
The Hydrolytic Peptides of Soybean Protein Induce Cell Cycle Arrest and Apoptosis on Human Oral Cancer Cell Line HSC-3

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction of Gelatin from Fish Skins
2.3. Preparation of Protein Hydrolysates
2.4. Cell Culture
2.5. MTT Assay
2.6. Western Blotting
2.7. Cell Cycle Analysis
2.8. Apoptosis Analysis
2.9. Statistical Analysis
3. Results and Discussion
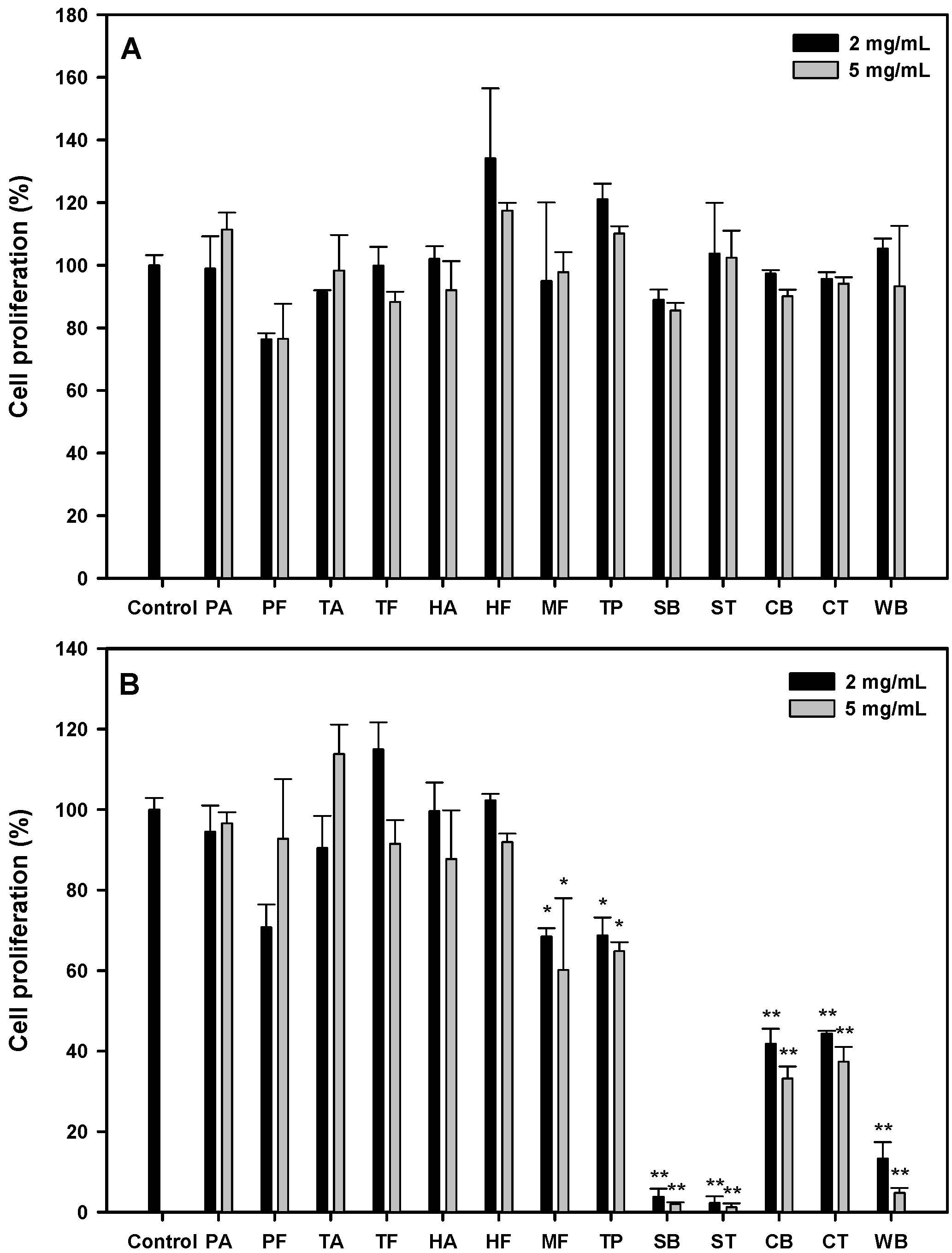
3.1. Antiproliferative Activities of Protein Hydrolysates
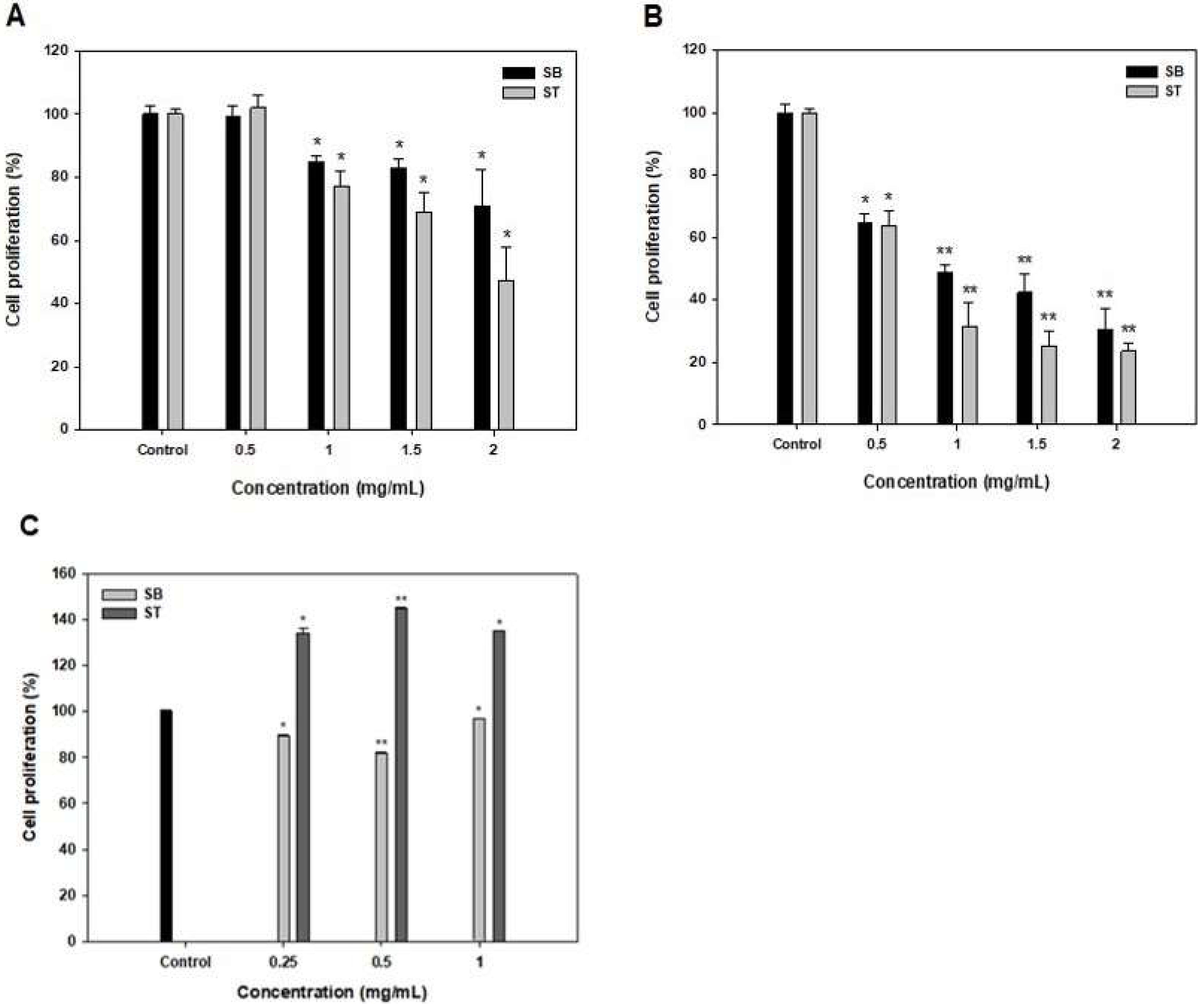
3.2. Antiproliferative Activity of SB and ST
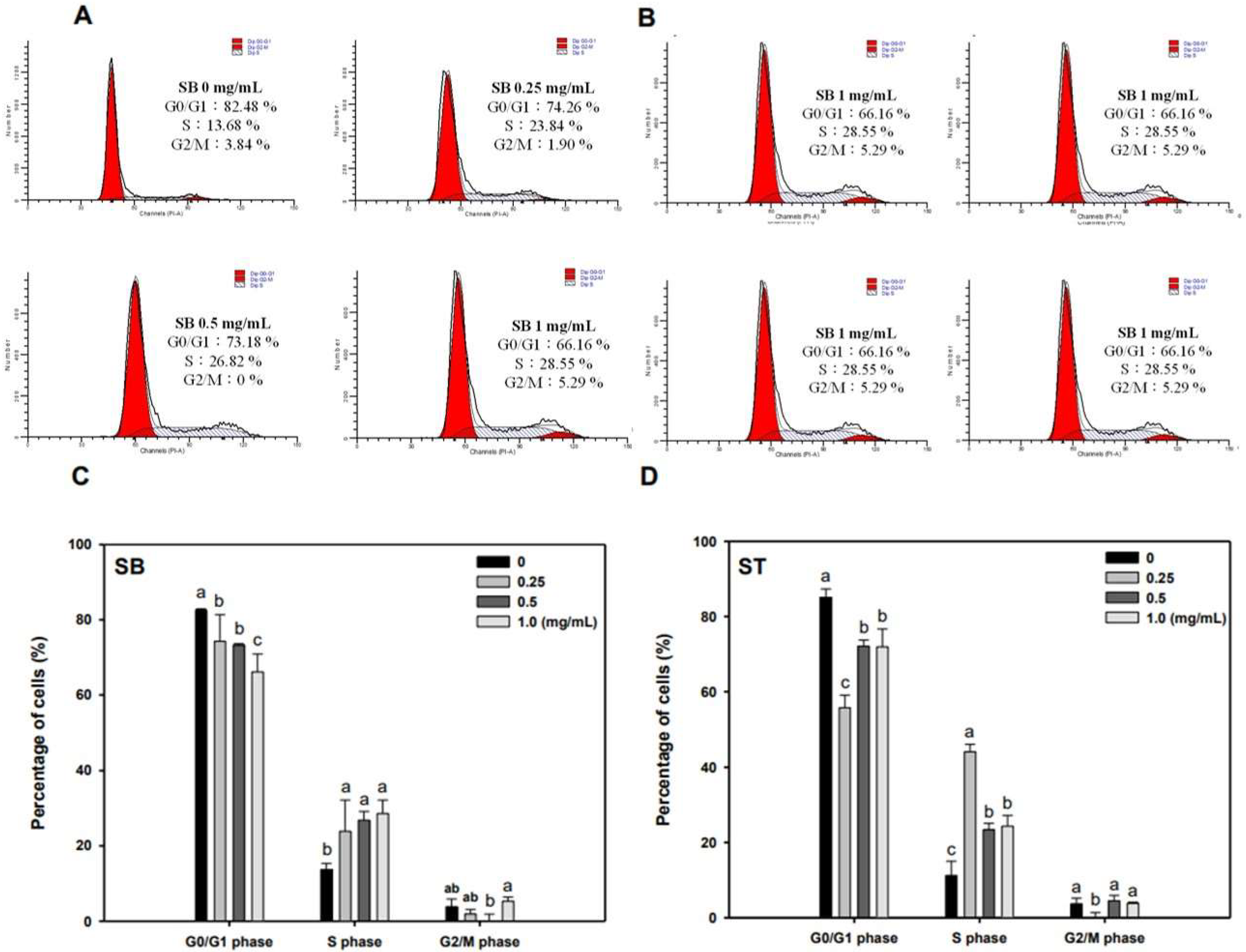
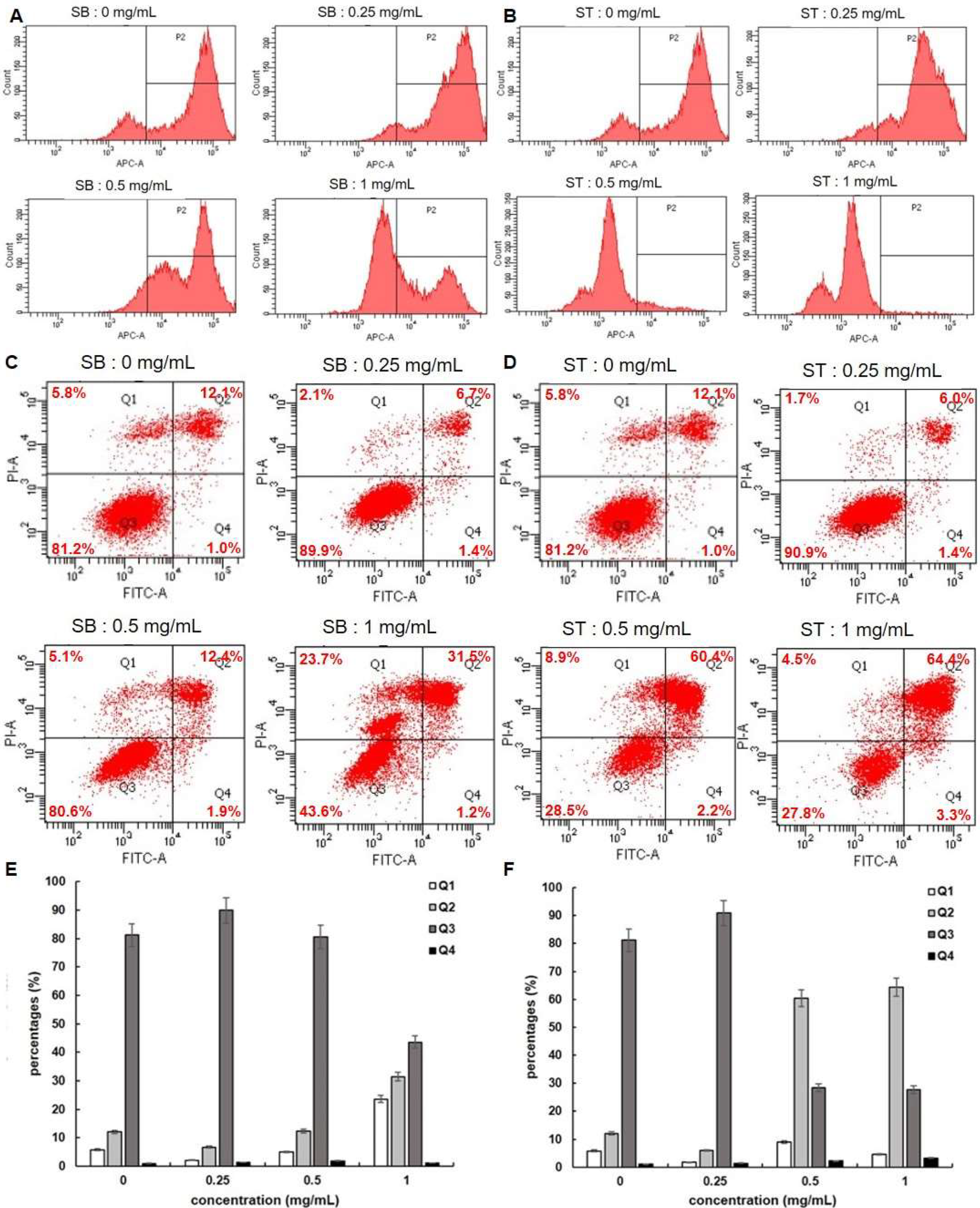
3.3. Cell Cycle
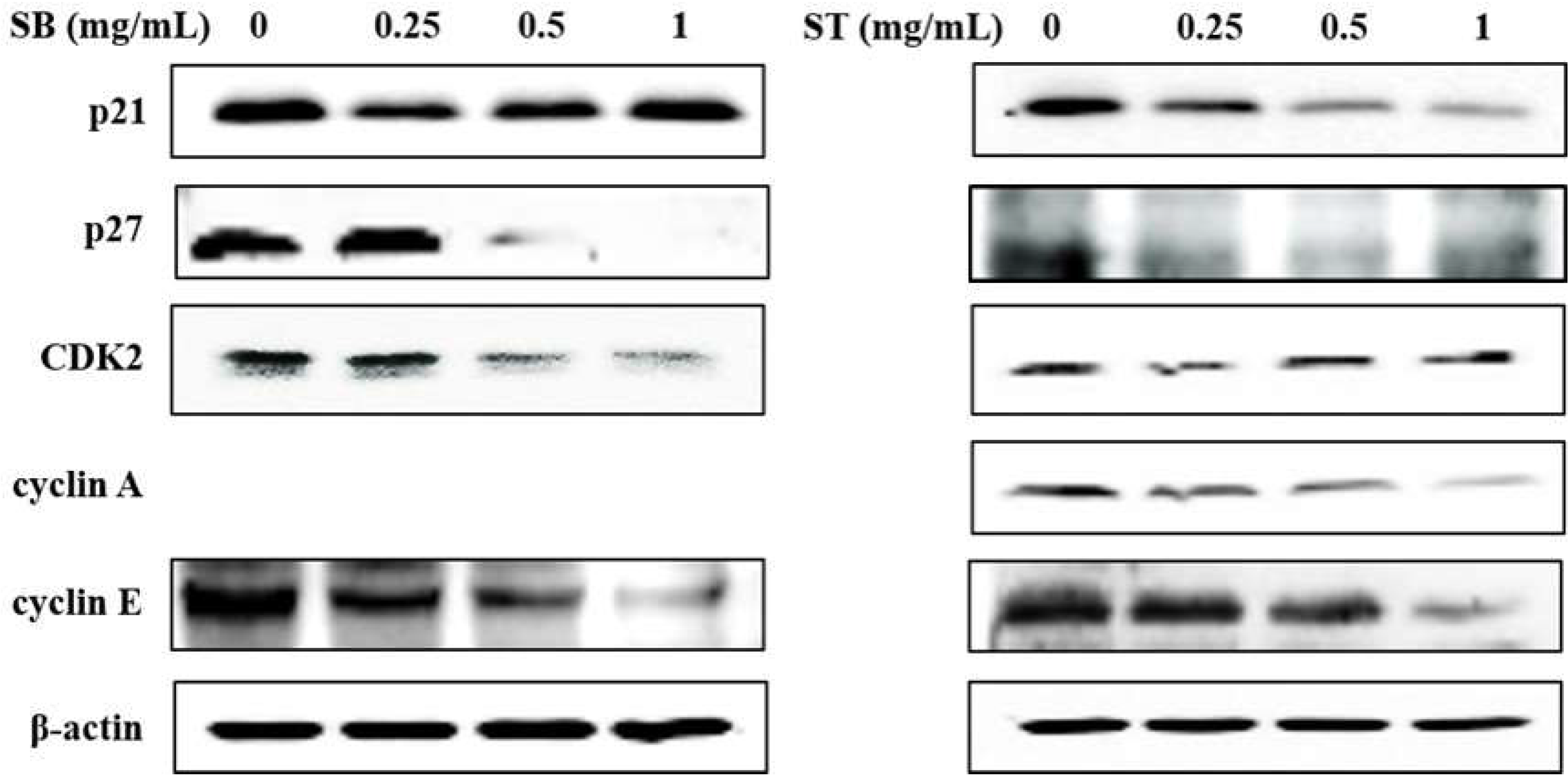
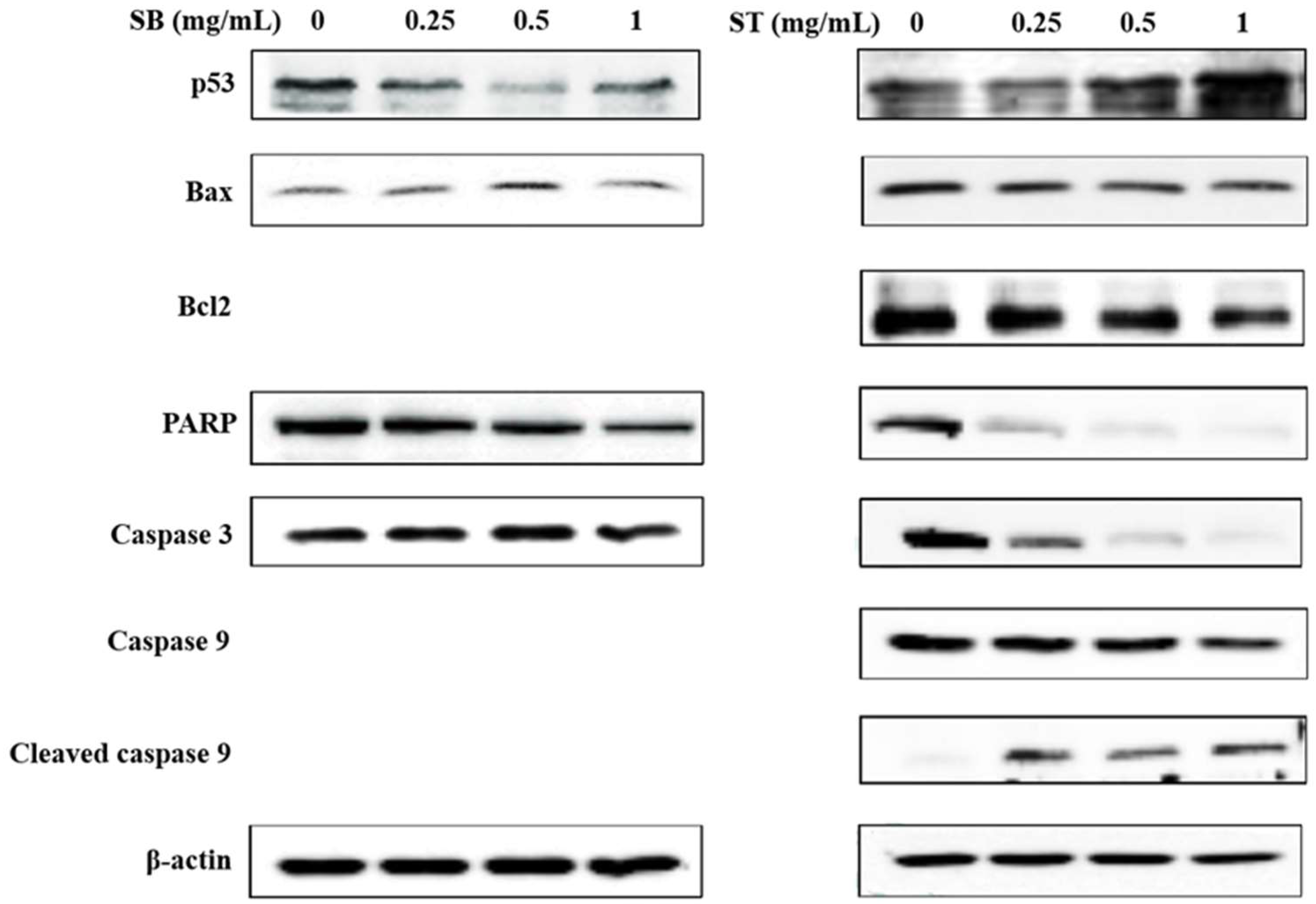
3.4. Apoptosis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tanaka, T.; Tanaka, M.; Tanaka, T. Oral carcinogenesis and oral cancer chemoprevention: A review. Pathol. Res. Int. 2011, 2011, 431246. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Saxena, M.; Srinivas, K.; Singh, V.K. Fucose: A biomarker in grading of oral cancer. Natl. J. Maxillofac. Surg. 2015, 6, 176. [Google Scholar] [PubMed] [Green Version]
