
Investigation Driven by Network Pharmacology on Potential Components and Mechanism of DGS, a Natural Vasoprotective Combination, for the Phytotherapy of Coronary Artery Disease

 , and
, and
Abstract
:1. Introduction
2. Results
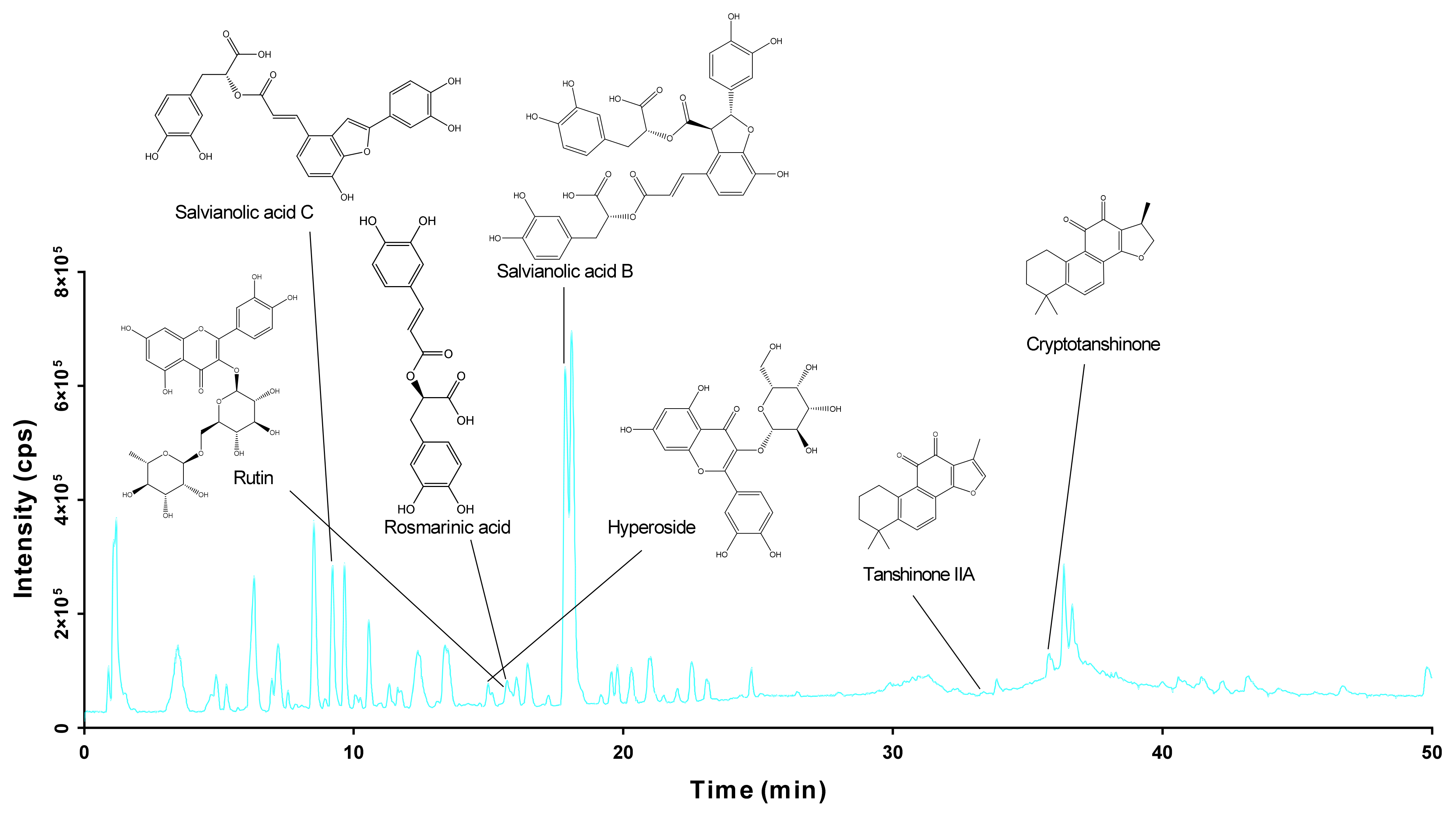
2.1. Identification of the Chemical Components of DGS
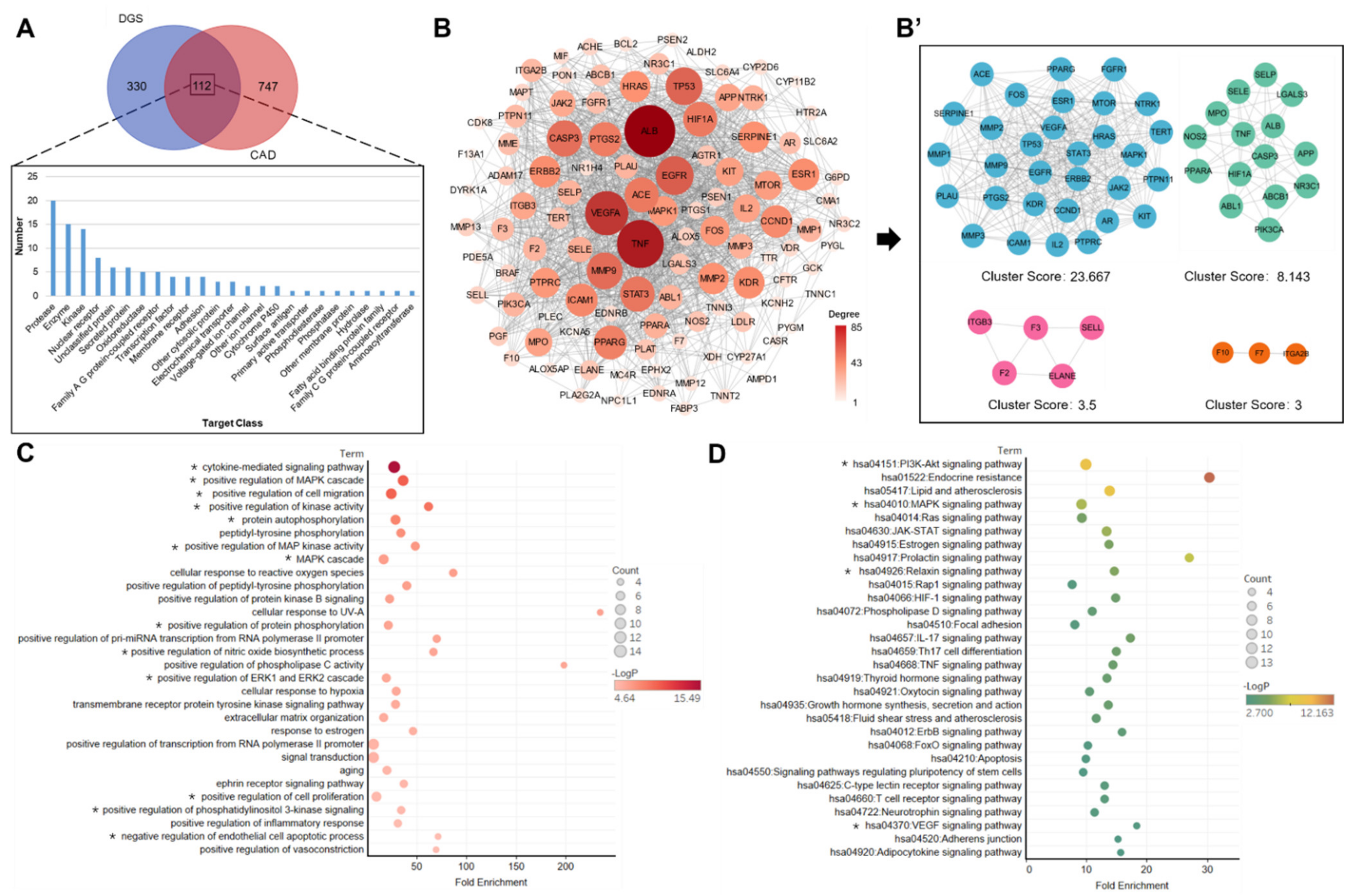
2.2. Key Component and Target Screening and Prediction Using Network Pharmacology Analysis
2.3. Pharmacological Mechanism Analysis in HUVECs
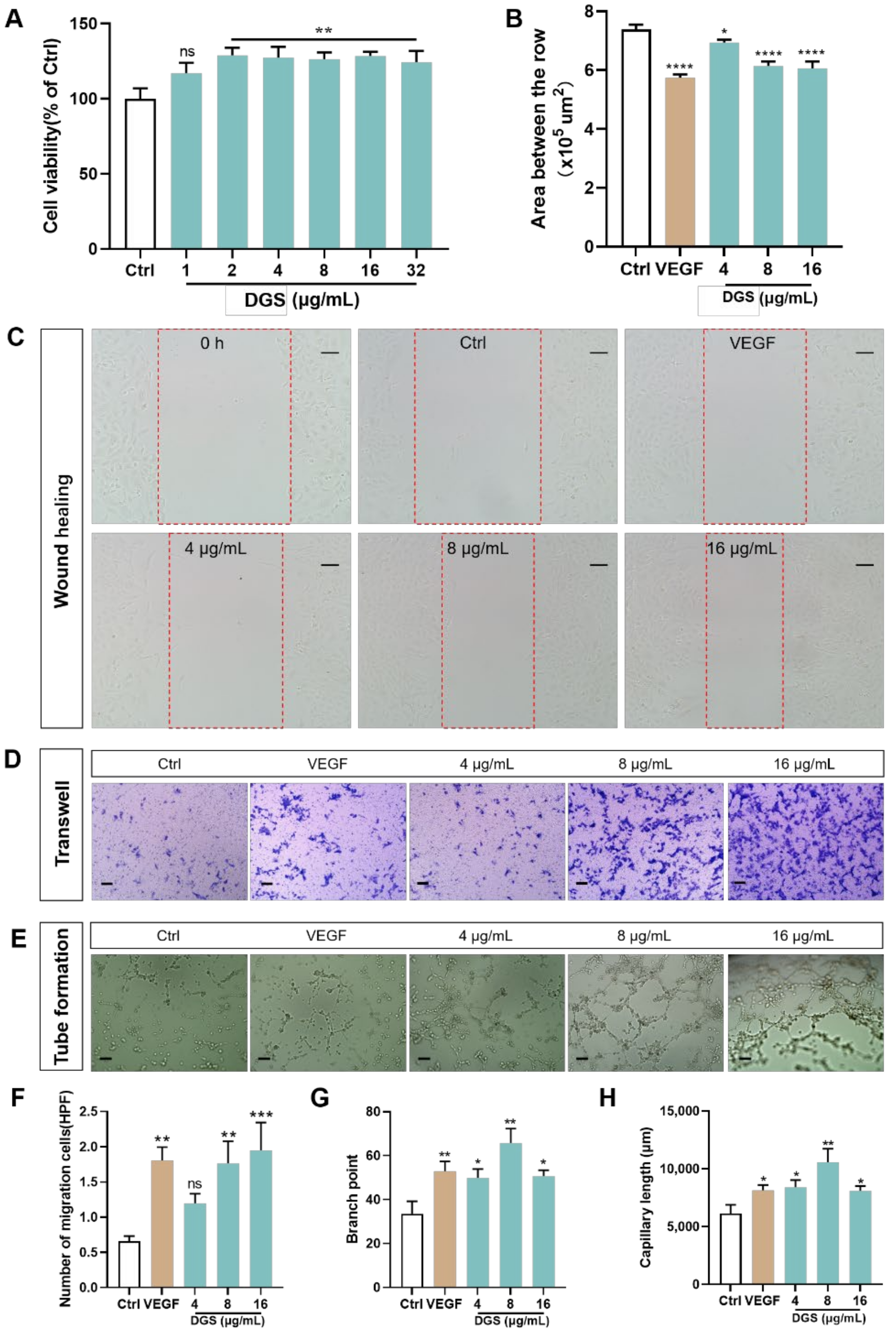
2.3.1. Effect of DGS on the Proliferation and Migration of HUVECs
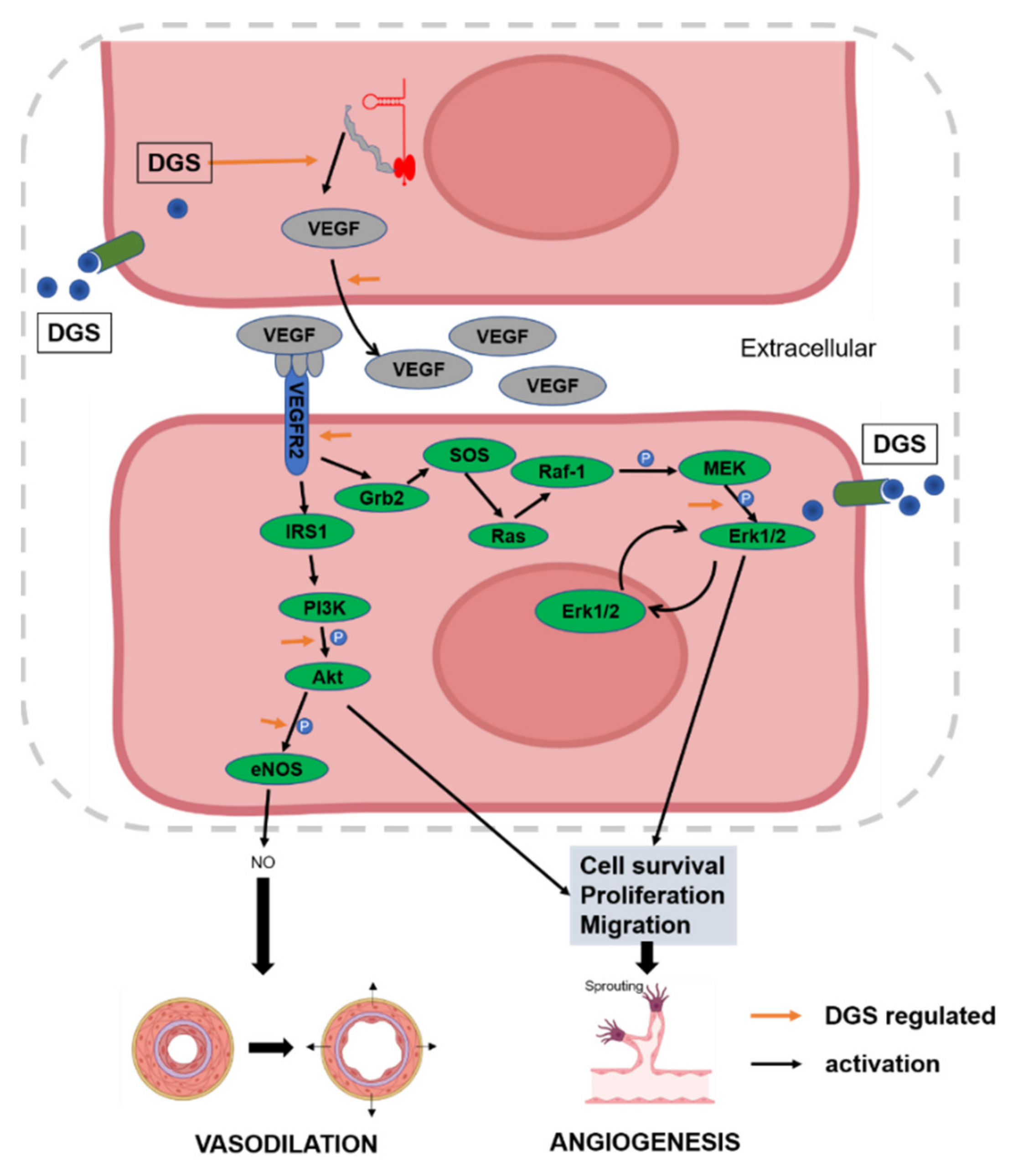
2.3.2. NO and VEGF Release and Protein Expression of DGS
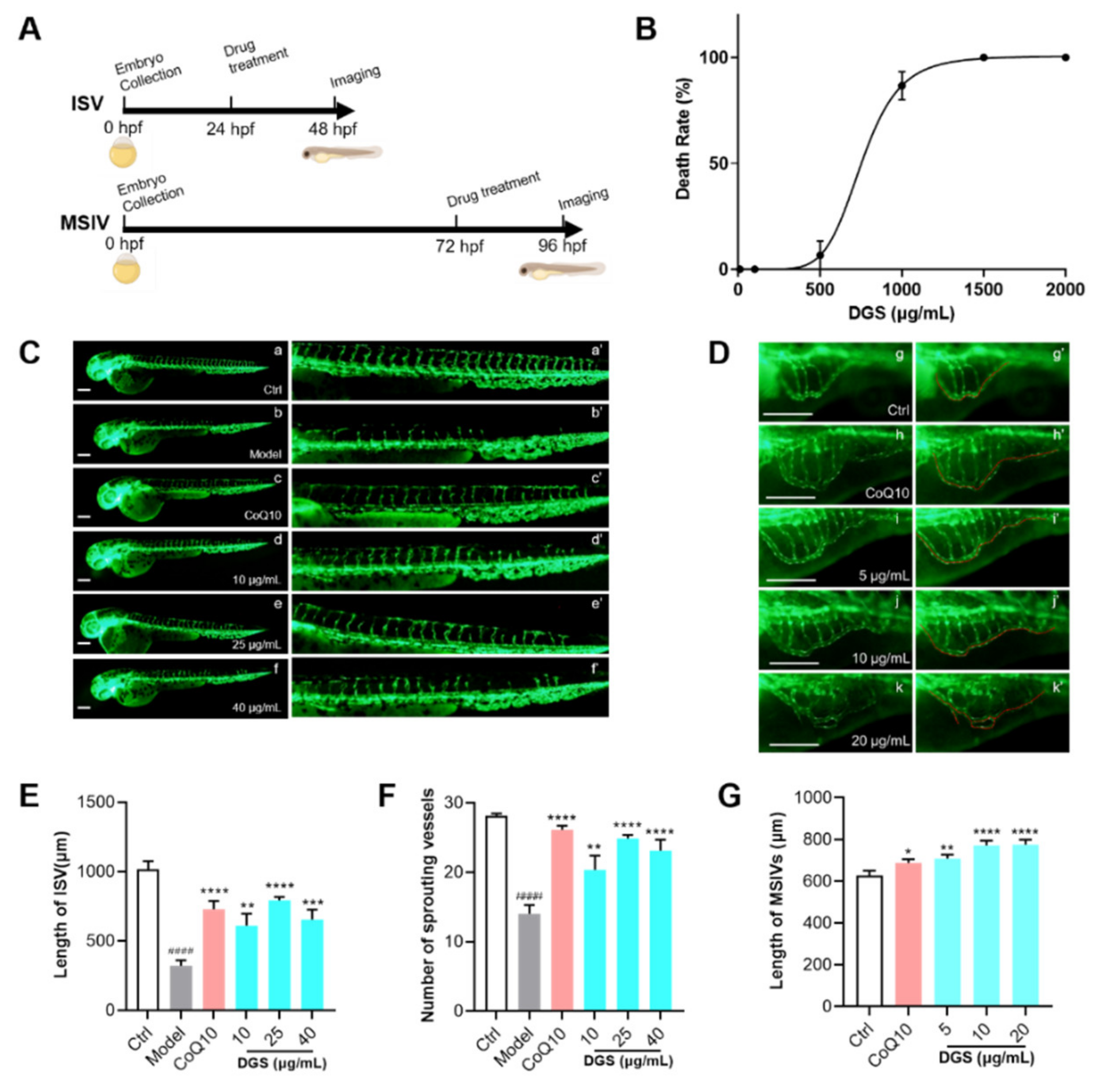
2.4. Bioactivity Evaluation of DGS Based on Zebrafish Model In Vivo
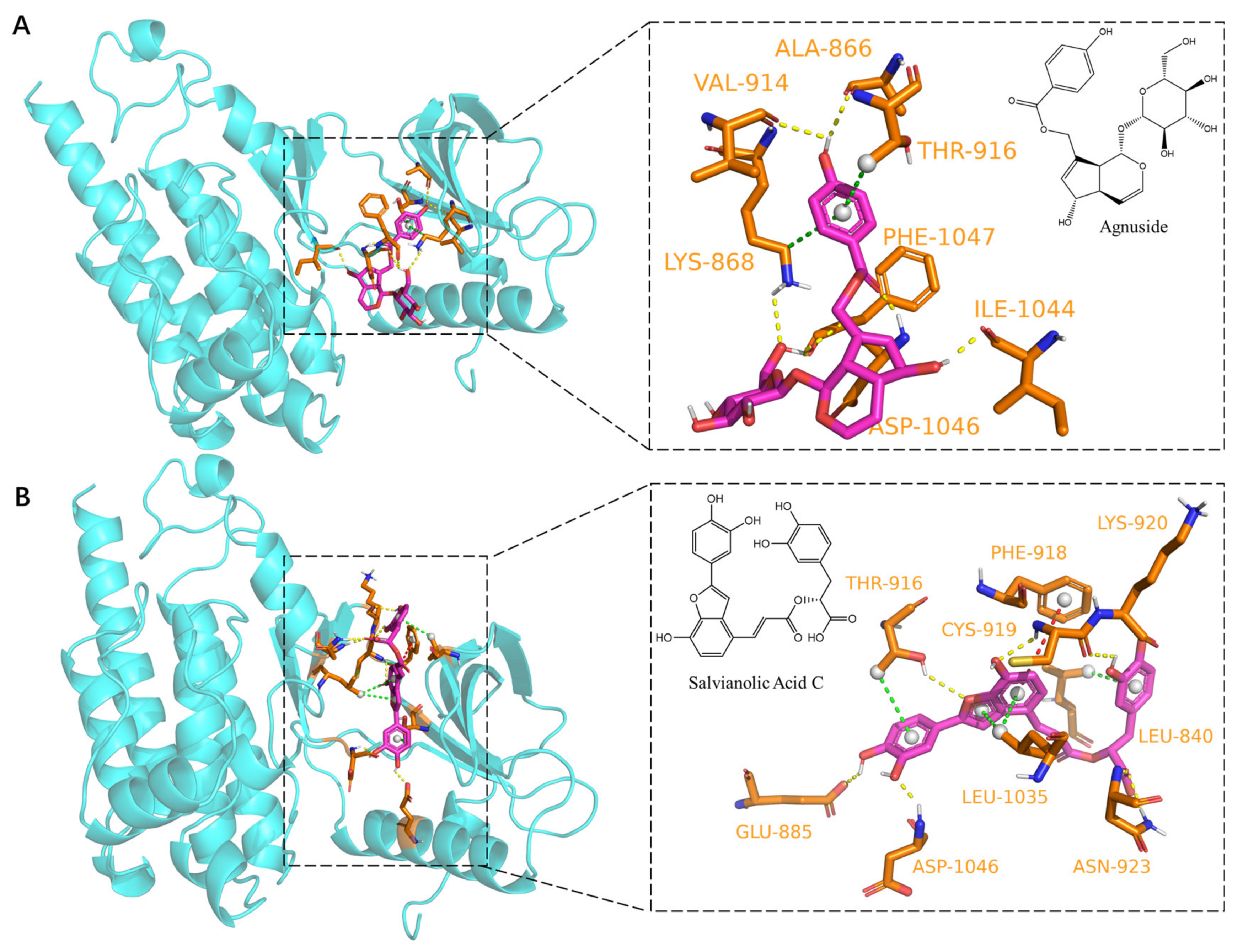
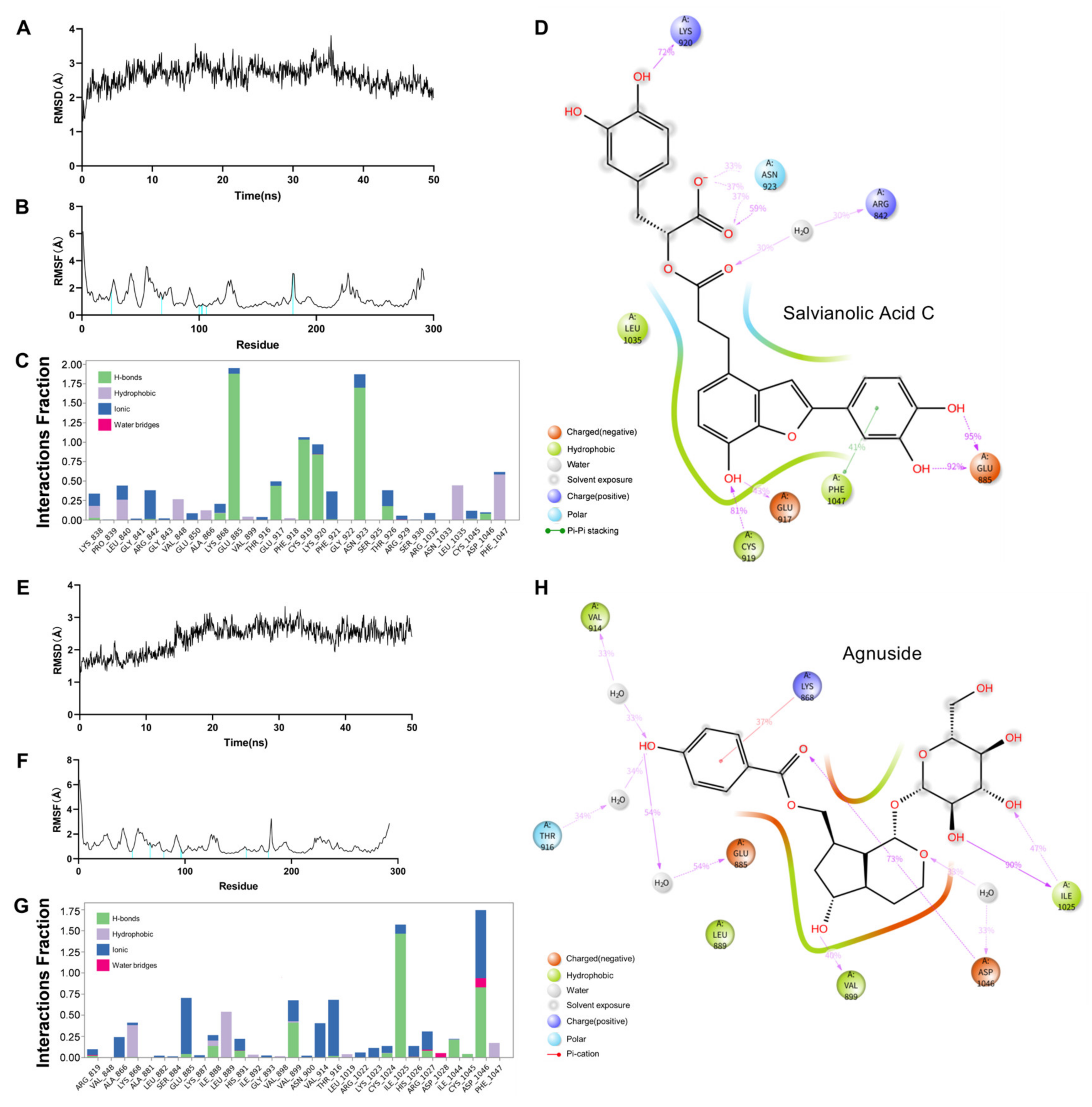
2.5. Dynamic Molecular Docking of Active Components of DGS with Key Targets
3. Discussion
4. Materials and Methods
4.1. Cell and Reagents
4.2. Plant Material and Extraction
4.3. Chromatographic and Mass Spectrometry Conditions
4.4. Collection of DGS-Related Targets and CAD-Related Targets
4.5. Protein–Protein Interaction (PPI) Network
4.6. Gene Ontology (GO) and KEGG Pathway Enrichment Analysis
4.7. Cell Culture and Treatment
4.8. CCK-8 Assay
4.9. Wound Healing Assay
4.10. Transwell Assay
4.11. Tube Formation Assay
4.12. VEGF and NO Level Detection
4.13. Western Blotting
4.14. Animals
4.15. Animal Grouping and Administration
4.16. Dynamics Molecular Docking
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Juan-Salvadores, P.; Jiménez Díaz, V.A.; Iglesia Carreño, C.; Guitián González, A.; Veiga, C.; Martínez Reglero, C.; Baz Alonso, J.A.; Caamaño Isorna, F.; Iñiguez Romo, A. Coronary Artery Disease in Very Young Patients: Analysis of Risk Factors and Long-Term Follow-Up. J. Cardiovasc. Dev. Dis. 2022, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Rocker, A.J.; Lee, D.J.; Shandas, R.; Park, D. Injectable Polymeric Delivery System for Spatiotemporal and Sequential Release of Therapeutic Proteins To Promote Therapeutic Angiogenesis and Reduce Inflammation. ACS Biomater. Sci. Eng. 2020, 6, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Gautam, N.; Saluja, P.; Malkawi, A.; Rabbat, M.G.; Al-Mallah, M.H.; Pontone, G.; Zhang, Y.; Lee, B.C.; Al’Aref, S.J. Current and Future Applications of Artificial Intelligence in Coronary Artery Disease. Healthcare 2022, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Hashim, M.J.; Mustafa, H.; Baniyas, M.Y.; Al Suwaidi, S.K.B.M.; AlKatheeri, R.; Alblooshi, F.M.K.; Almatrooshi, M.E.A.H.; Alzaabi, M.E.H.; Al Darmaki, R.S.; et al. Global Epidemiology of Ischemic Heart Disease: Results from the Global Burden of Disease Study. Cureus 2020, 12, e9349. