
The Role of Antimicrobial Peptides as Antimicrobial and Antibiofilm Agents in Tackling the Silent Pandemic of Antimicrobial Resistance

 , , ,
, , ,
Abstract
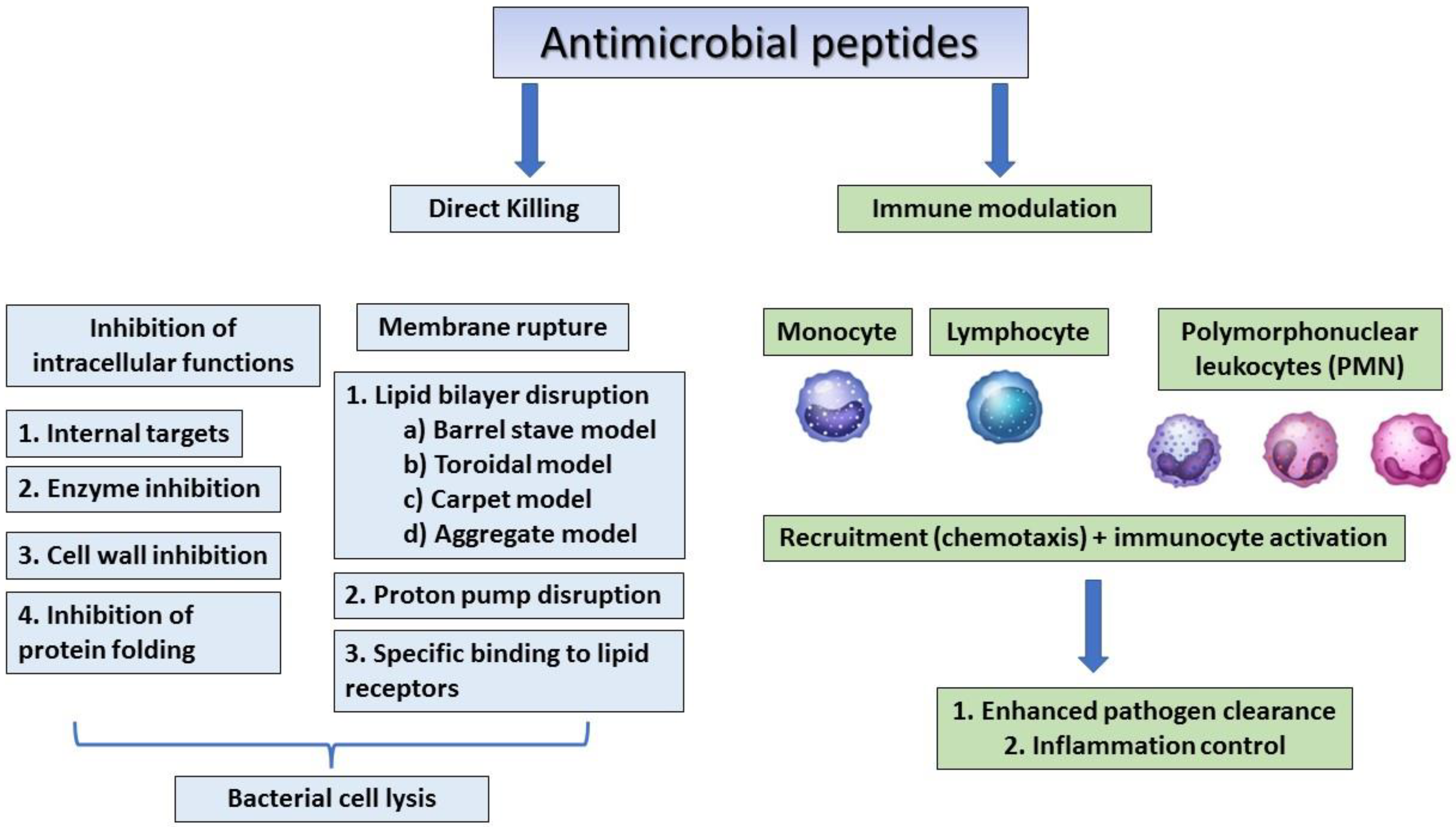
:1. Introduction
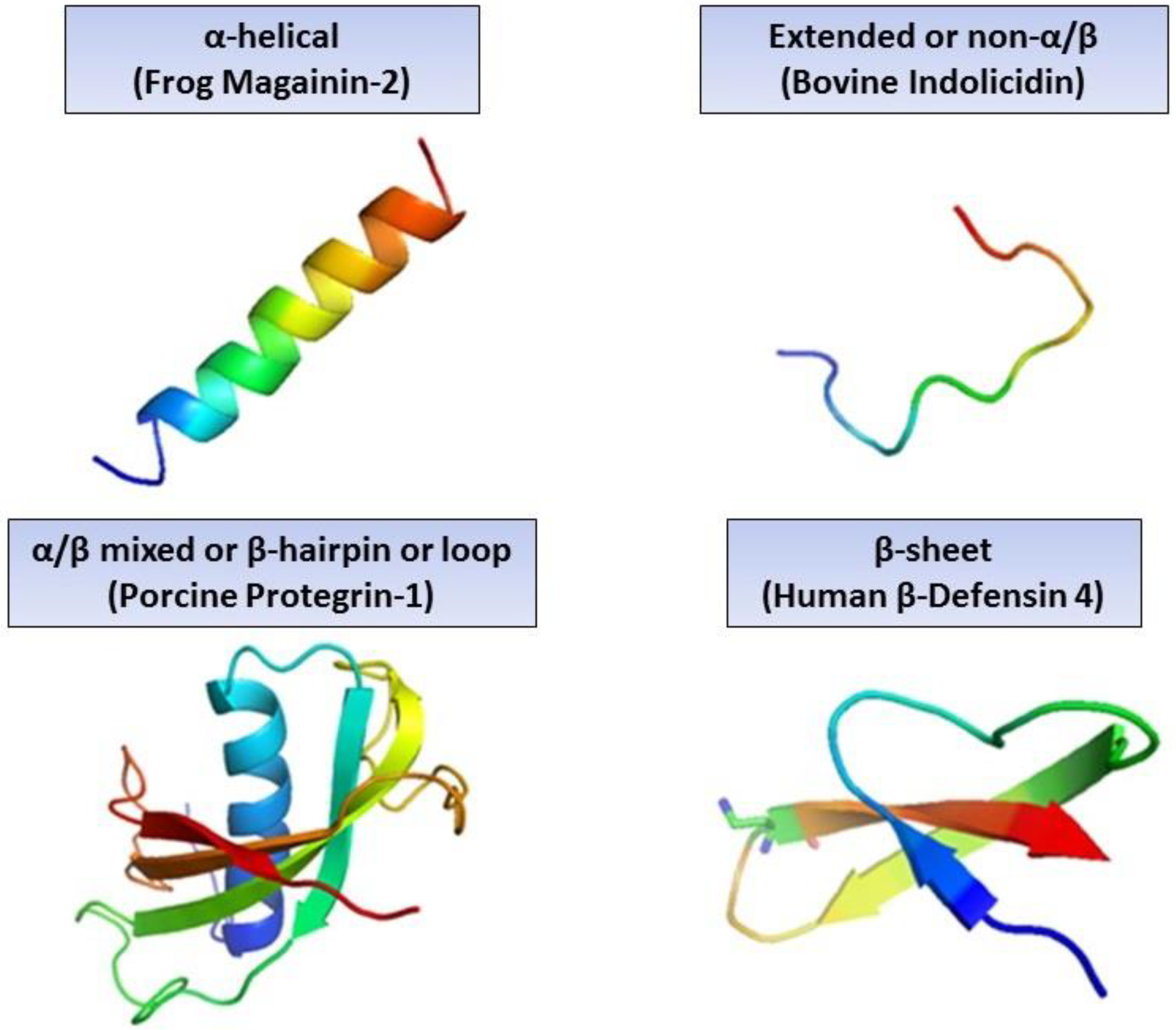
2. Different Classes of AMPs and Their Need in Modern Medicine
3. Sources of AMPs and Their Inhibitory Effects
3.1. Mammalian AMPs
3.2. Amphibian and Fish AMPs
3.3. AMPs from Microorganisms
3.3.1. AMPs from Gram-Positive Bacteria
3.3.2. AMPs from Gram-Negative Bacteria
3.3.3. Fungal AMPs
3.3.4. Viral and Bacteriophage AMPs
3.3.5. Insect AMPs
4. Bacterial Biofilm Formation in Priority Pathogens
4.1. Enterococcus spp.
4.2. Staphylococcus aureus
4.3. Klebsiella pneumoniae
4.4. Acinetobacter baumannii
4.5. Pseudomonas aeruginosa
4.6. Enterobacter spp.
4.7. Helicobacter pylori
4.8. Campylobacter spp.
5. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s Penicillin Producing Strain Is Not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, A. On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to their use in the Isolation of B. influenzae. Bull. World Health Organ. 2001, 79, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Lederberg, J. Infectious History. Science 2000, 288, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, B.S.; Strachan, N.J.C.; Ramjee, M.; Thomson, A.; MacRae, M.; Shaw, S.; Forbes, K.J. Nationwide Stepwise Emergence and Evolution of Multidrug-Resistant Campylobacter Jejuni Sequence Type 5136, United Kingdom. Emerg. Infect. Dis. 2019, 25, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Dubos, R.J. Studies on a Bactericidal Agent Extracted from a Soil Bacillus: I. Preparation of the Agent. Its Activity In Vitro. J. Exp. Med. 1939, 70, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dubos, R.J. Studies on a Bactericidal Agent Extracted from a Soil Bacillus: II. Protective Effect of the Bactericidal Agent against Experimental Pneumococcus Infections in Mice. J. Exp. Med. 1939, 70, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, R.D.; Dubos, R.J. Fractionation of the Bactericidal Agent from Cultures of a Soil Bacillus. J. Biol. Chem. 1940, 132, 791–792. [Google Scholar] [CrossRef]
- Van Epps, H.L. Rene Dubos: Unearthing Antibiotics. J. Exp. Med. 2006, 203, 259. [Google Scholar] [CrossRef] [Green Version]
- Van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and Mechanisms of Action of Naturally Occurring Antifungal Peptides. Cell. Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef]
- Kaur, J.; Sagaram, U.S.; Shah, D. Can Plant Defensins be used to Engineer Durable Commercially Useful Fungal Resistance in Crop Plants? Fungal Biol. Rev. 2011, 25, 128–135. [Google Scholar] [CrossRef]
- Tavares, P.M.; Thevissen, K.; Cammue, B.P.; François, I.E.; Barreto-Bergter, E.; Taborda, C.P.; Marques, A.F.; Rodrigues, M.L.; Nimrichter, L. In Vitro Activity of the Antifungal Plant Defensin RsAFP2 against Candida Isolates and its In Vivo Efficacy in Prophylactic Murine Models of Candidiasis. Antimicrob. Agents Chemother. 2008, 52, 4522–4525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.K.; Maurya, I.K.; Khan, S.I.; Jacob, M.R.; Kumar, V.; Tikoo, K.; Jain, R. Discovery of a Membrane-Active, Ring-Modified Histidine Containing Ultrashort Amphiphilic Peptide that Exhibits Potent Inhibition of Cryptococcus Neoformans. J. Med. Chem. 2017, 60, 6607–6621. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente-Nunez, C.; Silva, O.N.; Lu, T.K.; Franco, O.L. Antimicrobial Peptides: Role in Human Disease and Potential as Immunotherapies. Pharmacol. Ther. 2017, 178, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Gallo, R.L. Antimicrobial Peptides: Old Molecules with New Ideas. J. Investig. Dermatol. 2012, 132, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Powers, J.S.; Hancock, R.E. The Relationship between Peptide Structure and Antibacterial Activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial Peptides: Key Components of the Innate Immune System. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Hall, K.N.; Aguilar, M. Antimicrobial Peptide Structure and Mechanism of Action: A Focus on the Role of Membrane Structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef]
- Dhople, V.; Krukemeyer, A.; Ramamoorthy, A. The Human Beta-Defensin-3, an Antibacterial Peptide with Multiple Biological Functions. Biochim. Biophys. Acta (BBA)—Biomembr. 2006, 1758, 1499–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Hancock, R.E. Interaction of the Cyclic Antimicrobial Cationic Peptide Bactenecin with the Outer and Cytoplasmic Membrane. J. Biol. Chem. 1999, 274, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Villaruz, A.E.; Li, M.; Cha, D.J.; Sturdevant, D.E.; Otto, M. The Human Anionic Antimicrobial Peptide Dermcidin Induces Proteolytic Defence Mechanisms in Staphylococci. Mol. Microbiol. 2007, 63, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef] [PubMed]
- Schittek, B.; Hipfel, R.; Sauer, B.; Bauer, J.; Kalbacher, H.; Stevanovic, S.; Schirle, M.; Schroeder, K.; Blin, N.; Meier, F. Dermcidin: A Novel Human Antibiotic Peptide Secreted by Sweat Glands. Nat. Immunol. 2001, 2, 1133–1137. [Google Scholar] [CrossRef]
- Acharya, K.P.; Subramanya, S.H.; Lopes, B.S. Combatting Antimicrobial Resistance in Nepal: The Need for Precision Surveillance Programmes and Multi-Sectoral Partnership. JAC—Antimicrob. Resist. 2019, 1, dlz066. [Google Scholar] [CrossRef]
- Sukri, A.; Lopes, B.S.; Hanafiah, A. The Emergence of Multidrug-Resistant Helicobacter pylori in Southeast Asia: A Systematic Review on the Trends and Intervention Strategies using Antimicrobial Peptides. Antibiotics 2021, 10, 1061. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- O′Neill, J. Review on Antimicrobial Resistance: Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Wellcome Trust: London, UK, 2016. [Google Scholar]