- Yen, C.-Y.; Hou, M.-F.; Yang, Z.-W.; Tang, J.-Y.; Li, K.-T.; Huang, H.-W.; Huang, Y.-H.; Lee, S.-Y.; Fu, T.-F.; Hsieh, C.-Y. Concentration effects of grape seed extracts in anti-oral cancer cells involving differential apoptosis, oxidative stress, and DNA damage. BMC Complement. Altern. Med. 2015, 15, 94. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.-C.; Tseng, C.-N.; Yang, J.-I.; Huang, H.-W.; Fang, Y.; Tang, J.-Y.; Chang, F.-R.; Chang, H.-W. Antiproliferation and induction of apoptosis in Ca9-22 oral cancer cells by ethanolic extract of Gracilaria tenuistipitata. Molecules 2012, 17, 10916–10927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.-C.; Yang, J.-I.; Lee, J.-C.; Tseng, C.-N.; Chan, Y.-C.; Hseu, Y.-C.; Tang, J.-Y.; Chuang, L.-Y.; Huang, H.-W.; Chang, F.-R. Anti-proliferative effect of methanolic extract of Gracilaria tenuistipitata on oral cancer cells involves apoptosis, DNA damage, and oxidative stress. BMC Complement. Altern. Med. 2012, 12, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, Y.-H.; Farooqi, A.A.; Li, K.-T.; Butt, G.; Tang, J.-Y.; Wu, C.-Y.; Cheng, Y.-B.; Hou, M.-F.; Chang, H.-W. Methanolic extracts of Solieria robusta inhibits proliferation of oral cancer Ca9-22 cells via apoptosis and oxidative stress. Molecules 2014, 19, 18721–18732. [Google Scholar] [CrossRef] [Green Version]
