
A Comprehensive Review on Therapeutic Perspectives of Phytosterols in Insulin Resistance: A Mechanistic Approach

 ,
,  , , , ,
, , , ,  , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
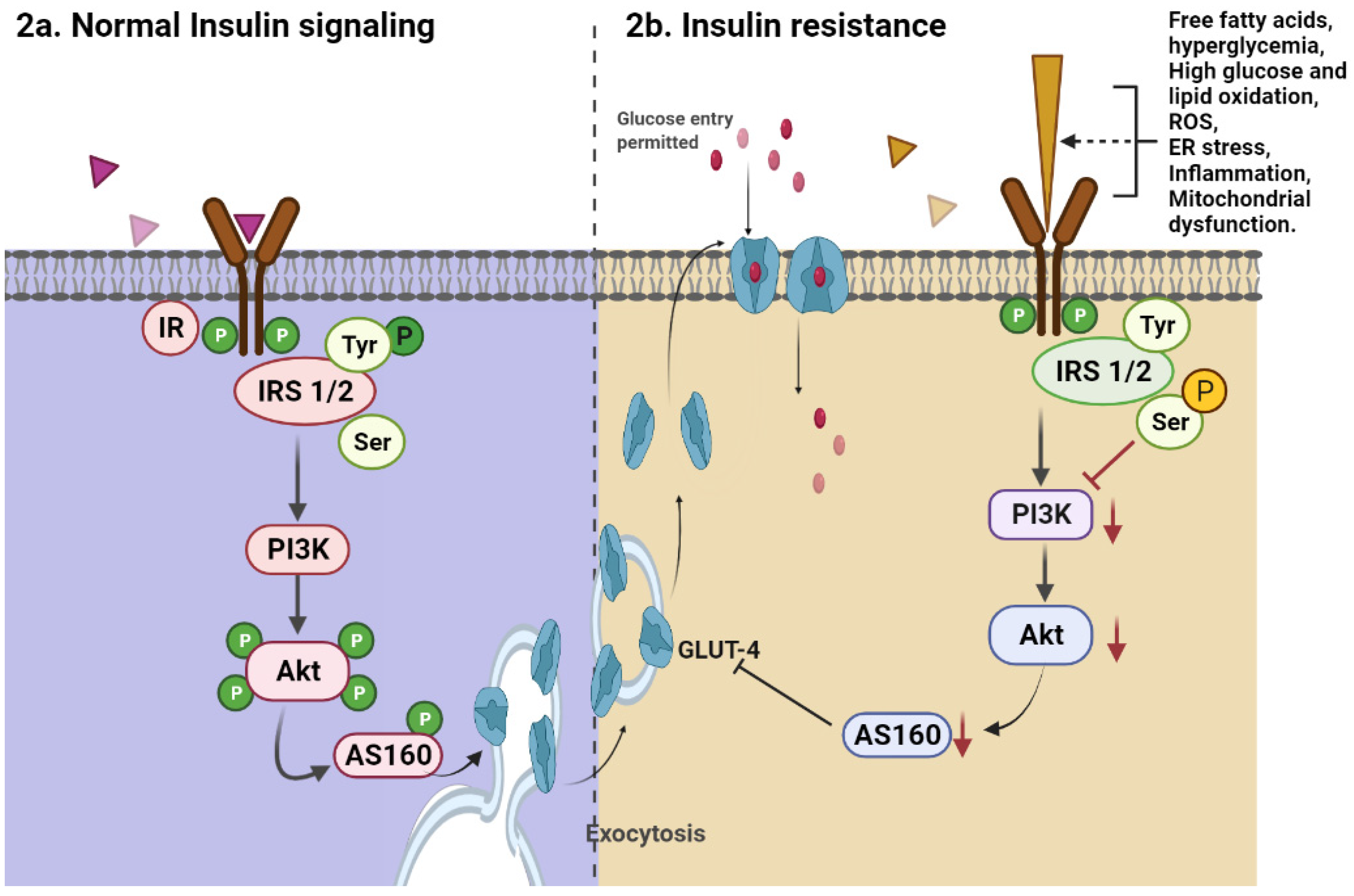
2. Insulin Pathophysiology
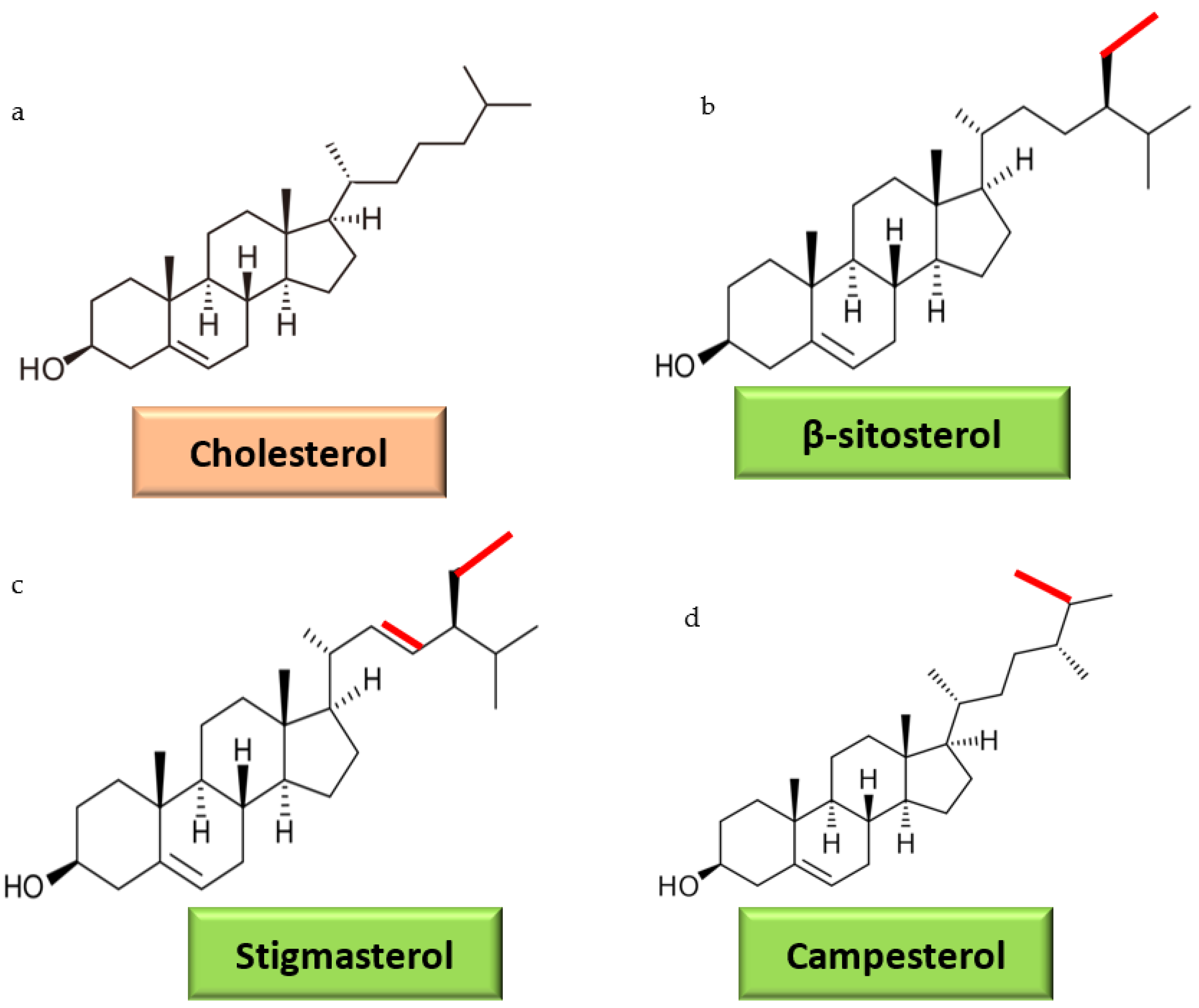