- Manohar, P.; Nachimuthu, R.; Lopes, B.S. The Therapeutic Potential of Bacteriophages Targeting Gram-Negative Bacteria Using Galleria Mellonella Infection Model. BMC Microbiol. 2018, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Vallet-Regí, M.; González, B.; Izquierdo-Barba, I. Nanomaterials as Promising Alternative in the Infection Treatment. Int. J. Mol. Sci. 2019, 20, 3806. [Google Scholar] [CrossRef] [Green Version]
- Silva, O.; De La Fuente-Núñez, C.; Haney, E.; Fensterseifer, I.; Ribeiro, S.; Porto, W.; Brown, P.; Faria-Junior, C.; Rezende, T.; Moreno, S. An Anti-Infective Synthetic Peptide with Dual Antimicrobial and Immunomodulatory Activities. Sci. Rep. 2016, 6, 35465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anunthawan, T.; De La Fuente-Núñez, C.; Hancock, R.E.; Klaynongsruang, S. Cationic Amphipathic Peptides KT2 and RT2 are Taken Up into Bacterial Cells and Kill Planktonic and Biofilm Bacteria. Biochim. Biophys. Acta (BBA)—Biomembr. 2015, 1848, 1352–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duplantier, A.J.; van Hoek, M.L. The Human Cathelicidin Antimicrobial Peptide LL-37 as a Potential Treatment for Polymicrobial Infected Wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Ouellette, A.J.; Bevins, C.L. Paneth Cell Defensins and Innate Immunity of the Small Bowel. Inflamm. Bowel Dis. 2001, 7, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.M.; Chertov, O.; Lubkowski, J. The Structure of Human Β-Defensin-1: New Insights into Structural Properties of Β-Defensins. J. Biol. Chem. 2001, 276, 39021–39026. [Google Scholar] [CrossRef] [Green Version]
- Schröder, J.; Harder, J. Human Beta-Defensin-2. Int. J. Biochem. Cell Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef]
- Tu, Y.; Ho, Y.; Chuang, Y.; Chen, P.; Chen, C. Identification of Lactoferricin B Intracellular Targets using an Escherichia coli Proteome Chip. PLoS ONE 2011, 6, e28197. [Google Scholar] [CrossRef] [Green Version]
- Wang, G. Human Antimicrobial Peptides and Proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [Green Version]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Basañez, G.; Shinnar, A.E.; Zimmerberg, J. Interaction of Hagfish Cathelicidin Antimicrobial Peptides with Model Lipid Membranes. FEBS Lett. 2002, 532, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Broekman, D.C.; Zenz, A.; Gudmundsdottir, B.K.; Lohner, K.; Maier, V.H.; Gudmundsson, G.H. Functional Characterization of codCath, the Mature Cathelicidin Antimicrobial Peptide from Atlantic Cod (Gadus Morhua). Peptides 2011, 32, 2044–2051. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Zhang, Y.; Zou, J.; Nie, P.; Secombes, C.J. Two Cathelicidin Genes are Present in both Rainbow Trout (Oncorhynchus mykiss) and Atlantic Salmon (Salmo salar). Antimicrob. Agents Chemother. 2006, 50, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruangsri, J.; Kitani, Y.; Kiron, V.; Lokesh, J.; Brinchmann, M.F.; Karlsen, B.O.; Fernandes, J.M. A Novel Beta-Defensin Antimicrobial Peptide in Atlantic Cod with Stimulatory Effect on Phagocytic Activity. PLoS ONE 2013, 8, e62302. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and Characterization of Pleurocidin, an Antimicrobial Peptide in the Skin Secretions of Winter Flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diep, D.B.; Nes, I.F. Ribosomally Synthesized Antibacterial Peptides in Gram Positive Bacteria. Curr. Drug Targets 2002, 3, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Stansly, P.G.; Shepherd, R.G.; White, H.J. Polymyxin: A New Chemotherapeutic Agent. Bull. Johns Hopkins Hosp. 1947, 81, 43–54. [Google Scholar] [PubMed]
- Velkov, T.; Thompson, P.E.; Azad, M.A.; Roberts, K.D.; Bergen, P.J. History, Chemistry and Antibacterial Spectrum. In Polymyxin Antibiotics: From Laboratory Bench to Bedside; Springer: Cham, Switzerland, 2019; pp. 15–36. [Google Scholar]
- Riley, M.A.; Chavan, M.A. Bacteriocins; Springer: Cham, Switzerland, 2007. [Google Scholar]
- Johnstone, B.A.; Christie, M.P.; Morton, C.J.; Parker, M.W. X-ray crystallography shines a light on pore-forming toxins. Methods Enzymol. 2021, 649, 1–46. [Google Scholar]
- Gillor, O.; Kirkup, B.C.; Riley, M.A. Colicins and Microcins: The Next Generation Antimicrobials. Adv. Appl. Microbiol. 2004, 54, 129–146. [Google Scholar]
- Papadakos, G.; Wojdyla, J.A.; Kleanthous, C. Nuclease Colicins and their Immunity Proteins. Q. Rev. Biophys. 2012, 45, 57–103. [Google Scholar] [CrossRef]
- Ramachander Turaga, V. Peptaibols: Antimicrobial Peptides from Fungi. In Bioactive Natural Products in Drug Discovery; Springer: Singapore, 2020; pp. 713–730. [Google Scholar]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventós, D.S. Plectasin, a Fungal Defensin, Targets the Bacterial Cell Wall Precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Gao, B.; Harvey, P.J.; Craik, D.J. Dermatophytic Defensin with Antiinfective Potential. Proc. Natl. Acad. Sci. USA 2012, 109, 8495–8500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A Venom-Derived Peptide with Promising Anti-Viral Properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Pastagia, M.; Schuch, R.; Fischetti, V.A.; Huang, D.B. Lysins: The Arrival of Pathogen-Directed Anti-Infectives. J. Med. Microbiol. 2013, 62, 1506–1516. [Google Scholar] [CrossRef] [PubMed]
- Plotka, M.; Kapusta, M.; Dorawa, S.; Kaczorowska, A.; Kaczorowski, T. Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses 2019, 11, 657. [Google Scholar] [CrossRef] [Green Version]
