
A Multi-Omics Analysis Reveals Anti-Osteoporosis Mechanism of Four Components from Crude and Salt-Processed Achyranthes bidentata Blume in Ovariectomized Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
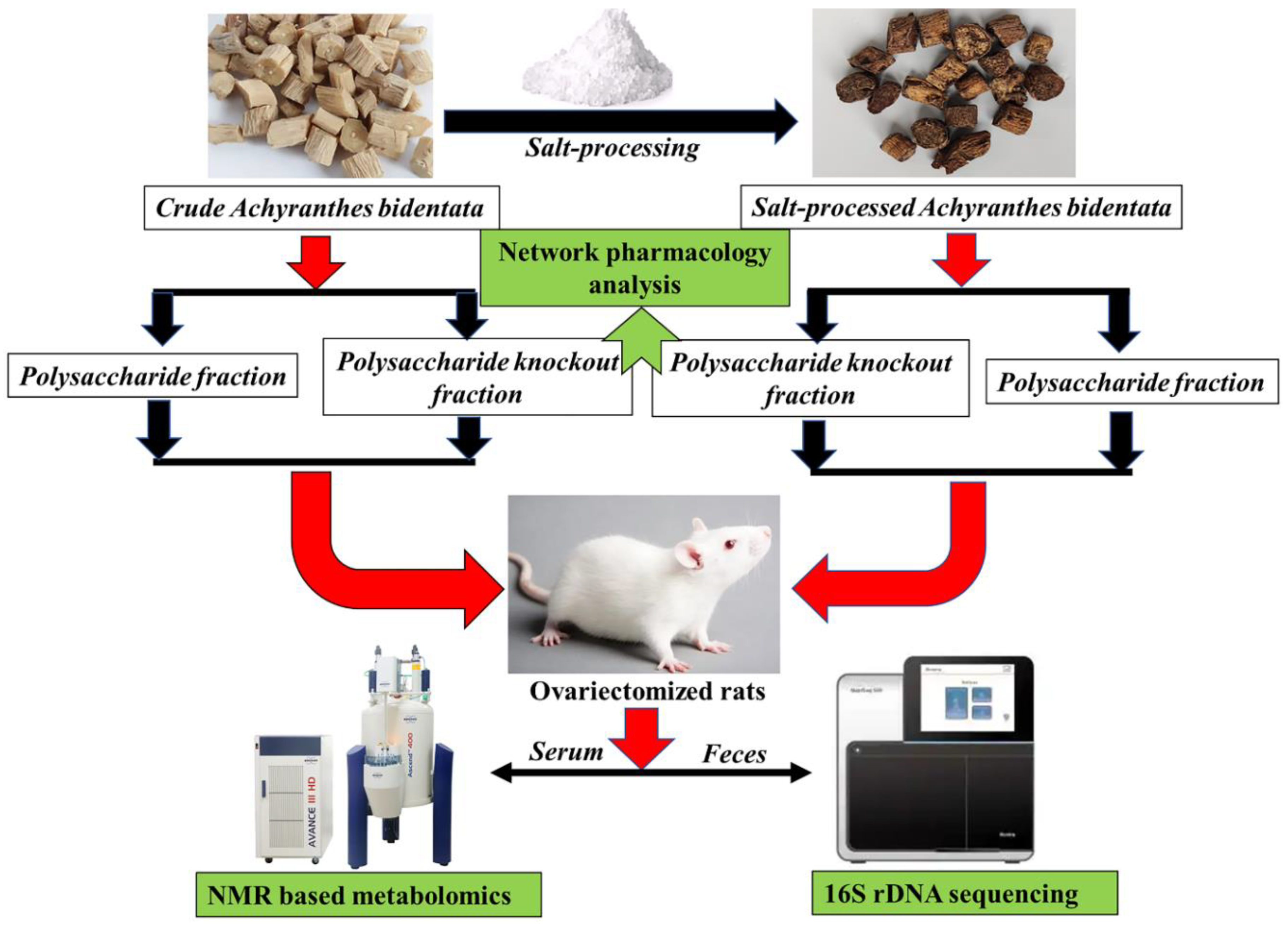
2.2. Preparation of Four Components of Crude and Salt-Processed AB
2.3. Experimental Design
2.4. Network Pharmacology Analysis
2.5. Animals and Drug Administration
2.6. Micro-CT and Dual-Energy X-ray Absorptiometry Analysis
2.7. Biochemical Index Determination
2.8. 1H-NMR Spectroscoy Based Metabolic Profiling
2.9. 16S rDNA Sequencing
2.10. Multivariate Statistical Analysis
3. Results and Discussion
3.1. Network Pharmacology Analysis
3.2. Biochemical Indexes
3.3. Micro-CT and Dual-Energy X-ray Absorptiometry Analysis
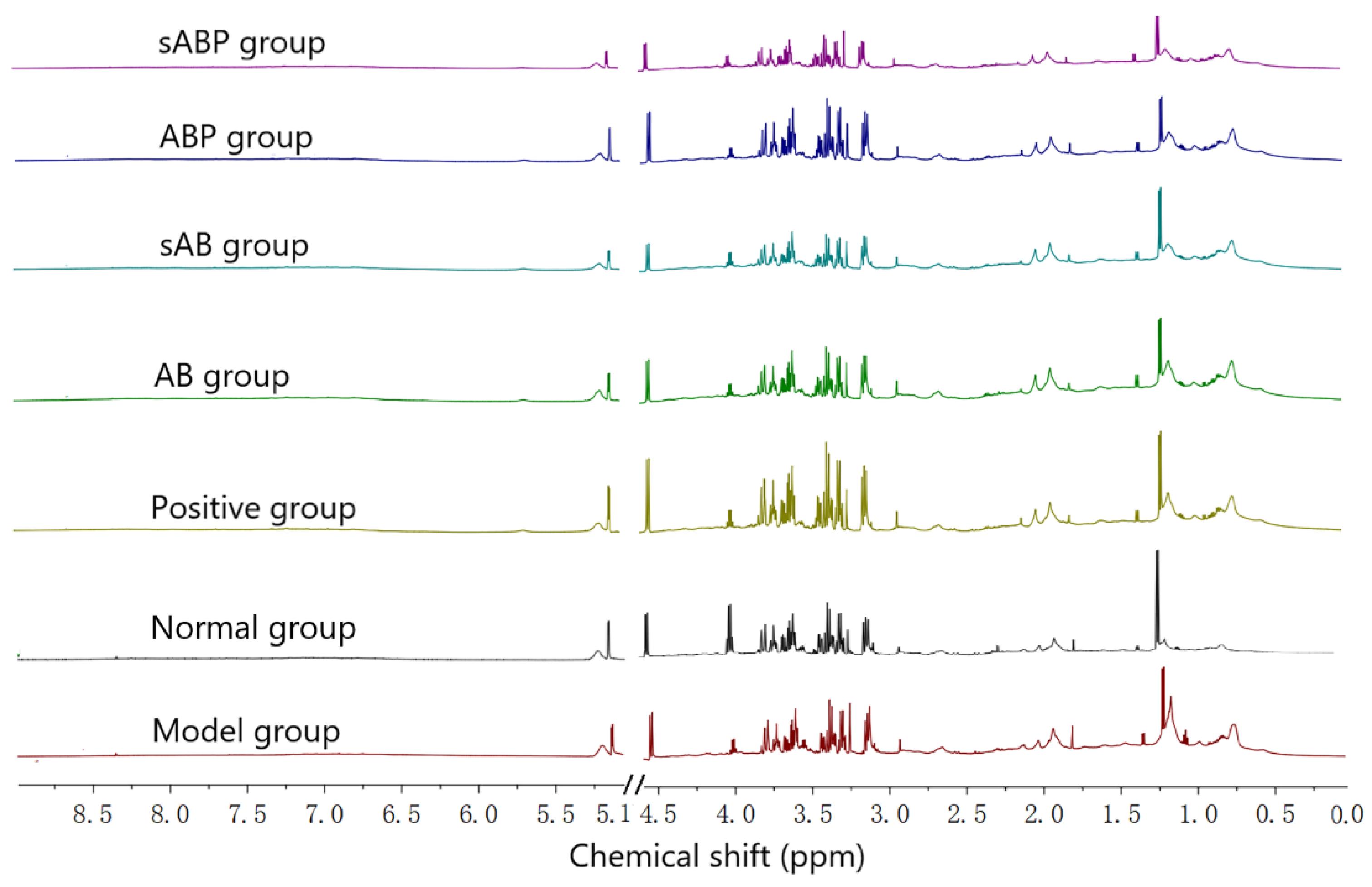
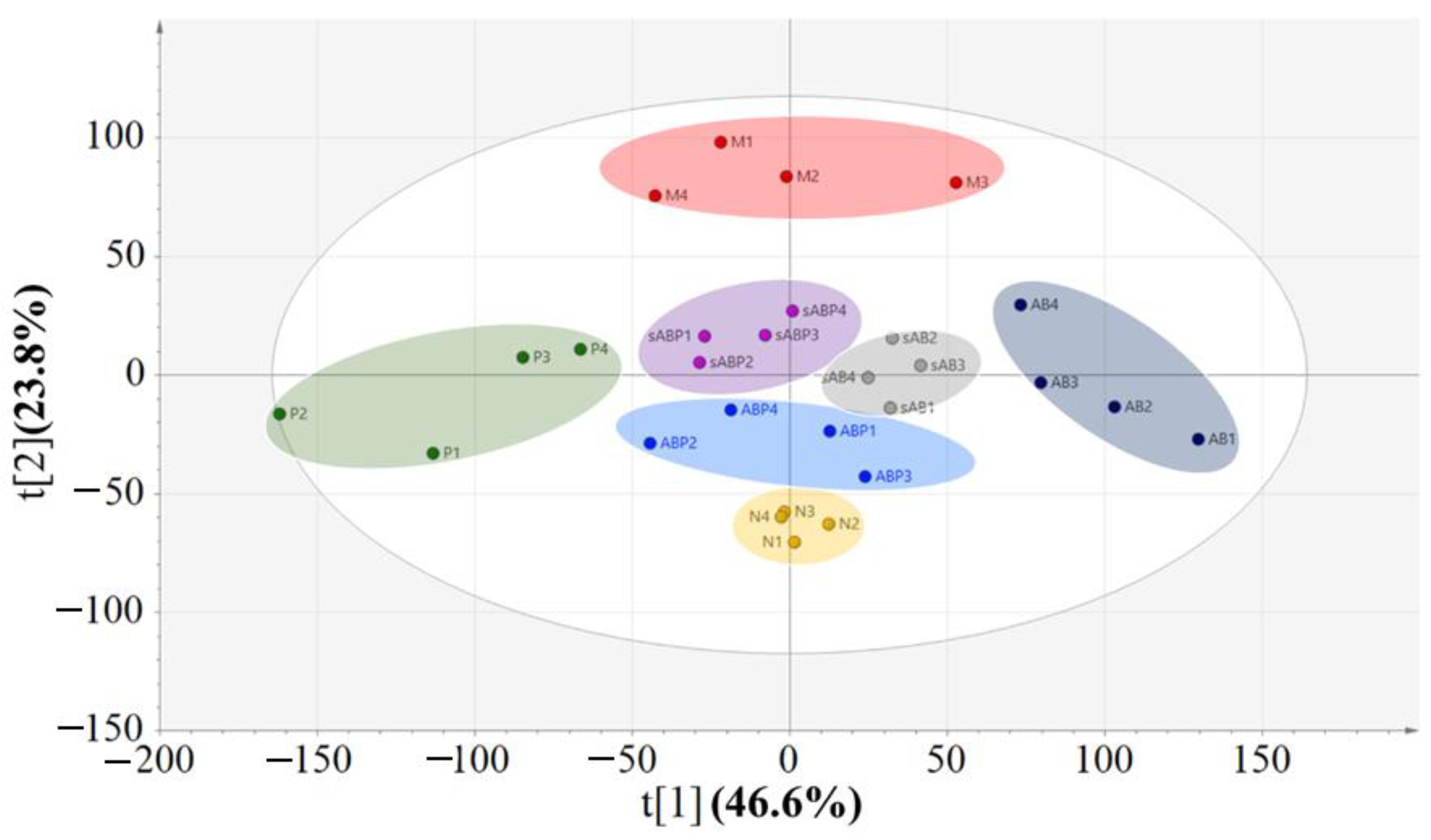
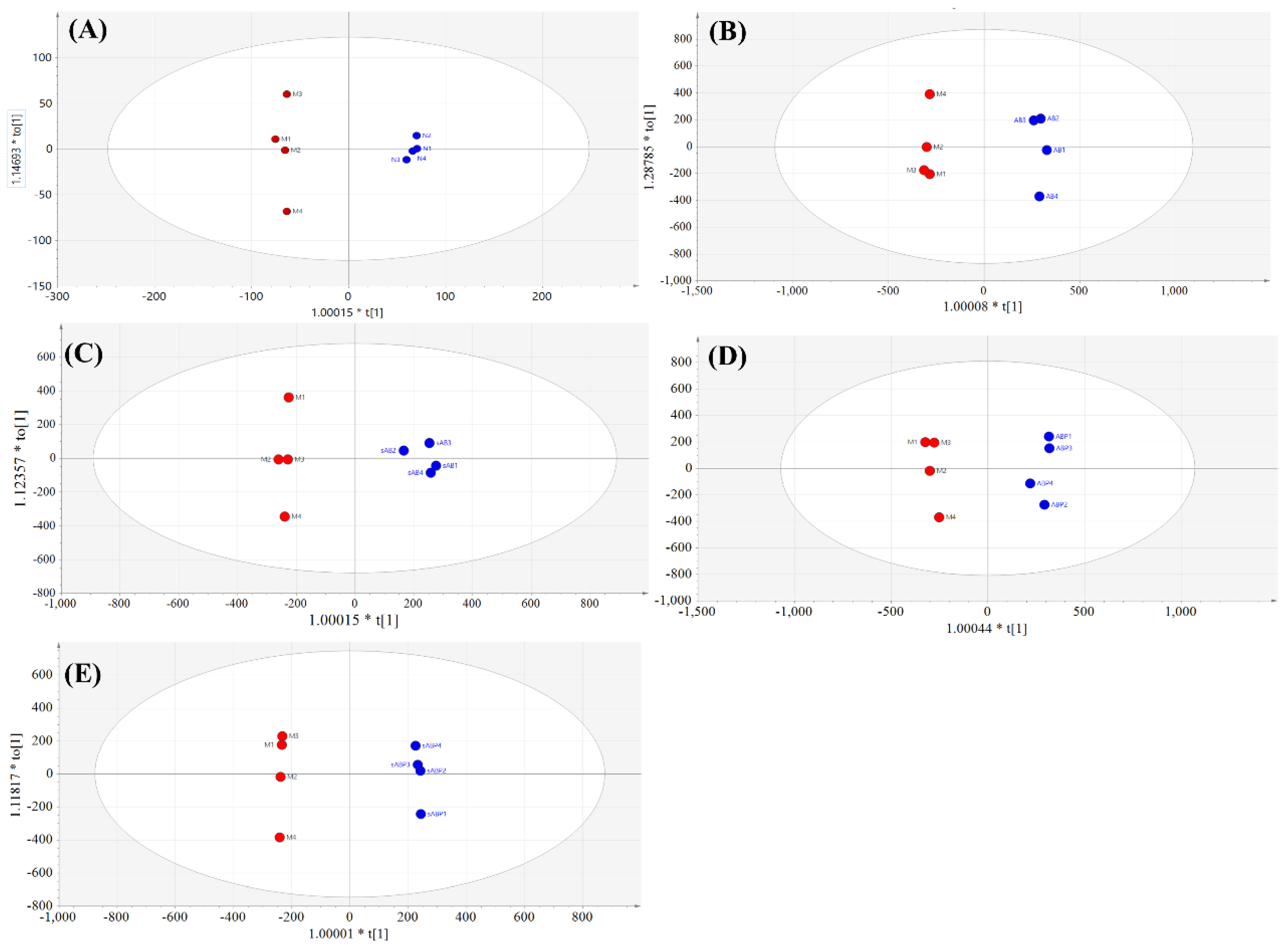
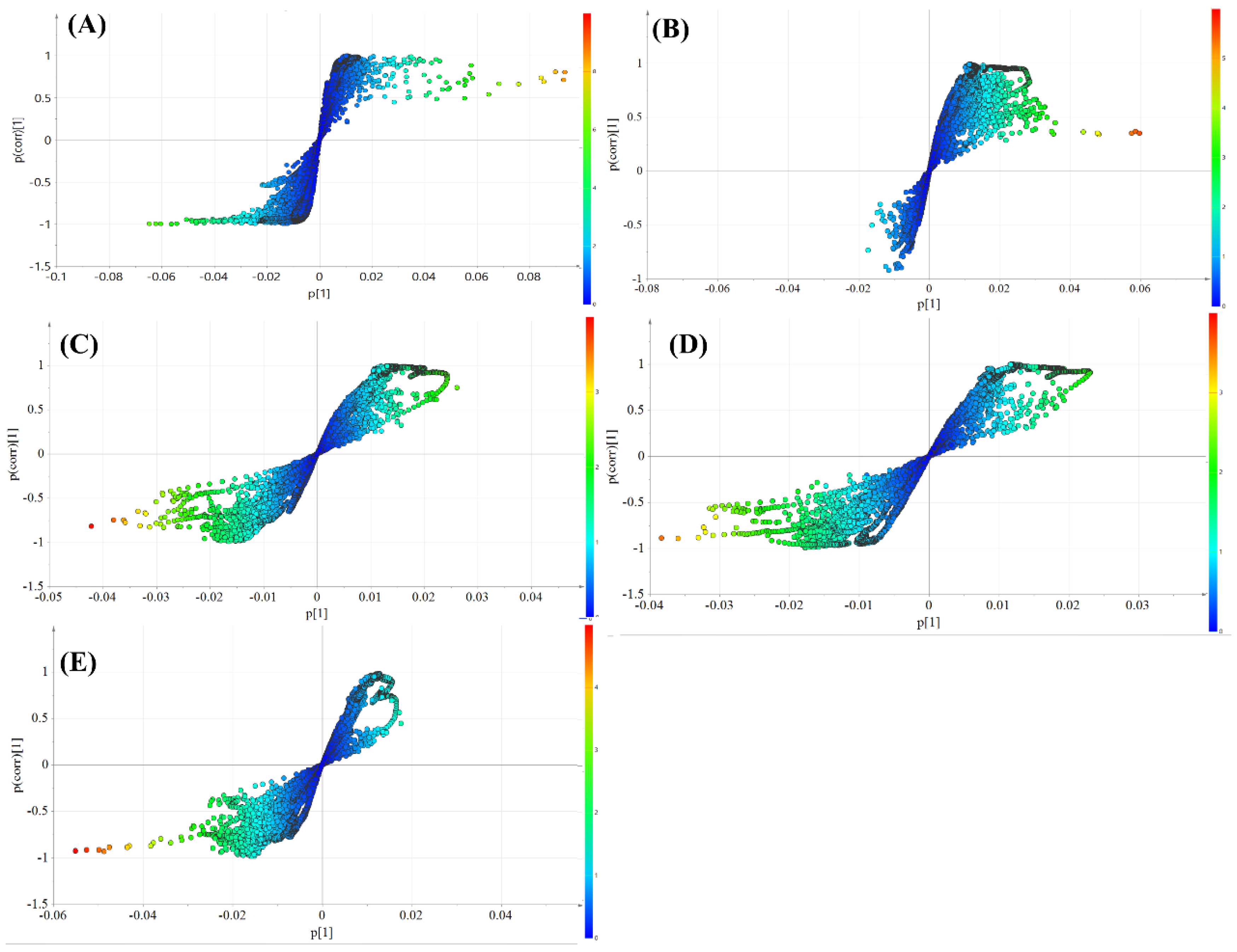
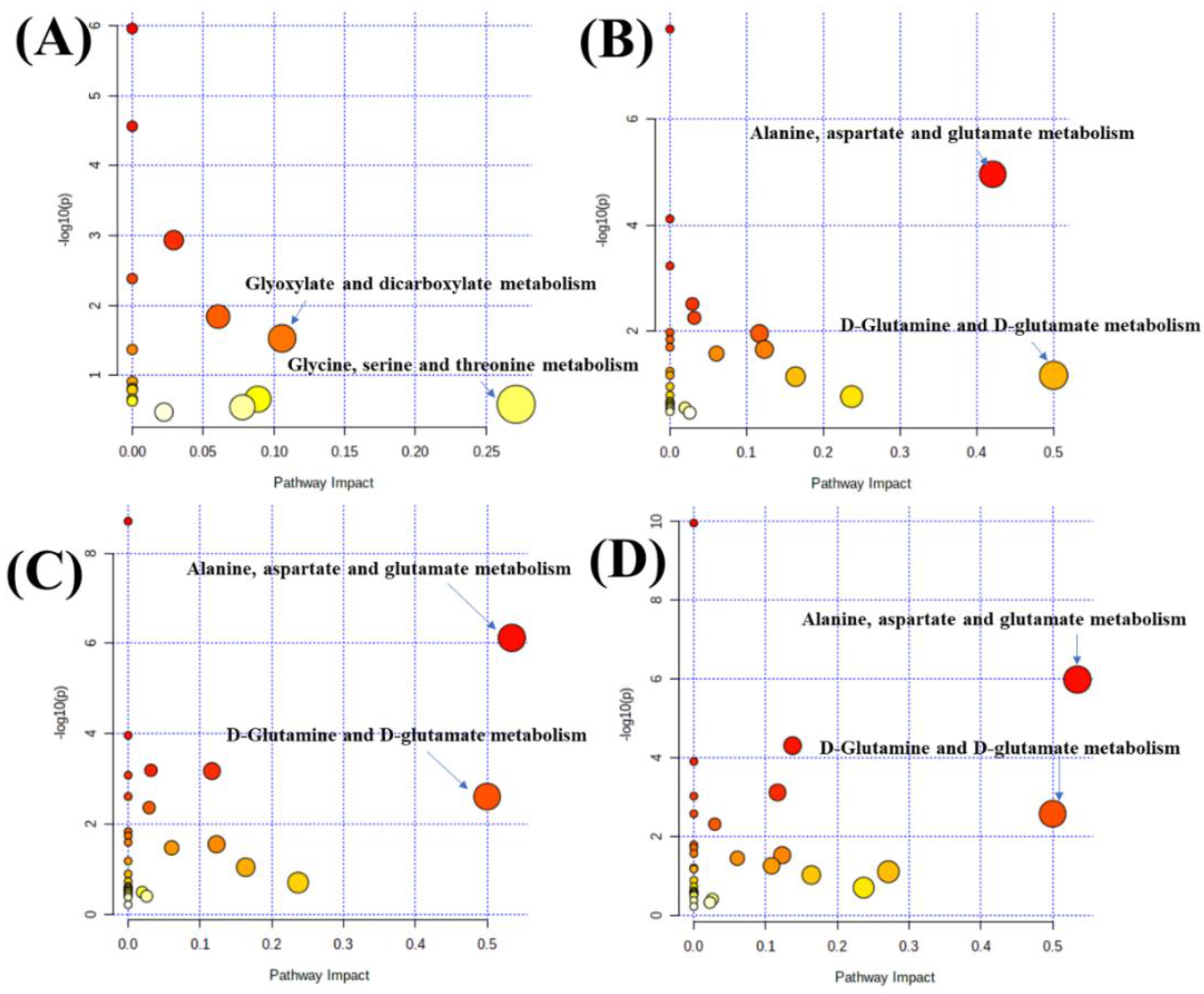
3.4. Metabolic Profile Analysis of Four Components from Crude and Salt-Processed AB
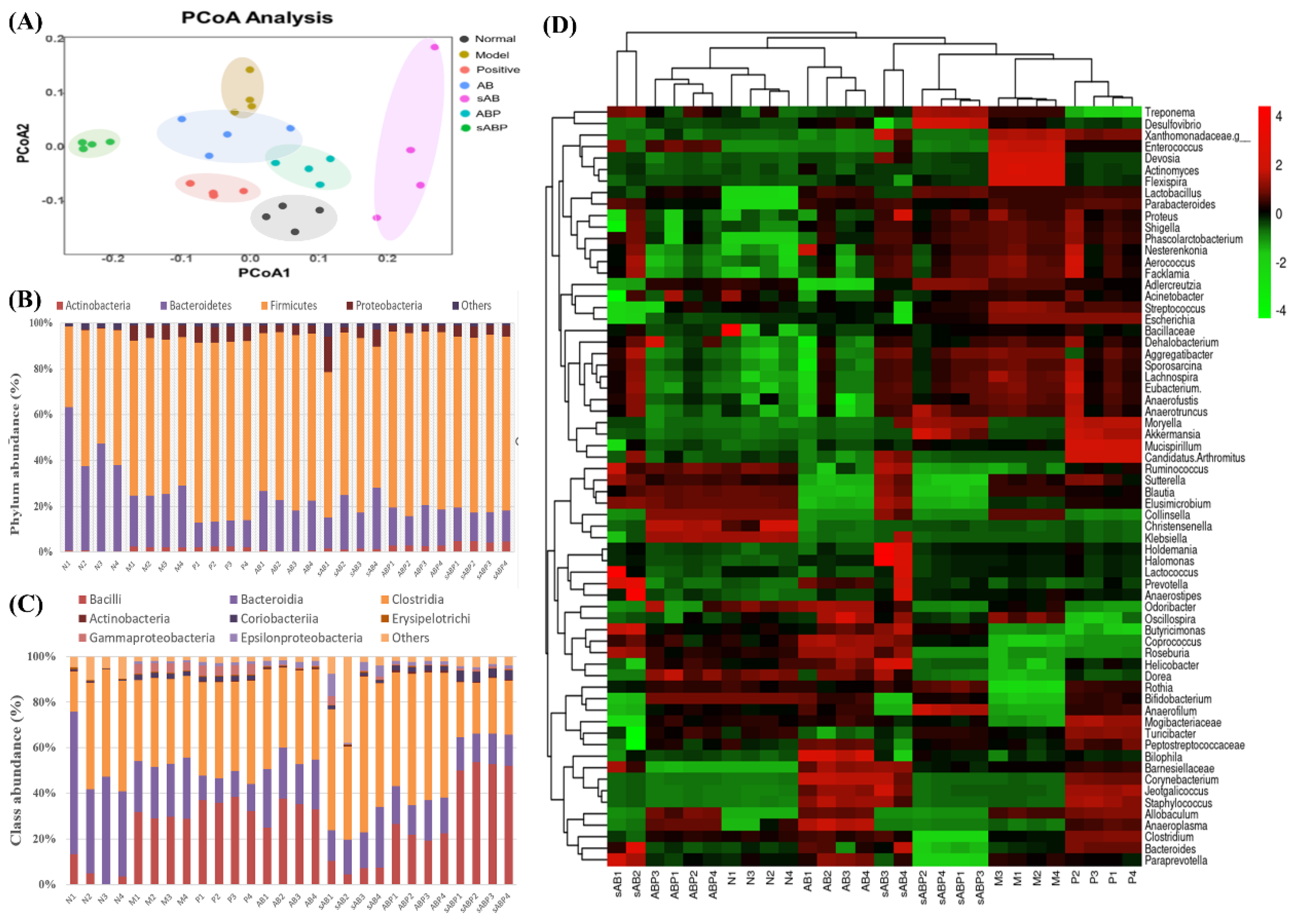
3.5. Gut Microbiota Composition of Different Components-Treated Groups
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fang, E.F.; Xie, C.; Schenkel, J.A.; Wu, C.; Long, Q.; Cui, H.; Aman, Y.; Frank, J.; Liao, J.; Zou, H.; et al. A research agenda for ageing in China in the 21st century (2nd edition): Focusing on basic and translational research, long-term care, policy and social networks. Ageing Res. Rev. 2020, 64, 101174. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.L.; Shen, H.; Liu, A.; Dong, S.S.; Zhang, L.; Deng, F.Y.; Zhao, Q.; Deng, H.W. A road map for understanding molecular and genetic determinants of osteoporosis. Nat. Rev. Endocrinol. 2020, 16, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Z. Ovarian Aging and Osteoporosis. Adv. Exp. Med. Biol. 2018, 1086, 199–215. [Google Scholar]
