRs (Table S1). Through the JVenn tool [64], a Venn diagram analysis was performed to identify GPCRs in human sperm (Table 1). The human sperm proteome was obtained by Santiago et al. [41].
3.2. Detailed Molecular Insight of the Sperm GPCRs
To identify sperm GPCRs with selective expression in the male reproductive tract, the Human Protein Atlas (HPA; Stockholm, Sweden) (version 21.0, downloaded on 10 March 2022) was used [65]. Information related to male infertility defects and phenotypes was retrieved from the Mouse Genome Informatics (MGI) database (downloaded on 10 March 2022) [66]. To gain molecular insight into the sperm GPCR functions, a gene ontology (GO) enrichment analysis was performed for biological processes (Table S2). This analysis was performed through the ClueGO plug-in (version 2.5.8; Bethesda, MD, USA) in Cytoscape (version 3.8.2; Bethesda, MD, USA) [67]. The GO range was fixed between 3 and 8, and the perfusion option was used. Only terms with a p-value < 0.05 were considered. Lastly, all sperm GPCRs were searched in the PubMed database (Bethesda, MD, USA) to identify studies that report their role in mammalian spermatozoa (search performed before 24 May 2022).
3.3. Identification of Sperm GPCRs Interactome
To predict the potential roles of the sperm GPCRs, a protein-protein interaction (PPI) network was imported from the IMEx database through Cytoscape (version 3.8.2; Bethesda, MD, USA) [67,68] (downloaded on 10 March 2022). The imported PPI network was filtered by taxonomic name (“human”), and only sperm proteins [41] were kept. Duplicate edges and self-loops were removed. The topological properties of the network were assessed using the Network Analyzer tool [69]. To gain molecular insight into the sperm GPCR interactors (Table S3), the GO terms related to biological processes were collected from the UniProt database (Cambridge, UK) (downloaded on 12 March 2022).
4. Conclusions and Future Perspectives
The identification of GPCRs and their ligands/interactors in human sperm, and the characterization of their functions and downstream signaling pathways in sperm physiology, may provide future directions for new developments in the treatment of male infertility. To date, some GPCRs have been identified in mammalian sperm and implicated in sperm-related processes such as motility and acrosome reactions. In this work, we applied a bioinformatic approach to identify human sperm GPCRs and predict their function. To our best knowledge, this study was the first approach to list which GPCRs are presented in human spermatozoa. As stated above, some sperm GPCRs identified in individual studies were not included in the sperm proteome. This is justified by the evolution of the proteomic field, where the early studies only allowed the identification of a small number of proteins due to restrictive criteria [70]. Further sperm proteomic analyses may provide new updates in sperm proteome and consequently lead to the identification of more GPCRs in human spermatozoa. Additionally, proteomics can be useful for identifying sperm-specific GPCRs, which would be good targets to modulate sperm function(s).
Taking together all the information collected from the searched databases, we believe that at least 8 sperm GPCRs—ADGRE5, ADGRL2, GLP1R, AGTR2, CELSR2, FZD3, CELSR3, and GABBR1—are strong candidates for being explored in further research, not only to characterize their role in sperm but also to be considered potential targets for drug therapies. Those 8 GPCRs were directly associated with sperm functions or interacted with proteins with known functions in those processes. Milardi and colleagues hypothesized that OR4C13 may be involved in acrosome reaction and motility [40], which also makes it a potential research target. Nonetheless, our results did not associate OR4C13 with the mentioned sperm functions. ADGRG1, ADGRV1, CELSR1, and VIPR2 were implicated in sperm-related processes (Figure 3), but they were also associated with spermatogenesis (Figure 3) and testicular histological alterations (Table 2). Despite those GPCRs being potential targets for modulation in sperm, their function should be carefully evaluated in the context of the testicular environment.
One limitation of the present analysis is the near absence of sperm-specific information in databases. Most of the identified GPCRs (and even the identified interactors) are exhaustively explored in somatic cells, but their roles are poorly described in spermatozoon. Consequently, it is possible to draw conjectures about the potential role of the identified GPCRs, but additional experimental studies are needed for a complete characterization of their functions, interactions, and signaling pathways in human sperm.