- Saier, M.H., Jr.; Reddy, B.L. Holins in Bacteria, Eukaryotes, and Archaea: Multifunctional Xenologues with Potential Biotechnological and Biomedical Applications. J. Bacteriol. 2015, 197, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Niu, W.; Wu, R.; Wang, J.; Lei, L.; Han, W.; Gu, J. The Phage Holin HolGH15 Exhibits Potential as an Antibacterial Agent to Control Listeria Monocytogenes. Foodborne Pathog. Dis. 2021, 18, 574–581. [Google Scholar] [CrossRef]
- Dutta, P.; Sahu, R.K.; Dey, T.; Lahkar, M.D.; Manna, P.; Kalita, J. Beneficial Role of Insect-Derived Bioactive Components against Inflammation and its Associated Complications (Colitis and Arthritis) and Cancer. Chem. Biol. Interact. 2019, 313, 108824. [Google Scholar] [CrossRef]
- Zahedifard, F.; Lee, H.; No, J.H.; Salimi, M.; Seyed, N.; Asoodeh, A.; Rafati, S. Comparative Study of Different Forms of Jellein Antimicrobial Peptide on Leishmania Parasite. Exp. Parasitol. 2020, 209, 107823. [Google Scholar] [CrossRef]
- Wang, G. Structures of Human Host Defense Cathelicidin LL-37 and its Smallest Antimicrobial Peptide KR-12 in Lipid Micelles. J. Biol. Chem. 2008, 283, 32637–32643. [Google Scholar] [CrossRef] [Green Version]
- Dürr, U.H.; Sudheendra, U.; Ramamoorthy, A. LL-37, the Only Human Member of the Cathelicidin Family of Antimicrobial Peptides. Biochim. Biophys. Acta (BBA)—Biomembr. 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, J.G. Phagocytin: A Bactericidal Substance from Polymorphonuclear Leucocytes. J. Exp. Med. 1956, 103, 589–611. [Google Scholar] [CrossRef] [PubMed]
- Zeya, H.I.; Spitznagel, J.K. Cationic Proteins of Polymorphonuclear Leukocyte Lysosomes. I. Resolution of Antibacterial and Enzymatic Activities. J. Bacteriol. 1966, 91, 750–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gschwandtner, M.; Zhong, S.; Tschachler, A.; Mlitz, V.; Karner, S.; Elbe-Bürger, A.; Mildner, M. Fetal Human Keratinocytes Produce Large Amounts of Antimicrobial Peptides: Involvement of Histone-Methylation Processes. J. Investig. Dermatol. 2014, 134, 2192–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J. A Peptide Antibiotic from Human Skin. Nature 1997, 387, 861. [Google Scholar] [CrossRef]
- Trend, S.; Strunk, T.; Hibbert, J.; Kok, C.H.; Zhang, G.; Doherty, D.A.; Richmond, P.; Burgner, D.; Simmer, K.; Davidson, D.J. Antimicrobial Protein and Peptide Concentrations and Activity in Human Breast Milk Consumed by Preterm Infants at Risk of Late-Onset Neonatal Sepsis. PLoS ONE 2015, 10, e0117038. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A. The Role of Amphibian Antimicrobial Peptides in Protection of Amphibians from Pathogens Linked to Global Amphibian Declines. Biochim. Biophys. Acta (BBA)—Biomembr. 2009, 1788, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.F.; Bui-Marinos, M.P.; Katzenback, B.A. Frog Skin Innate Immune Defences: Sensing and Surviving Pathogens. Front. Immunol. 2019, 9, 3128. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, Y.; Wang, X.; Liang, J.; Zhang, C.; Zhang, K.; Lin, G.; Lai, R. The First Antimicrobial Peptide from Sea Amphibian. Mol. Immunol. 2008, 45, 678–681. [Google Scholar] [CrossRef]
- Masso-Silva, J.A.; Diamond, G. Antimicrobial Peptides from Fish. Pharmaceuticals 2014, 7, 265–310. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Mercier, C.; Koussounadis, A.; Secombes, C. Discovery of Multiple Beta-Defensin Like Homologues in Teleost Fish. Mol. Immunol. 2007, 44, 638–647. [Google Scholar] [CrossRef]
- Guo, M.; Wei, J.; Huang, X.; Huang, Y.; Qin, Q. Antiviral Effects of Β-Defensin Derived from Orange-Spotted Grouper (Epinephelus coioides). Fish Shellfish. Immunol. 2012, 32, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Cai, J.; Liu, H.; Fan, D.; Peng, H.; Wang, K. Recombinant Medaka (Oryzias melastigmus) Pro-Hepcidin: Multifunctional Characterization. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 161, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.M.; Ruangsri, J.; Kiron, V. Atlantic Cod Piscidin and its Diversification through Positive Selection. PLoS ONE 2010, 5, e9501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; de la Fuente-Nunez, C.; Ou, R.W.; Torres, M.D.T.; Pande, S.G.; Sinskey, A.J.; Lu, T.K. Yeast-Based Synthetic Biology Platform for Antimicrobial Peptide Production. ACS Synth. Biol. 2018, 7, 896–902. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [Green Version]
- Brötz, H.; Sahl, H. New Insights into the Mechanism of Action of Lantibiotics—Diverse Biological Effects by Binding to the Same Molecular Target. J. Antimicrob. Chemother. 2000, 46, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Devi, S.M.; Halami, P.M. Detection and Characterization of Pediocin PA-1/AcH Like Bacteriocin Producing Lactic Acid Bacteria. Curr. Microbiol. 2011, 63, 181–185. [Google Scholar] [CrossRef]
- Cintas, L.M.; Casaus, P.; Havarstein, L.S.; Hernandez, P.E.; Nes, I.F. Biochemical and Genetic Characterization of Enterocin P, a Novel Sec-Dependent Bacteriocin from Enterococcus faecium P13 with a Broad Antimicrobial Spectrum. Appl. Environ. Microbiol. 1997, 63, 4321–4330. [Google Scholar] [CrossRef] [Green Version]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. Structure and Mode-of-Action of the Two-Peptide (Class-IIb) Bacteriocins. Probiotics Antimicrob. Proteins 2010, 2, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Rea, M.C.; Ross, R.P.; Cotter, P.D.; Hill, C. Classification of Bacteriocins from Gram-Positive Bacteria. In Prokaryotic Antimicrobial Peptides; Springer: New York, NY, USA, 2011; pp. 29–53. [Google Scholar]
- Heng, N.C.; Tagg, J.R. What’s in a Name? Class Distinction for Bacteriocins. Nat. Rev. Microbiol. 2006, 4, 160. [Google Scholar] [CrossRef]