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Singh, A.; Roy, A.; Mohanty, S.; Naik, N.; Kalaivani, M.; Ramakrishnan, L. Role of Endothelial Colony Forming Cells (ECFCs) Tetrahydrobiopterin (BH4) in Determining ECFCs Functionality in Coronary Artery Disease (CAD) Patients. Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Jamaiyar, A.; Juguilon, C.; Wan, W.; Richardson, D.; Chinchilla, S.; Gadd, J.; Enrick, M.; Wang, T.; McCabe, C.; Wang, Y.; et al. The Essential Role for Endothelial Cell Sprouting in Coronary Collateral Growth. J. Mol. Cell. Cardiol. 2022, 165, 158–171. [Google Scholar] [CrossRef]
- Brendler, T.; Al-Harrasi, A.; Bauer, R.; Gafner, S.; Hardy, M.L.; Heinrich, M.; Hosseinzadeh, H.; Izzo, A.A.; Michaelis, M.; Nassiri-Asl, M.; et al. Botanical Drugs and Supplements Affecting the Immune Response in the Time of COVID-19: Implications for Research and Clinical Practice. Phytother. Res. 2021, 35, 3013–3031. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Ivanova, E.A. Cellular Models of Atherosclerosis and Their Implication for Testing Natural Substances with Anti-Atherosclerotic Potential. Phytomedicine 2016, 23, 1190–1197. [Google Scholar] [CrossRef]
- Ling, Y.; Shi, J.; Ma, Q.; Yang, Q.; Rong, Y.; He, J.; Chen, M. Vasodilatory Effect of Guanxinning Tablet on Rabbit Thoracic Aorta Is Modulated by Both Endothelium-Dependent and -Independent Mechanism. Front. Pharmacol. 2021, 12, 754527. [Google Scholar] [CrossRef]
- Wang, X.-P.; Wang, P.-F.; Bai, J.-Q.; Gao, S.; Wang, Y.-H.; Quan, L.-N.; Wang, F.; Wang, X.-T.; Wang, J.; Xie, Y.-D. Investigating the Effects and Possible Mechanisms of Danshen-Honghua Herb Pair on Acute Myocardial Ischemia Induced by Isoproterenol in Rats. Biomed. Pharmacother. 2019, 118, 109268. [Google Scholar] [CrossRef]
- Wu, W.; Wang, Y. Pharmacological Actions and Therapeutic Applications of Salvia Miltiorrhiza Depside Salt and Its Active Components. Acta Pharmacol. Sin. 2012, 33, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.-J.; Hung, Y.-C.; Kuo, C.-E.; Chung, C.-J.; Hsu, C.Y.; Muo, C.-H.; Hsu, S.-F.; Hu, W.-L. Prescription of Radix Salvia Miltiorrhiza in Taiwan: A Population-Based Study Using the National Health Insurance Research Database. Front. Pharmacol. 2021, 12, 719519. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Colletti, A.; Bellentani, S. Nutraceutical Approach to Non-Alcoholic Fatty Liver Disease (NAFLD): The Available Clinical Evidence. Nutrients 2018, 10, E1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Cui, Y.; Xu, B.; Wang, Y.; Lv, F.; Li, Z.; Li, H.; Chen, X.; Peng, X.; Chen, Y.; et al. Main Active Components of Jiawei Gegen Qinlian Decoction Protects against Ulcerative Colitis under Different Dietary Environments in a Gut Microbiota-Dependent Manner. Pharmacol. Res. 2021, 170, 105694. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Lv, Z.; Huang, Z.; Ye, N.; Li, S.; Jiang, J.; Cheng, Y.; Shi, F. Dietary Hawthorn-Leaves Flavonoids Improves Ovarian Function and Liver Lipid Metabolism in Aged Breeder Hens. Poult. Sci. 2021, 100, 101499. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Q.; Pan, Y.; Xu, L. Sal B Alleviates Myocardial Ischemic Injury by Inhibiting TLR4 and the Priming Phase of NLRP3 Inflammasome. Molecules 2019, 24, 4416. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Yi, B.; Chen, M.; Wang, N.; Chen, P.; Guo, C.; Sun, J. Induction of Nur77 by Hyperoside Inhibits Vascular Smooth Muscle Cell Proliferation and Neointimal Formation. Biochem. Pharmacol. 2014, 92, 590–598. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Cheung, M.-K.; Yue, G.G.-L.; Leung, P.-C.; Wong, C.-K.; Lau, C.B.-S. Integrated Network Pharmacology Analysis and In Vitro Validation Revealed the Potential Active Components and Underlying Mechanistic Pathways of Herba Patriniae in Colorectal Cancer. Molecules 2021, 26, 6032. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhang, W.; Wang, R.; Li, C.; Lin, X.; Wang, L. Screening and Identification of Natural α-Glucosidase and α-Amylase Inhibitors from Partridge Tea (Mallotus Furetianus Muell-Arg) and in Silico Analysis. Food Chem. 2022, 388, 133004. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, N.; Ma, J.; Zhu, Y.; Wang, M.; Wang, X.; Zhang, P. A Platelet/CMC Coupled with Offline UPLC-QTOF-MS/MS for Screening Antiplatelet Activity Components from Aqueous Extract of Danshen. J. Pharm. Biomed. Anal. 2016, 117, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Pillarisetti, P.; Myers, K.A. Identification and Characterization of Agnuside, a Natural Proangiogenic Small Molecule. Eur. J. Med. Chem. 2018, 160, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Suo, T.-C.; Yu, H.; Li, A.-P.; Zhang, S.-Q.; Wang, C.