To summarize, despite the activation/inhibition of specific GPCRs associated with sperm motility, acrosome reaction, or fertilization being particularly interesting for male contraception, more studies should be conducted to clarify the role of these GPCRs in human spermatozoa and the implications of their modulation for the whole organism. Additionally, we should take advantage of the several drugs targeting GPCRs already approved and on the market. After the identification of relevant GPCRs involved in poor sperm quality and infertility, high-throughput drug screening should be performed to assess the impact of these compounds in spermatozoa as a first step. The development of more potent and specific drugs targeting sperm GPCRs should then be investigated.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules27196503/s1. Table S1: Human G-protein coupled receptors described in GPCR-Ligand Association (GLASS) and GPCRdb databases; Table S2: Biological processes associated with human sperm GPCRs; Table S3: Human sperm GPCR interactors.
Author Contributions
Conceptualization, P.O.C., J.S. and M.F.; methodology, P.O.C.; formal analysis, P.O.C. and J.S.; investigation, P.O.C. and J.S.; writing—original draft preparation, P.O.C. and J.S.; writing—review and editing, P.O.C., J.S. and M.F.; supervision, M.F.; funding acquisition, M.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Institute for Biomedicine—iBiMED grant number UIDB/04501/2020 and by an individual grant to P.O.C. grant number 2020.10111.BD and J.S. grant number SFRH/BD/136896/2018, both financed by the Portuguese Foundation for Science and Technology (FCT) of the Portuguese Ministry of Science and Higher Education and by the European Union (QREN, FEDER and COMPETE frameworks).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in supplementary material here.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Not applicable.
References
- Turek, P.J. Male Reproductive Physiology. In Campbell-Walsh Urology; Wein, A.J., Kavoussi, L.R., Mcdougal, W.S., Peters, C.A., Partin, A.W., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2016; pp. 516–537. [Google Scholar]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.C. Fertilizing Capacity of Spermatozoa deposited into the Fallopian Tubes. Nature 1951, 168, 1039–1040. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.R. Observations on the penetration of the sperm in the mammalian egg. Aust. J. Sci. Res. B. 1951, 4, 581–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadnais, M.L.; Aghajanian, H.K.; Lin, A.; Gerton, G.L. Signaling in Sperm: Toward a Molecular Understanding of the Acquisition of Sperm Motility in the Mouse Epididymis1. Biol. Reprod. 2013, 89, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Vijayaraghavan, S. Increased Phosphorylation of a Distinct Subcellular Pool of Protein Phosphatase, PP1gamma2, During Epididymal Sperm Maturation. Biol. Reprod. 2004, 70, 439–447. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, C. Biological basis for human capacitation-revisited. Hum. Reprod. Update 2017, 23, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Puga Molina, L.C.; Buffone, M.G.; Luque, G.M.; Balestrini, P.A.; Marín-Briggiler, C.I.; Romarowski, A. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72. [Google Scholar] [CrossRef] [Green Version]
- Gangwar, D.; Atreja, S. Signalling Events and Associated Pathways Related to the Mammalian Sperm Capacitation. Reprod. Domest. Anim. 2015, 50, 705–711. [Google Scholar] [CrossRef]
- Bridges, T.M.; Lindsley, C.W. G-Protein-Coupled Receptors: From Classical Modes of Modulation to Allosteric Mechanisms. ACS Chem. Biol. 2008, 3, 530–541. [Google Scholar] [CrossRef]
- Cahill, T.J.; Thomsen, A.R.B.; Tarrasch, J.T.; Plouffe, B.; Nguyen, A.H.; Yang, F.; Huang, L.-Y.; Kahsai, A.W.; Bassoni, D.L.; Gavino, B.J.; et al. Distinct conformations of GPCR–β-arrestin complexes mediate desensitization, signaling, and endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, 2562–2567. [Google Scholar] [CrossRef]
