
The Anti-Inflammatory Activities of Fermented Curcuma That Contains Butyrate Mitigate DSS-Induced Colitis in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Chemicals and Reagents
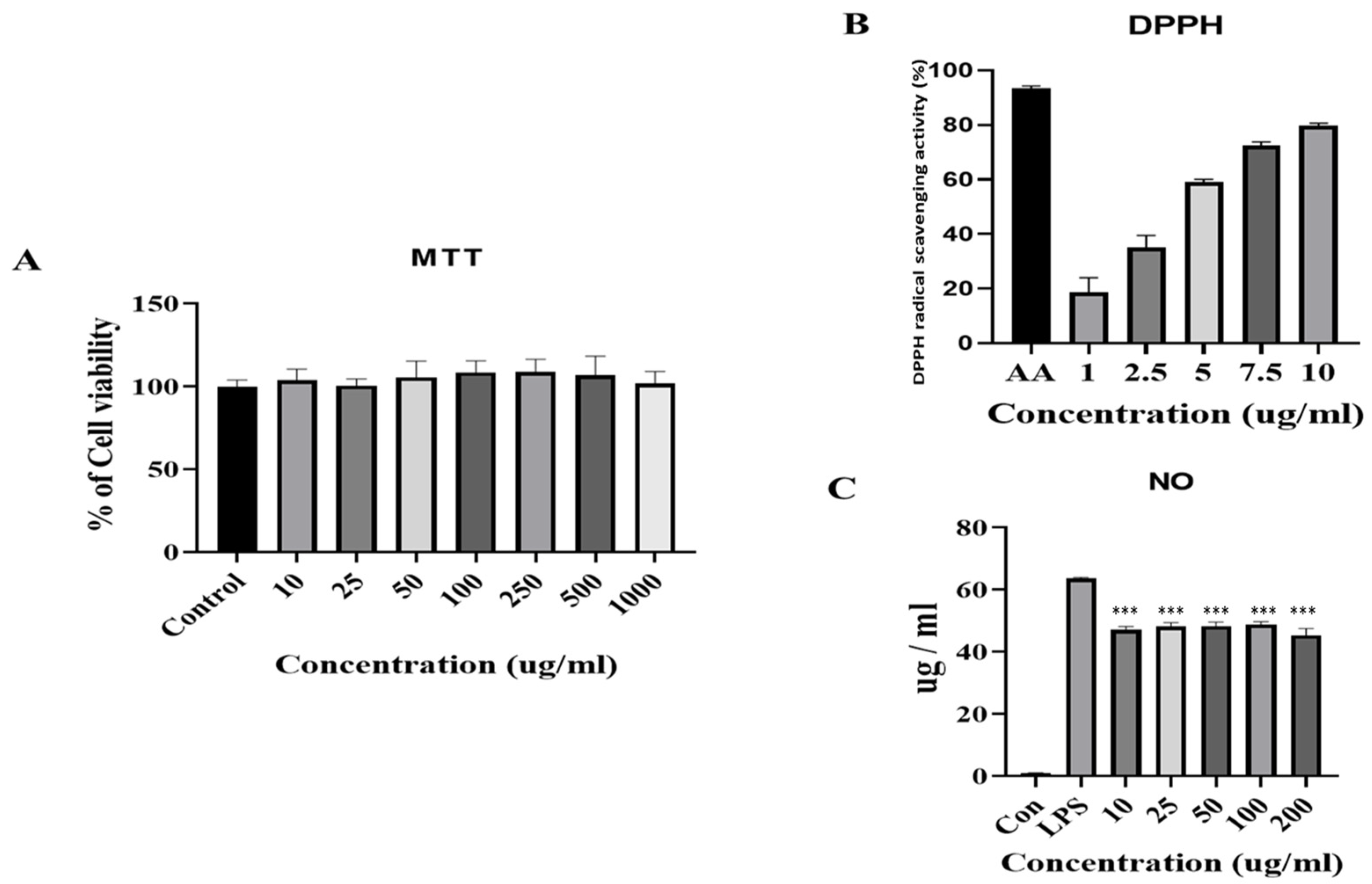
2.3. DPPH Radical Scavenging Activities
2.4. Cell Viability
2.5. Determination of Nitric Oxide (NO) Production
2.6. Animals
2.7. Measurement of Body Weight and Gut Length
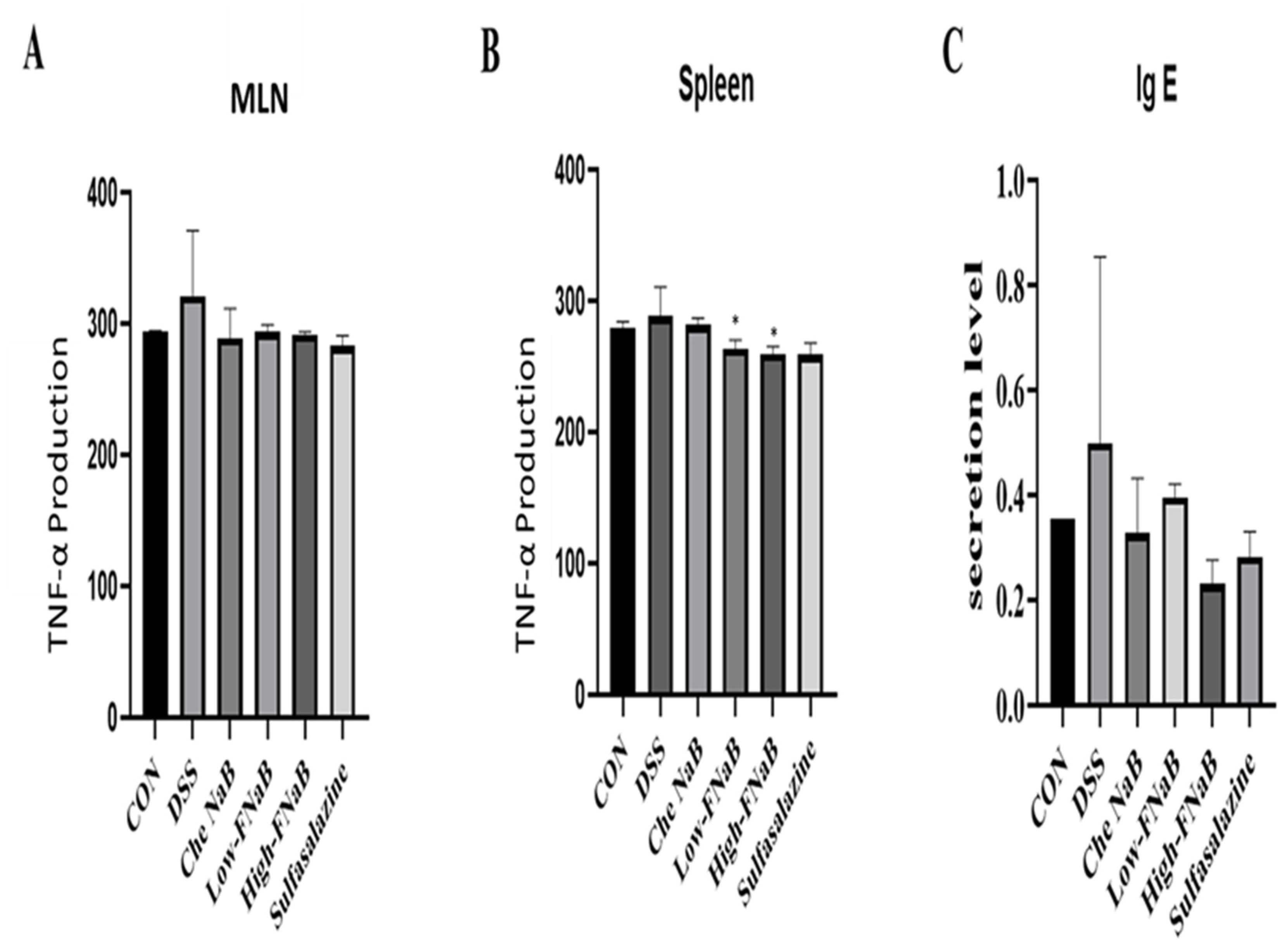
2.8. Determination of TNF-α
2.9. Serum IgE Analysis
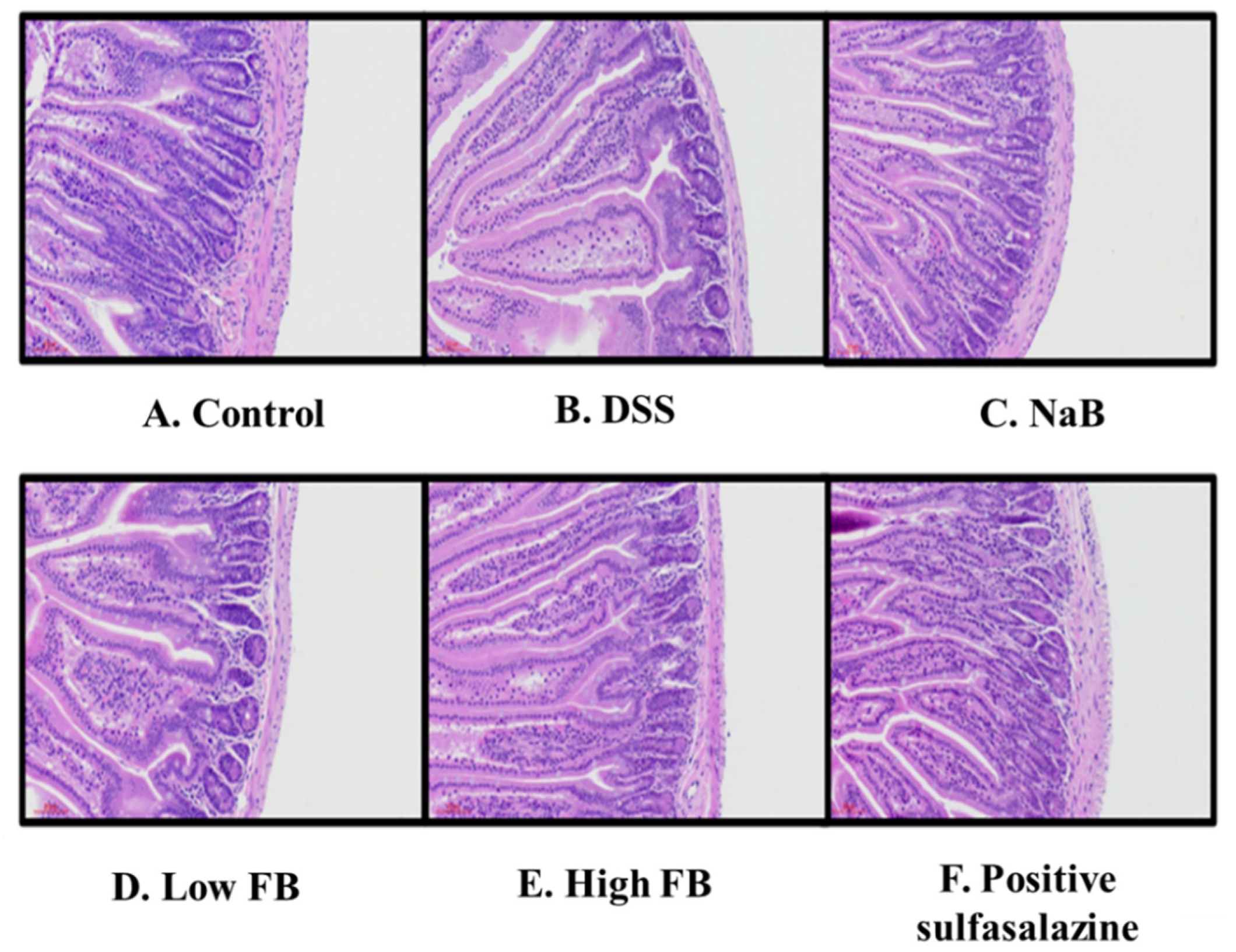
2.10. Hematoxylin and Eosin (H&E) Staining of Colon Tissue
2.11. Statistical Analysis
3. Results
3.1. Gas Chromatography and Mass Spectrometry (GC/MS) of FB
3.2. Antioxidant and Anti-Inflammatory Activities of FB and the Effect on Cell Viability
3.3. The Effect of FB in DSS-Induced Colitis Mice
3.4. TNF-α and IgE Release in DSS-Induced Colitis Mice
3.5. Histology of Large Intestine in DSS-Induced Colitis Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability Statement
References
- Dalal, S.R.; Chang, E.B. Chang The microbial basis of inflammatory bowel diseases. J. Clin. Investig. 2014, 124, 4190–4196. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; de la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Steidler, L.; Hans, W.; Schotte, L.; Neirynck, S.; Obermeier, F.; Falk, W.; Fiers, W.; Remaut, E. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 2000, 289, 1352–1355. [Google Scholar] [CrossRef] [Green Version]
- Bayazid, A.B.; Kim, J.G.; Azam, S.; Jeong, S.A.; Kim, D.H.; Park, C.W.; Lim, B.O. Sodium butyrate ameliorates neurotoxicity and exerts anti-inflammatory effects in high fat diet-fed mice. Food Chem. Toxicol. 2022, 159, 112743. [Google Scholar] [CrossRef]