- Starling, S. Gene linked to adverse effects of osteoporosis drug. Nat. Rev. Endocrinol. 2020, 16, 402–403. [Google Scholar] [CrossRef]
- Zhuo, Y.; Li, M.; Jiang, Q.; Ke, H.; Liang, Q.; Zeng, L.F.; Fang, J. Evolving Roles of Natural Terpenoids from Traditional Chinese Medicine in the Treatment of Osteoporosis. Front. Endocrinol. 2022, 13, 901545. [Google Scholar] [CrossRef]
- He, X.; Wang, X.; Fang, J.; Chang, Y.; Ning, N.; Guo, H.; Huang, L.; Huang, X. The genus Achyranthes: A review on traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2017, 203, 260–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, Y.; Zhang, Q.; Wang, C.; Zhang, D.; Wan, J.B.; Yan, C. UPLC/Q-TOF-MS-based metabolomics study of the anti-osteoporosis effects of Achyranthes bidentata polysaccharides in ovariectomized rats. Int. J. Biol. Macromol. 2018, 112, 433–441. [Google Scholar] [CrossRef]
- Kiuchi, F. Saponin constituents of Achyranthes root. J. Nat. Med. 2022, 76, 343–351. [Google Scholar] [CrossRef]
- Tao, Y.; Ni, J.; Li, W.; Cai, B. Integrated response surface methodology and UHPLC coupled with triple quadrupole time-of-flight MS quantitation to investigate the salt-processing chemistry of traditional Chinese medicines: A case study on Achyranthes bidentata. Sep. Sci. Plus 2018, 1, 439–445. [Google Scholar] [CrossRef]
- Tao, Y.; Du, Y.; Li, W.; Cai, B. Development and validation of an UHPLC-MS/MS approach for simultaneous quantification of five bioactive saponins in rat plasma: Application to a comparative pharmacokinetic study of aqueous extracts of raw and salt-processed Achyranthes bidentata. J. Pharm. Biomed. Anal. 2018, 151, 164–169. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, M.; Li, B.; Kan, Y.; Zhang, B.; Zheng, X.; Feng, W. Raw and salt-processed Achyranthes bidentata attenuate LPS-induced acute kidney injury by inhibiting ROS and apoptosis via an estrogen-like pathway. Biomed. Pharmacother. 