
Determination of Free Amino Acids in Milk, Colostrum and Plasma of Swine via Liquid Chromatography with Fluorescence and UV Detection

 , , and
, , and
Abstract
:1. Introduction
2. Results
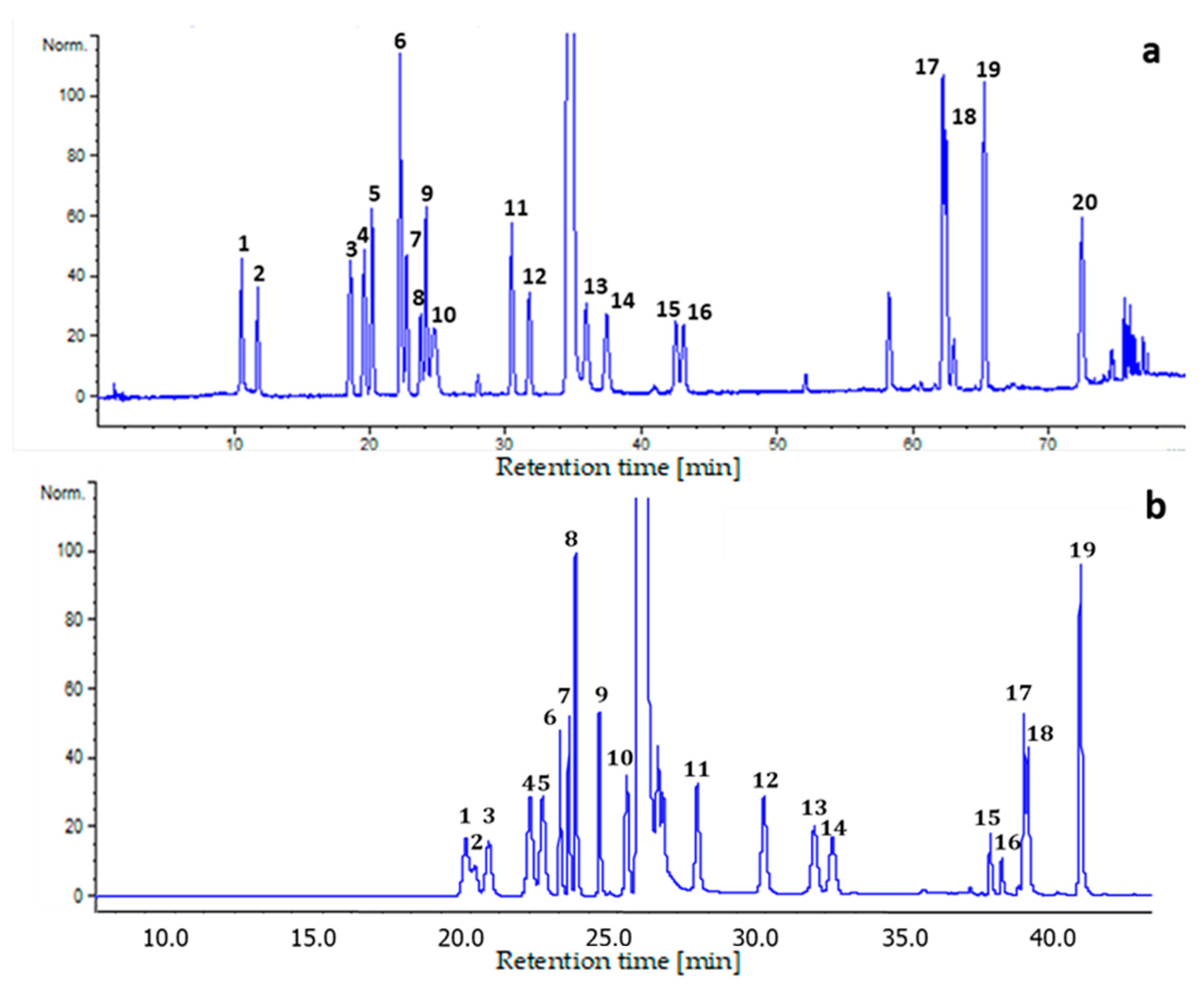
2.1. Chromatographic Conditions Optimization
2.2. Optimization of the Derivatization Reaction
2.3. Method Validation
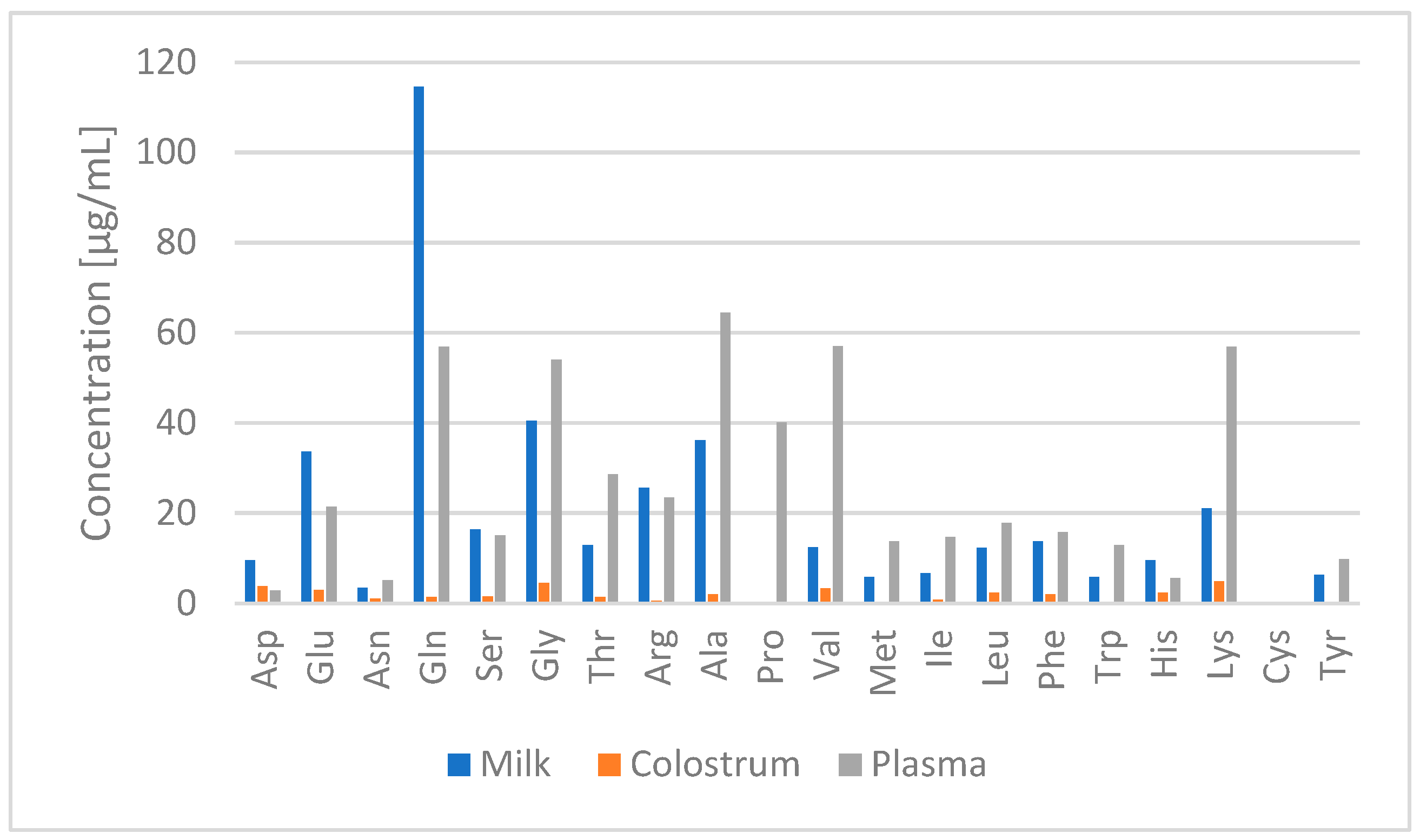
2.4. Application to Real Samples
3. Discussion
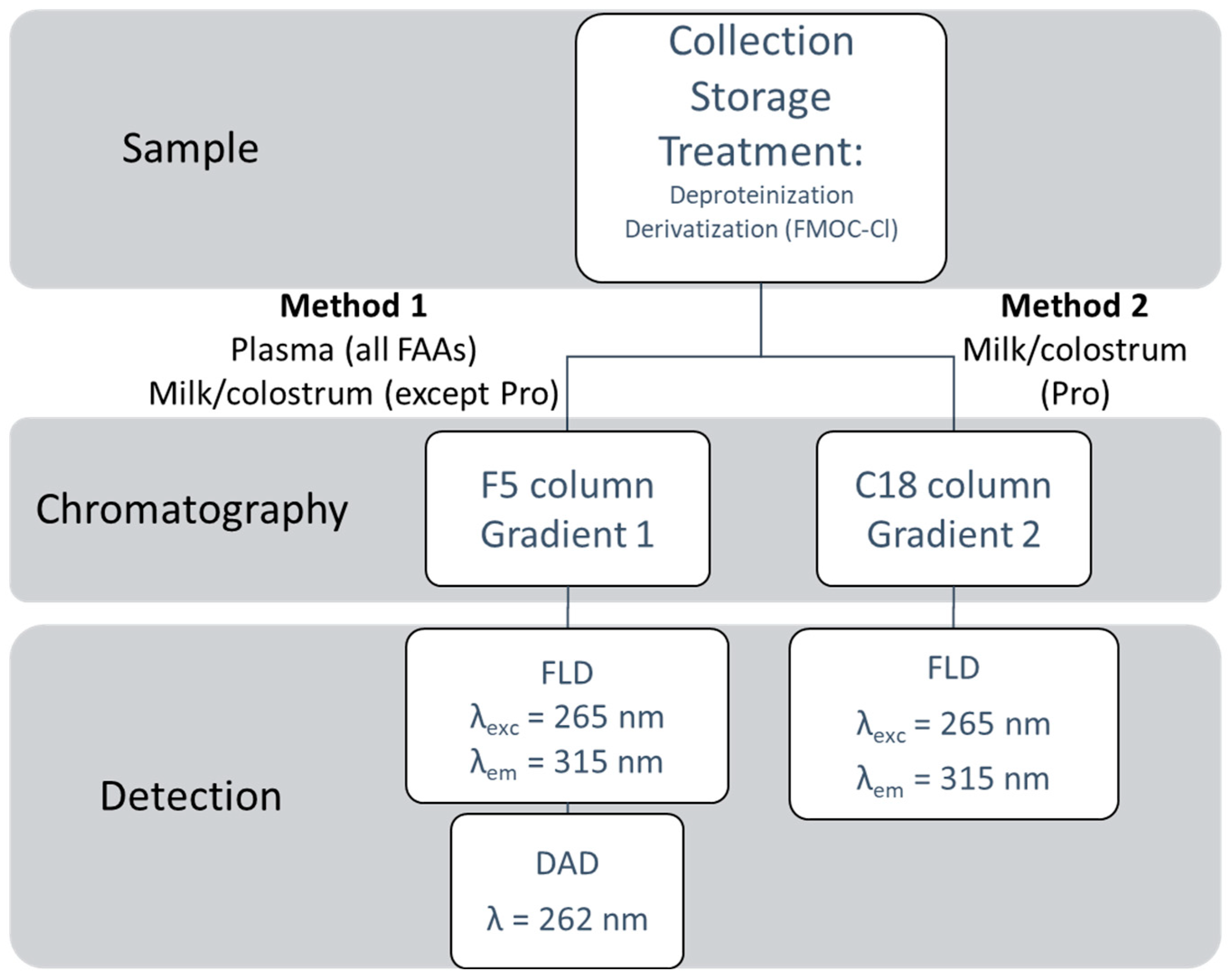
4. Materials and Methods
4.1. Chemicals and Solutions
4.2. Sample Collection
4.3. Standard Solutions
4.4. Sample Preparation
4.5. Derivatization Reaction
4.6. HPLC-UV/FLD Conditions
4.7. Method Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yin, B.; Li, T.; Zhang, S.; Li, Z.; He, P. Sensitive Analysis of 33 Free Amino Acids in Serum, Milk, and Muscle by Ultra-High Performance Liquid Chromatography-Quadrupole-Orbitrap High Resolution Mass Spectrometry. Food Anal. Methods 2016, 9, 2814–2823. [Google Scholar] [CrossRef]
- Wu, G. Functional Amino Acids in Nutrition and Health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.; Fang, L.H.; Jeong, J.H.; Kim, Y.Y. Effects of L-Arginine Supplementation during Late Gestation on Reproductive Performance, Piglet Uniformity, Blood Profiles, and Milk Composition in High Prolific Sows. Animals 2020, 10, 1313. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yue, Z.; Ao, Z.; Zhao, C.; Shi, J.; Zhao, C.; Zeng, F.; Cai, G.; Zheng, E.; Yang, J.; et al. Maternal Dietary Supplementation of Arginine Increases the Ratio of Total Cloned Piglets Born to Total Transferred Cloned Embryos by Improving the Pregnancy Rate of Recipient Sows. Anim. Reprod. Sci. 2018, 196, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Csapó, J.; Martin, T.G.; Csapó-Kiss, Z.S.; Házas, Z. Protein, Fats, Vitamin and Mineral Concentrations in Porcine Colostrum and Milk from Parturition to 60 Days. Int. Dairy J. 1996, 6, 881–902. [Google Scholar] [CrossRef]