-H.; Zhu, Y.; Li, Z. A New Strategy for Choosing “Q-Markers” via Network Pharmacology, Application to the Quality Control of a Chinese Medical Preparation. J. Food Drug Anal. 2018, 26, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, B.; Jiang, D.; Wei, Y.; Zhang, N. Herb Network Construction and Co-Module Analysis for Uncovering the Combination Rule of Traditional Chinese Herbal Formulae. BMC Bioinform. 2010, 11, S6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhang, B. Traditional Chinese Medicine Network Pharmacology: Theory, Methodology and Application. Chin. J. Nat. Med. 2013, 11, 110–120. [Google Scholar] [CrossRef]
- Yu, L.-J.; Zhang, K.-J.; Zhu, J.-Z.; Zheng, Q.; Bao, X.-Y.; Thapa, S.; Wang, Y.; Chu, M.-P. Salvianolic Acid Exerts Cardioprotection through Promoting Angiogenesis in Animal Models of Acute Myocardial Infarction: Preclinical Evidence. Oxidative Med. Cell. Longev. 2017, 2017, 8192383. [Google Scholar] [CrossRef]
- Syahputra, R.A.; Harahap, U.; Dalimunthe, A.; Nasution, M.P.; Satria, D. The Role of Flavonoids as a Cardioprotective Strategy against Doxorubicin-Induced Cardiotoxicity: A Review. Molecules 2022, 27, 1320. [Google Scholar] [CrossRef]
- Ghigo, A.; Laffargue, M.; Li, M.; Hirsch, E. PI3K and Calcium Signaling in Cardiovascular Disease. Circ. Res. 2017, 121, 282–292. [Google Scholar] [CrossRef]
- Kazakov, A.; Hall, R.; Jagoda, P.; Bachelier, K.; Müller-Best, P.; Semenov, A.; Lammert, F.; Böhm, M.; Laufs, U. Inhibition of Endothelial Nitric Oxide Synthase Induces and Enhances Myocardial Fibrosis. Cardiovasc. Res. 2013, 100, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, Y.; Li, C.; Xie, J.; Liu, Y.; Zhu, W.; Zhang, X.; Jiang, S.; Liu, L.; Ding, Z. HSPA12B Attenuates Cardiac Dysfunction and Remodelling after Myocardial Infarction through an ENOS-Dependent Mechanism. Cardiovasc. Res. 2013, 99, 674–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.-N.; Li, P.; Liu, W.-H.; Shang, J.-J.; Qiu, S.-L.; Zhou, M.-X.; Liu, H.-X. Danhong Injection Enhances Angiogenesis after Myocardial Infarction by Activating MiR-126/ERK/VEGF Pathway. Biomed. Pharmacother. 2019, 120, 109538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, Y.; Wang, L.; Li, X.; Zhang, S.; Wang, X.; Jin, M.; Hsiao, C.-D.; Lin, H.; Han, L.; et al. Metabolomics for Biomarker Discovery in Fermented Black Garlic and Potential Bioprotective Responses against Cardiovascular Diseases. J. Agric. Food Chem. 2019, 67, 12191–12198. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Tian, Q.; Shi, Y.; Chen, S.; Zhang, Y.; Deng, Z.; Wang, X.; Yao, Q.; Han, L. An Integrated Strategy for Rapid Discovery and Identification of Quality Markers in Gardenia Fructus Using an Omics Discrimination-Grey Correlation-Biological Verification Method. Front. Pharmacol. 2021, 12, 705498. [Google Scholar] [CrossRef]