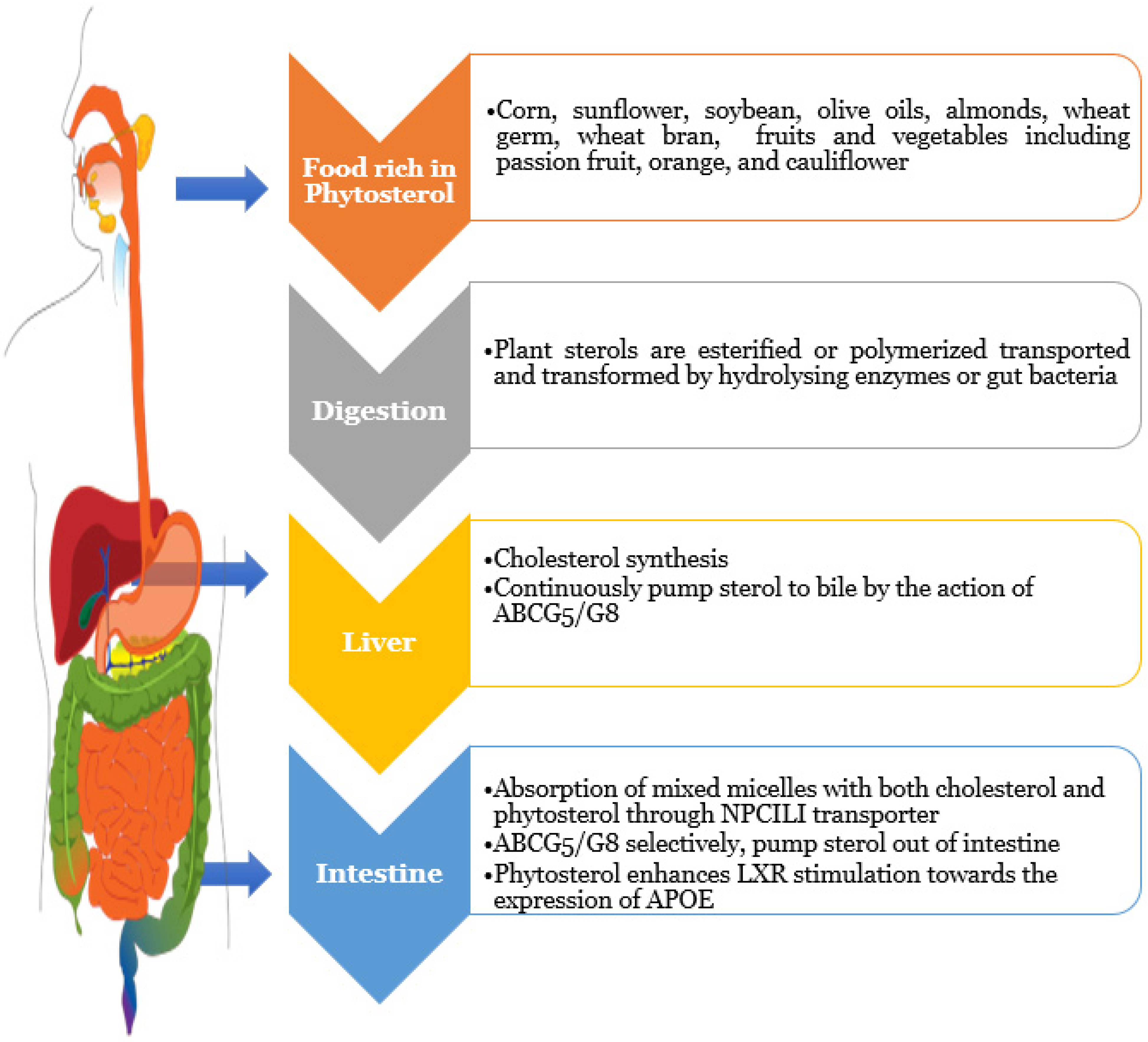
3. Dietary Sterols Overview
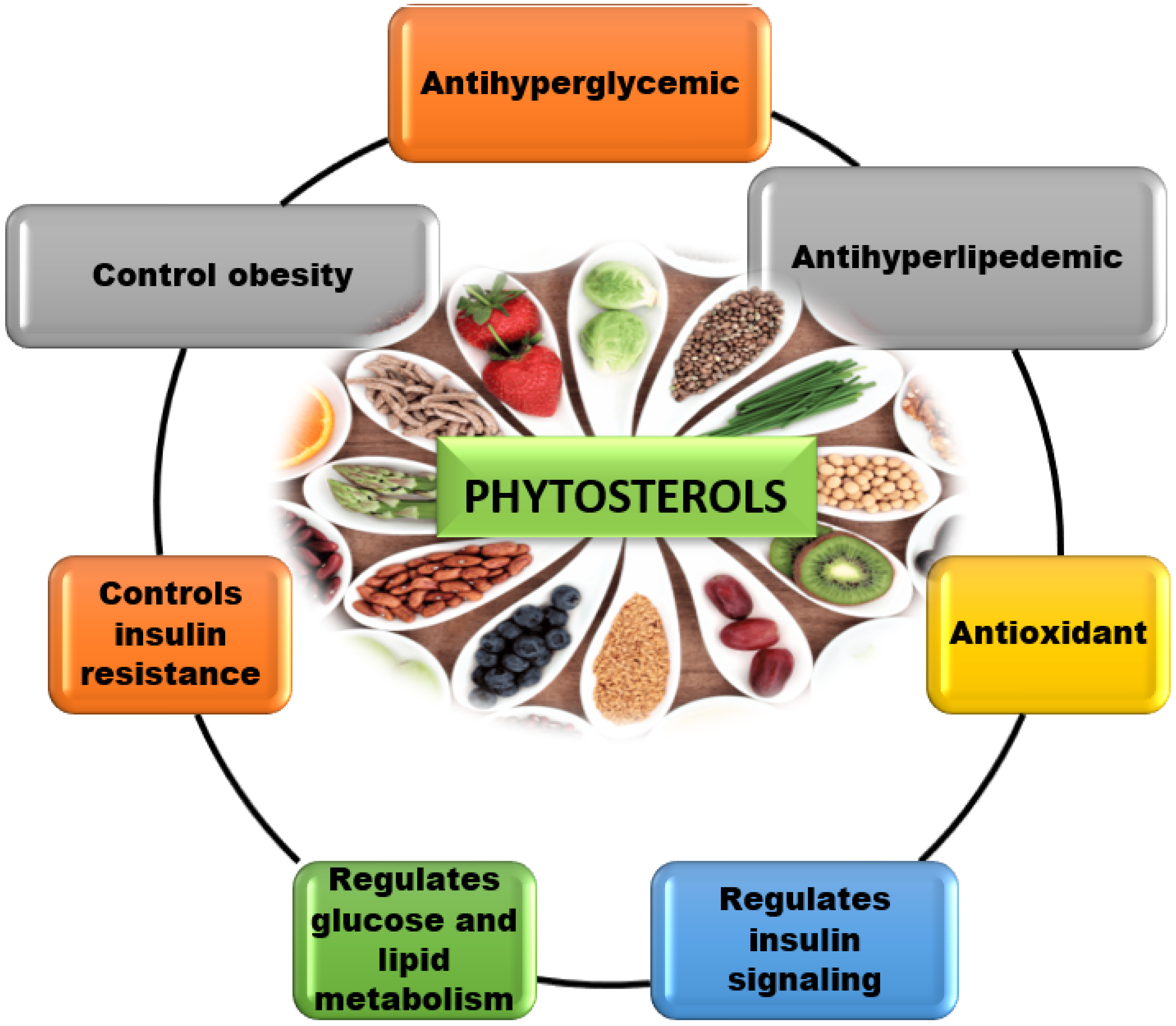
4. Therapeutic Functions of phytosterols
4.1. Reduces Intestinal Glucose Absorption
4.2. Reduces Intestinal Lipid Absorption
4.3. Alters Insulin Resistance in Tissues and Organs
4.4. Antidiabetic Activity of Phytosterol Derivatives
5. Conclusions and Prospective Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- IDF Diabetes Atlas 2021—10th Edition. Available online: https://diabetesatlas.org/idfawp/resource-files/2021/07/IDF_Atlas_10th_Edition_2021.pdf. (accessed on 10 February 2022).