- Bertocchi, M.; Bosi, P.; Luise, D.; Motta, V.; Salvarani, C.; Ribani, A.; Bovo, S.; Simongiovanni, A.; Matsunaga, K.; Takimoto, T.; et al. Dose-Response of Different Dietary Leucine Levels on Growth Performance and Amino Acid Metabolism in Piglets Differing for Aminoadipate-Semialdehyde Synthase Genotypes. Sci. Rep. 2019, 9, 18496. [Google Scholar] [CrossRef]
- Floc’h, N.L.; Gondret, F.; Matte, J.J.; Quesnel, H. Towards Amino Acid Recommendations for Specific Physiological and Patho-Physiological States in Pigs. Proc. Nutr. Soc. 2012, 71, 425–432. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, F.; Zhang, Y.; Lv, Y.; Heng, J.; Min, T.; Li, L.; Guan, W. Recent Progress of Porcine Milk Components and Mammary Gland Function. J. Anim. Sci. Biotechnol. 2018, 9, 77. [Google Scholar] [CrossRef]
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino Acids and Mammary Gland Development: Nutritional Implications for Milk Production and Neonatal Growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Luise, D.; Correa, F.; Fusco, L.; Bosi, P.; Trevisi, P. Productive Effects of a Colostrum-Oriented Amino Acid Dietary Supply for Sows in Transition from Gestation to Lactation. Ital. J. Anim. Sci. 2021, 20, 1837–1850. [Google Scholar] [CrossRef]
- Le Boucher, J.; Charret, C.; Coudray-Lucas, C.; Giboudeau, J.; Cynober, L. Amino Acid Determination in Biological Fluids by Automated Ion-Exchange Chromatography: Performance of Hitachi L-8500A. Clin. Chem. 1997, 43, 1421–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Martín, E.; Ruiz, J.; Pérez-Palacios, T.; Silva, A.; Antequera, T. Gas Chromatography–Mass Spectrometry Method for the Determination of Free Amino Acids as Their Dimethyl- Tert -Butylsilyl (TBDMS) Derivatives in Animal Source Food. J. Agric. Food Chem. 2012, 60, 2456–2463. [Google Scholar] [CrossRef]