- Lawson, N.D.; Weinstein, B.M. In Vivo Imaging of Embryonic Vascular Development Using Transgenic Zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Huang, X.; Kanisicak, O.; Li, Y.; Wang, Y.; Li, Y.; Pu, W.; Liu, Q.; Zhang, H.; Tian, X.; et al. Preexisting Endothelial Cells Mediate Cardiac Neovascularization after Injury. J. Clin. Investig. 2017, 127, 2968–2981. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, H.; He, L.; Huang, X.; Li, Y.; Pu, W.; Yu, W.; Zhang, L.; Cai, D.; Lui, K.O.; et al. Genetic Fate Mapping Defines the Vascular Potential of Endocardial Cells in the Adult Heart. Circ. Res. 2018, 122, 984–993. [Google Scholar] [CrossRef]
- Greenspan, L.J.; Weinstein, B.M. To Be or Not to Be: Endothelial Cell Plasticity in Development, Repair, and Disease. Angiogenesis 2021, 24, 251–269. [Google Scholar] [CrossRef]
- Xiang, Y.; Yao, X.; Wang, X.; Zhao, H.; Zou, H.; Wang, L.; Zhang, Q.-X. Houshiheisan Promotes Angiogenesis via HIF-1α/VEGF and SDF-1/CXCR4 Pathways: In Vivo and in Vitro. Biosci. Rep. 2019, 39, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Monika, P.; Chandraprabha, M.N.; Rangarajan, A.; Waiker, P.V.; Chidambara Murthy, K.N. Challenges in Healing Wound: Role of Complementary and Alternative Medicine. Front. Nutr. 2021, 8, 791899. [Google Scholar] [CrossRef]
- Salo, T.; Dourado, M.R.; Sundquist, E.; Apu, E.H.; Alahuhta, I.; Tuomainen, K.; Vasara, J.; Al-Samadi, A. Organotypic Three-Dimensional Assays Based on Human Leiomyoma-Derived Matrices. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160482. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Gómez, J.M.; Santiago-Mora, R.; Navarro-Valverde, C.; Dorado, G.; Casado-Díaz, A. Stimulation of In-Vitro Angiogenesis by Low Concentrations of Risedronate Is Mitigated by 1,25-Dihydroxyvitamin D3 or 24,25-Dihydroxyvitamin D3. J. Steroid Biochem. Mol. Biol. 2015, 148, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Lucas-Herald, A.K.; Montezano, A.C.; Alves-Lopes, R.; Haddow, L.; Alimussina, M.; O’Toole, S.; Flett, M.; Lee, B.; Amjad, S.B.; Steven, M.; et al. Vascular Dysfunction and Increased Cardiovascular Risk in Hypospadias. Eur. Heart J. 2022, 43, 1832–1845. [Google Scholar] [CrossRef]