- Lozano, I.; Werf, R.V.D.; Bietiger, W.; Seyfritz, E.; Peronet, C.; Pinget, M.; Jeandidier, N.; Maillard, E.; Marchioni, E.; Sigrist, S.; et al. High-fructose and high-fat diet-induced disorders in rats: Impact on diabetes risk, hepatic and vascular complications. Nutr. Metab. 2016, 13, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Tang, Y.M.; Yu, S.L.; Han, Y.W.; Kou, J.P.; Liu, B.L.; Yu, B.Y. Advances in the pharmacological activities and mechanisms of diosgenin. Chin. J. Nat. Med. 2015, 13, 578–587. [Google Scholar] [CrossRef]
- New WHO Guidelines to Promote Proper Use of Alternative Medicines. Available online: https://www.who.int/mediacentre/news/releases/2004/pr44/en/ (accessed on 28 November 2021).
- Roy, N.K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A.K.; Thakur, K.K.; Padmavathi, G.; Shakibaei, M.; Fan, L.; Sethi, G.; et al. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. Int. J. Mol. Sci. 2019, 20, 4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y. Artemisinin-A Gift from Traditional Chinese Medicine to the World (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2016, 55, 10210–10226. [Google Scholar] [CrossRef]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant Sterols: Diversity, Biosynthesis, and Physiological Functions. Biochem. Biokhimiia 2016, 81, 819–834. [Google Scholar] [CrossRef]
- Sharmila, R.; Sindhu, G. Evaluate the Antigenotoxicity and Anticancer Role of β-Sitosterol by Determining Oxidative DNA Damage and the Expression of Phosphorylated Mitogen-activated Protein Kinases’, C-fos, C-jun, and Endothelial Growth Factor Receptor. Pharmacogn. Mag. 2017, 13, 95–101. [Google Scholar]
- Pascual-Garrigos, A.; Maruthamuthu, M.K.; Ault, A.; Davidson, J.L.; Rudakov, G.; Pillai, D.; Koziol, J.; Schoonmaker, J.P.; Johnson, T.; Verma, M.S. On-farm colorimetric detection of Pasteurella multocida, Mannheimia haemolytica, and Histophilus somni in crude bovine nasal samples. Vet. Res. 2021, 52, 126. [Google Scholar] [CrossRef]
- Feng, S.; Gan, L.; Yang, C.S.; Liu, A.B.; Lu, W.; Shao, P.; Dai, Z.; Sun, P.; Luo, Z. Effects of Stigmasterol and β-Sitosterol on Nonalcoholic Fatty Liver Disease in a Mouse Model: A Lipidomic Analysis. J. Agric. Food Chem. 2018, 66, 3417–3425. [Google Scholar] [CrossRef]
- García, M.D.; Sáenz, M.T.; Gómez, M.A.; Fernández, M.A. Topical antiinflammatory activity of phytosterols isolated from Eryngium foetidum on chronic and acute inflammation models. Phytother. Res. PTR 1999, 13, 78–80. [Google Scholar] [CrossRef]
- Raju, L.; Lipin, R.; Eswaran, R. Identification, ADMET evaluation and molecular docking analysis of Phytosterols from Banaba (Lagerstroemia speciosa (L.)Pers) seed extract against breast cancer. Silico Pharmacol. 2021, 9, 43. [Google Scholar] [CrossRef]
- Jayaraman, S.; Devarajan, N.; Rajagopal, P.; Babu, S.; Ganesan, S.K.; Veeraraghavan, V.P.; Palanisamy, C.P.; Cui, B.; Periyasamy, V.; Chandrasekar, K. β-Sitosterol Circumvents Obesity Induced Inflammation and Insulin Resistance by down-Regulating IKK/NF-B and JNK Signaling Pathway in Adipocytes of Type 2 Diabetic Rats. Molecules 2021, 26, 2101. [Google Scholar] [CrossRef] [PubMed]
- Ponnulakshmi, R.; Shyamaladevi, B.; Vijayalakshmi, P.; Selvaraj, J. In silico and in vivo analysis to identify the antidiabetic activity of beta sitosterol in adipose tissue of high fat diet and sucrose induced type-2 diabetic experimental rats. Toxicol. Mech. Methods 2019, 29, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Babu, S.; Krishnan, M.; Rajagopal, P.; Periyasamy, V.; Veeraraghavan, V.; Govindan, R.; Jayaraman, S. Beta-sitosterol attenuates insulin resistance in adipose tissue via IRS-1/Akt mediated insulin signaling in high fat diet and sucrose induced type-2 diabetic rats. Eur. J. Pharmacol. 2020, 873, 173004. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.; Babu, S.; Rajagopal, P.; Nazar, S.P.; Chinnaiyan, M.; Jayaraman, S. Effect of β-sitosterol on Insulin Receptor, Glucose Transporter 4 Protein Expression and Glucose Oxidation in the Gastrocnemius Muscle of High Fat Diet Induced Type -2 Diabetic Experimental Rats. Indian J. Pharm. Educ. Res. 2021, 55, s479–s491. [Google Scholar] [CrossRef]