- Kaspar, H.; Dettmer, K.; Gronwald, W.; Oefner, P.J. Automated GC–MS Analysis of Free Amino Acids in Biological Fluids. J. Chromatogr. B 2008, 870, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Leggio, A.; Belsito, E.L.; De Marco, R.; Liguori, A.; Siciliano, C.; Spinella, M. Simultaneous Extraction and Derivatization of Amino Acids and Free Fatty Acids in Meat Products. J. Chromatogr. A 2012, 1241, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, M.P.; Dudzik, D.; Varas, E.; Gibellini, M.; Skotnicki, M.; Zorawski, M.; Zarzycki, W.; Pellati, F.; García, A. Optimization and Validation of a Chiral GC–MS Method for the Determination of Free d-Amino Acids Ratio in Human Urine: Application to a Gestational Diabetes Mellitus Study. J. Pharm. Biomed. Anal. 2015, 107, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.L.; Roberts, W.L.; Pasquali, M. Analysis of Plasma Amino Acids by HPLC with Photodiode Array and Fluorescence Detection. Clin. Chim. Acta 2005, 354, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Fürst, P.; Pollack, L.; Graser, T.A.; Godel, H.; Stehle, P. Appraisal of Four Pre-Column Derivatization Methods for the High-Performance Liquid Chromatographic Determination of Free Amino Acids in Biological Materials. J. Chromatogr. 1990, 499, 557–569. [Google Scholar] [CrossRef]
- Fierabracci, V.; Masiello, P.; Novelli, M.; Bergamini, E. Application of Amino Acid Analysis by High-Performance Liquid Chromatography with Phenyl Isothiocyanate Derivatization to the Rapid Determination of Free Amino Acids in Biological Samples. J. Chromatogr. B Biomed. Sci. Appl. 1991, 570, 285–291. [Google Scholar] [CrossRef]
- Dai, Z.; Wu, Z.; Jia, S.; Wu, G. Analysis of Amino Acid Composition in Proteins of Animal Tissues and Foods as Pre-Column o-Phthaldialdehyde Derivatives by HPLC with Fluorescence Detection. J. Chromatogr. B 2014, 964, 116–127. [Google Scholar] [CrossRef]
- Cohen, S.A.; Bidlingmeyer, B.A.; Tarvin, T.L. PITC Derivatives in Amino Acid Analysis. Nature 1986, 320, 769–770. [Google Scholar] [CrossRef]
- Cigić, I.; Vodošek, T.V.; Kosmerl, T.; Strlič, M. Amino Acid Quantification in the Presence of Sugars Using HPLC and Pre-Column Derivatization with 3-MPA/OPA and FMOC-Cl. Acta Chim. Slov. 2008, 55, 660–664. [Google Scholar]
