
Natural Small Molecules in Gastrointestinal Tract and Associated Cancers: Molecular Insights and Targeted Therapies

 , ,
, ,  ,
, 
 , ,
, ,  and
and
Abstract
:1. Introduction
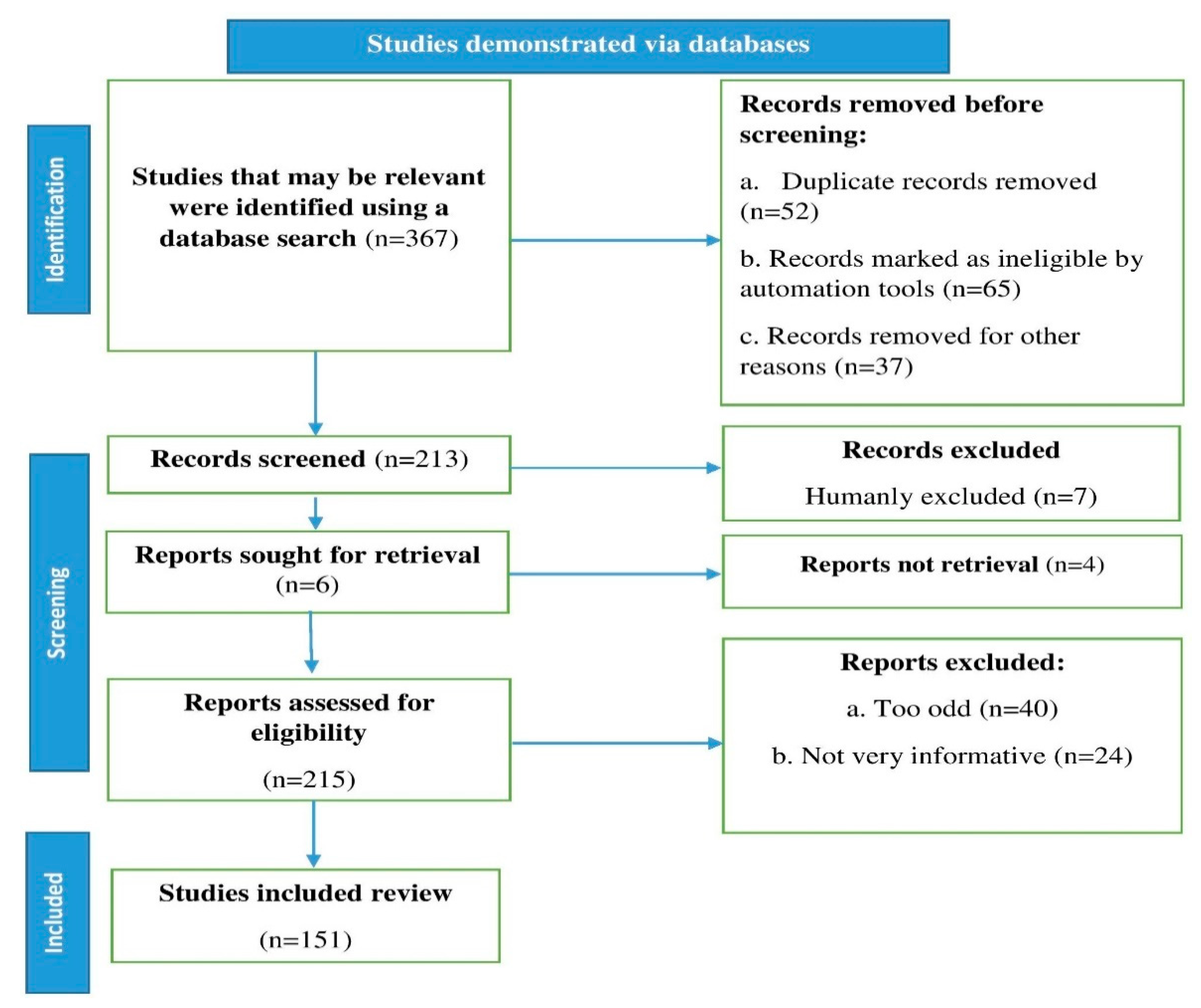
2. Methodology
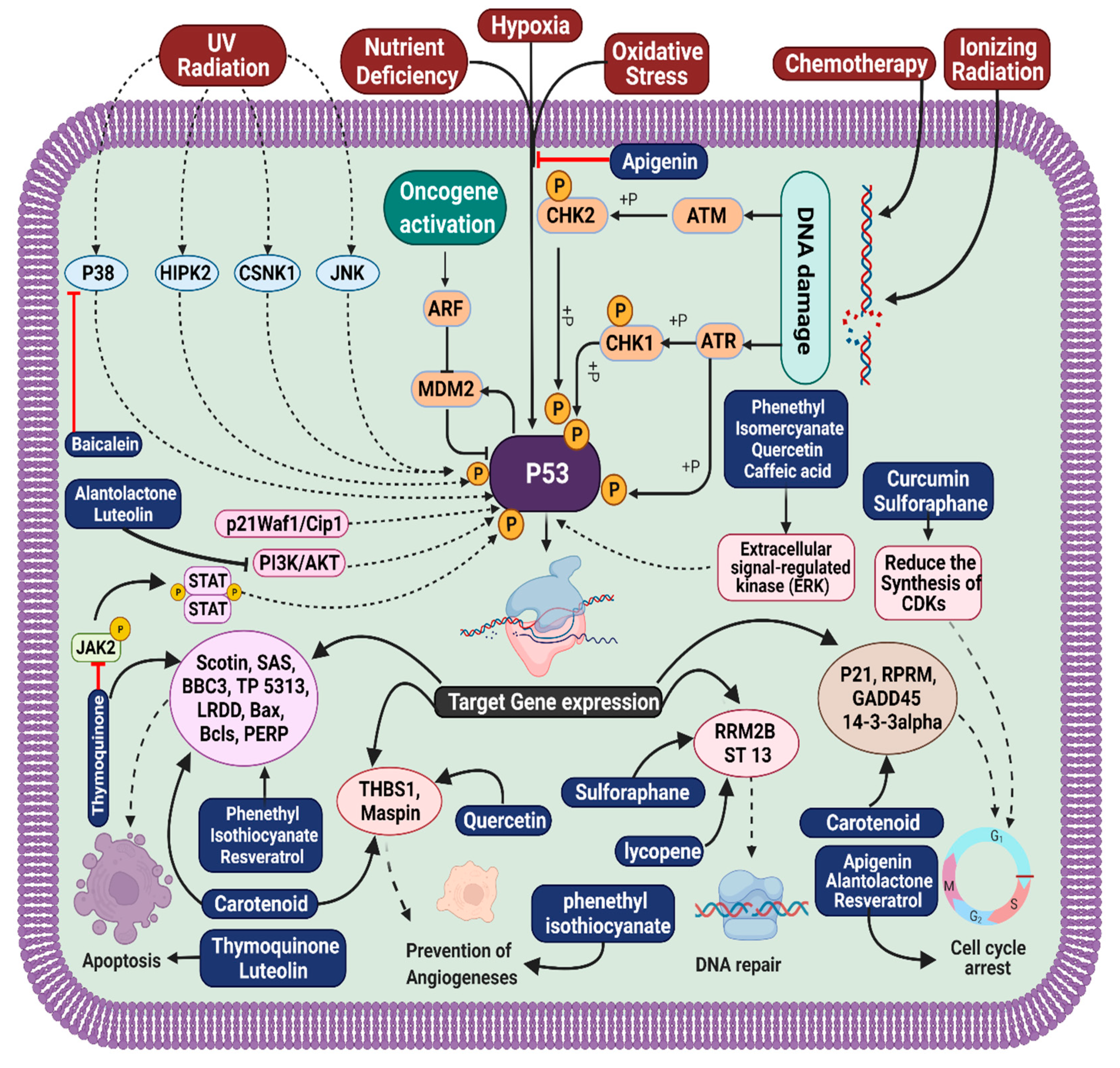
3. Molecular Pathogenesis of Gastrointestinal Cancer
4. Natural Small Molecules in Gastrointestinal Cancer
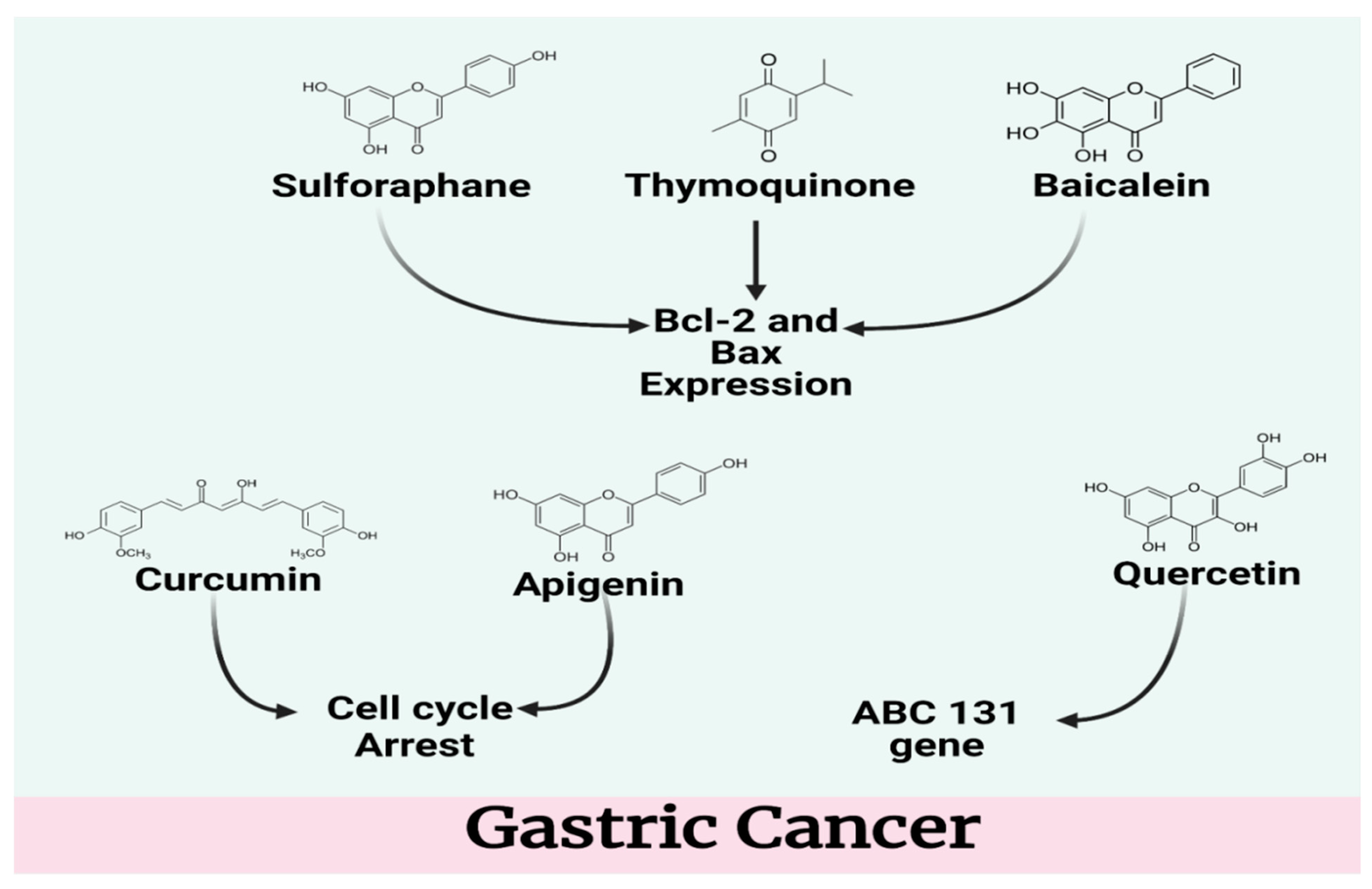
4.1. Gastric Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Compound | Study Model | Dose | Results | Mechanism of Action | References |
|---|---|---|---|---|---|
| Resveratrol | In vitro (HP Strains) | 6.25 µg to 100 µg | MIC50 and MIC90 were 12.5 and 25 µg | Inhibited the replication of H. pylori | [47] |
| In vitro (gastric adenocarcinoma SNU-1 cells) | 10 and 100 μM | Apoptosis | Decreased protein kinase C activity, promoted cell cycle arrest, and reduced gastric cancer cell growth induced by nitrosamine. | [48] | |
| Quercetin | In vitro (Parental EPG85-257P cell line) | 12 μM | Induced apoptosis | Downregulation of ABCB1 gene | [49] |
| In vitro (AGS cells) | 0, 10, 20, 40, 80, 160, and 320 μM | Induced apoptosis | Quercetin promoted morphological alterations and lowered vitality | [50] | |
| Apigenin | In vitro (HGC-27 and SGC-7901 cells) | 10 μg/mL | Induced Apoptosis | Suppressed the growth of stomach cancer | [51] |
| In vivo (Mongolian gerbils) | 30–60 mg/kg/day | Mongolian gerbils with atrophic gastritis and dysplasia/gastric cancer had their cancer rates considerably reduced | [32] | ||
| isothiocyanates (ITCs) | In vitro (AGS cell line) | 0.25 or 0.50 μM | Antiproliferative | Decreased AGS cell invasion and migration | [33] |
| In vitro (Human gastric cell lines MKN45, AGS, MKN74 and KATO-III) | 3–5 µmol PEITC/g | Chemopreventive effect | - | [52] | |
| Sulforaphane | In vitro (BGC-823 and MGC-803 cell lines) | 5 and 10 μM | Apoptosis | Cell cycle arrest and apoptosis induction in GC cells | [34] |
| In vitro (Gastric cancer cell lines, BGC-823 and SGC-7901) | 0 to 10 μM | Apoptosis | Both BGC-823 and SGC-7901 tumor spheres showed a reduction in the proliferation-related proteins PCNA and Cyclin D1. Bcl-2 expression was reduced in tumor spheres, whereas the expression of the pro-apoptosis proteins Bax and Caspase 8 was elevated | [53] | |
| Alantolactone (ALT) | In vitro (BGC-823 cells) | 0, 10, 20, 40 or 60 µM | Apoptosis | Via reduction of AKT signaling, ROS formation was successfully suppressed by the ROS scavenger N-acetyl cysteine, leading to the induction of apoptosis through the action of ALT. | [54] |
| In vitro (BGC-823 and SGC-7901 cells) | 10 and 20 mM | Anti-proliferation and apoptosis | Gastric cancer cells were killed by alantolactone, perhaps through control of the expression of MMPs. | [35] | |
| Baicalein | In vitro (gastric cancer cell line SGC-7901) | 0, 15, 30, and 60 μmol/L | Apoptosis | Revealed Bcl-2 downregulation and Bax overexpression after treatment with baicalein. According to these findings, baicalein causes apoptosis in gastric cancer cells through the mitochondrial mechanism. | [55] |
| Thymoquinone(TQ) | In vitro (MGC80-3 and SGC-7901) | 0–10, 25 µM | Apoptosis | To promote apoptosis, TQ modulates the expression of pro- and anti-apoptotic markers in gastric cancer cell lines. Bacterial Bax and caspase-3 were highly up-regulated whereas Bcl-2 was dramatically decreased. | [56] |
| Lycopene | In vitro (gastric adenocarcinoma; ATCC CRL 1739) | 0.5, 1, or 2 μM | Apoptosis | Increased apoptotic indices (DNA fragmentation, AIF, caspase-3 and caspase-9 cleavage, Bax/Bcl-2 ratio) were seen in the presence of lycopene. | [38] |
| In vitro (The gastric epithelial cell line AGS) | 25 μg/mL | Anti-proliferative | - | [57] | |
| Alliin and allicin | In vitro (The human gastric adenocarcinoma cell line SGC7901) | 15–120 μg/mL | Anti-proliferative | - | [40] |
| In vitro (SGC-7901 cells) | 30 μg/mL | Apoptosis | - | [40] |
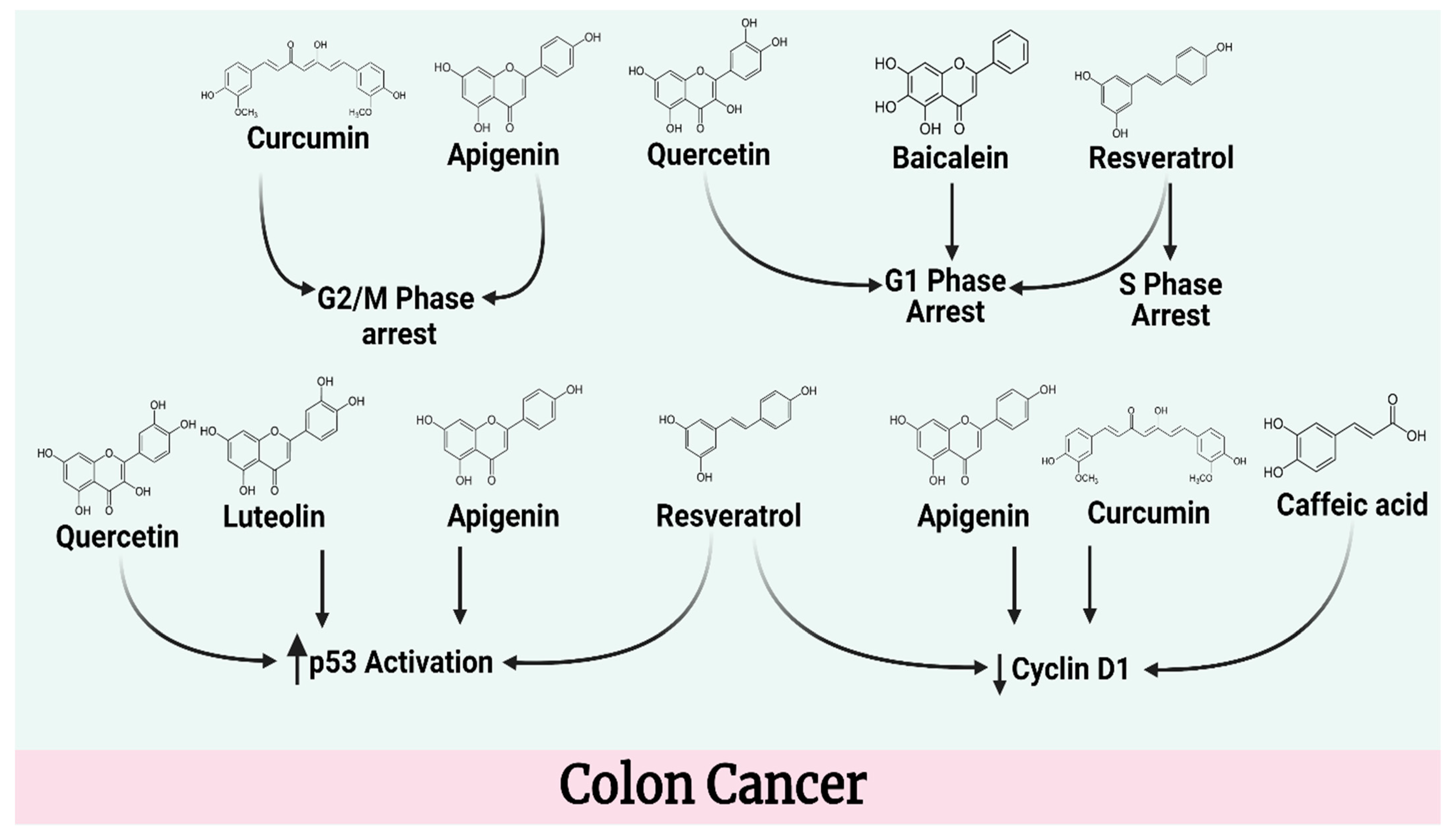
4.2. Colon Cancer
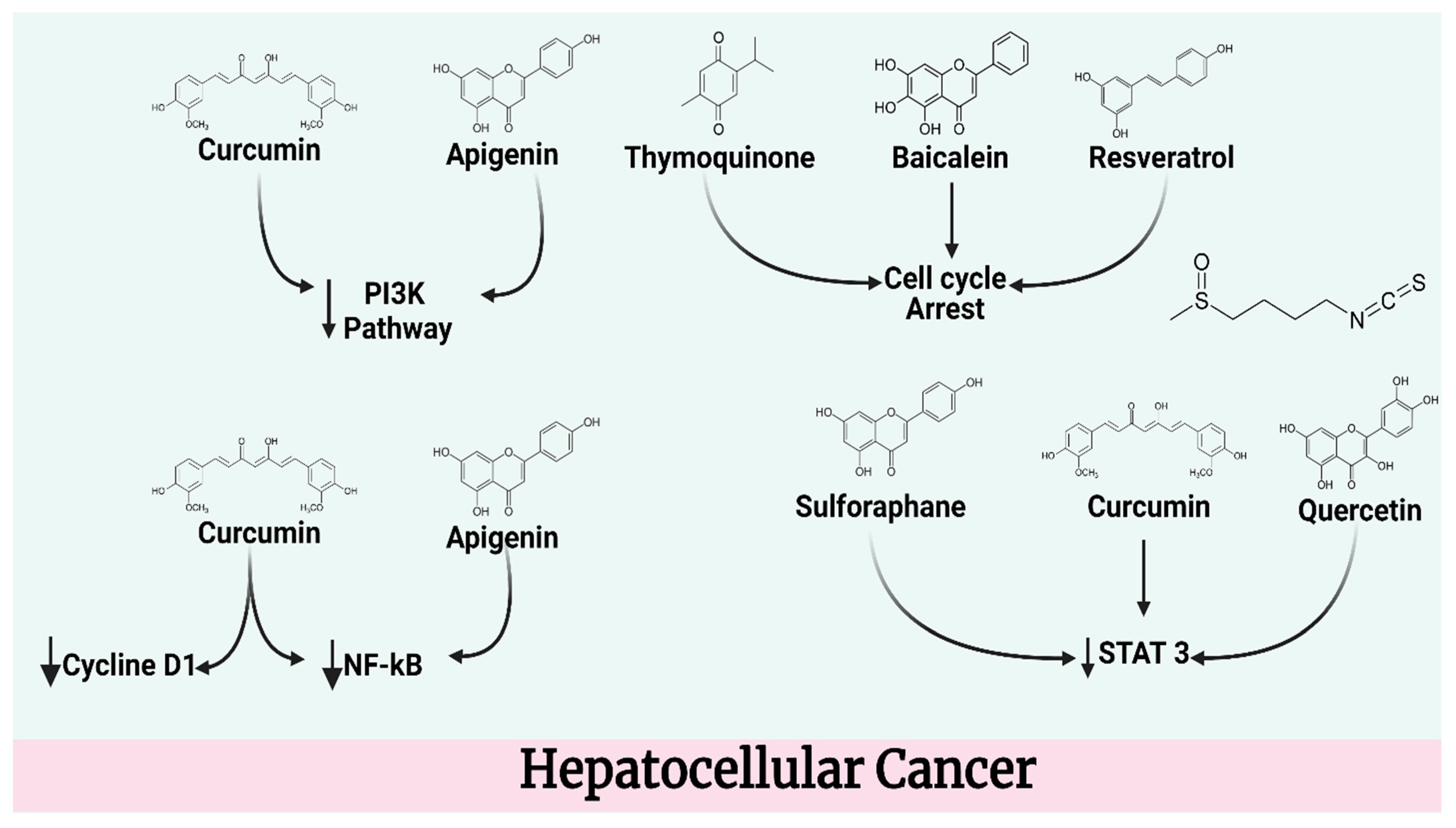
4.3. Hepatocellular Cancer
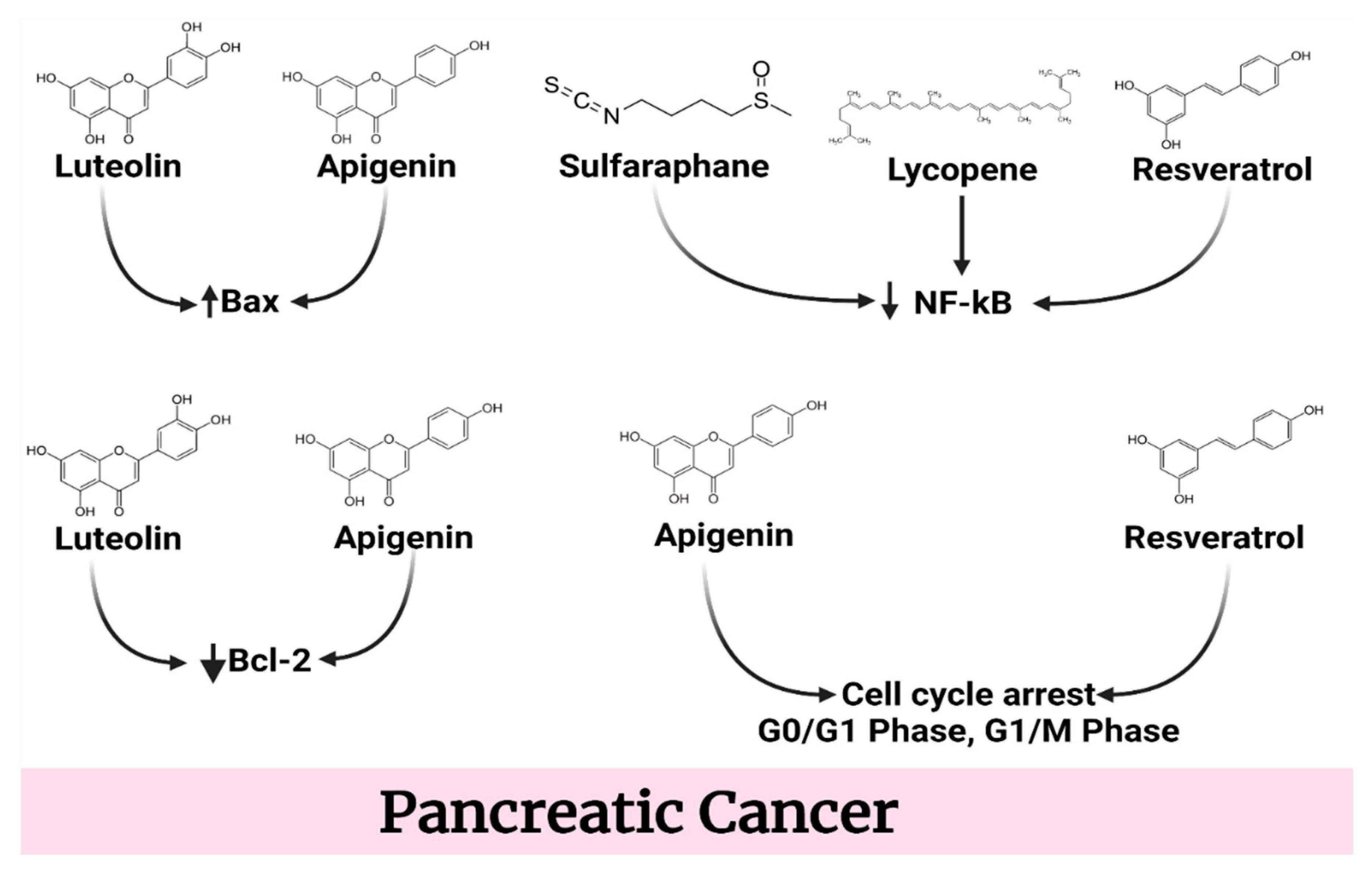
4.4. Pancreatic Cancer
5. Synergistic Effects
6. Clinical Evidences
7. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parkin, D.; Whelan, S.; Ferlay, J.; Teppo, L.; Thomas, D. Cancer Incidence in Five Continents Volume VIII; IARC Scientific Publication: Lyon, France, 2002. [Google Scholar]
- Parkin, D.M. Cancer Incidence in Five Continents; World Health Organization: Lyon, France, 1997. [Google Scholar]
- Percy, C.; Young, J., Jr.; Muir, C.; Ries, L.; Hankey, B.; Sobin, L.; Berg, J. Cancer. Introduction. Cancer 1995, 75, 140–146. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for preventing gastrointestinal cancers. Cochrane Database Syst. Rev. 2008, 16, Cd004183. [Google Scholar] [CrossRef]
- Yamada, T.A.D.H. Textbook of Gastroenterology, 5th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Katona, B.W.; Lynch, J.P. Chapter 66—Mechanisms of Gastrointestinal Malignancies. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H.M., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1615–1642. [Google Scholar]
- Link, A.; Goel, A. Chapter Six—MicroRNA in Gastrointestinal Cancer: A Step Closer to Reality. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 62, pp. 221–268. [Google Scholar]
- Govardhanagiri, S.; Bethi, S.; Nagaraju, G.P. Chapter 8—Small Molecules and Pancreatic Cancer Trials and Troubles. In Breaking Tolerance to Pancreatic Cancer Unresponsiveness to Chemotherapy; Nagaraju, G.P., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 5, pp. 117–131. [Google Scholar]
- Jeswani, G.; Paul, S.D. Chapter 15—Recent Advances in the Delivery of Chemotherapeutic Agents. In Nano- and Microscale Drug Delivery Systems; Grumezescu, A.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 281–298. [Google Scholar]
- Wynder, E.L.; Bross, I.J. A study of etiological factors in cancer of the esophagus. Cancer 1961, 14, 389–413. [Google Scholar] [CrossRef]
- Lynch, J.P.; Rustgi, A.K. Chapter 80—Mechanisms of GI Malignancies. In Physiology of the Gastrointestinal Tract, 5th ed.; Johnson, L.R., Ghishan, F.K., Kaunitz, J.D., Merchant, J.L., Said, H.M., Wood, J.D., Eds.; Academic Press: Boston, MA, USA, 2012; pp. 2129–2155. [Google Scholar]
- Heavey, P.M.; Rowland, I.R. Gastrointestinal cancer. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 323–336. [Google Scholar] [CrossRef]
- Chandradas, S.; Skillin, C.B.; Frenette, C.T. Chapter 9—Hepatocellular Carcinoma: Metastatic Disease. In Introduction to Cancer Metastasis; Ahmad, A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 163–175. [Google Scholar]
- Alharthy, S.A.; Tabrez, S.; Mirza, A.A.; Zughaibi, T.A.; Firoz, C.K.; Dutta, M. Sugiol Suppresses the Proliferation of Human U87 Glioma Cells via Induction of Apoptosis and Cell Cycle Arrest. Evid. -Based Complementary Altern. Med. 2022, 2022, 7658899. [Google Scholar] [CrossRef]