- Bayazid, A.B.; Jang, Y.A. The Role of Andrographolide on Skin Inflammations and Modulation of Skin Barrier Functions in Human Keratinocyte. Biotechnol. Bioprocess Eng. 2021, 26, 804–813. [Google Scholar] [CrossRef]
- Podosky, D. Inflammatory bowel disease (first of two parts). N. Engl. J. Med. 1991, 325, 928–937. [Google Scholar] [CrossRef]
- Bayazid, A.B.; Chun, E.M.; Al Mijan, M.; Park, S.H.; Moon, S.-K.; Lim, B.O. Anthocyanins profiling of bilberry (Vaccinium myrtillus L.) extract that elucidates antioxidant and anti-inflammatory effects. Food Agric. Immunol. 2021, 32, 713–726. [Google Scholar] [CrossRef]
- Bayazid, A.B.; Jang, Y.A.; Jeong, S.A.; Lim, B.O. Cypress tree (Chamaecyparis obtusa) Bark extract inhibits melanogenesis through repressing CREB and MITF signalling pathways in α-MSH-stimulated B16F10 cells. Food Agric. Immunol. 2022, 33, 498–510. [Google Scholar] [CrossRef]
- Williams, L.; Bradley, L.; Smith, A.; Foxwell, B. Signal transducer and activator of transcription 3 is the dominant mediator of the anti-inflammatory effects of IL-10 in human macrophages. J. Immunol. 2004, 172, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. A J. Tech. Methods Pathol. 1993, 69, 238–249. [Google Scholar]
- Dignass, A.; Preiss, J.C.; Aust, D.E.; Autschbach, F.; Ballauff, A.; Barretton, G.; Bokemeyer, B.; Fichtner-Feigl, S.; Hagel, S.; Herrlinger, K.R.; et al. Updated German guideline on diagnosis and treatment of ulcerative colitis, 2011. Z. Fur Gastroenterol. 