- Fagundes, M.B.; Alvarez-Rivera, G.; Mendiola, J.A.; Bueno, M.; Sánchez-Martínez, J.D.; Wagner, R.; Jacob-Lopes, E.; Zepka, L.Q.; Ibañez, E.; Cifuentes, A. Phytosterol-rich compressed fluids extracts from Phormidium autumnale cyanobacteria with neuroprotective potential. Algal Res. 2021, 55, 102264. [Google Scholar] [CrossRef]
- Wang, J.; Huang, M.; Yang, J.; Ma, X.; Zheng, S.; Deng, S.; Huang, Y.; Yang, X.; Zhao, P. Anti-diabetic activity of stigmasterol from soybean oil by targeting the GLUT4 glucose transporter. Food Nutr. Res. 2017, 61, 1364117. [Google Scholar] [CrossRef] [Green Version]
- Misawa, E.; Tanaka, M.; Nomaguchi, K.; Yamada, M.; Toida, T.; Takase, M.; Iwatsuki, K.; Kawada, T. Administration of phytosterols isolated from Aloe vera gel reduce visceral fat mass and improve hyperglycemia in Zucker diabetic fatty (ZDF) rats. Obes. Res. Clin. Pract. 2008, 2, I–II. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Hu, F.B.; Tapsell, L.C.; Josse, A.R.; Kendall, C.W. Possible benefit of nuts in type 2 diabetes. J. Nutr. 2008, 138, 1752S–1756S. [Google Scholar] [CrossRef] [Green Version]
- Tiganis, T. PTP1B and TCPTP--nonredundant phosphatases in insulin signaling and glucose homeostasis. FEBS J. 2013, 280, 445–458. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poitout, V.; Robertson, R.P. Minireview: Secondary E-cell failure in type 2 diabetes. A convergence of glucotoxicity and lipotoxicity. Endocrinology 2002, 143, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Polonsky, K.S.; Sturis, J.; Bell, G.I. Seminars in Medicine of the Beth Israel Hospital, Boston: Non-insulin-dependent diabetes mellitus—A genetically programmed failure of the beta cell to compensate for insulin resistance. N. Engl. J. Med. 1996, 334, 777–783. [Google Scholar] [CrossRef]
- Wei, L.; Hubbard, S.R.; Hendrickson, W.A.; Ellis, L. Expression, characterization, and crystallization of the catalytic core of the human insulin receptor protein-tyrosine kinase domain. J. Biol. Chem. 1995, 270, 8122–8130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E. The importance of E-cell failure in the development and progression of type 2 diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 4047–4058. [Google Scholar]
- Bergsten, P. Pathophysiology of impaired pulsatile insulin release. Diabetes Metab. Res. Rev. 2000, 16, 179–191. [Google Scholar] [CrossRef]
- Froguel, P.; Velho, G. Genetic determinants of type 2 diabetes. Recent Prog. Horm. Res. 2001, 56, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Maruthamuthu, M.K.; Ganesh, I.; Ravikumar, S.; Hong, S.H. Evaluation of zraP gene expression characteristics and construction of a lead (Pb) sensing and removal system in a recombinant Escherichia coli. Biotechnol. Lett. 2015, 37, 659–664. [Google Scholar] [CrossRef]
- Cavaghan, M.K.; Ehrmann, D.A.; Polonsky, K.S. Interactions between insulin resistance and insulin secretion in the development of glucose intolerance. J. Clin. Investig. 2000, 106, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Maruthamuthu, M.K.; Raffiee, A.H.; De Oliveira, D.M.; Ardekani, A.M.; Verma, M.S. Raman spectra-based deep learning: A tool to identify microbial contamination. Microbiol. Open 2020, 9, e1122. [Google Scholar] [CrossRef]
- Hansen, T. Genetics of type 2 diabetes. Curr. Sci. 2002, 83, 1477–1482. [Google Scholar]