- Ritter, S.L.; Hall, R.A. Fine-tuning of GPCR activity by receptor-interacting proteins. Nat. Rev. Mol. Cell Biol. 2009, 10, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and dynamics of GPCR signaling complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Kobilka, B.K. The Molecular Basis of G Protein–Coupled Receptor Activation. Annu. Rev. Biochem. 2018, 87, 897–919. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xiao, P.; Qu, C.; Liu, Q.; Wang, L.; Liu, Z.; He, Q.; Liu, C.; Xu, J.; Li, R.; et al. Allosteric mechanisms underlie GPCR signaling to SH3-domain proteins through arrestin. Nat. Chem. Biol. 2018, 14, 876–886. [Google Scholar] [CrossRef]
- Flock, T.; Ravarani, C.N.J.; Sun, D.; Venkatakrishnan, A.J.; Kayikci, M.; Tate, C.G.; Veprintsev, D.B.; Babu, M.M. Universal allosteric mechanism for Gα activation by GPCRs. Nature 2015, 524, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Flock, T.; Hauser, A.S.; Lund, N.; Gloriam, D.E.; Balaji, S.; Babu, M.M. Selectivity determinants of GPCR–G-protein binding. Nature 2017, 545, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Furness, S.G.B.; Liang, Y.-L.; Nowell, C.J.; Halls, M.L.; Wookey, P.J.; Dal Maso, E.; Inoue, A.; Christopoulos, A.; Wootten, D.; Sexton, P.M. Ligand-Dependent Modulation of G Protein Conformation Alters Drug Efficacy. Cell 2016, 167, 739–749.e11. [Google Scholar] [CrossRef] [Green Version]
- Venkatakrishnan, A.J.; Deupi, X.; Lebon, G.; Heydenreich, F.M.; Flock, T.; Miljus, T.; Balaji, S.; Bouvier, M.; Veprintsev, D.B.; Tate, C.G.; et al. Diverse activation pathways in class A GPCRs converge near the G-protein-coupling region. Nature 2016, 536, 484–487. [Google Scholar] [CrossRef] [Green Version]
- Homan, K.T.; Tesmer, J.J. Structural insights into G protein-coupled receptor kinase function. Curr. Opin. Cell Biol. 2014, 27, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Komolov, K.E.; Du, Y.; Duc, N.M.; Betz, R.M.; Rodrigues, J.P.G.L.M.; Leib, R.D.; Patra, D.; Skiniotis, G.; Adams, C.M.; Dror, R.O.; et al. Structural and Functional Analysis of a β 2-Adrenergic Receptor Complex with GRK5. Cell 2017, 169, 407–421.e16. [Google Scholar] [CrossRef]
- Yang, F.; Yu, X.; Liu, C.; Qu, C.-X.; Gong, Z.; Liu, H.-D.; Li, F.-H.; Wang, H.-M.; He, D.-F.; Yi, F.; et al. Phospho-selective mechanisms of arrestin conformations and functions revealed by unnatural amino acid incorporation and 19F-NMR. Nat. Commun. 2015, 6, 8202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, E.; Lefkowitz, R.J. GRKs and β-arrestins: Roles in receptor silencing, trafficking and signaling. Trends Endocrinol. Metab. 2006, 17, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Desimine, V.L.; McCrink, K.A.; Parker, B.M.; Wertz, S.L.; Maning, J.; Lymperopoulos, A. Biased Agonism/Antagonism of Cardiovascular GPCRs for Heart Failure Therapy. Int. Rev. Cell Mol. Biol. 2018, 339, 41–61. [Google Scholar] [PubMed]
- Dong, J.-H.; Wang, Y.-J.; Cui, M.; Wang, X.-J.; Zheng, W.-S.; Ma, M.-L.; Yang, F.; He, D.-F.; Hu, Q.-X.; Zhang, D.-L.; et al. Adaptive Activation of a Stress Response Pathway Improves Learning and Memory Through Gs and β-Arrestin-1–Regulated Lactate Metabolism. Biol. Psychiatry 2017, 81, 654–670. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J.; Shenoy, S.K. Transduction of Receptor Signals by ß-Arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef]
- Shukla, A.K.; Westfield, G.H.; Xiao, K.; Reis, R.I.; Huang, L.-Y.; Tripathi-Shukla, P.; Qian, J.; Li, S.; Blanc, A.; Oleskie, A.N.; et al. Visualization of arrestin recruitment by a G-protein-coupled receptor. Nature 2014, 512, 218–222. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Grotegut, C.A.; Wisler, J.W.; Mao, L.; Rosenberg, P.B.; Rockman, H.A.; Lefkowitz, R.J. The β-arrestin-biased β-adrenergic receptor blocker carvedilol enhances skeletal muscle contractility. Proc. Natl. Acad. Sci. USA 2020, 117, 12435–12443. [Google Scholar] [CrossRef]