2011, 49, 1276–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-Chain Fatty Acids Suppress Lipopolysaccharide-Induced Production of Nitric Oxide and Proinflammatory Cytokines Through Inhibition of NF-κB Pathway in RAW264.7 Cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Yuandani; Jantan, I.; Rohani, A.S.; Sumantri, I.B. Immunomodulatory Effects and Mechanisms of Curcuma Species and Their Bioactive Compounds: A Review. Front. Pharmacol. 2021, 12, 643119. [Google Scholar] [CrossRef] [PubMed]
- Colin, T.; Bories, A.; Lavigne, C.; Moulin, G. Effects of Acetate and Butyrate During Glycerol Fermentation by Clostridium butyricum. Curr. Microbiol. 2001, 43, 238–243. [Google Scholar] [CrossRef]
- Purwani, E.Y.; Purwadaria, T.; Suhartono, M.T. Fermentation RS3 derived from sago and rice starch with Clostridium butyricum BCC B2571 or Eubacterium rectale DSM 17629. Anaerobe 2012, 18, 55–61. [Google Scholar] [CrossRef]
- Barreto, M.J.d.S. Current Overview of the Role of Short-Chain Fatty Acids in Prevention or Therapy of Colorectal Carcinoma. Master’s Thesis, Universidade do Minho-Campus de Gualtar, Minho, Portugal, 2013. [Google Scholar]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Cao, S.; Feehley, T.J.; Nagler, C.R. The role of commensal bacteria in the regulation of sensitization to food allergens. FEBS Lett. 2014, 588, 4258–4266. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, F.; Olijnyk, O.R.; Tilley, C.J.; Tamaoki, T. Interactions between dextran sulfate and Escherichia coli ribosomes. Biochim. Et Biophys. Acta (BBA)-Nucleic Acids Protein Synth. 1967, 145, 96–104. [Google Scholar] [CrossRef]