2020, 129, 110403. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, Y.; Chen, W.; Liu, C.; Li, X.; Sun, D.; Liu, Z.; Xu, Y.; Mao, X.; Guo, Q.; et al. Achyranthes bidentata extract exerts osteoprotective effects on steroid-induced osteonecrosis of the femoral head in rats by regulating RANKL/RANK/OPG signaling. J. Transl. Med. 2014, 12, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Li, Y.; Xiao, W.; Peng, L.; Wang, L.; Liao, Z.; Hu, L. Achyranthes bidentata extract protects chondrocytes functions through suppressing glycolysis and apoptosis via MAPK/AKT signaling axis. Am. J. Transl. Res. 2020, 12, 142–152. [Google Scholar] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, H.; Wu, X.; Li, D.; Huang, J. An NMR metabolomics investigation of perturbations after treatment with Chinese herbal medicine formula in an experimental model of sepsis. Omics J. Integrat. Biol. 2013, 17, 252–258. [Google Scholar] [CrossRef]
- Wei, L.; Xue, R.; Zhang, P.; Wu, Y.; Li, X.; Pei, F. 1H NMR-Based Metabolomics and Neurotoxicity Study of Cerebrum and Cerebellum in Rats Treated with Cinnabar, a Traditional Chinese Medicine. Omics J. Integrat. Biol. 2015, 19, 490–498. [Google Scholar] [CrossRef]
- Seely, K.D.; Kotelko, C.A.; Douglas, H.; Bealer, B.; Brooks, A.E. The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. Int. J. Mol. Sci. 2021, 22, 9452. [Google Scholar] [CrossRef]
- He, J.; Xu, S.; Zhang, B.; Xiao, C.; Chen, Z.; Si, F.; Fu, J.; Lin, X.; Zheng, G.; Yu, G.; et al. Gut microbiota and metabolite alterations associated with reduced bone mineral density or bone metabolic indexes in postmenopausal osteoporosis. Aging 2020, 12, 8583–8604. [Google Scholar] [CrossRef]