- Nakamuva, K.; Ueyarna, T.; Yao, T.; Xuan, Z.X.; Ambe, K.; Adachi, Y.; Yakeishi, Y.; Matsukuma, A.; Enjoji, M. Pathology and prognosis of gastric carcinoma. Findings in 10,000 patients who underwent primary gastrectomy. Cancer 1992, 70, 1030–1037. [Google Scholar] [CrossRef]
- Tang, G.X.; Wang, Z.S.; Liu, D.G.; Liu, S.S. Gastroesophageal anastomotic leakage following resection of carcinoma of the esophagus and gastric cardia: Analysis of ten cases. J. Surg. Oncol. 1990, 43, 50–52. [Google Scholar] [CrossRef]
- Liccioni, A.; Reig, M.E.; Bruix, J. Chapter 91—Hepatocellular carcinoma. In Blumgart’s Surgery of the Liver, Biliary Tract and Pancreas, 2-Volume Set, 6th ed.; Jarnagin, W.R., Ed.; Elsevier: Philadelphia, PA, USA, 2017; pp. 1333–1338.e2. [Google Scholar]
- Joo, M.K.; Park, J.-J.; Chun, H.J. Impact of homeobox genes in gastrointestinal cancer. World J. Gastroenterol. 2016, 22, 8247. [Google Scholar] [CrossRef]
- Harmon, R.L.; Sugarbaker, P.H. Prognostic Indicators in Peritoneal Carcinomatosis from Gastrointestinal Cancer. In International Seminars in Surgical Oncology; BioMed Central: London, UK, 2005; pp. 1–10. [Google Scholar]
- Mitra, S.; Lami, M.S.; Ghosh, A.; Das, R.; Tallei, T.E.; Islam, F.; Dhama, K.; Begum, M.Y.; Aldahish, A.; Chidambaram, K. Hormonal therapy for gynecological cancers: How far has science progressed toward clinical applications? Cancers 2022, 14, 759. [Google Scholar] [CrossRef]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Díaz Marrero, A.R.; Deniz, I.; Erdoğan, A.; Lukić Bilela, L.; Moulin, C.; Taffin-de-Givenchy, E. Marine anticancer agents: An overview with a particular focus on their chemical classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Tabrez, S.; Zughaibi, T.A.; Hoque, M.; Suhail, M.; Khan, M.I.; Khan, A.U. Targeting glutaminase by natural compounds: Structure-based virtual screening and molecular dynamics simulation approach to suppress cancer progression. Molecules 2022, 27, 5042. [Google Scholar]
- Ding, Y.; Tong, Z.; Jin, L.; Ye, B.; Zhou, J.; Sun, Z.; Yang, H.; Hong, L.; Huang, F.; Wang, W. An NIR Discrete Metallacycle Constructed from Perylene Bisimide and Tetraphenylethylene Fluorophores for Imaging-Guided Cancer Radio-Chemotherapy. Adv. Mater. 2022, 34, 2106388. [Google Scholar] [CrossRef]
- Zhou, J.; Rao, L.; Yu, G.; Cook, T.R.; Chen, X.; Huang, F. Supramolecular cancer nanotheranostics. Chem. Soc. Rev. 2021, 50, 2839–2891. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, G.; Huang, F. Supramolecular chemotherapy based on host–guest molecular recognition: A novel strategy in the battle against cancer with a bright future. Chem. Soc. Rev. 2017, 46, 7021–7053. [Google Scholar] [CrossRef]
- Yamagishi, H.; Kuroda, H.; Imai, Y.; Hiraishi, H. Molecular pathogenesis of sporadic colorectal cancers. Chin. J. Cancer 2016, 35, 1–8. [Google Scholar] [CrossRef]
- Hansel, D.E.; Kern, S.E.; Hruban, R.H. Molecular pathogenesis of pancreatic cancer. Annu. Rev. Genom. Hum. Genet. 2003, 4, 237–256. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.; Yeoh, K.-G. Genetics and molecular pathogenesis of gastric adenocarcinoma. Gastroenterology 2015, 149, 1153–1162.e3. [Google Scholar] [CrossRef]
- Athar, M.; Back, J.H.; Tang, X.; Kim, K.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Resveratrol: A review of preclinical studies for human cancer prevention. Toxicol. Appl. Pharmacol. 2007, 224, 274–283. [Google Scholar] [CrossRef]
- Lee, H.H.; Lee, S.; Shin, Y.S.; Cho, M.; Kang, H.; Cho, H. Anti-cancer effect of quercetin in xenograft models with EBV-associated human gastric carcinoma. Molecules 2016, 21, 1286. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Weng, B.-C.; Wu, C.-C.; Yang, S.-F.; Wu, D.-C.; Wang, Y.-C. Apigenin has anti-atrophic gastritis and anti-gastric cancer progression effects in Helicobacter pylori-infected Mongolian gerbils. J. Ethnopharmacol. 2014, 151, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-D.; Lai, K.-C.; Lai, T.-Y.; Hsu, S.-C.; Kuo, C.-L.; Yu, C.-S.; Lin, M.-L.; Yang, J.-S.; Kuo, H.-M.; Wu, S.-H. Phenethyl isothiocyanate inhibits migration and invasion of human gastric cancer AGS cells through suppressing MAPK and NF-κB signal pathways. Anticancer Res. 2010, 30, 2135–2143. [Google Scholar] [PubMed]
- Wang, Y.; Wu, H.; Dong, N.; Su, X.; Duan, M.; Wei, Y.; Wei, J.; Liu, G.; Peng, Q.; Zhao, Y. Sulforaphane induces S-phase arrest and apoptosis via p53-dependent manner in gastric cancer cells. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cao, X.; Kong, Y.; Wang, S.; Xia, Y.; Bi, R.; Liu, J. Apoptosis-promoting and migration-suppressing effect of alantolactone on gastric cancer cell lines BGC-823 and SGC-7901 via regulating p38MAPK and NF-κB pathways. Hum. Exp. Toxicol. 2019, 38, 1132–1144. [Google Scholar] [CrossRef]
- Yan, X.; Rui, X.; Zhang, K. Baicalein inhibits the invasion of gastric cancer cells by suppressing the activity of the p38 signaling pathway. Oncol. Rep. 2015, 33, 737–743. [Google Scholar] [CrossRef]
- Zhu, W.-Q.; Wang, J.; Guo, X.-F.; Liu, Z.; Dong, W.-G. Thymoquinone inhibits proliferation in gastric cancer via the STAT3 pathway in vivo and in vitro. World J. Gastroenterol. 2016, 22, 4149. [Google Scholar] [CrossRef]
- Han, H.; Lim, J.W.; Kim, H. Lycopene inhibits activation of epidermal growth factor receptor and expression of cyclooxygenase-2 in gastric cancer cells. Nutrients 2019, 11, 2113. [Google Scholar] [CrossRef]
- Wu, C.-S.; Chen, M.-F.; Lee, I.-L.; Tung, S.-Y. Predictive role of nuclear factor-κB activity in gastric cancer: A promising adjuvant approach with caffeic acid phenethyl ester. J. Clin. Gastroenterol. 2007, 41, 894–900. [Google Scholar] [CrossRef]
- Zhang, W.; Ha, M.; Gong, Y.; Xu, Y.; Dong, N.; Yuan, Y. Allicin induces apoptosis in gastric cancer cells through activation of both extrinsic and intrinsic pathways. Oncol. Rep. 2010, 24, 1585–1592. [Google Scholar]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Selvendiran, K.; Koga, H.; Ueno, T.; Yoshida, T.; Maeyama, M.; Torimura, T.; Yano, H.; Kojiro, M.; Sata, M. Luteolin promotes degradation in signal transducer and activator of transcription 3 in human hepatoma cells: An implication for the antitumor potential of flavonoids. Cancer Res. 2006, 66, 4826–4834. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Gadad, S.S.; Kim, D.-S.; Kraus, W.L. Discovery, annotation, and functional analysis of long noncoding RNAs controlling cell-cycle gene expression and proliferation in breast cancer cells. Mol. Cell 2015, 59, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, M.; Ray, S.K. Anti-tumor activities of luteolin and silibinin in glioblastoma cells: Overexpression of miR-7-1-3p augmented luteolin and silibinin to inhibit autophagy and induce apoptosis in glioblastoma in vivo. Apoptosis 2016, 21, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Jain, H.; Dhingra, N.; Narsinghani, T.; Sharma, R. Insights into the mechanism of natural terpenoids as NF-κB inhibitors: An overview on their anticancer potential. Exp. Oncol. 2016, 38, 158–168. [Google Scholar] [CrossRef]