- Da Silva Sabo, S.; Vitolo, M.; González, J.M.D.; de Souza Oliveira, R.P. Overview of Lactobacillus Plantarum as a Promising Bacteriocin Producer among Lactic Acid Bacteria. Food Res. Int. 2014, 64, 527–536. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, E.; Alves, V.; Franco, B. Fundamentals and Perspectives for the use of Bacteriocins Produced by Lactic Acid Bacteria in Meat Products. Food Rev. Int. 2002, 18, 191–208. [Google Scholar] [CrossRef]
- Bann, S.J.; Ballantine, R.D.; Cochrane, S.A. The Tridecaptins: Non-Ribosomal Peptides that Selectively Target Gram-Negative Bacteria. RSC Med. Chem. 2021, 12, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parret, A.H.; De Mot, R. Bacteria Killing their Own Kind: Novel Bacteriocins of Pseudomonas and Other Γ-Proteobacteria. Trends Microbiol. 2002, 10, 107–112. [Google Scholar] [CrossRef]
- Denkovskienė, E.; Paškevičius, Š.; Misiūnas, A.; Stočkūnaitė, B.; Starkevič, U.; Vitkauskienė, A.; Hahn-Löbmann, S.; Schulz, S.; Giritch, A.; Gleba, Y. Broad and Efficient Control of Klebsiella Pathogens by Peptidoglycan-Degrading and Pore-Forming Bacteriocins Klebicins. Sci. Rep. 2019, 9, 15422. [Google Scholar] [CrossRef] [Green Version]
- Michel-Briand, Y.; Baysse, C. The Pyocins of Pseudomonas aeruginosa. Biochimie 2002, 84, 499–510. [Google Scholar] [CrossRef]
- Bissett, J.; Gams, W.; Jaklitsch, W.; Samuels, G.J. Accepted Trichoderma Names in the Year 2015. IMA Fungus 2015, 6, 263–295. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.; Reusser, F. A Polypeptide Antibacterial Agent Isolated from Trichoderma viride. Experientia 1967, 23, 85–86. [Google Scholar] [CrossRef]
- Chugh, J.; Wallace, B. Peptaibols: Models for Ion Channels. Biochem. Soc. Trans. 2001, 29, 565–570. [Google Scholar] [CrossRef]
- Chugh, J.; Brückner, H.; Wallace, B. Model for a Helical Bundle Channel Based on the High-Resolution Crystal Structure of trichotoxin_A50E. Biochemistry 2002, 41, 12934–12941. [Google Scholar] [CrossRef] [PubMed]
- Grishin, D.; Sokolov, N. Defensins are Natural Peptide Antibiotics of Higher Eukaryotes. Biochem. Suppl. Ser. B Biomed. Chem. 2014, 8, 11–18. [Google Scholar] [CrossRef]
- Nyanguile, O. Peptide Antiviral Strategies as an Alternative to Treat Lower Respiratory Viral Infections. Front. Immunol. 2019, 10, 1366. [Google Scholar] [CrossRef] [PubMed]
- Goodsell, D.S. Illustrations of the HIV Life Cycle. In The Future of HIV-1 Therapeutics; Springer: Cham, Switzerland, 2015; pp. 243–252. [Google Scholar]
- Mulder, K.; Lima, L.A.; Miranda, V.; Dias, S.C.; Franco, O.L. Current Scenario of Peptide-Based Drugs: The Key Roles of Cationic Antitumor and Antiviral Peptides. Front. Microbiol. 2013, 4, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matanic, V.C.A.; Castilla, V. Antiviral Activity of Antimicrobial Cationic Peptides against Junin Virus and Herpes Simplex Virus. Int. J. Antimicrob. Agents 2004, 23, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.H.; Jenssen, H.; Sandvik, K.; Gutteberg, T.J. Anti-HSV Activity of Lactoferrin and Lactoferricin Is Dependent on the Presence of Heparan Sulphate at the Cell Surface. J. Med. Virol. 2004, 74, 262–271. [Google Scholar] [CrossRef]
- Manohar, P.; Loh, B.; Namasivayam, E.; Loganathan, A.; Nachimuthu, R.; Leptihn, S. A Multiwell-Plate Caenorhabditis elegans Assay for Assessing the Therapeutic Potential of Bacteriophages against Clinical Pathogens. Microbiol. Spectr. 2022, 23, e0139321. [Google Scholar] [CrossRef]
- Mirski, T.; Mizak, L.; Nakonieczna, A.; Gryko, R. Bacteriophages, Phage Endolysins and Antimicrobial Peptides-the Possibilities for their Common use to Combat Infections and in the Design of New Drugs. Ann. Agric. Environ. Med. 2019, 26, 203–209. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage Endolysins as Novel Antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Parisien, A.; Allain, B.; Zhang, J.; Mandeville, R.; Lan, C. Novel Alternatives to Antibiotics: Bacteriophages, Bacterial Cell Wall Hydrolases, and Antimicrobial Peptides. J. Appl. Microbiol. 2008, 104, 1–13. [Google Scholar] [CrossRef]
- Gondil, V.S.; Harjai, K.; Chhibber, S. Endolysins as Emerging Alternative Therapeutic Agents to Counter Drug-Resistant Infections. Int. J. Antimicrob. Agents 2020, 55, 105844. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Donovan, D.M. Antimicrobial Bacteriophage-Derived Proteins and Therapeutic Applications. Bacteriophage 2015, 5, e1062590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briers, Y.; Volckaert, G.; Cornelissen, A.; Lagaert, S.; Michiels, C.W.; Hertveldt, K.; Lavigne, R. Muralytic Activity and Modular Structure of the Endolysins of Pseudomonas aeruginosa Bacteriophages φKZ and EL. Mol. Microbiol. 2007, 65, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; You, R.; Lai, M.; Lin, N.; Chen, L.; Chang, K. Highly Potent Antimicrobial Modified Peptides Derived from the Acinetobacter baumannii Phage Endolysin LysAB2. Sci. Rep. 2017, 7, 11477. [Google Scholar] [CrossRef] [Green Version]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A. Identification of a Broadly Active Phage Lytic Enzyme with Lethal Activity against Antibiotic-Resistant Enterococcus Faecalis and Enterococcus faecium. J. Bacteriol. 2004, 186, 4808–4812. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; Donovan, D.M.; García, P. Enhanced Staphylolytic Activity of the Staphylococcus aureus Bacteriophage vB_SauS-phiIPLA88 HydH5 Virion-Associated Peptidoglycan Hydrolase: Fusions, Deletions, and Synergy with LysH5. Appl. Environ. Microbiol. 2012, 78, 2241–2248. [Google Scholar] [CrossRef] [Green Version]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-Encoded Virion-Associated Enzymes to Overcome the Carbohydrate Barriers during the Infection Process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [Green Version]