- Chevalier, C.; Kieser, S.; Çolakoğlu, M.; Hadadi, N.; Brun, J.; Rigo, D.; Suárez-Zamorano, N.; Spiljar, M.; Fabbiano, S.; Busse, B.; et al. Warmth Prevents Bone Loss through the Gut Microbiota. Cell Metab. 2020, 32, 575–590. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, X.; Ou, S.; Arowolo, M.A.; Hou, D.X.; He, J. Effects of Achyranthes bidentata Polysaccharides on Intestinal Morphology, Immune Response, and Gut Microbiome in Yellow Broiler Chickens Challenged with Escherichia coli K88. Polymers 2018, 10, 1233. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Chen, X.; Li, W.; Cai, B.; Di, L.; Shi, L.; Hu, L. Global and untargeted metabolomics evidence of the protective effect of different extracts of Dipsacus asper Wall. ex C.B. Clarke on estrogen deficiency after ovariectomia in rats. J. Ethnopharmacol. 2017, 199, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Gao, M.; Bao, B.; Cao, Y.; Cheng, F.; Zhang, L.; Li, Z.; Shan, J.; Yao, W. Metabolomics-driven of relationships among kidney, bone marrow and bone of rats with postmenopausal osteoporosis. Bone 2022, 156, 116306. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Heiden, M.G.V. Aerobic Glycolysis: Meeting the Metabolic Requirements of Cell Proliferation. Annu. Rev. Cell Dev. Bl. 2011, 27, 441–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, J.; He, C.; Cui, L.; Zhang, Z.; Wang, W.; Tan, L.; Liu, D.; Zheng, W.; Gu, W.; Xia, N. Discovery of Potential Biomarkers for Postmenopausal Osteoporosis Based on Untargeted GC/LC-MS. Front. Endocrinol. 2022, 13, 849076. [Google Scholar] [CrossRef]
- Qin, Q.; Yan, S.; Yang, Y.; Chen, J.; Yan, H.; Li, T.; Gao, X.; Wang, Y.; Li, A.; Wang, S.; et al. The Relationship between Osteoporosis and Intestinal Microbes in the Henan Province of China. Front. Cell. Dev. Biol. 2021, 9, 752990. [Google Scholar] [CrossRef]
- Shi, Z.; Zou, S.; Shen, Z.; Luan, F.; Yan, J. High-throughput metabolomics using UPLC/Q-TOF-MS coupled with multivariate data analysis reveals the effect and mechanism of syringin against ovariectomized osteoporosis. J. Chromatogr. B 2021, 1183, 122957. [Google Scholar] [CrossRef]