- Onoda, C.; Kuribayashi, K.; Nirasawa, S.; Tsuji, N.; Tanaka, M.; Kobayashi, D.; Watanabe, N. (−)-Epigallocatechin-3-gallate induces apoptosis in gastric cancer cell lines by down-regulating survivin expression. Int. J. Oncol. 2011, 38, 1403–1408. [Google Scholar]
- Mahady, G.B.; Pendland, S.L. Resveratrol inhibits the growth of Helicobacter pylori in vitro. Am. J. Gastroenterol. 2000, 95, 1849. [Google Scholar] [CrossRef]
- Palit, S.; Bhuiyan, R.H.; Aklima, J.; Emran, T.B.; Dash, R. A study of the prevalence of thalassemia and its correlation with liver function test in different age and sex group in the Chittagong district of Bangladesh. J. Basic Clin. Pharm. 2012, 3, 352. [Google Scholar]
- Borska, S.; Chmielewska, M.; Wysocka, T.; Drag-Zalesinska, M.; Zabel, M.; Dziegiel, P. In vitro effect of quercetin on human gastric carcinoma: Targeting cancer cells death and MDR. Food Chem. Toxicol. 2012, 50, 3375–3383. [Google Scholar] [CrossRef]
- Dash, R.; Uddin, M.M.N.; Islam, A.; Junaid, M. Molecular docking of fisetin with AD associated AChE, ABAD and BACE1 proteins. Bioinformation 2014, 10, 562. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.; Li, Z.; Liu, C.; Yin, L. The apoptotic effect of apigenin on human gastric carcinoma cells through mitochondrial signal pathway. Tumor Biol. 2014, 35, 7719–7726. [Google Scholar] [CrossRef]
- Rabben, H.-L.; Kodama, Y.; Nakamura, M.; Bones, A.M.; Wang, T.C.; Chen, D.; Zhao, C.-M.; Øverby, A. Chemopreventive effects of dietary isothiocyanates in animal models of gastric cancer and synergistic anticancer effects with cisplatin in human gastric cancer cells. Front. Pharmacol. 2021, 12, 613458. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Zhang, L.; Cao, L.; Xie, C.; Li, X.; Li, Y.; Meng, Y.; Chen, Y.; Wang, X.; Chen, J. Sulforaphane inhibits gastric cancer stem cells via suppressing sonic hedgehog pathway. Int. J. Food Sci. Nutr. 2019, 70, 570–578. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.M. Alantolactone induces gastric cancer BGC-823 cell apoptosis by regulating reactive oxygen species generation and the AKT signaling pathway. Oncol. Lett. 2019, 17, 4795–4802. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Liu, T.; Jiang, L.; Wu, X.; Cao, Y.; Li, M.; Dong, Q.; Liu, Y.; Xu, H. The traditional Chinese medicine baicalein potently inhibits gastric cancer cells. J. Cancer 2016, 7, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.-M.; Wang, X.-F.; Huang, Q.-X. Thymoquinone induces cytotoxicity and reprogramming of EMT in gastric cancer cells by targeting PI3K/Akt/mTOR pathway. J. Biosci. 2017, 42, 547–554. [Google Scholar] [CrossRef]
- Abdel-Latif, M.M.; Windle, H.J.; Homasany, B.S.E.; Sabra, K.; Kelleher, D. Caffeic acid phenethyl ester modulates Helicobacter pylori-induced nuclear factor-kappa B and activator protein-1 expression in gastric epithelial cells. Br. J. Pharmacol. 2005, 146, 1139–1147. [Google Scholar] [CrossRef]
- Favoriti, P.; Carbone, G.; Greco, M.; Pirozzi, F.; Pirozzi, R.E.M.; Corcione, F. Worldwide burden of colorectal cancer: A review. Updates Surg. 2016, 68, 7–11. [Google Scholar] [CrossRef]
- Schneider, Y.; Vincent, F.; Duranton, B.; Badolo, L.; Gossé, F.; Bergmann, C.; Seiler, N.; Raul, F. Anti-proliferative effect of resveratrol, a natural component of grapes and wine, on human colonic cancer cells. Cancer Lett. 2000, 158, 85–91. [Google Scholar] [CrossRef]
- Juneja, M.; Kobelt, D.; Walther, W.; Voss, C.; Smith, J.; Specker, E.; Neuenschwander, M.; Gohlke, B.-O.; Dahlmann, M.; Radetzki, S. Statin and rottlerin small-molecule inhibitors restrict colon cancer progression and metastasis via MACC1. PLoS Biol. 2017, 15, e2000784. [Google Scholar] [CrossRef]
- Delmas, D.; Rébé, C.; Lacour, S.; Filomenko, R.; Athias, A.; Gambert, P.; Cherkaoui-Malki, M.; Jannin, B.; Dubrez-Daloz, L.; Latruffe, N. Resveratrol-induced apoptosis is associated with Fas redistribution in the rafts and the formation of a death-inducing signaling complex in colon cancer cells. J. Biol. Chem. 2003, 278, 41482–41490. [Google Scholar] [CrossRef]
- Wolter, F.; Akoglu, B.; Clausnitzer, A.; Stein, J. Downregulation of the cyclin D1/Cdk4 complex occurs during resveratrol-induced cell cycle arrest in colon cancer cell lines. J. Nutr. 2001, 131, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- del Mar Blanquer-Rosselló, M.; Hernández-López, R.; Roca, P.; Oliver, J.; Valle, A. Resveratrol induces mitochondrial respiration and apoptosis in SW620 colon cancer cells. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2017, 1861, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Fuggetta, M.; Lanzilli, G.; Tricarico, M.; Cottarelli, A.; Falchetti, R.; Ravagnan, G.; Bonmassar, E. Effect of resveratrol on proliferation and telomerase activity of human colon cancer cells in vitro. J. Exp. Clin. Cancer Res. 2006, 25, 189. [Google Scholar] [PubMed]
- Aires, V.; Limagne, E.; Cotte, A.K.; Latruffe, N.; Ghiringhelli, F.; Delmas, D. Resveratrol metabolites inhibit human metastatic colon cancer cells progression and synergize with chemotherapeutic drugs to induce cell death. Mol. Nutr. Food Res. 2013, 57, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Woo, K.J.; Lee, J.-T.; Lim, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Kwon, T.K. Resveratrol induces pro-apoptotic endoplasmic reticulum stress in human colon cancer cells. Oncol. Rep. 2007, 18, 1269–1273. [Google Scholar] [CrossRef]
- Saunier, E.; Antonio, S.; Regazzetti, A.; Auzeil, N.; Laprévote, O.; Shay, J.W.; Coumoul, X.; Barouki, R.; Benelli, C.; Huc, L. Resveratrol reverses the Warburg effect by targeting the pyruvate dehydrogenase complex in colon cancer cells. Sci. Rep. 2017, 7, 6945. [Google Scholar] [CrossRef]
- Vanamala, J.; Reddivari, L.; Radhakrishnan, S.; Tarver, C. Resveratrol suppresses IGF-1 induced human colon cancer cell proliferation and elevates apoptosis via suppression of IGF-1R/Wnt and activation of p53 signaling pathways. BMC Cancer 2010, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, G.; Jin, G.; Yao, K.; Zhao, Z.; Bie, L.; Guo, Y.; Li, N.; Deng, W.; Chen, X. Resveratrol suppresses colon cancer growth by targeting the AKT/STAT3 signaling pathway. Int. J. Mol. Med. 2019, 43, 630–640. [Google Scholar] [CrossRef]