- Silva, J.P.B.; Navegantes-Lima, K.C.; de Oliveira, A.L.B.; Rodrigues, D.V.S.; Gaspar, S.L.F.; Monteiro, V.V.S.; Moura, D.P.; Monteiro, M.C. Protective Mechanisms of Butyrate on Inflammatory Bowel Disease. Curr. Pharm. Des. 2018, 24, 4154–4166. [Google Scholar] [CrossRef]
- Tsang, S.W.; Ip, S.P.; Wu, J.C.-Y.; Ng, S.-C.; Yung, K.K.-L.; Bian, Z.-X. A Chinese medicinal formulation ameliorates dextran sulfate sodium-induced experimental colitis by suppressing the activity of nuclear factor-kappaB signaling. J. Ethnopharmacol. 2015, 162, 20–30. [Google Scholar] [CrossRef]
- Atreya, R.; Zimmer, M.; Bartsch, B.; Waldner, M.J.; Atreya, I.; Neumann, H.; Hildner, K.; Hoffman, A.; Kiesslich, R.; Rink, A.D.; et al. Antibodies Against Tumor Necrosis Factor (TNF) Induce T-Cell Apoptosis in Patients With Inflammatory Bowel Diseases via TNF Receptor 2 and Intestinal CD14+ Macrophages. Gastroenterology 2011, 141, 2026–2038. [Google Scholar] [CrossRef] [PubMed]
- Powrie, F.; Leach, M.W.; Mauze, S.; Menon, S.; Caddle, L.B.; Coffman, R.L. Inhibition of Thl responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity 1994, 1, 553–562. [Google Scholar] [CrossRef]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The Immunology of Mucosal Models of Inflammation. Annu. Rev. Immunol. 2002, 20, 495–549. [Google Scholar] [CrossRef] [PubMed]
- Blando, F.; Calabriso, N.; Berland, H.; Maiorano, G.; Gerardi, C.; Carluccio, M.A.; Andersen, Ø.M. Radical Scavenging and Anti-Inflammatory Activities of Representative Anthocyanin Groupings from Pigment-Rich Fruits and Vegetables. Int. J. Mol. Sci. 2018, 19, 169. [Google Scholar] [CrossRef] [Green Version]