- Turiák, G.; Volicer, L. Stability of O-Phthalaldehyde-Sulfite Derivatives of Amino Acids and Their Methyl Esters: Electrochemical and Chromatographic Properties. J. Chromatogr. A 1994, 668, 323–329. [Google Scholar] [CrossRef]
- Heinrikson, R.L.; Meredith, S.C. Amino Acid Analysis by Reverse-Phase High-Performance Liquid Chromatography: Precolumn Derivatization with Phenylisothiocyanate. Anal. Biochem. 1984, 136, 65–74. [Google Scholar] [CrossRef]
- Einarsson, S.; Josefsson, B.; Lagerkvist, S. Determination of Amino Acids with 9-Fluorenylmethyl Chloroformate and Reversed-Phase High-Performance Liquid Chromatography. J. Chromatogr. A 1983, 282, 609–618. [Google Scholar] [CrossRef]
- Jámbor, A.; Molnár-Perl, I. Amino Acid Analysis by High-Performance Liquid Chromatography after Derivatization with 9-Fluorenylmethyloxycarbonyl Chloride Literature Overview and Further Study. J. Chromatogr. A 2009, 1216, 3064–3077. [Google Scholar] [CrossRef]
- Zanker, I.A.; Hammon, H.M.; Blum, J.W. Plasma Amino Acid Pattern during the First Month of Life in Calves Fed the First Colostrum at 0-2 h or at 24-25 h after Birth. J. Vet. Med. Ser. A 2000, 47, 107–121. [Google Scholar] [CrossRef]
- Samaei, S.P.; Ghorbani, M.; Tagliazucchi, D.; Martini, S.; Gotti, R.; Themelis, T.; Tesini, F.; Gianotti, A.; Gallina Toschi, T.; Babini, E. Functional, Nutritional, Antioxidant, Sensory Properties and Comparative Peptidomic Profile of Faba Bean (Vicia faba, L.) Seed Protein Hydrolysates and Fortified Apple Juice. Food Chem. 2020, 330, 127120. [Google Scholar] [CrossRef]
- Chuang, C.-K.; Lin, S.-P.; Lee, H.-C.; Wang, T.-J.; Shih, Y.-S.; Huang, F.-Y.; Yeung, C.-Y. Free Amino Acids in Full-Term and Pre-Term Human Milk and Infant Formula. J. Pediatric Gastroenterol. Nutr. 2005, 40, 496–500. [Google Scholar] [CrossRef]