- Caldentey, J.; Bamford, D.H. The Lytic Enzyme of the Pseudomonas Phage Φ6. Purification and Biochemical Characterization. Biochim. Biophys. Acta (BBA)—Protein Struct. Mol. Enzymol. 1992, 1159, 44–50. [Google Scholar] [CrossRef]
- Leiman, P.G.; Molineux, I.J. Evolution of a New Enzyme Activity from the Same Motif Fold. Mol. Microbiol. 2008, 69, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, A.; Ceyssens, P.; Krylov, V.N.; Noben, J.; Volckaert, G.; Lavigne, R. Identification of EPS-Degrading Activity within the Tail Spikes of the Novel Pseudomonas Putida Phage AF. Virology 2012, 434, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Lin, T.; Lin, Y.; Su, P.; Chen, C.; Hsieh, P.; Hsu, C.; Chen, C.; Hsieh, Y.; Wang, J. Identification of Capsular Types in Carbapenem-Resistant Klebsiella Pneumoniae Strains by Wzc Sequencing and Implications for Capsule Depolymerase Treatment. Antimicrob. Agents Chemother. 2015, 59, 1038–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Lin, T.; Chen, C.; Tsai, Y.; Cheng, Y.; Chen, Y.; Hsieh, P.; Lin, Y.; Wang, J. Klebsiella Phage ΦK64–1 Encodes Multiple Depolymerases for Multiple Host Capsular Types. J. Virol. 2017, 91, e02457-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, L.; Gorman, S.P.; Gilmore, B.F. The use of Lytic Bacteriophages in the Prevention and Eradication of Biofilms of Proteus Mirabilis and Escherichia coli. FEMS Immunol. Med. Microbiol. 2010, 59, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibeu, A.; Lingohr, E.J.; Masson, L.; Manges, A.; Harel, J.; Ackermann, H.; Kropinski, A.M.; Boerlin, P. Bacteriophages with the Ability to Degrade Uropathogenic Escherichia Coli Biofilms. Viruses 2012, 4, 471–487. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; Lu, C.; Sun, J.; Yan, Y. Application of a Bacteriophage Lysin to Disrupt Biofilms Formed by the Animal Pathogen Streptococcus Suis. Appl. Environ. Microbiol. 2011, 77, 8272–8279. [Google Scholar] [CrossRef] [Green Version]
- Topka-Bielecka, G.; Dydecka, A.; Necel, A.; Bloch, S.; Nejman-Faleńczyk, B.; Węgrzyn, G.; Węgrzyn, A. Bacteriophage-Derived Depolymerases against Bacterial Biofilm. Antibiotics 2021, 10, 175. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as Weapons against Bacterial Biofilms in the Food Industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef]
- Wang, I.; Smith, D.L.; Young, R. Holins: The Protein Clocks of Bacteriophage Infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Vilcinskas, A. Evolutionary Plasticity of Insect Immunity. J. Insect Physiol. 2013, 59, 123–129. [Google Scholar] [CrossRef]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Buonocore, F.; Fausto, A.M.; Pelle, G.D.; Roncevic, T.; Gerdol, M.; Picchietti, S. Attacins: A Promising Class of Insect Antimicrobial Peptides. Antibiotics 2021, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Swain, S.S.; Behera, A.; Sahoo, G.; Mahapatra, P.K.; Panda, S.K. Antimicrobial Peptides Derived from Insects Offer a Novel Therapeutic Option to Combat Biofilm: A Review. Front. Microbiol. 2021, 12, 661195. [Google Scholar] [CrossRef] [PubMed]
- Shirtliff, M.E.; Peters, B.M.; Jabra-Rizk, M.A. Cross-Kingdom Interactions: Candida albicans and Bacteria. FEMS Microbiol. Lett. 2009, 299, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coquant, G.; Grill, J.; Seksik, P. Impact of N-Acyl-Homoserine Lactones, Quorum Sensing Molecules, on Gut Immunity. Front. Immunol. 2020, 11, 1827. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial Biofilm Development as a Multicellular Adaptation: Antibiotic Resistance and New Therapeutic Strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Rice, L.B. Federal Funding for the Study of Antimicrobial Resistance in Nosocomial Pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- Mogasale, V.V.; Saldanha, P.; Pai, V.; Rekha, P.; Mogasale, V. A Descriptive Analysis of Antimicrobial Resistance Patterns of WHO Priority Pathogens Isolated in Children from a Tertiary Care Hospital in India. Sci. Rep. 2021, 11, 5116. [Google Scholar] [CrossRef]
- Eckhard, L.H.; Sol, A.; Abtew, E.; Shai, Y.; Domb, A.J.; Bachrach, G.; Beyth, N. Biohybrid Polymer-Antimicrobial Peptide Medium Against Enterococcus Faecalis. PLoS ONE 2014, 9, e109413. [Google Scholar] [CrossRef] [Green Version]
- Mergoni, G.; Manfredi, M.; Bertani, P.; Ciociola, T.; Conti, S.; Giovati, L. Activity of Two Antimicrobial Peptides against Enterococcus faecalis in a Model of Biofilm-Mediated Endodontic Infection. Antibiotics 2021, 10, 1220. [Google Scholar] [CrossRef] [PubMed]
- Oyama, L.B.; Crochet, J.; Edwards, J.E.; Girdwood, S.E.; Cookson, A.R.; Fernandez-Fuentes, N.; Hilpert, K.; Golyshin, P.N.; Golyshina, O.V.; Privé, F. Buwchitin: A Ruminal Peptide with Antimicrobial Potential against Enterococcus Faecalis. Front. Chem. 2017, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Hsueh, J.; Yip, B.; Chih, Y.; Peng, K.; Cheng, J. Antimicrobial Peptides Display Strong Synergy with Vancomycin against Vancomycin-Resistant, E. Faecium, S. Aureus, and Wild-Type E. coli. Int. J. Mol. Sci. 2020, 21, 4578. [Google Scholar] [CrossRef] [PubMed]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; de Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; van der Heijde, T.; Boekema, B.K. The Antimicrobial Peptide SAAP-148 Combats Drug-Resistant Bacteria and Biofilms. Sci. Transl. Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef] [Green Version]