- Zullino, S.; Buzzella, F.; Simoncini, T. Nitric Oxide and the Biology of Pregnancy. Vasc. Pharmacol. 2018, 110, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Ambari, A.M.; Lilihata, G.; Zuhri, E.; Ekawati, E.; Wijaya, S.A.; Dwiputra, B.; Sukmawan, R.; Radi, B.; Haryana, S.M.; Adiarto, S.; et al. External Counterpulsation Improves Angiogenesis by Preserving Vascular Endothelial Growth Factor-A and Vascular Endothelial Growth Factor Receptor-2 but Not Regulating MicroRNA-92a Expression in Patients With Refractory Angina. Front. Cardiovasc. Med. 2021, 8, 761112. [Google Scholar] [CrossRef]
- Li, J.-Z.; Zhou, X.-X.; Wu, W.-Y.; Qiang, H.-F.; Xiao, G.-S.; Wang, Y.; Li, G. Concanavalin A Promotes Angiogenesis and Proliferation in Endothelial Cells through the Akt/ERK/Cyclin D1 Axis. Pharm. Biol. 2022, 60, 65–74. [Google Scholar] [CrossRef]
- Gfeller, D.; Michielin, O.; Zoete, V. Shaping the Interaction Landscape of Bioactive Molecules. Bioinformatics 2013, 29, 3073–3079. [Google Scholar] [CrossRef]
- Safran, M.; Rosen, N.; Twik, M.; BarShir, R.; Stein, T.I.; Dahary, D.; Fishilevich, S.; Lancet, D. The GeneCards Suite. In Practical Guide to Life Science Databases; Abugessaisa, I., Kasukawa, T., Eds.; Springer Nature Singapore: Singapore, 2021; pp. 27–56. ISBN 978-981-16-5812-9. [Google Scholar]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2–27. [Google Scholar] [CrossRef] [Green Version]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 10, 1–6. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | Formular | Calc. Mass | Molecular Ion (m/z) | Mass Error (ppm) | Fragment Ion | Compound Name |
|---|---|---|---|---|---|---|---|
| 1 | 1.256 | C7H12O6 | 191.05611 | 191.0576 | 7.8 | 173.0375, 93.0279, 87.0043, 85.0251 | Quinic acid |
| 2 | 3.268 | C11H12O7 | 255.0510 | 255.0505 | −2.1 | 165.0451, 147.0418, 131.0459 | Piscidic acid |
| 3 | 4.823 | C26H28O13 | 547.14571 | 547.1468 | 2.0 | 295.0490, 267.0542 | Mirificin |
| 4 | 4.908 | C30H26O12 | 577.1352 | 577.138 | 3.2 | 425.5437, 255.4897 | Procyanidin B2 |
| 5 | 5.23 | C15H14O6 | 289.07176 | 289.0737 | 6.9 | 205.2053, 189.8794 | (−)-Epicatechin |
| 6 | 6.283 | C21H20O10 | 431.0937 | 431.0899 | −8.8 | 311.0481, 283.0500 | Vitexin |
| 7 | 6.425 | C16H18O9 | 353.0878 | 353.08501 | −7.9 | 191.0235, 179.0027 | Chlorogenic acid |
| 8 | 6.866 | C11H12O6 | 239.0561 | 239.0551 | −4.2 | 179.0277, 177.0486, 107.0453 | 2-(Carboxymethyl)-4,5-dimethoxybenzoic acid |
| 9 | 7.199 | C22H22O11 | 461.1035 | 461.1033 | −0.4 | 253.0397 | Tectoridin |
| 10 | 7.55 | C26H28O14 | 563.1406 | 563.1428 | 3.9 | 311.0433, 283.0471, 227.0628 | Schaftoside |
| 11 | 8.52 | C21H20O9 | 415.1005 | 415.0963 | −7.2 | 296.0397, 267.0549, 207.0577, 193.0550 | Puerarin |