- Shen, L.; Yu, Y.; Zhou, Y.; Pruett-Miller, S.M.; Zhang, G.F.; Karner, C.M. SLC38A2 provides proline to fulfill unique synthetic demands arising during osteoblast differentiation and bone formation. eLife 2022, 11, e76963. [Google Scholar] [CrossRef]
- Si, Z.; Zhou, S.; Shen, Z.; Luan, F. High-Throughput Metabolomics Discovers Metabolic Biomarkers and Pathways to Evaluating the Efficacy and Exploring Potential Mechanisms of Osthole against Osteoporosis Based on UPLC/Q-TOF-MS Coupled with Multivariate Data Analysis. Front. Pharmacol. 2020, 11, 741. [Google Scholar] [CrossRef]
- Wan, X.; Eguchi, A.; Fujita, Y.; Ma, L.; Wang, X.; Yang, Y.; Qu, Y.; Chang, L.; Zhang, J.; Mori, C.; et al. Effects of (R)-ketamine on reduced bone mineral density in ovariectomized mice: A role of gut microbiota. Neuropharmacology 2022, 213, 109139. [Google Scholar] [CrossRef]
- Xie, H.; Hua, Z.; Guo, M.; Lin, S.; Zhou, Y.; Weng, Z.; Wu, L.; Chen, Z.; Xu, Z.; Li, W. Gut microbiota and metabonomics used to explore the mechanism of Qing’e Pills in alleviating osteoporosis. Pharma. Biol. 2022, 60, 785–800. [Google Scholar] [CrossRef]
- Hao, Y.M.; He, D.W.; Gao, Y.; Fang, L.N.; Zhang, P.P.; Lu, K.; Lu, R.Z.; Li, C. Association of Hydrogen Sulfide with Femoral Bone Mineral Density in Osteoporosis Patients: A Preliminary Study. Med. Sci. Monit. 2021, 27, e929389. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Model Group | E2-Treated Group | AB-Treated Group | sAB-Treated Group | ABP-Treated Group | sABP-Treated Group |
|---|---|---|---|---|---|---|
| S-Ca (mM) | 2.50 ± 0.07 | 2.47 ± 0.03 | 2.52 ± 0.05 | 2.51 ± 0.05 | 2.54 ± 0.04 | 2.47 ± 0.04 |
| S-P (mM) | 1.58 ± 0.07 | 1.64 ± 0.11 | 1.60 ± 0.09 | 1.64 ± 0.07 | 1.62 ± 0.10 | 1.67 ± 0.11 |