- Dean, S.N.; Bishop, B.M.; Van Hoek, M.L. Natural and Synthetic Cathelicidin Peptides with Anti-Microbial and Anti-Biofilm Activity against Staphylococcus aureus. BMC Microbiol. 2011, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Herbert, S.; Bera, A.; Nerz, C.; Kraus, D.; Peschel, A.; Goerke, C.; Meehl, M.; Cheung, A.; Götz, F. Molecular Basis of Resistance to Muramidase and Cationic Antimicrobial Peptide Activity of Lysozyme in Staphylococci. PLoS Pathog. 2007, 3, e102. [Google Scholar] [CrossRef]
- Herpers, B. Endolysins: Redefining Antibacterial Therapy. Future Microbiol. 2015, 10, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The Preclinical and Clinical Progress of Bacteriophages and their Lytic Enzymes: The Parts are Easier than the Whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Fleeman, R.M.; Macias, L.A.; Brodbelt, J.S.; Davies, B.W. Defining Principles that Influence Antimicrobial Peptide Activity against Capsulated Klebsiella pneumoniae. Proc. Natl. Acad. Sci. USA 2020, 117, 27620–27626. [Google Scholar] [CrossRef]
- Cebrián, R.; Xu, C.; Xia, Y.; Wu, W.; Kuipers, O.P. The Cathelicidin-Derived Close-to-Nature Peptide D-11 Sensitises Klebsiella pneumoniae to a Range of Antibiotics In Vitro, Ex Vivo and In Vivo. Int. J. Antimicrob. Agents 2021, 58, 106434. [Google Scholar] [CrossRef]
- Ribeiro, S.M.; De La Fuente-Núñez, C.; Baquir, B.; Faria-Junior, C.; Franco, O.L.; Hancock, R.E. Antibiofilm Peptides Increase the Susceptibility of Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates to Β-Lactam Antibiotics. Antimicrob. Agents Chemother. 2015, 59, 3906–3912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente-Núñez, C.; Reffuveille, F.; Haney, E.F.; Straus, S.K.; Hancock, R.E. Broad-Spectrum Anti-Biofilm Peptide that Targets a Cellular Stress Response. PLoS Pathog. 2014, 10, e1004152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial Activity of Selected Antimicrobial Peptides against Planktonic Culture and Biofilm of Acinetobacter baumannii. Probiotics Antimicrob. Proteins 2019, 11, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wu, Z.; Mao, C.; Guo, G.; Zeng, Z.; Fei, Y.; Wan, S.; Peng, J.; Wu, J. Antimicrobial Peptide Cec4 Eradicates the Bacteria of Clinical Carbapenem-Resistant Acinetobacter baumannii Biofilm. Front. Microbiol. 2020, 11, 1532. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Sambanthamoorthy, K.; Palys, T.; Paranavitana, C. The Human Antimicrobial Peptide LL-37 and its Fragments Possess both Antimicrobial and Antibiofilm Activities against Multidrug-Resistant Acinetobacter baumannii. Peptides 2013, 49, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Liao, Y.; Hsu, C.; Huang, T.; Chuang, Y.; Chen, J.; Kuo, Y.; Chia, J. Identification of Potential Therapeutic Antimicrobial Peptides against Acinetobacter baumannii in a Mouse Model of Pneumonia. Sci. Rep. 2021, 11, 7318. [Google Scholar] [CrossRef]
- Huang, G.; Shen, X.; Gong, Y.; Dong, Z.; Zhao, X.; Shen, W.; Wang, J.; Hu, F.; Peng, Y. Antibacterial Properties of Acinetobacter baumannii Phage Abp1 Endolysin (PlyAB1). BMC Infect. Dis. 2014, 14, 681. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Hu, K.; Xie, Y.; Liu, Y.; Mu, D.; Guo, H.; Zhang, Z.; Zhang, Y.; Chang, D.; Shi, Y. A Novel Phage PD-6A3, and its Endolysin Ply6A3, with Extended Lytic Activity against Acinetobacter baumannii. Front. Microbiol. 2019, 9, 3302. [Google Scholar] [CrossRef]
- Lood, R.; Winer, B.Y.; Pelzek, A.J.; Diez-Martinez, R.; Thandar, M.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Novel Phage Lysin Capable of Killing the Multidrug-Resistant Gram-Negative Bacterium Acinetobacter baumannii in a Mouse Bacteremia Model. Antimicrob. Agents Chemother. 2015, 59, 1983–1991. [Google Scholar] [CrossRef] [Green Version]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.; Rehm, B.H.; Hancock, R.E. Human Host Defense Peptide LL-37 Prevents Bacterial Biofilm Formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, C.; Lin, X.; Yi, G.; Zhang, Y.; Rowe-Magnus, D.A.; Bush, K. Cathelicidin Antimicrobial Peptides with Reduced Activation of Toll-Like Receptor Signaling have Potent Bactericidal Activity against Colistin-Resistant Bacteria. mBio 2016, 7, e01418-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hase, K.; Murakami, M.; Iimura, M.; Cole, S.P.; Horibe, Y.; Ohtake, T.; Obonyo, M.; Gallo, R.L.; Eckmann, L.; Kagnoff, M.F. Expression of LL-37 by Human Gastric Epithelial Cells as a Potential Host Defense Mechanism against Helicobacter pylori. Gastroenterology 2003, 125, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Yu, J.; Zhang, L.; Wu, W. The Role of Cathelicidin in Control of Helicobacter Pylori Colonisation in the Stomach. Hong Kong Med. J. 2015, 21 (Suppl. 4), 17–19. [Google Scholar]
- Chow, J.Y.C.; Li, Z.J.; Wu, W.K.K.; Cho, C.H. Cathelicidin a Potential Therapeutic Peptide for Gastrointestinal Inflammation and Cancer. World J. Gastroenterol. WJG 2013, 19, 2731. [Google Scholar] [CrossRef]
- Narayana, J.L.; Huang, H.N.; Wu, C.J.; Chen, J.Y. Efficacy of the Antimicrobial Peptide TP4 against Helicobacter pylori Infection: In Vitro Membrane Perturbation Via Micellization and In Vivo Suppression of Host Immune Responses in a Mouse Model. Oncotarget 2015, 6, 12936–12954. [Google Scholar] [CrossRef] [Green Version]
- Leszczyńska, K.; Namiot, A.; Fein, D.E.; Wen, Q.; Namiot, Z.; Savage, P.B.; Diamond, S.; Janmey, P.A.; Bucki, R. Bactericidal Activities of the Cationic Steroid CSA-13 and the Cathelicidin Peptide LL-37 against Helicobacter pylori in Simulated Gastric Juice. BMC Microbiol. 2009, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Guzman, J.; Téné, N.; Touchard, A.; Castillo, D.; Belkhelfa, H.; Haddioui-Hbabi, L.; Treilhou, M.; Sauvain, M. Anti-Helicobacter pylori Properties of the Ant-Venom Peptide Bicarinalin. Toxins 2018, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Windham, I.H.; Servetas, S.L.; Whitmire, J.M.; Pletzer, D.; Hancock, R.E.; Merrell, D.S. Helicobacter pylori Biofilm Formation Is Differentially Affected by Common Culture Conditions, and Proteins Play a Central Role in the Biofilm Matrix. Appl. Environ. Microbiol. 2018, 84, e00391-18. [Google Scholar] [CrossRef] [Green Version]