- Lämmermann, T.; Kastenmüller, W. Concepts of GPCR-controlled navigation in the immune system. Immunol. Rev. 2019, 289, 205–231. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Yu, B.; Liu, Z.; Li, J.; Ma, M.; Wang, Y.; Zhu, M.; Yin, H.; Wang, X.; Fu, Y.; et al. Homocysteine directly interacts and activates the angiotensin II type I receptor to aggravate vascular injury. Nat. Commun. 2018, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Hauser, A.S.; Chavali, S.; Masuho, I.; Jahn, L.J.; Martemyanov, K.A.; Gloriam, D.E.; Babu, M.M. Pharmacogenomics of GPCR Drug Targets. Cell 2018, 172, 41–54.e19. [Google Scholar] [CrossRef]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.I.; et al. A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Muroi, T.; Matsushima, Y.; Kanamori, R.; Inoue, H.; Fujii, W.; Yogo, K. GPR62 constitutively activates cAMP signaling but is dispensable for male fertility in mice. Reproduction 2017, 154, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenker, C.; Goodwin, N.; Weyand, I.; Kashikar, N.D.; Naruse, M.; Krähling, M.; Müller, A.; Kaupp, U.B.; Strünker, T. The CatSper channel: A polymodal chemosensor in human sperm. EMBO J. 2012, 31, 1654–1665. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Wang, Y.; Qin, Z.; Yuan, X.; Zhang, Y.; Zeng, C. Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility. Genes 2021, 12, 488. [Google Scholar] [CrossRef]
- Pinto, F.M.; Cejudo-Román, A.; Ravina, C.G.; Fernández-Sánchez, M.; Martín-Lozano, D.; Illanes, M.; Tena-Sempere, M.; Candenas, M.L. Characterization of the kisspeptin system in human spermatozoa. Int. J. Androl. 2012, 35, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Michailov, Y.; Ickowicz, D.; Breitbart, H. Zn2+-stimulation of sperm capacitation and of the acrosome reaction is mediated by EGFR activation. Dev. Biol. 2014, 396, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Flegel, C.; Vogel, F.; Hofreuter, A.; Wojcik, S.; Schoeder, C.; Kieć-Kononowicz, K.; Brockmeyer, N.H.; Müller, C.E.; Becker, C.; Altmüller, J.; et al. Characterization of non-olfactory GPCRs in human sperm with a focus on GPR18. Sci. Rep. 2016, 6, 32255. [Google Scholar] [CrossRef]
- Milardi, D.; Colussi, C.; Grande, G.; Vincenzoni, F.; Pierconti, F.; Mancini, F.; Baroni, S.; Castagnola, M.; Marana, R.; Pontecorvi, A. Olfactory Receptors in Semen and in the Male Tract: From Proteome to Proteins. Front. Endocrinol. 2018, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.; Silva, J.V.; Fardilha, M. First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa. Int. J. Mol. Sci. 2019, 20, 5518. [Google Scholar] [CrossRef]
- Vinson, G.P.; Puddefoot, J.R.; Ho, M.M.; Barker, S.; Mehta, J.; Saridogan, E.; Djahanbakhch, O. Type 1 angiotensin II receptors in rat and human sperm. J. Endocrinol. 1995, 144, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Gianzo, M.; Muñoa-Hoyos, I.; Urizar-Arenaza, I.; Larreategui, Z.; Quintana, F.; Garrido, N.; Subirán, N.; Irazusta, J. Angiotensin II type 2 receptor is expressed in human sperm cells and is involved in sperm motility. Fertil. Steril. 2016, 105, 608–616. [Google Scholar] [CrossRef] [Green Version]
- El-Ariny, A.F.; El-Garem, Y.F.; Eid, A.A.; Donia, H.M.; Aly, R.G.; Abdelmeniem, I.M. Seminal levels of angiotensin II and angiotensin II type 2 receptor expression on spermatozoa in varicocele patients: Relation to fertility status. Andrologia 2020, 52, e13513. [Google Scholar] [CrossRef] [PubMed]
- Rago, V.; De Rose, D.; Santoro, M.; Panza, S.; Malivindi, R.; Andò, S.; D’Agata, R.; Aquila, S. Human Sperm Express the Receptor for Glucagon-like Peptide-1 (GLP-1), Which Affects Sperm Function and Metabolism. Endocrinology 2020, 161, bqaa031. [Google Scholar] [CrossRef] [PubMed]
- Kooistra, A.J.; Mordalski, S.; Pándy-Szekeres, G.; Esguerra, M.; Mamyrbekov, A.; Munk, C.; Keserű, G.M.; Gloriam, D.E. GPCRdb in 2021: Integrating GPCR sequence, structure and function. Nucleic Acids Res. 2021, 49, D335–D343. [Google Scholar] [CrossRef]
- La Sala, G.; Marazziti, D.; Di Pietro, C.; Golini, E.; Matteoni, R.; Tocchini-Valentini, G.P. Modulation of Dhh signaling and altered Sertoli cell function in mice lacking the GPR37-prosaposin receptor. FASEB J. 2015, 29, 2059–2069. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zou, Y.; Li, H.; Zhang, Y.; Liu, X.; Zhao, X.; Wu, X.; Fei, W.; Xu, Z.; Yang, X. Decreased angiotensin receptor 1 expression in ± AT1 Knockout mice testis results in male infertility and GnRH reduction. Reprod. Biol. Endocrinol. 2021, 19, 120. [Google Scholar] [CrossRef]