- Mudd, A.T.; Alexander, L.S.; Johnson, S.K.; Getty, C.M.; Malysheva, O.V.; Caudill, M.A.; Dilger, R.N. Perinatal Dietary Choline Deficiency in Sows Influences Concentrations of Choline Metabolites, Fatty Acids, and Amino Acids in Milk throughout Lactation. J. Nutr. 2016, 146, 2216–2223. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Y.; Cao, M.; Li, Y.; Tang, L.; Li, W.; Jiang, X.; Xiao, W.; Liu, S.; Jiang, X.; Fang, Z.; et al. Soybean Bioactive Peptides Supplementation during Late Gestation and Lactation Affect the Reproductive Performance, Free Amino Acid Composition in Plasma and Milk of Sows. Livest. Sci. 2020, 237, 104064. [Google Scholar] [CrossRef]
- van Sadelhoff, J.H.J.; Wiertsema, S.P.; Garssen, J.; Hogenkamp, A. Free Amino Acids in Human Milk: A Potential Role for Glutamine and Glutamate in the Protection Against Neonatal Allergies and Infections. Front. Immunol. 2020, 11, 1007. [Google Scholar] [CrossRef] [PubMed]
- Klein, K.; Bancher-Todesca, D.; Graf, T.; Garo, F.; Roth, E.; Kautzky-Willer, A.; Worda, C. Concentration of Free Amino Acids in Human Milk of Women with Gestational Diabetes Mellitus and Healthy Women. Breastfeed. Med. 2013, 8, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Ihara, T.; Furuse, M. Differences in Free Amino Acid Concentrations in Milk between Wistar and Wistar Kyoto Rats. J. Vet. Med. Sci. 2019, 81, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Knabe, D.A. Free and Protein-Bound Amino Acids in Sow’s Colostrum and Milk. J. Nutr. 1994, 124, 415–424. [Google Scholar] [CrossRef]
- How, Z.T.; Busetti, F.; Linge, K.L.; Kristiana, I.; Joll, C.A.; Charrois, J.W.A. Analysis of Free Amino Acids in Natural Waters by Liquid Chromatography-Tandem Mass Spectrometry. J. Chromatogr. A 2014, 1370, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, A.; Ng, A.; Kwan, T.; Cusmano-Ozog, K.; Cowan, T.M. A Rapid, Sensitive Method for Quantitative Analysis of Underivatized Amino Acids by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 944, 166–174. [Google Scholar] [CrossRef]
- Moldoveanu, S.C.; Zhu, J.; Qian, N. Free Amino Acids Analysis by Liquid Chromatography with Tandem Mass Spectrometry in Several Botanicals with Antioxidant Character. J. Sep. Sci. 2015, 38, 2208–2222. [Google Scholar] [CrossRef]