- Ma, J.; Nakagawa, Y.; Kojima, I.; Shibata, H. Prolonged insulin stimulation down-regulates GLUT4 through oxidative stressmediated retromer inhibition by a protein kinase CK2-dependent mechanism in 3T3-L1 adipocytes. J. Biol. Chem. 2013, 298, 133–142. [Google Scholar]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Savopoulos, C.G.; Ahuja, J.; Hatzitolios, A.I. Role of phytosterols in lipid lowering: Current perspectives. QJM 2011, 104, 301–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, T.A.; Sivaraman, R.M.; Sheeba Thavamani, S.; Peter Amaladhas, T.; AlSalhi, M.S.; Devanesan, S.; Kannan, M.M. Green synthesis of plasmonic nanoparticles using Sargassum ilicifolium and application in photocatalytic degradation of cationic dyes. Environ. Res. 2022, 208, 112642. [Google Scholar] [CrossRef] [PubMed]
- Cabral, C.E.; Klein, M.R.S.T. Phytosterols in the treatment of hypercholesterolemia and prevention of cardiovascular diseases. Arq. Bras. Cardiol. 2017, 109, 475–482. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Izar, M.C.; Tegani, D.M.; Kasmas, S.H.; Fonseca, F.A. Phytosterols and phytosterolemia: Gene-diet interactions. Genes Nutr. 2011, 6, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, T.A.; Klett, E.L.; Gylling, H.; Isoniemi, H.; Patel, S.B. Liver transplantation in a patient with sitosterolemia and cirrhosis. Gastroenterology 2006, 130, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, W.J.; Hofmann, A.F.; Theodor, E. Absorption of cholesterol from a micellar solution: Intestinal perfusion studies in man. J. Clin. Investig. 1967, 46, 874–890. [Google Scholar] [CrossRef] [Green Version]
- Kopylov, A.T.; Malsagova, K.A.; Stepanov, A.A.; Kaysheva, A.L. Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols. Nutrients 2021, 13, 1623. [Google Scholar] [CrossRef] [PubMed]
- Schött, H.F.; Baumgartner, S.; Husche, C.; Luister, A.; Friedrichs, S.; Miller, C.M.; McCarthy, F.O.; Plat, J.; Laufs, U.; Weingärtner, O.; et al. Oxidation of sitosterol and transport of its 7-oxygenated products from different tissues in humans and ApoE knockout mice. J. Steroid Biochem. Mol. Biol. 2017, 169, 145–151. [Google Scholar] [CrossRef]
- Lin, D.S.; Connor, W.E.; Phillipson, B.E. Sterol composition of normal human bile. Effects of feeding shellfish (marine) sterols. Gastroenterology 1984, 86, 611–617. [Google Scholar] [CrossRef]
- Muthukumar, B.; Parthipan, P.; AlSalhi, M.S.; Prabhu, N.S.; Rao, T.N.; Devanesan, S.; Maruthamuthu, M.K.; Rajasekar, A. Characterization of bacterial community in oil-contaminated soil and its biodegradation efficiency of high molecular weight (>C40) hydrocarbon. Chemosphere 2022, 289, 133168. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Rajendiran, D.; Packirisamy, S.; Gunasekaran, K. A review on role of antioxidants in diabetes. Asian J. Pharm. Clin. Res. 2018, 11, 48. [Google Scholar] [CrossRef]
- Sathyavathi, G.V.; Gupta, A.K.; Tandon, N. Medicinal Plants of India; Indian Council of Medical Research: New Delhi, India, 1987. [Google Scholar]
- Chackrewarthy, S.; Thabrew, M.I.; Weerasuriya, M.; Jayasekera, S. Evaluation of the hypoglycemic and hypolipidemic effects of an ethylacetate fraction of Artocarpus heterophyllus (jak) leaves in streptozotocin-induced diabetic rats. Phcog. Mag. 2010, 6, 186–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Maruthamuthu, M.K.; Jeong, J.; Yoo, I.K.; Kim, T.W.; Hong, S.H. Development of fenitrothion adsorbing recombinant Escherichia coli by cell surface display of pesticide-binding peptide. J. Biotechnol. 2020, 322, 90–95. [Google Scholar] [CrossRef]
- Gupta, A.; Sharma, A.K.; Dobhal, M.P.; Sharma, M.C.; Gupta, R.S. Antidiabetic and antioxidant potential of b-sitosterol in treptozotocin-induced experimental hyperglycemia. J. Diabetes 2011, 3, 29–37. [Google Scholar] [CrossRef]