- Yoo, Y.C.; Watanabe, S.; Watanabe, R.; Hata, K.; Shimazaki, K.I.; Azuma, I. Bovine lactoferrin and lactoferricin, a peptide derived from bovine lactoferrin, inhibit tumor metastasis in mice. Jpn. J. Cancer Res. 1997, 88, 184–190. [Google Scholar] [CrossRef]
- Armstrong, W.B.; Kennedy, A.R.; Wan, X.S.; Atiba, J.; McLaren, C.E.; Meyskens, F.L. Single-dose administration of Bowman-Birk inhibitor concentrate in patients with oral leukoplakia. Cancer Epidemiol. Prev. Biomark. 2000, 9, 43–47. [Google Scholar]
- Kim, S.E.; Kim, H.H.; Kim, J.Y.; Im Kang, Y.; Woo, H.J.; Lee, H.J. Anticancer activity of hydrophobic peptides from soy proteins. Biofactors 2000, 12, 151–155. [Google Scholar] [CrossRef]
- Kobayashi, H.; Suzuki, M.; Kanayama, N.; Terao, T. A soybean Kunitz trypsin inhibitor suppresses ovarian cancer cell invasion by blocking urokinase upregulation. Clin. Exp. Metastasis 2004, 21, 159–166. [Google Scholar] [CrossRef]