| U-Ca/Cr (mmol/mmol) | 0.37 ± 0.01 | 0.20 ± 0.02 ** | 0.30 ± 0.02 ** | 0.29 ± 0.01 ** | 0.27 ± 0.01 ** | 0.25 ± 0.01 ** |
| U-P/Cr (mmol/mmol) | 4.76 ± 0.11 | 3.74 ± 0.17 ** | 4.41 ± 0.15 ** | 4.49 ± 0.09 ** | 4.20 ± 0.25 ** | 4.13 ± 0.24 ** |
| ALP (U/L) | 268.90 ± 9.11 | 129.92 ± 7.19 ** | 217.67 ± 14.65 ** | 210.36 ± 14.29 ** | 181.78 ± 11.47 ** | 173.48 ± 16.67 ** |
| Parameters | Model | E2 | AB | sAB | ABP | sABP |
|---|---|---|---|---|---|---|
| BMD (g/cm3) | 0.348 ± 0.011 | 0.768 ± 0.039 ** | 0.459 ± 0.013 ** | 0.401 ± 0.012 ** | 0.490 ± 0.014 ** | 0.543 ± 0.016 ** |
| BV/TV | 8.292 ± 0.331 | 43.345 ± 0.527 ** | 16.405 ± 0.857 ** | 13.174 ± 0.818 ** | 17.903 ± 0.587 ** | 23.402 ± 0.560 ** |
| Tb.Th (mm) | 0.079 ± 0.005 | 0.102 ± 0.004 ** | 0.080 ± 0.006 | 0.079 ± 0.002 | 0.080 ± 0.007 | 0.077 ± 0.004 |
| TB.N (1/mm) | 1.051 ± 0.050 | 4.178 ± 0.063 ** | 2.024 ± 0.039 ** | 1.590 ± 0.041 ** | 2.098 ± 0.061 ** | 2.799 ± 0.055 ** |
| Tb.Sp (mm) | 1.033 ± 0.029 | 0.137 ± 0.008 ** | 0.451 ± 0.054 ** | 0.501 ± 0.037 ** | 0.436 ± 0.013 ** | 0.250 ± 0.036 ** |
| Parameters | Model Group | E2-Treated Group | AB-Treated Group | sAB-Treated Group | ABP-Treated Group | sABP-Treated Group |
|---|---|---|---|---|---|---|
| Maximum load (N) | 103.12 ± 1.94 | 120.32 ± 1.68 ** | 109.61 ± 3.77 ** | 108.47 ± 0.90 ** | 112.86 ± 1.24 ** | 115.20 ± 3.84 ** |
| Stiffness (N/mm) | 144.97 ± 3.93 | 159.55 ± 3.87 ** | 151.80 ± 1.72 ** | 153.03 ± 4.15 ** | 155.06 ± 0.90 ** | 156.09 ± 1.51 ** |
| Energy (N × mm) | 39.78 ± 0.59 | 54.93 ± 3.29 ** | 44.63 ± 1.96 ** | 42.37 ± 1.40 ** | 46.59 ± 2.27 ** | 50.42 ± 1.28 ** |
| Maximum stress (MPa) | 147.88 ± 1.29 | 202.37 ± 3.70 ** | 153.51 ± 2.91 ** | 152.22 ± 4.10 ** | 159.60 ± 4.67 ** | 165.60 ± 6.35 ** |
| Young modulus (MPa) | 4450.53 ± 52.09 | 6580.19 ± 102.41 ** | 5675.56 ± 88.10 ** | 5153.65 ± 103.23 ** | 5795.70 ± 63.07 ** | 5914.54 ± 69.61 ** |