- Feng, M.; Zhong, L.-X.; Zhan, Z.-Y.; Huang, Z.-H.; Xiong, J.-P. Resveratrol treatment inhibits proliferation of and induces apoptosis in human colon cancer cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 1101. [Google Scholar] [CrossRef]
- Milacic, V.; Banerjee, S.; Landis-Piwowar, K.R.; Sarkar, F.H.; Majumdar, A.P.; Dou, Q.P. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res. 2008, 68, 7283–7292. [Google Scholar] [CrossRef]
- Van Erk, M.J.; Teuling, E.; Staal, Y.C.; Huybers, S.; Van Bladeren, P.J.; Aarts, J.M.; Van Ommen, B. Time-and dose-dependent effects of curcumin on gene expression in human colon cancer cells. J. Carcinog. 2004, 3, 8. [Google Scholar] [CrossRef]
- Chen, A.; Xu, J.; Johnson, A. Curcumin inhibits human colon cancer cell growth by suppressing gene expression of epidermal growth factor receptor through reducing the activity of the transcription factor Egr-1. Oncogene 2006, 25, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Chang, J.Y.; Hahm, E.R.; Park, S.; Kim, H.-K.; Yang, C.H. Quercetin, a potent inhibitor against β-catenin/Tcf signaling in SW480 colon cancer cells. Biochem. Biophys. Res. Commun. 2005, 328, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Kim, S.-K.; Kim, B.-S.; Lee, S.-H.; Park, Y.-S.; Park, B.-K.; Kim, S.-J.; Kim, J.; Choi, C.; Kim, J.-S. Apoptotic effect of quercetin on HT-29 colon cancer cells via the AMPK signaling pathway. J. Agric. Food Chem. 2010, 58, 8643–8650. [Google Scholar] [CrossRef]
- Kim, W.K.; Bang, M.H.; Kim, E.S.; Kang, N.E.; Jung, K.C.; Cho, H.J.; Park, J.H. Quercetin decreases the expression of ErbB2 and ErbB3 proteins in HT-29 human colon cancer cells. J. Nutr. Biochem. 2005, 16, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Sung, B.; Kang, Y.J.; Kim, D.H.; Jang, J.-Y.; Hwang, S.Y.; Kim, M.; Lim, H.S.; Yoon, J.-H.; Chung, H.Y. Apigenin-induced apoptosis is enhanced by inhibition of autophagy formation in HCT116 human colon cancer cells. Int. J. Oncol. 2014, 44, 1599–1606. [Google Scholar] [CrossRef]
- Turktekin, M.; Konac, E.; Onen, H.I.; Alp, E.; Yilmaz, A.; Menevse, S. Evaluation of the effects of the flavonoid apigenin on apoptotic pathway gene expression on the colon cancer cell line (HT29). J. Med. Food 2011, 14, 1107–1117. [Google Scholar] [CrossRef]
- Gamet-Payrastre, L.; Li, P.; Lumeau, S.; Cassar, G.; Dupont, M.-A.; Chevolleau, S.; Gasc, N.; Tulliez, J.; Tercé, F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000, 60, 1426–1433. [Google Scholar]
- Cao, P.; Xia, Y.; He, W.; Zhang, T.; Hong, L.; Zheng, P.; Shen, X.; Liang, G.; Cui, R.; Zou, P. Enhancement of oxaliplatin-induced colon cancer cell apoptosis by alantolactone, a natural product inducer of ROS. Int. J. Biol. Sci. 2019, 15, 1676. [Google Scholar] [CrossRef]
- Su, M.-Q.; Zhou, Y.-R.; Rao, X.; Yang, H.; Zhuang, X.H.; Ke, X.-J.; Peng, G.-Y.; Zhou, C.-L.; Shen, B.-Y.; Dou, J. Baicalein induces the apoptosis of HCT116 human colon cancer cells via the upregulation of DEPP/Gadd45a and activation of MAPKs. Int. J. Oncol. 2018, 53, 750–760. [Google Scholar]
- Kim, S.-J.; Kim, H.-J.; Kim, H.-R.; Lee, S.-H.; Cho, S.-D.; Choi, C.-S.; Nam, J.-S.; Jung, J.-Y. Antitumor actions of baicalein and wogonin in HT-29 human colorectal cancer cells. Mol. Med. Rep. 2012, 6, 1443–1449. [Google Scholar] [CrossRef]
- Hsu, H.-H.; Chen, M.-C.; Day, C.H.; Lin, Y.-M.; Li, S.-Y.; Tu, C.-C.; Padma, V.V.; Shih, H.-N.; Kuo, W.-W.; Huang, C.-Y. Thymoquinone suppresses migration of LoVo human colon cancer cells by reducing prostaglandin E2 induced COX-2 activation. World J. Gastroenterol. 2017, 23, 1171. [Google Scholar] [CrossRef]
- Chen, M.-C.; Lee, N.-H.; Hsu, H.-H.; Ho, T.-J.; Tu, C.-C.; Hsieh, D.J.-Y.; Lin, Y.-M.; Chen, L.-M.; Kuo, W.-W.; Huang, C.-Y. Thymoquinone induces caspase-independent, autophagic cell death in CPT-11-resistant lovo colon cancer via mitochondrial dysfunction and activation of JNK and p38. J. Agric. Food Chem. 2015, 63, 1540–1546. [Google Scholar] [CrossRef]
- Tang, F.Y.; Shih, C.J.; Cheng, L.H.; Ho, H.J.; Chen, H.J. Lycopene inhibits growth of human colon cancer cells via suppression of the Akt signaling pathway. Mol. Nutr. Food Res. 2008, 52, 646–654. [Google Scholar] [CrossRef]
- Lin, M.-C.; Wang, F.-Y.; Kuo, Y.-H.; Tang, F.-Y. Cancer chemopreventive effects of lycopene: Suppression of MMP-7 expression and cell invasion in human colon cancer cells. J. Agric. Food Chem. 2011, 59, 11304–11318. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Dharmalingam, P.; Sadagopan, S.K.A.; Ramar, M.; Munusamy, A. Luteolin induces growth arrest in colon cancer cells through involvement of Wnt/β-catenin/GSK-3β signaling. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.S.; Won, S.B.; Kwon, Y.H. Luteolin induces apoptosis and autophagy in HCT116 colon cancer cells via p53-dependent pathway. Nutr. Cancer 2022, 74, 677–686. [Google Scholar] [CrossRef]
- Xiang, D.; Wang, D.; He, Y.; Xie, J.; Zhong, Z.; Li, Z.; Xie, J. Caffeic acid phenethyl ester induces growth arrest and apoptosis of colon cancer cells via the β-catenin/T-cell factor signaling. Anti-Cancer Drugs 2006, 17, 753–762. [Google Scholar] [CrossRef]
- Jaganathan, S.K. Growth inhibition by caffeic acid, one of the phenolic constituents of honey, in HCT 15 colon cancer cells. Sci. World J. 2012, 2012, 372345. [Google Scholar] [CrossRef]
- Adachi, S.; Nagao, T.; Ingolfsson, H.I.; Maxfield, F.R.; Andersen, O.S.; Kopelovich, L.; Weinstein, I.B. The inhibitory effect of (−)-epigallocatechin gallate on activation of the epidermal growth factor receptor is associated with altered lipid order in HT29 colon cancer cells. Cancer Res. 2007, 67, 6493–6501. [Google Scholar] [CrossRef]
- Adachi, S.; Shimizu, M.; Shirakami, Y.; Yamauchi, J.; Natsume, H.; Matsushima-Nishiwaki, R.; To, S.; Weinstein, I.B.; Moriwaki, H.; Kozawa, O. (−)-Epigallocatechin gallate downregulates EGF receptor via phosphorylation at Ser1046/1047 by p38 MAPK in colon cancer cells. Carcinogenesis 2009, 30, 1544–1552. [Google Scholar] [CrossRef]
- Cha, K.H.; Koo, S.Y.; Lee, D.-U. Antiproliferative effects of carotenoids extracted from Chlorella ellipsoidea and Chlorella vulgaris on human colon cancer cells. J. Agric. Food Chem. 2008, 56, 10521–10526. [Google Scholar] [CrossRef]
- Liao, P.-C.; Ng, L.-T.; Lin, L.-T.; Richardson, C.D.; Wang, G.-H.; Lin, C.-C. Resveratrol arrests cell cycle and induces apoptosis in human hepatocellular carcinoma Huh-7 cells. J. Med. Food 2010, 13, 1415–1423. [Google Scholar] [CrossRef]
- Notas, G.; Nifli, A.-P.; Kampa, M.; Vercauteren, J.; Kouroumalis, E.; Castanas, E. Resveratrol exerts its antiproliferative effect on HepG2 hepatocellular carcinoma cells, by inducing cell cycle arrest, and NOS activation. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2006, 1760, 1657–1666. [Google Scholar] [CrossRef]
- Pan, Z.; Zhuang, J.; Ji, C.; Cai, Z.; Liao, W.; Huang, Z. Curcumin inhibits hepatocellular carcinoma growth by targeting VEGF expression. Oncol. Lett. 2018, 15, 4821–4826. [Google Scholar] [CrossRef]