- Galvez, A.F.; Chen, N.; Macasieb, J.; Ben, O. Chemopreventive property of a soybean peptide (lunasin) that binds to deacetylated histones and inhibits acetylation. Cancer Res. 2001, 61, 7473–7478. [Google Scholar] [PubMed]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; de Lumen, B.O. Inhibition of core histone acetylation by the cancer preventive peptide lunasin. J. Agric. Food Chem. 2007, 55, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-C.; Li-Chan, E.C.; Jao, C.-L. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chem. 2011, 126, 617–622. [Google Scholar] [CrossRef]
- Hung, C.-C.; Yang, Y.-H.; Kuo, P.-F.; Hsu, K.-C. Protein hydrolysates from tuna cooking juice inhibit cell growth and induce apoptosis of human breast cancer cell line MCF-7. J. Funct. Foods 2014, 11, 563–570. [Google Scholar] [CrossRef]
- Yang, J.-I.; Tang, J.-Y.; Liu, Y.-S.; Wang, H.-R.; Lee, S.-Y.; Yen, C.-Y.; Chang, H.-W. Roe protein hydrolysates of giant grouper (Epinephelus lanceolatus) inhibit cell proliferation of oral cancer cells involving apoptosis and oxidative stress. BioMed Res. Int. 2016, 2016, 8305073. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Kaur, H.; Kehinde, B.A.; Chhikara, N.; Sharma, D.; Panghal, A. Food-derived anticancer peptides: A review. Int. J. Pept. Res. Ther. 2021, 27, 55–70. [Google Scholar] [CrossRef]