- Sipilä, P.; Jalkanen, J.; Huhtaniemi, I.T.; Poutanen, M. Novel epididymal proteins as targets for the development of post-testicular male contraception. Reproduction 2009, 137, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Davies, B.; Baumann, C.; Kirchhoff, C.; Ivell, R.; Nubbemeyer, R.; Habenicht, U.-F.; Theuring, F.; Gottwald, U. Targeted Deletion of the Epididymal Receptor HE6 Results in Fluid Dysregulation and Male Infertility. Mol. Cell. Biol. 2004, 24, 8642–8648. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-L.; Sun, Y.-J.; Ma, M.-L.; Wang, Y.; Lin, H.; Li, R.-R.; Liang, Z.-L.; Gao, Y.; Yang, Z.; He, D.-F.; et al. Gq activity- and β-arrestin-1 scaffolding-mediated ADGRG2/CFTR coupling are required for male fertility. Elife 2018, 7, e33432. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Dacheux, F. New insights into epididymal function in relation to sperm maturation. Reproduction 2014, 147, R27–R42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Wang, Y.; Lin, H.; Sun, Y.; Wang, M.; Jia, Y.; Yu, X.; Jiang, H.; Xu, W.; Sun, J.; et al. Function and therapeutic potential of G protein-coupled receptors in epididymis. Br. J. Pharmacol. 2020, 177, 5489–5508. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, M.; Joenväärä, S.; Jain, T.; Tomar, A.K.; Sinha, A.; Singh, S.; Yadav, S.; Renkonen, R. Human Spermatozoa Quantitative Proteomic Signature Classifies Normo- and Asthenozoospermia. Mol. Cell. Proteomics 2017, 16, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Adeoya-Osiguwa, S.A.; Fraser, L.R. Cathine, an amphetamine-related compound, acts on mammalian spermatozoa via beta1- and 2A-adrenergic receptors in a capacitation state-dependent manner. Hum. Reprod. 2006, 22, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabeur, K.; Vo, A.; Ball, B. Effects of angiotensin II on the acrosome reaction in equine spermatozoa. Reproduction 2000, 120, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Vedantam, S.; Atreja, S.K.; Garg, M. Angiotensin-II induced nitric oxide production during buffalo sperm capacitation and acrosome reaction. Res. Vet. Sci. 2012, 92, 207–212. [Google Scholar] [CrossRef]
- Mededovic, S.; Fraser, L.R. Angiotensin II stimulates cAMP production and protein tyrosine phosphorylation in mouse spermatozoa. Reproduction 2004, 127, 601–612. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H.; Lax, Y.; Rubinstein, S.; Zamir, N. Angiotensin II induces acrosomal exocytosis in bovine spermatozoa. Am. J. Physiol. Metab. 1998, 275, E87–E93. [Google Scholar] [CrossRef]
- Etkovitz, N.; Tirosh, Y.; Chazan, R.; Jaldety, Y.; Daniel, L.; Rubinstein, S.; Breitbart, H. Bovine sperm acrosome reaction induced by G protein-coupled receptor agonists is mediated by epidermal growth factor receptor transactivation. Dev. Biol. 2009, 334, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, A.A.; Yeste, M.; Salazar, E.; Ramírez-Reveco, A.; Rodriguez Gil, J.E.; Concha, I.I. The Wnt1 ligand/Frizzled 3 receptor system plays a regulatory role in the achievement of the ‘in vitro’ capacitation and subsequent ‘in vitro’ acrosome exocytosis of porcine spermatozoa. Andrology 2015, 3, 357–367. [Google Scholar] [CrossRef]
- Zhang, E.; Xu, F.; Liang, H.; Yan, J.; Xu, H.; Li, Z.; Wen, X.; Weng, J. GLP-1 Receptor Agonist Exenatide Attenuates the Detrimental Effects of Obesity on Inflammatory Profile in Testis and Sperm Quality in Mice. Am. J. Reprod. Immunol. 2015, 74, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.K.B.; Zhang, H.; Yang, J.; Brender, J.R.; Hur, J.; Özgür, A.; Zhang, Y. GLASS: A comprehensive database for experimentally validated GPCR-ligand associations. Bioinformatics 2015, 31, 3035–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinformatics 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Bult, C.J.; Blake, J.A.; Smith, C.L.; Kadin, J.A.; Richardson, J.E.; Anagnostopoulos, A.; Asabor, R.; Baldarelli, R.M.; Beal, J.S.; Bello, S.M.; et al. Mouse Genome Database (MGD) 2019. Nucleic Acids Res. 2019, 47, D801–D806. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; Del-Toro, N.; et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef] [Green Version]
- Assenov, Y.; Ramírez, F.; Schelhorn, S.E.S.E.; Lengauer, T.; Albrecht, M. Computing topological parameters of biological networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef]
Figure 1.