- Leong, H.Y.; Show, P.L.; Lim, M.H.; Ooi, C.W.; Ling, T.C. Natural red pigments from plants and their health benefits: A review. Food Rev. Int. 2017, 34, 463–482. [Google Scholar] [CrossRef]
- Chang, L.; Neu, J. Early Factors Leading to Later Obesity: Interactions of the Microbiome, Epigenome, and Nutrition. Curr. Probl. Pediatric Adolesc. Health Care 2015, 45, 134–142. [Google Scholar] [CrossRef]
- Bayazid, A.B.; Jang, Y.A.; Kim, Y.M.; Kim, J.G.; Lim, B.O. Neuroprotective Effects of Sodium Butyrate through Suppressing Neuroinflammation and Modulating Antioxidant Enzymes. Neurochem. Res. 2021, 46, 2348–2358. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Content |
|---|---|
| Butyrate | 0.19 g per 100 g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayazid, A.B.; Jeong, S.A.; Park, C.W.; Kim, D.H.; Lim, B.O. The Anti-Inflammatory Activities of Fermented Curcuma That Contains Butyrate Mitigate DSS-Induced Colitis in Mice. Molecules 2022, 27, 4745. https://doi.org/10.3390/molecules27154745
Bayazid AB, Jeong SA, Park CW, Kim DH, Lim BO. The Anti-Inflammatory Activities of Fermented Curcuma That Contains Butyrate Mitigate DSS-Induced Colitis in Mice. Molecules. 2022; 27(15):4745. https://doi.org/10.3390/molecules27154745
Chicago/Turabian StyleBayazid, Al Borhan, Soo Ah Jeong, Chae Won Park, Da Hee Kim, and Beong Ou Lim. 2022. "The Anti-Inflammatory Activities of Fermented Curcuma That Contains Butyrate Mitigate DSS-Induced Colitis in Mice" Molecules 27, no. 15: 4745. https://doi.org/10.3390/molecules27154745