- Li, M.; Li, W.; Kong, F.; Kang, S.; Liang, X.; Han, H.; Wu, J.; Zheng, Y.; Li, Q.; Yue, X.; et al. Metabolomics Methods to Analyze Full Spectrum of Amino Acids in Different Domains of Bovine Colostrum and Mature Milk. Eur. Food Res. Technol. 2020, 246, 213–224. [Google Scholar] [CrossRef]
- Gałęzowska, G.; Ratajczyk, J.; Wolska, L. Determination of Amino Acids in Human Biological Fluids by High-Performance Liquid Chromatography: Critical Review. Amino Acids 2021, 53, 993–1009. [Google Scholar] [CrossRef]
- Vakili, H.; Talebpour, Z.; Haghighi, F. Development, Validation, and Uncertainty Measurement of HPLC-DAD Method for Determination of Some Free Amino Acids in Infant Formula and Medical Food Products for Inborn Errors of Metabolism. Food Chem. 2022, 390, 133204. [Google Scholar] [CrossRef]
- Kang, M.; Yue, Q.; Jia, S.; Wang, J.; Zheng, M.; Suo, R. Identification of Geographical Origin of Milk by Amino Acid Profile Coupled with Chemometric Analysis. J. Food Qual. 2022, 2022, 2001253. [Google Scholar] [CrossRef]
- He, M.; Nie, X.; Wang, H.; Yan, S.; Zhang, Y. Effects of a High-Grain Diet With a Buffering Agent on Milk Protein Synthesis in Lactating Goats. Front. Vet. Sci. 2021, 8, 696703. [Google Scholar] [CrossRef] [PubMed]
- Alu’datt, M.H.; Al-u’datt, D.G.F.; Alhamad, M.N.; Tranchant, C.C.; Rababah, T.; Gammoh, S.; Althnaibat, R.M.; Daradkeh, M.G.; Kubow, S. Characterization and Biological Properties of Peptides Isolated from Dried Fermented Cow Milk Products by RP-HPLC: Amino Acid Composition, Antioxidant, Antihypertensive, and Antidiabetic Properties. J. Food Sci. 2021, 86, 3046–3060. [Google Scholar] [CrossRef] [PubMed]