- Tyagi, A.; Kapoor, P.; Kumar, R.; Chaudhary, K.; Gautam, A.; Raghava, G. In silico models for designing and discovering novel anticancer peptides. Sci. Rep. 2013, 3, 2984. [Google Scholar] [CrossRef]
- Lee, Y.K.; Kim, S.Y.; Kim, K.H.; Chun, B.-H.; Lee, K.-H.; Oh, D.J.; Chung, N. Use of soybean protein hydrolysates for promoting proliferation of human keratinocytes in serum-free medium. Biotechnol. Lett. 2008, 30, 1931–1936. [Google Scholar] [CrossRef]
- Marcela, G.-M.; Eva, R.-G.; Del Carmen, R.-R.M.; Rosalva, M.-E. Evaluation of the antioxidant and antiproliferative effects of three peptide fractions of germinated soybeans on breast and cervical cancer cell lines. Plant Foods Hum. Nutr. 2016, 71, 368–374. [Google Scholar] [CrossRef]
- Freitas, R.D.; Dias, R.B.; Vidal, M.T.A.; Valverde, L.D.F.; Gomes Alves Costa, R.; Damasceno, A.K.A.; Sales, C.B.S.; Siquara da Rocha, L.D.O.; Dos Reis, M.G.; Soares, M.B.P. Inhibition of CAL27 oral squamous carcinoma cell by targeting hedgehog pathway with vismodegib or itraconazole. Front. Oncol. 2020, 10, 563838. [Google Scholar] [CrossRef]