- Radika, M.K.; Viswanathan, P.; Anuradha, C.V. Nitric oxide mediates the insulin sensitizing effects of β-sitosterol in high fat diet-fed rats. Nitric Oxide 2013, 32, 43–53. [Google Scholar] [CrossRef]
- Katan, M.B.; Grundy, S.M.; Jones, P.; Law, M.; Miettinen, T.; Paoletti, R. Efficacy and safety of plant stanols and sterols in the management of blood cholesterol levels. Mayo Clin. Proc. 2003, 78, 965–978. [Google Scholar]
- Berger, A.; Jones, P.J.; Abumweis, S.S. Plant sterols: Factors affecting their efficacy and safety as functional food ingredients. Lipids Health Dis. 2004, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Prommaban, A.; Utama-Ang, N.; Chaikitwattana, A.; Uthaipibull, C.; Porter, J.B.; Srichairatanakool, S. Phytosterol, Lipid and Phenolic Composition, and Biological Activities of Guava Seed Oil. Molecules 2020, 25, 2474. [Google Scholar] [CrossRef]
- Maruthamuthu, M.K.; Rudge, S.R.; Ardekani, A.M.; Ladisch, M.R.; Verma, M.S. Process Analytical Technologies and Data Analytics for the Manufacture of Monoclonal Antibodies. Trends Biotechnol. 2020, 38, 1169–1186. [Google Scholar] [CrossRef]
- Alves da Silva, S.; Sampaio, G.R.; Ferraz da Silva Torres, E.A. Phytosterols Content in Vegetable Oils of Brazil:Coconut, Safflower, Linseed and Evening Primrose. Braz. Arch. Biol. Technol. 2020, 63, e20190216. [Google Scholar] [CrossRef]
- Ajayi, T.O.; Moody, J.O.; Odumuwagun, O.J.; Olugbuyiro, J.A.O. Lipid Altering Potential of Moringa oleifera Lam Seed Extract and Isolated Constituents In Wistar Rats. Afr. J. Biomed. Res. 2020, 23, 77–85. [Google Scholar]
- Qureshi, J.A.; Memon, Z.; Ismail, K.; Saher, F.; Motiani, V.; Mushtaq, Z. Anti-Hyperglycemic and Anti-Dyslipidemic Activities of Glycyrrhiza Glabra Root Extract In Diabetic Rats. JIIMC 2020, 15, 93–108. [Google Scholar]
- De Jong, A.; Plat, J.; Bast, A.; Godschalk, R.W.; Basu, S.; Mensink, R.P. Effects of plant sterol and stanol ester consumption on lipid metabolism, antioxidant status and markers of oxidative stress, endothelial function and low-grade inflammation in patients on current statin treatment. Eur. J. Clin. Nutr. 2008, 62, 263–273. [Google Scholar] [CrossRef]
- Mayes, P.A.; Botham, K.M. Lipid transport and storage. In Harper’s Illustrated Biochemistry; Murray, R.K., Granner, D.K., Mayes, P.A., Rodwell, V.W., Eds.; Lange Medical Books/McGraw-Hill: New York, NY, USA, 2003; Volume 26. [Google Scholar]
- Al-Dosari, M.S. Hypolipidemic and antioxidant activities of avocado fruit pulp on high cholesterol fed diet in rats. Afr. J. Pharm. Pharmacol. 2011, 5, 1475–1483. [Google Scholar] [CrossRef] [Green Version]
- Oliveira Godoy Ilha, A.; Sutti Nunes, V.; Silva Afonso, M.; Regina Nakandakare, E.; da Silva Ferreira, G.; de Paula Assis Bombo, R.; Rodrigues Giorgi, R.; Marcondes Machado, R.; Carlos Rocha Quintão, E.; Lottenberg, A.M. Phytosterols Supplementation Reduces Endothelin-1 Plasma Concentration in Moderately Hypercholesterolemic Individuals Independently of Their Cholesterol-Lowering Properties. Nutrients 2020, 12, 1507. [Google Scholar] [CrossRef]
- Hatting, M.; Tavares, C.D.J.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. N. Y. Acad. Sci. 2018, 1411, 21–35. [Google Scholar] [CrossRef]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Ogbe, R.J.; Ochalefu, D.O.; Mafulul, S.G.; Olaniru, O.B. A review on dietary phytosterols: Their occurrence, metabolism and health benefits. Asian J. Plant Sci. 2015, 5, 10–21. [Google Scholar]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6, a009191. [Google Scholar]