| 12 | 9.137 | C26H20O10 | 491.0984 | 491.0985 | 0.3 | 363.1897, 351.6277, 320.0439 | Salvianolic acid C |
| 13 | 9.202 | C22H22O10 | 445.1140 | 445.1103 | −8.3 | 283.0402 | 3′-Methoxypuerarin |
| 14 | 10.518 | C15H10O4 | 253.0506 | 253.0487 | −7.6 | 223.0306, 209.0538 | Daidzein |
| 15 | 11.318 | C29H34O14 | 605.1876 | 605.1836 | −6.6 | 297.0657, 253.0771 | Pueroside A |
| 16 | 11.706 | C21H20O10 | 431.0984 | 431.0955 | −6.7 | 269.0337 | Genistin |
| 17 | 12.261 | C27H30O14 | 577.1563 | 577.1557 | −1.1 | 413.0696, 313.0373, 293.0346 | Vitexin 2″-O-rhamnoside |
| 18 | 13.302 | C15H10O5 | 269.0456 | 269.0478 | 8.4 | 252.0337 | Genistein |
| 19 | 13.341 | C27H30O15 | 593.1512 | 593.1483 | −4.9 | 311.0412, 283.0351 | Tectorigenin 7-O-xylosylglucoside |
| 20 | 14.927 | C21H20O12 | 463.0882 | 463.0851 | −6.7 | 303.0245 | Hyperoside |
| 21 | 15.64 | C27H30O16 | 609.1461 | 609.1415 | −7.6 | 301.03118 | Rutin |
| 22 | 15.74 | C18H16O8 | 359.0772 | 359.0761 | −3.2 | 161.0169, 135.0388, 119.4798 | Rosmarinic acid |
| 23 | 16.1 | C26H22O10 | 493.1140 | 493.1100 | −8.2 | 295.0498, 313.0585, 197.0355, 162.0195 | Salvianolic acid A |
| 24 | 16.452 | C27H22O12 | 537.1038 | 537.0986 | −9.8 | 313.0583, 295.0497 | Lithospermic acid |
| 25 | 17.822 | C36H30O16 | 717.14611 | 717.1420 | −5.7 | 429.1065, 339.0366, 320.0352, 279.0567, 185.0106 | Salvianolic acid B |
| 26 | 19.44 | C27H30O15 | 593.1512 | 593.1471 | −6.9 | 414.0734, 311.0541, 293.0322 | Vitexin-4″-O-glucoside |
| 27 | 20.929 | C36H30O16 | 717.1461 | 717.1420 | −5.7 | 537.0832, 493.0698, 339.0386, 295.0556 | Salvianolic acid L |
| 28 | 23.049 | C29H26O12 | 565.1352 | 565.1376 | 4.3 | 339.0376, 321.0272, 293.0328 | Dimethyl Lithospermate |
| 29 | 24.749 | C26H20O10 | 491.0984 | 491.0939 | −9.1 | 295.0532 | isosalvianolic acid C |
| 30 | 31.259 | C20H28O2 | 301.2162 | 301.2156 | −2.1 | 271.6406, 259.5924 | Sugiol |
| 31 | 33.13 | C16H16O5 | 287.0925 | 287.0906 | −6.8 | 269.2039, 258.1443 | Shikonin |
| 32 | 33.32 | C19H18O3 | 293.1138 | 293.1192 | 3 | 231.3145, 221.1474 | Tanshinone IIA |
| 33 | 33.845 | C19H22O4 | 313.14453 | 313.1455 | 3.0 | 227.0246, 212.0280, 267.0357 | Tanshinone V |
| 34 | 35.93 | C19H20O3 | 295.1340 | 295.1362 | 7.6 | 277.0811, 265.0752, 209.0584 | Cryptotanshinone |
| 35 | 36.129 | C19H22O3 | 297.1496 | 297.1519 | 7.7 | 270.3796, 253.1184 | 2-[2-(6-methoxy-3,4-dihydro-2H-naphthalen-1-ylidene)ethyl]-2-methylcyclopentane-1,3-dione |
| 36 | 36.555 | C17H14O6 | 313.0717 | 313.0703 | −4.4 | 295.1204, 283.1224, 268, 255.1049 | Salvianolic acid F |