- Milan, A.M.; Samuelsson, L.M.; Shrestha, A.; Sharma, P.; Day, L.; Cameron-Smith, D. Circulating Branched Chain Amino Acid Concentrations Are Higher in Dairy-Avoiding Females Following an Equal Volume of Sheep Milk Relative to Cow Milk: A Randomized Controlled Trial. Front. Nutr. 2020, 7, 553674. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Nguyen, H.V.; Garcia-Bravo, R.; Fiorotto, M.L.; Jackson, E.M.; Reeds, P.J. Amino Acid Composition of the Milk of Some Mammalian Species Changes with Stage of Lactation. Br. J. Nutr. 1994, 72, 845–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, W.L. 9. Composition of Sow Colostrum and Milk. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, Germany, 2014; pp. 193–230. ISBN 978-90-8686-253-5. [Google Scholar]
- Gao, K.; Li, G.; Zhu, C.; Wang, L.; Yang, X.; Wen, X.; Wu, Z.; Jiang, Z. Effect of Optimizing Dietary Valine-to-Lysine Ratio in Late Gestation or Lactation on Biochemical Indices and Performance of Lactating Primiparous Sows. Anim. Feed Sci. Technol. 2019, 253, 13–21. [Google Scholar] [CrossRef]
- Richert, B.T.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; Campbell, R.G.; Kershaw, S. The Effect of Dietary Lysine and Valine Fed during Lactation on Sow and Litter Performance. J. Anim. Sci. 1997, 75, 1853–1860. [Google Scholar] [CrossRef]
- O’Quinn, P.R.; Knabe, D.A.; Wu, G. Arginine Catabolism in Lactating Porcine Mammary Tissue. J. Anim. Sci. 2002, 80, 467–474. [Google Scholar] [CrossRef]
- Bengtsson, G. Sow Colostral Free Amino Acids and Their Importance for the Newborn Piglet. Acta Vet. Scand. 1972, 13, 7–13. [Google Scholar]
- Yu, M.; Mu, C.; Yang, Y.; Zhang, C.; Su, Y.; Huang, Z.; Yu, K.; Zhu, W. Increases in Circulating Amino Acids with In-Feed Antibiotics Correlated with Gene Expression of Intestinal Amino Acid Transporters in Piglets. Amino Acids 2017, 49, 1587–1599. [Google Scholar] [CrossRef]
- Yang, Z.; Hasan, M.S.; Htoo, J.K.; Burnett, D.D.; Feugang, J.M.; Crenshaw, M.A.; Liao, S.F. Effects of Dietary Supplementation of L-Methionine vs. Dl-Methionine on Performance, Plasma Concentrations of Free Amino Acids and Other Metabolites, and Myogenesis Gene Expression in Young Growing Pigs. Transl. Anim. Sci. 2019, 3, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Cardenia, V.; Zappaterra, M.; Motta, V.; Bosi, P.; Rodriguez-Estrada, M.T.; Trevisi, P. Evaluation of Breed and Parity Order Effects on the Lipid Composition of Porcine Colostrum. J. Agric. Food Chem. 2018, 66, 12911–12920. [Google Scholar] [CrossRef] [PubMed]