| Metabolites | δ(1H) | AB-Treated Group vs. Model Group | sAB-Treated Group vs. Model Group | ABP-Treated Group vs. Model Group | sABP-Treated Group vs. Model Group | ||||
|---|---|---|---|---|---|---|---|---|---|
| VIP | Change/Fold Change | VIP | Change/Fold Change | VIP | Change/Fold Change | VIP | Change/Fold Change | ||
| Lipid | 0.83–0.89 (bra) | 2.58 | ↑*/1.33 | 2.23 | ↑*/1.27 | 2.11 | ↑*/1.33 | 1.51 | ↑*/1.16 |
| Leucine | 0.96 (t), 1.70 (m), 3.73 (m) | 1.27 | ↓*/1.13 | 1.29 | ↑*/1.12 | 1.20 | ↓*/0.88 | 1.33 | ↓*/0.88 |
| Isoleucine | 1.02 (d), 3.73 (m) | 1.23 | ↓*/0.87 | 1.09 | ↓*/0.89 | 1.02 | ↓*/0.87 | 1.40 | ↓*/0.86 |
| Valine | 1.05 (d), 3.61 (d) | 1.39 | ↑*/1.21 | 1.33 | ↓*/0.77 | 1.42 | ↓*/0.67 | 1.52 | ↓*/0.73 |
| 3-Hydroxybutyrate | 1.20 (d), 2.28 (q),2.40 (q), 4.15 (m) | 1.05 | ↑*/1.30 | 1.62 | ↓*/0.78 | 1.34 | ↓*/0.77 | 1.56 | ↓*/0.79 |
| Lactate | 1.33 (d), 4.12 (q) | 5.52 | ↑*/2.04 | 3.94 | ↓*/0.87 | 1.12 | ↓*/0.82 | 4.98 | ↓*/0.79 |
| Alanine | 1.48 (d), 3.78 (q) | 1.62 | ↑*/1.36 | 2.03 | ↑*/1.31 | 1.34 | ↑*/1.21 | 1.41 | ↓*/0.84 |
| Acetate | 1.92 (s) | 1.29 | ↓*/0.83 | 2.69 | ↓*/0.72 | 1.67 | ↓*/0.80 | 1.67 | ↓*/0.81 |
| Glutamate | 2.08 (m), 2.34 (m) | — | — | 1.22 | ↓*/0.89 | 1.48 | ↓*/0.82 | 1.89 | ↓*/0.81 |
| Glutamine | 2.13 (m), 2.45 (m) | — | — | — | — | 1.22 | ↓*/0.72 | 1.61 | ↓*/0.84 |
| Succinate | 2.37 (s) | — | — | 1.48 | ↓*/0.81 | 1.45 | ↓*/0.73 | 1.71 | ↓*/0.77 |
| Citrate | 2.54 (d), 2.66 (d) | — | — | 1.74 | ↓*/0.66 | 1.66 | ↓*/0.56 | 1.37 | ↓*/0.70 |
| Aspartate | 2.87 (m), 2.94 (m) | — | — | 1.21 | ↓*/0.77 | 1.29 | ↓*/0.64 | 1.35 | ↓*/0.72 |
| Choline | 3.20 (s) | — | — | 1.82 | ↓*/0.78 | 1.60 | ↓*/0.72 | 1.47 | ↓*/0.78 |
| Proline | 3.36 (m) | 1.61 | ↓*/0.80 | 1.70 | ↓*/0.83 | 1.82 | ↓*/0.73 | 2.18 | ↓*/0.75 |
| Glycine | 3.57 (s) | 1.03 | ↑*/1.27 | — | — | — | — | 1.04 | ↓*/0.83 |