Sperm GPCRs. (a) Venn diagram illustrates the cross-comparison between the human sperm proteome and human GPCRs. For the subsequent analysis, only GPCRs identified in sperm were considered. (b) Classification of sperm GPCRs. (c) Integrative network of relevant processes associated with sperm GPCRs. Each node represents a GPCR (represented by gene name), and each square represents a class of biological processes.
Figure 1.
Sperm GPCRs. (a) Venn diagram illustrates the cross-comparison between the human sperm proteome and human GPCRs. For the subsequent analysis, only GPCRs identified in sperm were considered. (b) Classification of sperm GPCRs. (c) Integrative network of relevant processes associated with sperm GPCRs. Each node represents a GPCR (represented by gene name), and each square represents a class of biological processes.

Figure 2.
Protein-protein interaction network of sperm GPCRs. The green nodes represent sperm GPCRs (n = 47), the blue nodes represent their interactors (n = 163), and the orange nodes represent the interactors associated with relevant biological processes to sperm physiology (n = 49). The bigger green nodes correspond to the top 10 sperm GPCRs with a higher degree. All proteins are represented by the gene name.
Figure 2.
Protein-protein interaction network of sperm GPCRs. The green nodes represent sperm GPCRs (n = 47), the blue nodes represent their interactors (n = 163), and the orange nodes represent the interactors associated with relevant biological processes to sperm physiology (n = 49). The bigger green nodes correspond to the top 10 sperm GPCRs with a higher degree. All proteins are represented by the gene name.

Figure 3.
Integrative network of the relevant interactors and sperm GPCRs. The green nodes represent sperm GPCRs (n = 24), the orange nodes are the interactors associated with relevant biological processes to sperm physiology (n = 49), and the grey squares are the biological processes. All proteins are represented by the gene name.
Figure 3.
Integrative network of the relevant interactors and sperm GPCRs. The green nodes represent sperm GPCRs (n = 24), the orange nodes are the interactors associated with relevant biological processes to sperm physiology (n = 49), and the grey squares are the biological processes. All proteins are represented by the gene name.


{kind=link}
{kind=link}
{kind=link}
Table 1.
G-protein coupled receptors (GPCRs) identified in human spermatozoa. The sperm GPCRs were divided into families, and the subfamily for each receptor is indicated according to the GPCRdb.
Table 1.
G-protein coupled receptors (GPCRs) identified in human spermatozoa. The sperm GPCRs were divided into families, and the subfamily for each receptor is indicated according to the GPCRdb.
| UniProtKB | Gene Name | Protein Names | Subfamily |
|---|---|---|---|
| Class A: Rhodopsin (n = 42) | |||
| Q9BXC0 | HCAR1 | Hydroxycarboxylic acid receptor 1 | Alicarboxylic acid receptors |
| P41595 | HTR2B | 5-hydroxytryptamine receptor 2B | Aminergic receptors |
| P25100 | ADRA1D | Alpha-1D adrenergic receptor | Aminergic receptors |
| Q9NS75 | CYSLTR2 | Cysteinyl leukotriene receptor 2 | Lipid receptors |
| Q9H244 | P2RY12 | P2Y purinoceptor 12 | Nucleotide receptors |
| P51582 | P2RY4 | P2Y purinoceptor 4 | Nucleotide receptors |
| Q9NQS5 | GPR84 | G-protein coupled receptor 84 | Orphan receptors |
| Q96P66 | GPR101 | Probable G-protein coupled receptor 101 | Orphan receptors |
| Q5T848 | GPR158 | Probable G-protein coupled receptor 158 | Orphan receptors |
| O15354 | GPR37 | Prosaposin receptor GPR37 | Orphan receptors |