- Kabir, M.S.H.; Hossain, M.M.; Kabir, M.I.; Rahman, M.M.; Hasanat, A.; Emran, T.B.; Rahman, M.A. Phytochemical screening, Antioxidant, Thrombolytic, alpha-amylase inhibition and cytotoxic activities of ethanol extract of Steudnera colocasiifolia K. Koch leaves. J. Young Pharm. 2016, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Li, J.; Liu, T.; Li, S.; Feng, J.; Yu, Q.; Zhang, J.; Chen, J.; Zhou, Y.; Ji, J. Quercetin shows anti-tumor effect in hepatocellular carcinoma LM3 cells by abrogating JAK2/STAT3 signaling pathway. Cancer Med. 2019, 8, 4806–4820. [Google Scholar] [CrossRef]
- Dai, W.; Gao, Q.; Qiu, J.; Yuan, J.; Wu, G.; Shen, G. Quercetin induces apoptosis and enhances 5-FU therapeutic efficacy in hepatocellular carcinoma. Tumor Biol. 2016, 37, 6307–6313. [Google Scholar] [CrossRef]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, D.; Zhou, H.-g.; Wang, X.-h.; Zhong, W.-l.; Chen, S.; Gu, W.-g.; Wang, W.; Zhang, C.-h.; Liu, Y.-r. Apigenin inhibits NF-κB and snail signaling, EMT and metastasis in human hepatocellular carcinoma. Oncotarget 2016, 7, 41421. [Google Scholar] [CrossRef]
- Liu, P.; Atkinson, S.J.; Akbareian, S.E.; Zhou, Z.; Munsterberg, A.; Robinson, S.D.; Bao, Y. Sulforaphane exerts anti-angiogenesis effects against hepatocellular carcinoma through inhibition of STAT3/HIF-1α/VEGF signalling. Sci. Rep. 2017, 7, 12651. [Google Scholar] [CrossRef]
- Moon, D.-O.; Kang, S.-H.; Kim, K.-C.; Kim, M.-O.; Choi, Y.H.; Kim, G.-Y. Sulforaphane decreases viability and telomerase activity in hepatocellular carcinoma Hep3B cells through the reactive oxygen species-dependent pathway. Cancer Lett. 2010, 295, 260–266. [Google Scholar] [CrossRef]
- Rauf, A.; Abu-Izneid, T.; Khalil, A.A.; Imran, M.; Shah, Z.A.; Emran, T.B.; Mitra, S.; Khan, Z.; Alhumaydhi, F.A.; Aljohani, A.S. Berberine as a potential anticancer agent: A comprehensive review. Molecules 2021, 26, 7368. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, S.; Ji, Y.; Li, J.; An, P.; Ren, H.; Liang, R.; Yang, J.; Li, Z. Baicalein inhibits the invasion and metastatic capabilities of hepatocellular carcinoma cells via down-regulation of the ERK pathway. PLoS ONE 2013, 8, e72927. [Google Scholar] [CrossRef]
- Islam, M.R.; Islam, F.; Nafady, M.H.; Akter, M.; Mitra, S.; Das, R.; Urmee, H.; Shohag, S.; Akter, A.; Chidambaram, K. Natural small molecules in breast cancer treatment: Understandings from a therapeutic viewpoint. Molecules 2022, 27, 2165. [Google Scholar] [CrossRef]
- Bie, B.; Sun, J.; Li, J.; Guo, Y.; Jiang, W.; Huang, C.; Yang, J.; Li, Z. Baicalein, a natural anti-cancer compound, alters microRNA expression profiles in Bel-7402 human hepatocellular carcinoma cells. Cell. Physiol. Biochem. 2017, 41, 1519–1531. [Google Scholar] [CrossRef]
- Ahmed, W.A.; Hassan, S.A.; Galeb, F.M.; El-Taweel, M.A.; Abu-Bedair, F.A. The in vitro promising therapeutic activity of thymoquinone on hepatocellular carcinoma (HepG2) cell line. Glob. Vet. 2008, 2, 233–241. [Google Scholar]
- Tallei, T.E.; Niode, N.J.; Idroes, R.; Zidan, B.M.R.M.; Mitra, S.; Celik, I.; Nainu, F.; Ağagündüz, D.; Emran, T.B.; Capasso, R. A Comprehensive Review of the Potential Use of Green Tea Polyphenols in the Management of COVID-19. Evid-Based Complement. Altern. Med. 2021, 2021, 7170736. [Google Scholar] [CrossRef]
- Gupta, P.; Bhatia, N.; Bansal, M.P.; Koul, A. Lycopene modulates cellular proliferation, glycolysis and hepatic ultrastructure during hepatocellular carcinoma. World J. Hepatol. 2016, 8, 1222. [Google Scholar] [CrossRef]
- Balamurugan, K.; Karthikeyan, J. Evaluation of luteolin in the prevention of N-nitrosodiethylamine-induced hepatocellular carcinoma using animal model system. Indian J. Clin. Biochem. 2012, 27, 157–163. [Google Scholar] [CrossRef]
- Gu, W.; Yang, Y.; Zhang, C.; Zhang, Y.; Chen, L.; Shen, J.; Li, G.; Li, Z.; Li, L.; Li, Y. Caffeic acid attenuates the angiogenic function of hepatocellular carcinoma cells via reduction in JNK-1-mediated HIF-1α stabilization in hypoxia. RSC Adv. 2016, 6, 82774–82782. [Google Scholar] [CrossRef]
- Wang, L.; Lu, M.; Yi, M.; Chen, L.; Shen, J.; Li, Z.; Li, L.; Yang, Y.; Zhang, J.; Li, Y. Caffeic acid attenuates the autocrine IL-6 in hepatocellular carcinoma via the epigenetic silencing of the NF-κB-IL-6-STAT-3 feedback loop. RSC Adv. 2015, 5, 52952–52957. [Google Scholar] [CrossRef]
- Shirakami, Y.; Shimizu, M.; Adachi, S.; Sakai, H.; Nakagawa, T.; Yasuda, Y.; Tsurumi, H.; Hara, Y.; Moriwaki, H. (–)-Epigallocatechin gallate suppresses the growth of human hepatocellular carcinoma cells by inhibiting activation of the vascular endothelial growth factor–vascular endothelial growth factor receptor axis. Cancer Sci. 2009, 100, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Helmy, S.A.; El-Mesery, M.; El-Karef, A.; Eissa, L.A.; El Gayar, A.M. Thymoquinone upregulates TRAIL/TRAILR2 expression and attenuates hepatocellular carcinoma in vivo model. Life Sci. 2019, 233, 116673. [Google Scholar] [CrossRef]
- Bernhaus, A.; Ozsvar-Kozma, M.; Saiko, P.; Jaschke, M.; Lackner, A.; Grusch, M.; Horvath, Z.; Madlener, S.; Krupitza, G.; Handler, N. Antitumor effects of KITC, a new resveratrol derivative, in AsPC-1 and BxPC-3 human pancreatic carcinoma cells. Investig. New Drugs 2009, 27, 393–401. [Google Scholar] [CrossRef]
- Harikumar, K.B.; Kunnumakkara, A.B.; Sethi, G.; Diagaradjane, P.; Anand, P.; Pandey, M.K.; Gelovani, J.; Krishnan, S.; Guha, S.; Aggarwal, B.B. Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabine in vitro and in orthotopic mouse model of human pancreatic cancer. Int. J. Cancer 2010, 127, 257–268. [Google Scholar]
- Hidaka, H.; Ishiko, T.; Furuhashi, T.; Kamohara, H.; Suzuki, S.; Miyazaki, M.; Ikeda, O.; Mita, S.; Setoguchi, T.; Ogawa, M. Curcumin inhibits interleukin 8 production and enhances interleukin 8 receptor expression on the cell surface: Impact on human pancreatic carcinoma cell growth by autocrine regulation. Cancer 2002, 95, 1206–1214. [Google Scholar] [CrossRef]
- Borska, S.; Drag-Zalesinska, M.; Wysocka, T.; Sopel, M.; Dumanska, M.; Zabel, M.; Dziegiel, P. Antiproliferative and pro-apoptotic effects of quercetin on human pancreatic carcinoma cell lines EPP85-181P and EPP85-181RDB. Folia Histochem. Et Cytobiol. 2010, 48, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Lan, C.-Y.; Chen, S.-Y.; Kuo, C.-W.; Lu, C.-C.; Yen, G.-C. Quercetin facilitates cell death and chemosensitivity through RAGE/PI3K/AKT/mTOR axis in human pancreatic cancer cells. J. Food Drug Anal. 2019, 27, 887–896. [Google Scholar] [CrossRef]
- Ma, J.; Li, Q.; Zhao, J.; Guo, Y.; Su, Q.; Ji, Z. Effects of apigenin on cell proliferation of human pancreatic carcinoma cell line BxPC-3 in vitro. J. Nanjing Med. Univ. 2007, 21, 94–98. [Google Scholar] [CrossRef]