- Chen, Z.; Li, W.; Santhanam, R.K.; Wang, C.; Gao, X.; Chen, Y.; Wang, C.; Xu, L.; Chen, H. Bioactive peptide with antioxidant and anticancer activities from black soybean [Glycine max (L.) Merr.] byproduct: Isolation, identification and molecular docking study. Eur. Food Res. Technol. 2019, 245, 677–689. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Lin, K.-C.; Lin, J.-P.; Tang, N.-Y.; Yang, J.-S.; Lu, K.-W.; Chung, J.-G. Phenethyl Isothiocyanate (PEITC) inhibits the growth of human oral squamous carcinoma HSC-3 cells through G0/G1 phase arrest and mitochondria-mediated apoptotic cell death. Evid.-Based Complement. Altern. Med. 2012, 2012, 718320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Liu, H.; Li, X. Orlistat treatment induces apoptosis and arrests cell cycle in HSC-3 oral cancer cells. Microb. Pathog. 2017, 112, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Choi, K.-M.; Kim, W.; Jeon, Y.-S.; Lee, Y.-M.; Hong, J.-T.; Yun, Y.-P.; Yoo, H.-S. Hinokitiol inhibits cell growth through induction of S-phase arrest and apoptosis in human colon cancer cells and suppresses tumor growth in a mouse xenograft experiment. J. Nat. Prod. 2013, 76, 2195–2202. [Google Scholar] [CrossRef]