- Elgamoudi, B.A.; Korolik, V. Campylobacter Biofilms: Potential of Natural Compounds to Disrupt Campylobacter jejuni Transmission. Int. J. Mol. Sci. 2021, 22, 12159. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Turner, K.L.; Crockett, T.M.; Lu, X.; Morris, C.F.; Konkel, M.E. Inhibitory Effect of Puroindoline Peptides on Campylobacter jejuni Growth and Biofilm Formation. Front. Microbiol. 2021, 12, 702762. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Oh, J.H.; Kang, H.K.; Choi, M.; Seo, C.H.; Park, Y. Scorpion-Venom-Derived Antimicrobial Peptide Css54 Exerts Potent Antimicrobial Activity by Disrupting Bacterial Membrane of Zoonotic Bacteria. Antibiotics 2020, 9, 831. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, S.A.; Lohans, C.T.; Brandelli, J.R.; Mulvey, G.; Armstrong, G.D.; Vederas, J.C. Synthesis and Structure–activity Relationship Studies of N-Terminal Analogues of the Antimicrobial Peptide Tridecaptin A1. J. Med. Chem. 2014, 57, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, A.; Herrebout, M.; Tersteeg-Zijderveld, M.H.; Tjeerdsma-van Bokhoven, J.L.; Bleumink-Pluym, N.; Jansman, A.J.; Veldhuizen, E.J.; Haagsman, H.P. Campylobacter jejuni Is Highly Susceptible to Killing by Chicken Host Defense Peptide Cathelicidin-2 and Suppresses Intestinal Cathelicidin-2 Expression in Young Broilers. Vet. Microbiol. 2012, 160, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical Relevance of the ESKAPE Pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic Resistance of Bacteria in Biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Taylor, T.A.; Unakal, C.G. Staphylococcus aureus. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2022. [Google Scholar]
- Jun, S.Y.; Jung, G.M.; Yoon, S.J.; Oh, M.; Choi, Y.; Lee, W.J.; Kong, J.; Seol, J.G.; Kang, S.H. Antibacterial Properties of a Pre-Formulated Recombinant Phage Endolysin, SAL-1. Int. J. Antimicrob. Agents 2013, 41, 156–161. [Google Scholar] [CrossRef]
- Ashurst, J.V.; Dawson, A. Klebsiella pneumonia; StatPearls Publishing LLC: Treasure Island, FL, USA, 2018. [Google Scholar]
- Bellich, B.; Lagatolla, C.; Tossi, A.; Benincasa, M.; Cescutti, P.; Rizzo, R. Influence of Bacterial Biofilm Polysaccharide Structure on Interactions with Antimicrobial Peptides: A Study on Klebsiella pneumoniae. Int. J. Mol. Sci. 2018, 19, 1685. [Google Scholar] [CrossRef] [Green Version]
- Llobet, E.; March, C.; Giménez, P.; Bengoechea, J.A. Klebsiella pneumoniae OmpA Confers Resistance to Antimicrobial Peptides. Antimicrob. Agents Chemother. 2009, 53, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Lopes, B.S.; Al-Agamy, M.H.; Ismail, M.A.; Shibl, A.M.; Al-Qahtani, A.A.; Al-Ahdal, M.N.; Forbes, K.J. The Transferability of blaOXA-23 Gene in Multidrug-Resistant Acinetobacter baumannii Isolates from Saudi Arabia and Egypt. Int. J. Med. Microbiol. 2015, 305, 581–588. [Google Scholar] [CrossRef]
- Akbari, R.; Hakemi Vala, M.; Hashemi, A.; Aghazadeh, H.; Sabatier, J.; Pooshang Bagheri, K. Action Mechanism of Melittin-Derived Antimicrobial Peptides, MDP1 and MDP2, De Novo Designed against Multidrug Resistant Bacteria. Amino Acids 2018, 50, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Akbari, R.; Hakemi-Vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Pooshang Bagheri, K. Highly Synergistic Effects of Melittin with Conventional Antibiotics against Multidrug-Resistant Isolates of Acinetobacter baumannii and Pseudomonas aeruginosa. Microb. Drug Resist. 2019, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieprawska-Lupa, M.; Mydel, P.; Krawczyk, K.; Wójcik, K.; Puklo, M.; Lupa, B.; Suder, P.; Silberring, J.; Reed, M.; Pohl, J. Degradation of Human Antimicrobial Peptide LL-37 by Staphylococcus Aureus-Derived Proteinases. Antimicrob. Agents Chemother. 2004, 48, 4673–4679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidtchen, A.; Frick, I.; Andersson, E.; Tapper, H.; Björck, L. Proteinases of Common Pathogenic Bacteria Degrade and Inactivate the Antibacterial Peptide LL-37. Mol. Microbiol. 2002, 46, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, T.; Sharma, P.K.; Busscher, H.J.; Van Der Mei, H.C.; Krom, B.P. Role of Extracellular DNA in Initial Bacterial Adhesion and Surface Aggregation. Appl. Environ. Microbiol. 2010, 76, 3405–3408. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.N.; Klein, D.R.; Kazi, M.I.; Guérin, F.; Cattoir, V.; Brodbelt, J.S.; Boll, J.M. Colistin Heteroresistance in Enterobacter cloacae Is Regulated by PhoPQ-dependent 4-amino-4-deoxy-l-arabinose Addition to Lipid A. Mol. Microbiol. 2019, 111, 1604–1616. [Google Scholar] [CrossRef]
- Soren, O.; Brinch, K.S.; Patel, D.; Liu, Y.; Liu, A.; Coates, A.; Hu, Y. Antimicrobial Peptide Novicidin Synergizes with Rifampin, Ceftriaxone, and Ceftazidime against Antibiotic-Resistant Enterobacteriaceae In Vitro. Antimicrob. Agents Chemother. 2015, 59, 6233–6240. [Google Scholar] [CrossRef] [Green Version]
- Nuding, S.; Gersemann, M.; Hosaka, Y.; Konietzny, S.; Schaefer, C.; Beisner, J.; Schroeder, B.O.; Ostaff, M.J.; Saigenji, K.; Ott, G. Gastric Antimicrobial Peptides Fail to Eradicate Helicobacter pylori Infection due to Selective Induction and Resistance. PLoS ONE 2013, 8, e73867. [Google Scholar] [CrossRef] [Green Version]