| Q9P1P5 | TAAR2 | Trace amine-associated receptor 2 | Orphan receptors |
| P24530 | EDNRB | Endothelin receptor type B | Peptide receptors |
| P25101 | EDNRA | Endothelin-1 receptor | Peptide receptors |
| P32239 | CCKBR | Gastrin/cholecystokinin type B receptor | Peptide receptors |
| Q01726 | MC1R | Melanocyte-stimulating hormone receptor | Peptide receptors |
| Q9GZQ6 | NPFFR1 | Neuropeptide FF receptor 1 | Peptide receptors |
| Q9Y5X5 | NPFFR2 | Neuropeptide FF receptor 2 | Peptide receptors |
| P25090 | FPR2 | N-formyl peptide receptor 2 | Peptide receptors |
| P25116 | F2R | Proteinase-activated receptor 1 | Peptide receptors |
| P30874 | SSTR2 | Somatostatin receptor type 2 | Peptide receptors |
| P30556 | AGTR1 | Type-1 angiotensin II receptor | Peptide receptors |
| P50052 | AGTR2 | Type-2 angiotensin II receptor | Peptide receptors |
| Q9GZK3 | OR2B2 | Olfactory receptor 2B2 | Peptide receptors |
| Q8NGT1 | OR2K2 | Olfactory receptor 2K2 | Peptide receptors |
| Q8NH16 | OR2L2 | Olfactory receptor 2L2 | Peptide receptors |
| Q8NG97 | OR2Z1 | Olfactory receptor 2Z1 | Peptide receptors |
| Q8NGL6 | OR4A15 | Olfactory receptor 4A15 | Peptide receptors |
| Q8NGP0 | OR4C13 | Olfactory receptor 4C13 | Peptide receptors |
| Q8NH41 | OR4K15 | Olfactory receptor 4K15 | Peptide receptors |
| Q8NGK0 | OR51G2 | Olfactory receptor 51G2 | Peptide receptors |
| Q8NGH9 | OR52E4 | Olfactory receptor 52E4 | Peptide receptors |
| Q8NGH8 | OR56A4 | Olfactory receptor 56A4 | Peptide receptors |
| Q8NHB8 | OR5K2 | Olfactory receptor 5K2 | Peptide receptors |
| A6NET4 | OR5K3 | Olfactory receptor 5K3 | Peptide receptors |
| A6NMS3 | OR5K4 | Olfactory receptor 5K4 | Peptide receptors |
| Q8NGP4 | OR5M3 | Olfactory receptor 5M3 | Peptide receptors |
| Q8NGP3 | OR5M9 | Olfactory receptor 5M9 | Peptide receptors |
| Q8NGE1 | OR6C4 | Olfactory receptor 6C4 | Peptide receptors |
| Q8NG99 | OR7G2 | Olfactory receptor 7G2 | Peptide receptors |
| Q9GZM6 | OR8D2 | Olfactory receptor 8D2 | Peptide receptors |
| Q8NG78 | OR8G5 | Olfactory receptor 8G5 | Peptide receptors |
| Q8NGP2 | OR8J1 | Olfactory receptor 8J1 | Peptide receptors |
| Class B1: Secretin (n = 2) | |||
| P43220 | GLP1R | Glucagon-like peptide 1 receptor | Peptide receptors |
| P41587 | VIPR2 | Vasoactive intestinal polypeptide receptor 2 | Peptide receptors |
| Class B2: Adhesion (n = 16) | |||
| Q96PE1 | ADGRA2 | Adhesion G protein-coupled receptor A2 | Adhesion receptors |
| O60241 | ADGRB2 | Adhesion G protein-coupled receptor B2 | Adhesion receptors |
| Q14246 | ADGRE1 | Adhesion G protein-coupled receptor E1 | Adhesion receptors |
| P48960 | ADGRE5 | Adhesion G protein-coupled receptor E5 | Adhesion receptors |
| Q8IZF2 | ADGRF5 | Adhesion G protein-coupled receptor F5 | Adhesion receptors |
| Q86Y34 | ADGRG3 | Adhesion G protein-coupled receptor G3 | Adhesion receptors |
| O95490 | ADGRL2 | Adhesion G protein-coupled receptor L2 | Adhesion receptors |
| Q9Y653 | ADGRG1 | Adhesion G-protein coupled receptor G1 | Adhesion receptors |
| Q8IZP9 | ADGRG2 | Adhesion G-protein coupled receptor G2 | Adhesion receptors |
| Q8IZF6 | ADGRG4 | Adhesion G-protein coupled receptor G4 | Adhesion receptors |
| Q86SQ4 | ADGRG6 | Adhesion G-protein coupled receptor G6 | Adhesion receptors |
| Q8WXG9 | ADGRV1 | Adhesion G-protein coupled receptor V1 | Adhesion receptors |
| P08913 | ADRA2A | Alpha-2A adrenergic receptor | Aminergic receptors |
| Q9NYQ6 | CELSR1 | Cadherin EGF LAG seven-pass G-type receptor 1 | Adhesion receptors |
| Q9HCU4 | CELSR2 | Cadherin EGF LAG seven-pass G-type receptor 2 | Adhesion receptors |