- Lin, C.-C.; Yang, J.-S.; Chen, J.-T.; Fan, S.; Yu, F.-S.; Yang, J.-L.; Lu, C.-C.; Kao, M.-C.; Huang, A.-C.; Lu, H.-F. Berberine induces apoptosis in human HSC-3 oral cancer cells via simultaneous activation of the death receptor-mediated and mitochondrial pathway. Anticancer. Res. 2007, 27, 3371–3378. [Google Scholar]
- Hafidz, R.; Yaakob, C.; Amin, I.; Noorfaizan, A. Chemical and functional properties of bovine and porcine skin gelatin. Int. Food Res. J. 2011, 18, 787–791. [Google Scholar]
- Wang, T.-Y.; Hsieh, C.-H.; Hung, C.-C.; Jao, C.-L.; Chen, M.-C.; Hsu, K.-C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm-and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Rombouts, I.; Lamberts, L.; Celus, I.; Lagrain, B.; Brijs, K.; Delcour, J.A. Wheat gluten amino acid composition analysis by high-performance anion-exchange chromatography with integrated pulsed amperometric detection. J. Chromatogr. A 2009, 1216, 5557–5562. [Google Scholar] [CrossRef]
- Cervantes-Pahm, S.; Stein, H. Ileal digestibility of amino acids in conventional, fermented, and enzyme-treated soybean meal and in soy protein isolate, fish meal, and casein fed to weanling pigs. J. Anim. Sci. 2010, 88, 2674–2683. [Google Scholar] [CrossRef] [Green Version]