- Themelis, T.; Gotti, R.; Orlandini, S.; Gatti, R. Quantitative Amino Acids Profile of Monofloral Bee Pollens by Microwave Hydrolysis and Fluorimetric High Performance Liquid Chromatography. J. Pharm. Biomed. Anal. 2019, 173, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Walfish, S. A Statistical Perspective on the ICH Q2A and Q2B Guidelines for Validation of Analytical Methods. BioPharm Int. 2006, 19, 28–36. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gradient 1 * | Gradient 2 ⊥ | |||||
|---|---|---|---|---|---|---|
| Time (min) | A% | B% | Time (min) | A% | B% | C% |
| 0 | 80 | 20 | 0 | 90 | 0 | 10 |
| 28 | 57 | 43 | 10 | 85 | 0 | 15 |
| 38 | 57 | 43 | 11 | 80 | 0 | 20 |
| 70 | 25 | 75 | 20 | 76 | 0 | 24 |
| 75 | 0 | 100 | 21 | 0 | 55 | 45 |
| 31 | 0 | 55 | 45 | |||
| 36 | 0 | 30 | 70 | |||
| 42 | 0 | 15 | 85 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gotti, R.; Esposito, E.; Luise, D.; Tullio, S.; Interino, N.; Trevisi, P.; Fiori, J. Determination of Free Amino Acids in Milk, Colostrum and Plasma of Swine via Liquid Chromatography with Fluorescence and UV Detection. Molecules 2022, 27, 4153. https://doi.org/10.3390/molecules27134153
Gotti R, Esposito E, Luise D, Tullio S, Interino N, Trevisi P, Fiori J. Determination of Free Amino Acids in Milk, Colostrum and Plasma of Swine via Liquid Chromatography with Fluorescence and UV Detection. Molecules. 2022; 27(13):4153. https://doi.org/10.3390/molecules27134153
Chicago/Turabian StyleGotti, Roberto, Erika Esposito, Diana Luise, Stefano Tullio, Nicolò Interino, Paolo Trevisi, and Jessica Fiori. 2022. "Determination of Free Amino Acids in Milk, Colostrum and Plasma of Swine via Liquid Chromatography with Fluorescence and UV Detection" Molecules 27, no. 13: 4153. https://doi.org/10.3390/molecules27134153