- Téné, N.; Roche-Chatain, V.; Rifflet, A.; Bonnafé, E.; Lefranc, B.; Leprince, J.; Treilhou, M. Potent Bactericidal Effects of Bicarinalin against Strains of the Enterobacter and Cronobacter Genera. Food Control 2014, 42, 202–206. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, A.; Qi, B.; Yu, H.; Xiong, Y.; Zhou, G.; Qin, M.; Dou, J.; Wang, J. Secretion Expression of Human Neutrophil Peptide 1 (HNP1) in Pichia Pastoris and its Functional Analysis against Antibiotic-Resistant Helicobacter pylori. Appl. Microbiol. Biotechnol. 2018, 102, 4817–4827. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.K.; Dallas, J.F.; Strachan, N.J.; MacRae, M.; McCarthy, N.D.; Wilson, D.J.; Gormley, F.J.; Falush, D.; Ogden, I.D.; Maiden, M.C. Campylobacter Genotyping to Determine the Source of Human Infection. Clin. Infect. Dis. 2009, 48, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sproston, E.L.; Wimalarathna, H.M.; Sheppard, S.K. Trends in Fluoroquinolone Resistance in Campylobacter. Microb. Genom. 2018, 4, e000198. [Google Scholar] [CrossRef] [PubMed]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus Subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Lohans, C.T.; van Belkum, M.J.; Li, J.; Vederas, J.C. Characterization of Bacterial Antimicrobial Peptides Active against Campylobacter jejuni. Can. J. Chem. 2015, 93, 381–388. [Google Scholar] [CrossRef]
- Rodríguez-Rojas, A.; Baeder, D.Y.; Johnston, P.; Regoes, R.R.; Rolff, J. Bacteria Primed by Antimicrobial Peptides Develop Tolerance and Persist. PLoS Pathog. 2021, 17, e1009443. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| References | Antimicrobial Peptide | Secondary Structure | Source | Function/Mode of Action |
|---|---|---|---|---|
| [17] | Protegrins | α-helix and β-sheet | Porcine |
|
| [17] | Thanatin | Loop structure | Insect (Podisus maculiventris) |
|
| [19,20] | Tachyplesins | β-sheet | Horseshoe crab |
|
| [22] | Bactenecin | Loop structure | Bovine |
|
| [35] | Human neutrophil peptides (HNP 1-4) | β-sheet | Human |
|
| [36,37] | Human β-defensins (HBD 1-4) | β-sheet | Human |
|
| [38] | Lactoferricin B | Loop structure | Human |
|
| [39] | Cathelidicin (LL-37) | α-helix | Human |
|
| [39] | Human defensin (HD5-6) | α-helix | Human |
|
| [40] | Magainin-2 | α-helix | Frog |
|
| [41] | hagfish intestinal AMP (HFIAP)-1, -2, and -3 | α-helix | Myxine glutinosa (Atlantic hagfish) |
|
| [42,43] | cathelicidin codCATH | α-helix | Atlantic Cod (Gadus morhua) |
|
| [42,43] | rtCATH1 (R146-P181) and rtCATH2 (R143-I178) | α-helix | Rainbow Trout |
|
| [44] | cod-defensin | β-sheet | Atlantic cod |
|
| [45] | Pleurocidin | α-helix | winter flounder (Pleuronectes americanus) |
|
| [46] | Gramicidin | β-sheet | Gram-positive bacteria (Lactococcus lactis and Bacillus brevis) |
|
| [47,48] | Polymyxin | α-helix | Paenibacillus polymyxa |
|
| [49] | Bacteriocin | α-helix | Gram-positive bacteria, Gram-negative bacteria |
|
| [50,51,52] | Colicins | α-helix and β-sheet | E. coli |
|
| [53] | Peptaibols | α-helix | Fungal |
|
| [54] | Plectasin | α-helix | Fungal (Pseudoplectania nigrella) |
|
| [55] | Micasin | α-helix | Microsporum canis |
|
| [56] | Melittin | α-helix | European honeybee Apis mellifera venom |
|
| [57,58] | Phage lysins | α-helix | Bacteriophages |
|
| [59] | Holins | α-helix | dsDNA bacteriophages |
|
| [60] | HolGH15 | α-helix | S. aureus bacteriophage GH15 |
|
| [61] | Cecropin A | α-helix | Cecropia moth (Hyalophora cecropia) and bees |
|
| [62] | Jellein | α-helix | Honey bee royal jelly |
|
| Bacterial Species | Antimicrobial Peptides | References |
|---|---|---|
| Enterococcus faecalis | C16-KGGK KP and L18R (antifungal peptides) Buwchitin P-113 Bip-P-113 SAAP-148 (LL-37 derivative) | [136] [137] [138] [139] [140] |
| Staphylococcus aureus | NA-CATH:ATRA1-ATRA1 human lysozyme-derived peptide (LP9) polymyxin B gallidermin phage-derived bacteriocins SAAP-148 (LL-37 derivative) | [141] [142] [142] [142] [143,144] [140] |
| Klebsiella pneumoniae | Analog PepC (A6, A12, and A19) cathelicidin-derived peptide D-11 DJK-6 IDR-1018 citropin 1.1 CAMEL SAAP-148 (LL-37 derivative) | [145] [146] [147] [148] [149] [140] |
| Acinetobacter baumannii | Cec4 LL-37 SMAP-29 Tilapia Piscidin 4 (TP4) Combination of melittin and doripenem plyAB1 ply6A3 plyF307 SAAP-148 (LL-37 derivative) | [150] [151] [152] [152] [153] [154] [155] [140] |
| Pseudomonas aeruginosa | Lactoferrin LL-37 LysPA26 SAAP-148 (LL-37 derivative) | [129] [156] [157] |
| Enterobacter spp. | BMAP-27B SMAP-29D SAAP-148 (LL-37 derivative) | [158] [158] [140] |
| Helicobacter pylori | Cathelicidin mCRAMP Tilapia Piscidin 4 (TP4) ceragenin CSA-13 bicarinalin IDR-1018 DJK-5 | [159,160,161] [162] [163] [164] [165] [165] |
| Campylobacter spp. | puroindoline A (PinA) Css54 (Scorpion-venom-derived AMP) Tridecaptins chicken cathelicidin-2 (CATH-2) | [166,167] [168] [169] [170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, B.S.; Hanafiah, A.; Nachimuthu, R.; Muthupandian, S.; Md Nesran, Z.N.; Patil, S. The Role of Antimicrobial Peptides as Antimicrobial and Antibiofilm Agents in Tackling the Silent Pandemic of Antimicrobial Resistance. Molecules 2022, 27, 2995. https://doi.org/10.3390/molecules27092995
Lopes BS, Hanafiah A, Nachimuthu R, Muthupandian S, Md Nesran ZN, Patil S. The Role of Antimicrobial Peptides as Antimicrobial and Antibiofilm Agents in Tackling the Silent Pandemic of Antimicrobial Resistance. Molecules. 2022; 27(9):2995. https://doi.org/10.3390/molecules27092995
Chicago/Turabian StyleLopes, Bruno S., Alfizah Hanafiah, Ramesh Nachimuthu, Saravanan Muthupandian, Zarith Nameyrra Md Nesran, and Sandip Patil. 2022. "The Role of Antimicrobial Peptides as Antimicrobial and Antibiofilm Agents in Tackling the Silent Pandemic of Antimicrobial Resistance" Molecules 27, no. 9: 2995. https://doi.org/10.3390/molecules27092995