- Mackle, T.; Dwyer, D.; Bauman, D. Effects of branched-chain amino acids and sodium caseinate on milk protein concentration and yield from dairy cows. J. Dairy Sci. 1999, 82, 161–171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treated Time (h) | IC50 (mg/mL) | |
|---|---|---|
| SB | ST | |
| 48 | 4.01 | 1.69 |
| 72 | 0.74 | 0.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, C.-H.; Wang, T.-Y.; Tung, B.-C.; Liu, H.-P.; Yeh, L.-T.; Hsu, K.-C. The Hydrolytic Peptides of Soybean Protein Induce Cell Cycle Arrest and Apoptosis on Human Oral Cancer Cell Line HSC-3. Molecules 2022, 27, 2839. https://doi.org/10.3390/molecules27092839
Hsieh C-H, Wang T-Y, Tung B-C, Liu H-P, Yeh L-T, Hsu K-C. The Hydrolytic Peptides of Soybean Protein Induce Cell Cycle Arrest and Apoptosis on Human Oral Cancer Cell Line HSC-3. Molecules. 2022; 27(9):2839. https://doi.org/10.3390/molecules27092839
Chicago/Turabian StyleHsieh, Cheng-Hong, Tzu-Yuan Wang, Bo-Chen Tung, Hui-Ping Liu, Lien-Te Yeh, and Kuo-Chiang Hsu. 2022. "The Hydrolytic Peptides of Soybean Protein Induce Cell Cycle Arrest and Apoptosis on Human Oral Cancer Cell Line HSC-3" Molecules 27, no. 9: 2839. https://doi.org/10.3390/molecules27092839