- Rahman, M.M.; Islam, M.R.; Shohag, S.; Hossain, M.E.; Rahaman, M.S.; Islam, F.; Ahmed, M.; Mitra, S.; Khandaker, M.U.; Idris, A.M. The multifunctional role of herbal products in the management of diabetes and obesity: A comprehensive review. Molecules 2022, 27, 1713. [Google Scholar] [CrossRef]
- Kallifatidis, G.; Rausch, V.; Baumann, B.; Apel, A.; Beckermann, B.M.; Groth, A.; Mattern, J.; Li, Z.; Kolb, A.; Moldenhauer, G. Sulforaphane targets pancreatic tumour-initiating cells by NF-κB-induced antiapoptotic signalling. Gut 2009, 58, 949–963. [Google Scholar] [CrossRef]
- Li, S.-H.; Fu, J.; Watkins, D.N.; Srivastava, R.K.; Shankar, S. Sulforaphane regulates self-renewal of pancreatic cancer stem cells through the modulation of Sonic hedgehog–GLI pathway. Mol. Cell. Biochem. 2013, 373, 217–227. [Google Scholar] [CrossRef]
- Zheng, H.; Yang, L.; Kang, Y.; Chen, M.; Lin, S.; Xiang, Y.; Li, C.; Dai, X.; Huang, X.; Liang, G. Alantolactone sensitizes human pancreatic cancer cells to EGFR inhibitors through the inhibition of STAT3 signaling. Mol. Carcinog. 2019, 58, 565–576. [Google Scholar] [CrossRef]
- He, R.; Shi, X.; Zhou, M.; Zhao, Y.; Pan, S.; Zhao, C.; Guo, X.; Wang, M.; Li, X.; Qin, R. Alantolactone induces apoptosis and improves chemosensitivity of pancreatic cancer cells by impairment of autophagy-lysosome pathway via targeting TFEB. Toxicol. Appl. Pharmacol. 2018, 356, 159–171. [Google Scholar] [CrossRef]
- Wang, Y.-M. Inhibitory effects of thymoquinone on human pancreatic carcinoma orthotopically implanted in nude mice. Zhonghua Yi Xue Za Zhi 2011, 91, 3111–3114. [Google Scholar]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene inhibits reactive oxygen species-mediated NF-κB signaling and induces apoptosis in pancreatic cancer cells. Nutrients 2019, 11, 762. [Google Scholar] [CrossRef]
- Cai, X.; Lu, W.; Ye, T.; Lu, M.; Wang, J.; Huo, J.; Qian, S.; Wang, X.; Cao, P. The molecular mechanism of luteolin-induced apoptosis is potentially related to inhibition of angiogenesis in human pancreatic carcinoma cells. Oncol. Rep. 2012, 28, 1353–1361. [Google Scholar] [CrossRef]
- Chen, M.-J.; Chang, W.-H.; Lin, C.-C.; Liu, C.-Y.; Wang, T.-E.; Chu, C.-H.; Shih, S.-C.; Chen, Y.-J. Caffeic acid phenethyl ester induces apoptosis of human pancreatic cancer cells involving caspase and mitochondrial dysfunction. Pancreatology 2008, 8, 566–576. [Google Scholar] [CrossRef]
- Chen, M.-J.; Shih, S.-C.; Wang, H.-Y.; Lin, C.-C.; Liu, C.-Y.; Wang, T.-E.; Chu, C.-H.; Chen, Y.-J. Caffeic acid phenethyl ester inhibits epithelial-mesenchymal transition of human pancreatic cancer cells. Evid.-Based Complement. Altern. Med. 2013, 2013, 270906. [Google Scholar] [CrossRef]
- Ujiki, M.B.; Ding, X.-Z.; Salabat, M.R.; Bentrem, D.J.; Golkar, L.; Milam, B.; Talamonti, M.S.; Bell, R.H.; Iwamura, T.; Adrian, T.E. Apigenin inhibits pancreatic cancer cell proliferation through G2/M cell cycle arrest. Mol. Cancer 2006, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. WJG 2006, 12, 354. [Google Scholar] [CrossRef] [PubMed]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma: An attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Gu, J.; Wu, H.; Rauf, A.; Emran, T.B.; Khan, Z.; Mitra, S.; Aljohani, A.S.M.; Alhumaydhi, F.A.; Al-Awthan, Y.S.; et al. Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants 2022, 11, 1158. [Google Scholar] [CrossRef]
- Mavroudis, D.; Kourousis, C.; Androulakis, N.; Kalbakis, K.; Agelaki, S.; Kakolyris, S.; Souglakos, J.; Sarra, E.; Vardakis, N.; Hatzidaki, D. Frontline treatment of advanced gastric cancer with docetaxel and granulocyte colony-stimulating factor (G-CSF): A phase II trial. Am. J. Clin. Oncol. 2000, 23, 341–344. [Google Scholar] [CrossRef]
- Woell, E.; Devries, A.; Eisterer, W.; Hejna, M.; Keil, F.; Stein, H.; Zacherl, J.; Greil, R. Chemotherapy in gastric cancer. Anticancer Res. 2008, 28, 1213–1219. [Google Scholar]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Shariati, M.A.; Rebezov, M.; Rengasamy, K.R.; Mubarak, M.S. Superoxide dismutase: An updated review on its health benefits and industrial applications. Crit. Rev. Food Sci. Nutr. 2021, 2021, 1–19. [Google Scholar] [CrossRef]
- Liu, W.; Guo, Q.-L.; You, Q.-D.; Zhao, L.; Gu, H.-Y.; Yuan, S.-T. Anticancer effect and apoptosis induction of gambogic acid in human gastric cancer line BGC-823. World J. Gastroenterol. WJG 2005, 11, 3655. [Google Scholar] [CrossRef]
- Patel, B.B.; Majumdar, A.P. Synergistic role of curcumin with current therapeutics in colorectal cancer: Minireview. Nutr. Cancer 2009, 61, 842–846. [Google Scholar] [CrossRef]
- Zou, Z.; Xie, L.; Wei, J.; Yu, L.; Qian, X.; Chen, J.; Wang, T.; Liu, B. Synergistic anti-proliferative effects of gambogic acid with docetaxel in gastrointestinal cancer cell lines. BMC Complement. Altern. Med. 2012, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Rakib, A.; Islam, M.A.; Khanam, B.H.; Faiz, F.B.; Paul, A.; Chy, M.N.U.; Bhuiya, N.M.M.A.; Uddin, M.M.N.; Ullah, S.M.A.; et al. In vivo and in vitro pharmacological activities of Tacca integrifolia rhizome and investigation of possible lead compounds against breast cancer through in silico approaches. Clin. Phytosci. 2019, 5, 36. [Google Scholar] [CrossRef]
- Tisman, G.; Wu, S. Effectiveness of intermediate-dose methotrexate and high-dose 5-fluorouracil as sequential combination chemotherapy in refractory breast cancer and as primary therapy in metastatic adenocarcinoma of the colon. Cancer Treat. Rep. 1980, 64, 829–835. [Google Scholar]
- Mulder, J.; Smink, T.; Van Putten, L. 5-Fluorouracil and methotrexate combination chemotherapy: The effect of drug scheduling. Eur. J. Cancer Clin. Oncol. 1981, 17, 831–837. [Google Scholar] [CrossRef]
- Hibino, Y.; Kawarabayashi, Y.; Kohri, H.; Ueda, N.; Tsukagoshi, S. The mechanism of potentiation of the antitumor effect of 5-fluorouracil by methionine-free intravenous amino acid solution (AO-90) in rats. Gan Kagaku Ryoho. Cancer Chemother. 1994, 21, 2021–2028. [Google Scholar]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. eBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Mitra, S.; Tareq, A.M.; Das, R.; Emran, T.B.; Nainu, F.; Chakraborty, A.J.; Ahmad, I.; Tallei, T.E.; Idris, A.M.; Simal-Gandara, J. Polyphenols: A first evidence in the synergism and bioactivities. Food Rev. Int. 2022, 2022, 1–23. [Google Scholar] [CrossRef]
- Tareq, A.M.; Farhad, S.; Uddin, A.N.; Hoque, M.; Nasrin, M.S.; Uddin, M.M.R.; Hasan, M.; Sultana, A.; Munira, M.S.; Lyzu, C.; et al. Chemical profiles, pharmacological properties, and in silico studies provide new insights on Cycas pectinata. Heliyon 2020, 6, e04061. [Google Scholar] [CrossRef]