| 37 | 37.433 | C39H54O7 | 633.3797 | 633.3828 | 4.9 | 617.3838, 471.358 | 3-O-p-Coumaroyltormentic acid |
| Ligand | Binding Affinity (kcal/mol) | Type of Interaction |
|---|---|---|
| Agnuside | −8.8 | Hydrogen bonding: PHE-1047, ILE-1044, ASP-046, LYS-868, VAL-914, ALA-866; |
| π–Sigma: THR-916 | ||
| Salvianolic Acid C | −10.7 | Hydrogen bonding: LYS-920, CYS-919, ASN-923, ASP-1046, GLU-885, THR-916; |
| π–Sigma: LEU-840, LEU-1035, THR-916; | ||
| π–π stacked: PHE-918 | ||
| Isosalvianolic Acid C | −9.7 | Hydrogen bonding: GLU-917, CYS-919, ASN-923; |
| π–Sigma: PHE-1047; | ||
| π–π stacked: PHE-918, PHE-1047 | ||
| Genistin | −9.6 | Hydrogen bonding: GLU-885, THR-916, ASN-923; |
| π–Sigma: THR-916, VAL-848, LEU-1035; | ||
| π–π stacked: PHE-918 | ||
| 3-O-p-Coumaroyltormentic acid | -9.4 | Hydrogen bonding: ILE-1025; |
| π–π stacked: PHE-845 | ||
| Tanshinone IIA | −9.2 | π–Sigma: PHE-1047, PHE-845; |
| π–π stacked: PHE-845 | ||
| 2-[2-(6-Methoxy-3,4-Dihydro-2H-Naphthalen-1-Ylidene)Ethyl]-2-Methylcyclopentane-1,3-Dione | −9.2 | π–Sigma: LEU-840, |
| π–π stacked: PHE-845 | ||
| Daidzein | −9.1 | π–Sigma: THR-916, LEU-1035, LEU-840; |
| π–π stacked: PHE-918 | ||
| Rosmarinic Acid | −9 | Hydrogen bonding: GLU-917, CYS-919, ASP-1046, LYS-868; |
| π–Sigma: LEU-889, LEU-1035 | ||
| Lithospermic Acid | −9 | Hydrogen bonding: GLU-885, MET-869, ASP-1046; |
| π–π stacked: PHE-845 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-G.; Liu, X.-X.; Zong, J.-C.; Zhang, Y.-T.-J.; Dong, R.; Wang, N.; Ma, Z.-H.; Li, L.; Wang, S.-L.; Mu, Y.-L.; et al. Investigation Driven by Network Pharmacology on Potential Components and Mechanism of DGS, a Natural Vasoprotective Combination, for the Phytotherapy of Coronary Artery Disease. Molecules 2022, 27, 4075. https://doi.org/10.3390/molecules27134075
Zhang Y-G, Liu X-X, Zong J-C, Zhang Y-T-J, Dong R, Wang N, Ma Z-H, Li L, Wang S-L, Mu Y-L, et al. Investigation Driven by Network Pharmacology on Potential Components and Mechanism of DGS, a Natural Vasoprotective Combination, for the Phytotherapy of Coronary Artery Disease. Molecules. 2022; 27(13):4075. https://doi.org/10.3390/molecules27134075
Chicago/Turabian StyleZhang, You-Gang, Xia-Xia Liu, Jian-Cheng Zong, Yang-Teng-Jiao Zhang, Rong Dong, Na Wang, Zhi-Hui Ma, Li Li, Shang-Long Wang, Yan-Ling Mu, and et al. 2022. "Investigation Driven by Network Pharmacology on Potential Components and Mechanism of DGS, a Natural Vasoprotective Combination, for the Phytotherapy of Coronary Artery Disease" Molecules 27, no. 13: 4075. https://doi.org/10.3390/molecules27134075