| Q9NYQ7 | CELSR3 | Cadherin EGF LAG seven-pass G-type receptor 3 | Adhesion receptors |
| Class C: Glutamate (n = 10) | |||
| Q9UBS5 | GABBR1 | Gamma-aminobutyric acid type B receptor subunit 1 | Amino acid receptors |
| Q13255 | GRM1 | Metabotropic glutamate receptor 1 | Amino acid receptors |
| Q14416 | GRM2 | Metabotropic glutamate receptor 2 | Amino acid receptors |
| Q14832 | GRM3 | Metabotropic glutamate receptor 3 | Amino acid receptors |
| Q14833 | GRM4 | Metabotropic glutamate receptor 4 | Amino acid receptors |
| O15303 | GRM6 | Metabotropic glutamate receptor 6 | Amino acid receptors |
| O00222 | GRM8 | Metabotropic glutamate receptor 8 | Amino acid receptors |
| Q8TE23 | TAS1R2 | Taste receptor type 1 member 2 | Sensory receptors |
| P59544 | TAS2R50 | Taste receptor type 2 member 50 | Sensory receptors |
| Q9NYW3 | TAS2R7 | Taste receptor type 2 member 7 | Sensory receptors |
| Class F: Frizzled (n = 1) | |||
| Q9NPG1 | FZD3 | Frizzled-3 | Protein receptors |
Table 2.
GPCRs with selective expression in male reproductive tissues or associated with male infertility defects. For each GPCR, selective expression in male reproductive tissues and the male infertility phenotypes observed in knock-out mice are indicated.
Table 2.
GPCRs with selective expression in male reproductive tissues or associated with male infertility defects. For each GPCR, selective expression in male reproductive tissues and the male infertility phenotypes observed in knock-out mice are indicated.
| Gene Name | Protein Name | Tissue Expression | Male Fertility Phenotype |
|---|---|---|---|
| ADGRG1 | Adhesion G-protein coupled receptor G1 | Other tissues | Abnormal testis morphology; reduced male infertility |
| ADGRG2 | Adhesion G-protein coupled receptor G2 | Epididymis (enriched) | Abnormal vas deferens; azoospermia |
| ADGRV1 | Adhesion G-protein coupled receptor V1 | Other tissues | Abnormal testis morphology |
| ADRA2A | Alpha-2A adrenergic receptor | Other tissues | Abnormal testis morphology |
| CELSR1 | Cadherin EGF LAG seven-pass G-type receptor 1 | Seminal vesicle (enriched) | Abnormal testis morphology; reduced fertility |
| CYSLTR2 | Cysteinyl leukotriene receptor 2 | Seminal vesicle (enhanced) | N/A |
| EDNRA | Endothelin-1 receptor | Seminal vesicle (enhanced) | N/A |
| F2R | Proteinase-activated receptor 1 | Other tissues | Abnormal fertility/fecundity |
| FZD3 | Frizzled-3 | Seminal vesicle (enhanced) | N/A |
| GRM1 | Metabotropic glutamate receptor 1 | Other tissues | Abnormal fertility/fecundity; male infertility; reduced fertility |
| GRM2 | Metabotropic glutamate receptor 2 | Testis (enhanced) | N/A |
| HTR2B | 5-hydroxytryptamine receptor 2B | Other tissues | Reduced fertility |
| NPFFR2 | Neuropeptide FF receptor 2 | Seminal vesicle (enhanced) | N/A |
| VIPR2 | Vasoactive intestinal polypeptide receptor 2 | Other tissues | Abnormal testis morphology; male infertility |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Corda, P.O.; Santiago, J.; Fardilha, M. G-Protein Coupled Receptors in Human Sperm: An In Silico Approach to Identify Potential Modulatory Targets. Molecules 2022, 27, 6503. https://doi.org/10.3390/molecules27196503
AMA Style
Corda PO, Santiago J, Fardilha M. G-Protein Coupled Receptors in Human Sperm: An In Silico Approach to Identify Potential Modulatory Targets. Molecules. 2022; 27(19):6503. https://doi.org/10.3390/molecules27196503
Chicago/Turabian StyleCorda, Pedro O., Joana Santiago, and Margarida Fardilha. 2022. "G-Protein Coupled Receptors in Human Sperm: An In Silico Approach to Identify Potential Modulatory Targets" Molecules 27, no. 19: 6503. https://doi.org/10.3390/molecules27196503