- Al Mahmud, Z.; Emran, T.B.; Qais, N.; Bachar, S.C.; Sarker, M.; Uddin, M.M.N. Evaluation of analgesic, anti-inflammatory, thrombolytic and hepatoprotective activities of roots of Premna esculenta (Roxb). J. Basic Clin. Physiol. Pharmacol. 2016, 27, 63–70. [Google Scholar] [CrossRef]
- Dutta, T.; Paul, A.; Majumder, M.; Sultan, R.A.; Emran, T.B. Pharmacological evidence for the use of Cissus assamica as a medicinal plant in the management of pain and pyrexia. Biochem. Biophys. Rep. 2020, 21, 100715. [Google Scholar] [CrossRef]
- Rahman, J.; Tareq, A.M.; Hossain, M.; Sakib, S.A.; Islam, M.N.; Ali, M.H.; Uddin, A.B.M.N.; Hoque, M.; Nasrin, M.S.; Emran, T.B.; et al. Biological evaluation, DFT calculations and molecular docking studies on the antidepressant and cytotoxicity activities of Cycas pectinata Buch.-Ham. Compounds. Pharmaceuticals 2020, 13, 232. [Google Scholar] [CrossRef]






| Name of Compound | Subject | Dose | Result | Mechanism of Action | Reference |
|---|---|---|---|---|---|
| Resveratrol | In vitro (CaCo-2 human colon cancer cells) | 25 μM | 70% growth inhibition | Reduction in the amount of ornithine decarboxylase enzyme | [59] |
| In vitro (SW480, SW620, and HCT116 cell line) | 10–100 μm | Cell death | Conformational changes of Bax and caspase activation | [61] | |
| In vitro (Caco-2 cell) | 12.5–200 μmol/L | Inhibition of cell development and proliferation | Increased activity of caspase-3, S-phase arrest, reduced production of cdk-4 and cyclin D1 | [62] | |
| In vitro (SW620 cell) | 10 μM | Cell growth inhibition | Elevation of mitochondrial oxygen consumption rate and ATP production | [63] | |
| In vitro (WiDr and HT-29 cell) | 5, 10, 20 and 40 µg/mL | Reduction in cell viability | Inhibition of TLMA and reduced activity of telomerase | [64] | |
| In vitro (SW620 and SW480) | 30 µM | Cell proliferation inhibited | S-phase arrest, elevated initiation of cyclins B and A expression, increased level of Cdk 2 protein, increase in phosphorylated histone H2AX, activation of p53 protein, activation of ATR/p53 pathway | [65] | |
| In vitro (HT-29 cell) | 50 and 100 µM | Reduction in cell progression | Endoplasmic reticulum stress, inhibition of caspase-4 | [66] | |
| In vitro (Caco2 colon cell) | 10 µM | Reduction in cell progression | Inhibition of pentose phosphate pathway, S phase arrest | [67] | |
| In vitro (SW480 and HT-29 cells) | 100–150 μM | Reduction of cell proliferation | P27 stimulation, cyclin D1 suppression, IGF-1R suppression, p53 activation | [68] | |
| In vitro (HCT15 and DLD1 cells) | 10, 20, 30, and 40 μM | Reduction of cell proliferation | G1 phase arrest, reduced cyclin D1, E2, and BCL2 expression, elevated P53 level. | [69] | |
| In vitro (SW480 and HT-29 cells) | 30 μM | Cell death | Inhibition of COX-2 and reduced expression of PEG2 | [70] | |
| Curcumin | In vitro (SW480 and HCT116 cells) | 10, 20, and 30 μmol/L | Cell proliferation inhibition | Inhibition of 20S proteasome activity | [71] |
| In vitro (HT-29 cells) | - | Reduction in cancer cell | G2/M cell cycle arrest, downregulation of cytochrome P450 gene | [72] | |
| In vitro (Moser cell) | 0, 5, 10, 15 and 20 μM | Cell progression inhibition | Reduced expression of EGFR and cyclin D1 | [73] | |
| Quercetin | In vitro (HEK293 and SW480 cells) | 0–100 μM | Cell progression inhibition | Inhibition of Beta-catenin/Tcf signaling pathway | [74] |
| In vitro (HT-29 cells) | 100 mg/kg | Inhibition of cancer cell development | G1 cell cycle arrest, upregulation of p21, AMPK, and p53 | [75] | |
| In vitro (HT-29 cells) | 0, 25, 50, and 100 μmol/L | Apoptosis and cell viability decline | Restriction of ErbB2/ErbB3 signaling, Akt pathway, and Bcl-2 level | [76] | |
| Apigenin | In vitro (HCT116 cells) | 25 and 50 μM | Apoptosis induction | G2/M phase cell cycle inhibition, suppression of cyclin B1, elevated expression of P53 and p53-dependent p21CIP1/WAF1, decreased level of procaspase-8,9,3 | [77] |
| In vitro (HT29 cell) | 90 μM | Induced apoptosis | Caspase-3 and caspase-8 expression increased, mTOR and cyclin D1 expression decreased | [78] | |
| Sulforaphane | In vitro (HT29 cells) | 5–50 μM | Induced apoptosis | Increased initiation of cyclin A and cyclin B1 | [79] |
| Alantolactone | In vitro and In vivo (HCT116, RKO cells and BALB/c mice) | 40–120 µM | Cancer cell death | Activation of JNK and p38 MAPK signaling pathway | [80] |
| Baicalein | In vitro (panc-1, HTC116 and A549 cells) | 0, 10, 20 or 40 µM | Apoptosis | Upregulation of Gadd45a and DEPP | [81] |
| In vitro (HT29 cells) | 100 μM | Cell viability reduced | G1 cell cycle arrest, reduced Bcl-2 expression, increased Bax expression, and PI3K/AKT pathway inactivation | [82] | |
| Thymoquinone | In vitro (LoVo cells) | 20 μmol/L | Inhibited migration and cell growth | p-PI3K, p-Akt, p-GSK3, and beta-catenin levels were all reduced. | [83] |
| In vitro (LoVo cells) | 2 μM | Cell death | Mitochondrial outer membrane permeability increased when JNK and p38 were activated. | [84] | |
| Lycopene | In vitro (HT-29 cells) | 10 μM | Cell proliferation halted | Nonphosphorylated beta-catenin protein and reduced Akt activation | [85] |
| In vitro (HT-29 cells) | 250 nM | Inhibition of cancer cell progression | Inhibition of MMP-7 | [86] | |
| Luteolin | In vitro (HCT-15 cells) | 100 µM | Cell cycle arrest and apoptosis | Reduced expression of non-P-beta-catenin, phosphorylated glycogen synthase kinase-3beta, and cyclin D1. | [87] |
| In vitro (HT-29 cells) | 10–20 μM | Cell cycle blockade and apoptosis | Increased p53 phosphorylation and p53 target gene expression | [88] | |
| Caffeic acid | In vitro (HCT116 and SW480 cells) | 2.5, 5 or 10 mg/mL | Apoptosis and cell growth inhibition | Cyclin D1 and c-myc expression were reduced in a dose-dependent manner. | [89] |
| In vitro (HCT 15 cells) | 800 μM | Induced apoptosis | Increased generation of ROS and decreased mitochondrial membrane potential | [90] | |
| Epigallocatechin gallate | In vitro (HT-29 cells) | 5 μg/mL | Inhibits cancer cell growth | Inhibited epidermal growth factor receptor | [91] |
| In vitro (SW480 cells) | 25 μM | EGFR is downregulated | Activation of p38 MAPK resulted in phosphorylation of EGFR at serine 1046/1047 | [92] | |
| Carotenoids | In vitro (HCT116 cell) | - | Cell growth inhibited | Induced apoptosis | [93] |
| Name of Compound | Subject | Dose | Result | Mechanism of Action | Reference |
|---|---|---|---|---|---|
| Resveratrol | In vitro (Huh-7 cells) | 22.4 μg/mL | Cell cycle arrest and apoptosis induction | Although cyclin E, cyclin A, and cyclin-dependent kinase 2 expression was downregulated, p21/WAF1 expression was elevated in a p53-independent manner | [94] |
| In vitro (HepG2 cell lines) | 10−7 M | Cell progression inhibited | Cell cycle inhibited in G1 and G2/M phase | [95] | |