| Glucose | 3.24 (q), 3.48 (t), 3.90 (q), 3.54 (t), 3.71 (t), 3.83 (t) | 3.05 | ↑*/1.45 | 1.18 | ↓*/0.89 | 1.36 | ↑*/1.15 | 1.65 | ↓*/0.81 |
| Serine | 3.84 (m), 3.96 (m) | 2.47 | ↑*/1.46 | 1.43 | ↑*/1.17 | 1.55 | ↑*/1.22 | 1.21 | ↑*/1.14 |
| NAc | 2.03 (s) | 1.25 | ↑*/1.13 | — | — | 1.34 | ↓*/0.88 | 1.74 | ↓*/0.87 |
| PUFA | 2.76–2.83 (bra) | — | — | 1.17 | ↓*/0.78 | 1.23 | ↓*/0.76 | 1.02 | ↓*/0.86 |
| Glycerol | 3.61 (m), 3.65 (m) | — | — | 2.32 | ↓*/0.70 | 1.23 | ↓*/0.73 | 1.48 | ↓*/0.72 |
| Lysine | 1.45 (m), 1.71 (m), 1.89 (m), 3.02 (t), 3.75 (t) | — | — | 2.51 | ↓*/0.64 | 2.20 | ↑*/1.33 | 2.62 | ↓*/0.62 |
| Microbiota | r |
|---|---|
| Anaerofilum | 0.754 ** |
| Bifidobacterium | 0.556 ** |
| Blautia | −0.413 ** |
| Devosia | −0.469 ** |
| Lactobacillus | 0.465 ** |
| Mogibacteriaceae | 0.680 ** |
| Odoribacter | −0.565 ** |
| Oscillospira | −0.705 ** |
| Paraprevotella | −0.428 ** |
| Peptostreptococcaceae | 0.557 ** |
| Rothia | 0.743 ** |
| Turicibacter | 0.729 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Y.; Zhu, F.; Pan, M.; Bao, J.; Liu, Q.; Tao, Y. A Multi-Omics Analysis Reveals Anti-Osteoporosis Mechanism of Four Components from Crude and Salt-Processed Achyranthes bidentata Blume in Ovariectomized Rats. Molecules 2022, 27, 5012. https://doi.org/10.3390/molecules27155012
Yin Y, Zhu F, Pan M, Bao J, Liu Q, Tao Y. A Multi-Omics Analysis Reveals Anti-Osteoporosis Mechanism of Four Components from Crude and Salt-Processed Achyranthes bidentata Blume in Ovariectomized Rats. Molecules. 2022; 27(15):5012. https://doi.org/10.3390/molecules27155012
Chicago/Turabian StyleYin, Yuwen, Fei Zhu, Meiling Pan, Jiaqi Bao, Qing Liu, and Yi Tao. 2022. "A Multi-Omics Analysis Reveals Anti-Osteoporosis Mechanism of Four Components from Crude and Salt-Processed Achyranthes bidentata Blume in Ovariectomized Rats" Molecules 27, no. 15: 5012. https://doi.org/10.3390/molecules27155012