| Curcumin | In vitro and in vivo (H22 HCC cells and mice) | 50 and 100 mg/kg | Cell proliferation inhibited | The signaling pathways PI3K/AKT and vascular endothelial growth factor were inhibited. | [96] |
| In vitro (HepG2 cells) | 300–3000 mg/kg | Cytotoxic activity | Suppression of NF-kB, AP-1 STAT3, STAT4, peroxisome proliferators-associated receptor gamma, cyclin D1 | [97] | |
| Quercetin | In vitro and in vivo (LM3 cell line and mice) | 20–200 μmol/L | Induction of apoptosis | Down-regulation of JAK2 and STAT3 | [98] |
| In vitro (HepG2 and SMCC cell line) | 0.05, 0.10, or 0.15 mM | Cell proliferation inhibition and apoptosis induction | Increased expression of Bad and Bax, with a reduction in Bcl-2 expression | [99] | |
| Apigenin | In vitro (HepG2 cells) | 10, 20 and 40 μM | Apoptosis and autophagy induction | Eradication of PI3K/Akt/mTOR pathway | [100] |
| In vitro (HCC cell lines) | 10 and 20 μM | Inhibited the migration and metastasis | Snai1 and NF-kB expression were reduced, but EMT marker levels rose. | [101] | |
| Sulforaphane | In vitro and in vivo (HCC cell line, HepG2 and human endothelial cell) | 1.25, 2.5, 5, 10 and 20 μM | Reduced tumor growth | STAT3/HIF-1α/VEGF signaling inhibition | [102] |
| In vitro (Hep3B cells) | 20 μM | Cell viability inhibited | Downregulation of telomerase reverse transcriptase and suppression of phosphorylation of Akt | [103] | |
| Baicalein | In vitro and in vivo (HCC cell lines and mice) | 50 μM | Reduction in cell motility and migration | MMP-2, MMP-9, and u-PA levels fell, while MEK1 and ERK 1/2 were phosphorylated. | [105] |
| In vitro (HCC cell line, Bel-7402 and Hep3B) | 40 and 80 μM | Cell proliferation inhibited | During the S and G2/M phases, cell cycle arrest was accomplished by upregulating the expression of p21/CDKN1A and P27/CDKN1B and blocking the PI3K/Akt pathway | [107] | |
| Thymoquinone | In vivo (male Sprague rats) | 20 mg/kg | Inhibited cancer cell progression | Upregulation of TRAIL/TRAILR2, caspase-3, and Bcl-2 | [115] |
| In vitro (HepG2) | 25, 50, 100, 200 and 400 μM | Inhibition of cancer cell growth | Increase in Caspase 2 and 9; G1/S cell cycle arrest | [108] | |
| Lycopene | In vivo (Female BALB/c mice) | 5 mg/kg | Induction of apoptosis | Enhanced expression of PCNA and cyclin D1 | [110] |
| Luteolin | In vivo (Albino mice) | 0.2 mg/kg | Induction of apoptosis | Altered tissue-damaging enzymes and enzymatic antioxidants. | [111] |
| Caffeic acid | In vitro and in vivo (HepG2, HUVECs and BALB/c mice) | 20 μM | Attenuation of the angiogenic function | JNK-1-mediated HIF-1α stabilization was reduced.. | [112] |
| In vitro (HepG2 and MHCC97H) | 10, 20 or 40 μM | Inhibits cancer cell progression | Endogenous Interleukin-6 expression was inhibited. | [113] | |
| Epigallocatechin gallate | In vitro (HLF, PLC/PRF/5, HepG2, HuH7, HLE, and Hep3B) | 25 µg/mL | Inhibition of cancer cell growth | VEGFR-2 and p-VEGFR-2 protein expression reduced; ERK and Akt signaling pathways inhibited. | [114] |
| Name of Compound | Subject | Dose | Result | Mechanism of Action | Reference |
|---|---|---|---|---|---|
| Resveratrol | In vitro (AsPC-1 and BxPC-3) | 10, 20 and 30 μM | Induction of apoptosis | Arrested cells in the G0/G1 phase of the cell cycle 2 and depleted cells in the S phase | [116] |
| In vitro and in vivo (AsPC-1 and Male athymic nu/nu mice) | 10 μM | Cell proliferation inhibited | NF-kB activation, as well as the expression of bcl-2, bcl-xL, COX-2, cyclin D1, MMP-9, and VEGF, were all inhibited. | [117] | |
| Curcumin | In vitro (SUIT-2) | 10–100 μM | Cell proliferation inhibited | Reduces IL-8 | [118] |
| Quercetin | In vitro (EPP85-181P and EPP85-181RDB) | Antiproliferative and proapoptotic effect | Affecting ERK signal transduction pathway | [119] | |
| In vitro (A Paca-2, BxPC-3, AsPC-, HPAC and PANC-1) | 6.25, 12.5, 25, and 50 μM | Cancer cell death | Restricted RAGE in pancreatic cancer cell | [120] | |
| Apigenin | In vitro (BxPC-3) | 100, 200, and 400 μmol/L | Death of cancer cells | The G2/M cell cycle was halted and Bcl-2 expression was lowered, while Bax gene expression was increased. | [121] |
| In vitro (AsPC-1, CD18, MIA PaCa2, and S2-013) | 6.25–100 μM | Cancer cell death | DNA synthesis washindered, the G2/M cell cycle was arrested, and levels of cyclin A, cyclin B, cdc2, and cdc25 were all reduced. | [132] | |
| Sulforaphane | In vitro (AsPC-1, BxPc-3, Capan-1 and MIA-PaCa2) | 10 μM | Cell proliferation inhibited | NF-kB binding was prevented, downregulating apoptosis inhibitors and inducing apoptosis | [123] |
| In vitro and in vivo (Human PC stem cells and mice) | 10 μM | Cell proliferation inhibited | Blockade of hedghehog pathway, smo, Gli1, Gli2, Nanong, Oct-4, VEGF and PDGFRα | [124] | |
| Alantolactone | In vitro and in vivo (BxPC-3, AsPC1, PANC-1 and BALB/c mice) | 1.98 and 2.15 μM | Cancer cell death | Downregulation of STAT3 signaling pathway | [125] |
| In vitro and in vivo (MIA PaCa-2, PANC-1 and BALB/c mice) | 10 μM | Apoptosis | CTSB/CTSD protein activity and expression were inhibited and TFEB was reduced. | [126] | |
| Thymoquinone | In vivo (Nude mice) | 5 and 20 mg/kg | Apoptosis | Reduced XIAP and MMP-9 expression | [127] |
| Lycopene | In vitro (PANC-1) | 0.25 and 0.5 μM | Reduced cancer cell growth | Decreased ROS level and NF-kB expression and increased caspase-3 and Bax to Bcl-2 ratio | [128] |
| Luteolin | In vitro (PANC-1, CoLo-357 and BxPC-3) | 40 μmol/L | Induces programmed cell death | Increased Bax while reducing Bcl-2 protein and increasing caspase-3 | [129] |
| Caffeic acid | In vitro (BxPC-3 and PANC-1) | 10 µg/mL | Induces apoptosis | Mitochondrial dysfunction and activation of caspase-3/caspase-7 | [130] |
| In vitro and in vivo (PANC-1 and BALB/c mice) | 5 μg/mL | Reduces cancer cell progression | Inhibited the expression of Twist 2 and vimentin | [131] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, F.; Mitra, S.; Emran, T.B.; Khan, Z.; Nath, N.; Das, R.; Sharma, R.; Awadh, A.A.A.; Park, M.N.; Kim, B. Natural Small Molecules in Gastrointestinal Tract and Associated Cancers: Molecular Insights and Targeted Therapies. Molecules 2022, 27, 5686. https://doi.org/10.3390/molecules27175686
Islam F, Mitra S, Emran TB, Khan Z, Nath N, Das R, Sharma R, Awadh AAA, Park MN, Kim B. Natural Small Molecules in Gastrointestinal Tract and Associated Cancers: Molecular Insights and Targeted Therapies. Molecules. 2022; 27(17):5686. https://doi.org/10.3390/molecules27175686
Chicago/Turabian StyleIslam, Fahadul, Saikat Mitra, Talha Bin Emran, Zidan Khan, Nikhil Nath, Rajib Das, Rohit Sharma, Ahmed Abdullah Al Awadh, Moon Nyeo Park, and Bonglee Kim. 2022. "Natural Small Molecules in Gastrointestinal Tract and Associated Cancers: Molecular Insights and Targeted Therapies" Molecules 27, no. 17: 5686. https://doi.org/10.3390/molecules27175686