- Meliani, N.; Dib, M.; Allali, H.; Tabti, B. Hypoglycaemic effect of Berberis vulgaris L. in normal and streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2011, 1, 468–471. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.Y.R.; Adanlawo, I.G. In vitro and in vivo antioxidant activity of saponin extracted from the root of Garcinia kola (bitter kola) on alloxan–induced diabetic rats. WJPPS 2014, 3, 08–26. [Google Scholar]
- Sethi, G.; Shanmugam, M.K.; Warrier, S.; Merarchi, M.; Arfuso, F.; Kumar, A.P.; Bishayee, A. Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients 2018, 10, 645. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Go, J.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Kim, D.S.; Son, H.J.; Lee, H.S.; Lee, C.Y.; et al. Diosgenin effectively suppresses skin inflammation induced by phthalic anhydride in IL-4/Luc/CNS-1 transgenic mice. Biosci. Biotechnol. Biochem. 2016, 80, 891–901. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.C.; Yang, J.; Yao, F.; Xie, W.; Tang, Y.Y.; Ouyang, X.P.; He, P.P.; Tan, Y.L.; Li, L.; Zhang, M.; et al. Diosgenin inhibits atherosclerosis via suppressing the MiR-19b-induced downregulation of ATP-binding cassette transporter A1. Atherosclerosis 2015, 240, 80–89. [Google Scholar] [CrossRef]
- Pari, L.; Monisha, P.; Mohamed Jalaludeen, A. Beneficial role of diosgenin on oxidative stress in aorta of streptozotocin induced diabetic rats. Eur. J. Pharmacol. 2012, 691, 143–150. [Google Scholar]
- Gan, Q.; Wang, J.; Hu, J.; Lou, G.; Xiong, H.; Peng, C.; Zheng, S.; Huang, Q. The role of diosgenin in diabetes and diabetic complications. J. Steroid Biochem. Mol. Biol. 2020, 198, 105575. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
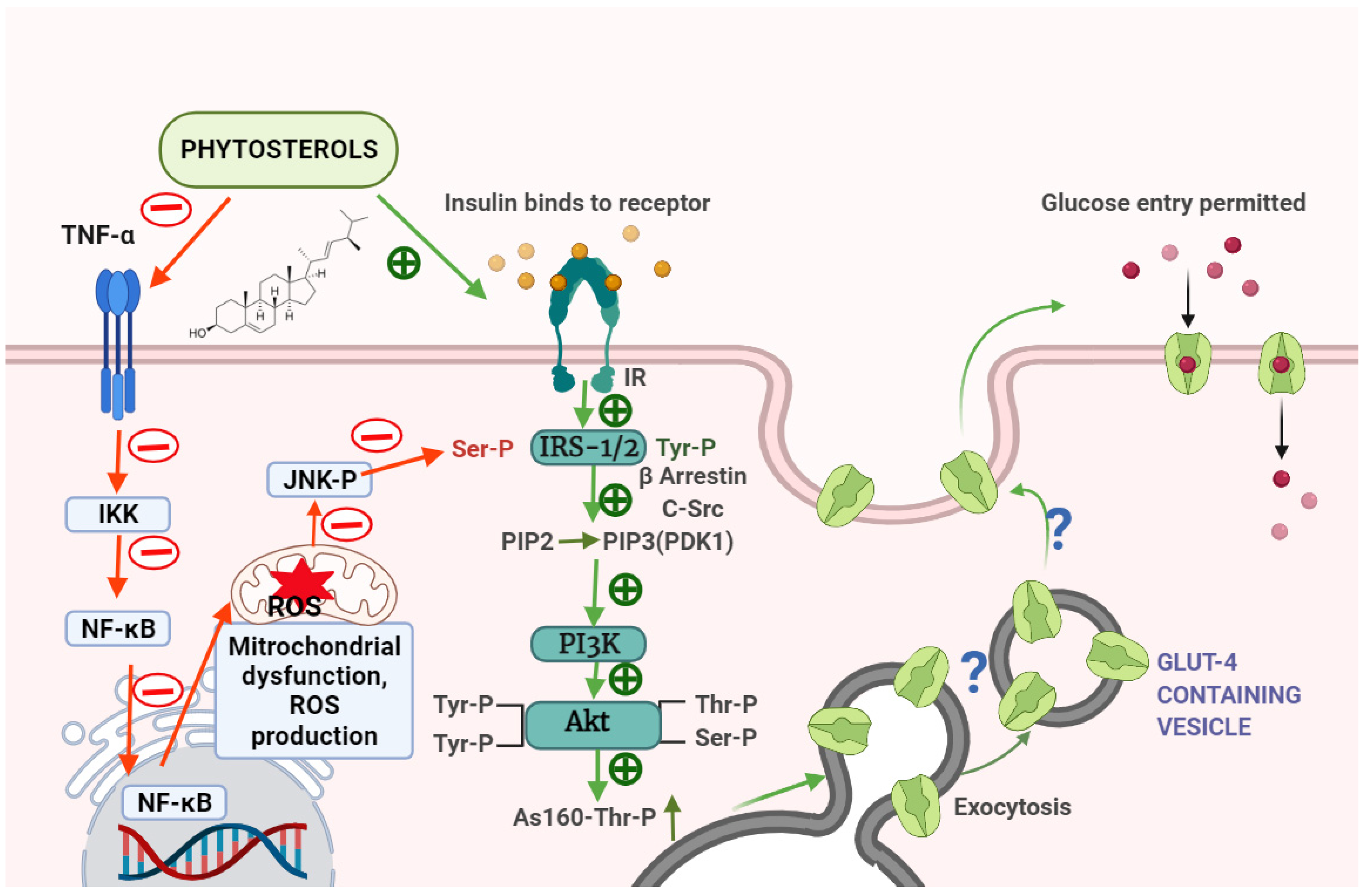{kind=link}
| S. No. | Phytosteroor Its Sources | Model | Type of Study | Biological Activity of Phytosterol | Implication | Reference |
|---|---|---|---|---|---|---|
| 1 | 0.4% stigmasterol and β-sitosterol in the diet | Mice fed a Western-style high-fat diet | In vivo | Antilipidemic activity | The study found that phytosterols were beneficial in preventing nonalcoholic fatty liver disease (NAFLD) caused by a high-fat Western diet (HFWD) (NAFLD). In this long-term (33-week) investigation, phytosterols, at a dose comparable to that advised for the alleviation of NAFLD, were predominantly related to the decreases in hepatic cholesterol, triglycerides with polyunsaturated fatty acids, and modifications of hepatic-free fatty acid. | [11] |
| 2 | Dry leaves of Eryngium foetidum L. hexane extract | Six female Swiss mice weighing 25–30 g each | In vivo | Anti-inflammatory activity | In the chromatographic fractionation of the Eryngium foetidum L. leaves isolate, a-cholesterol, brassicasterol, campesterol, stigmasterol (as the main component, 95%), clerosterol, β-sitosterol, D5-avenasterol, D524-stigmastadienol, and D7-avenasterol were all detected. The topical anti-inflammatory effects of hexane extract and stigmasterol reduced oedema in the same proportion in both animal tests (acute and chronic). Both the extract and the compound greatly reduced myeloperoxidase activity in the acute phase but not in the chronic phase, suggesting that Eryngium foetidum L. leaves could be used to treat contact dermatitis. Because stigmasterol possesses anti-inflammatory activities on the epidermis but is not a major anti-inflammatory agent, other bioactive components are likely involved in the hexane extract’s efficacy. | [12] |
| 3 | Lagerstroemia speciosa seed ethanolic extract | In silico | Breast cancer | The ADME (Adsorption, Distribution, Metabolism, Excretion) characteristics, pharmacokinetic features, drug-likeliness, and acute toxicity of the discovered phytosterols compounds are predicted by ethanolic extracts Lagerstroemia speciosa seeds GC–MS analysis and in silico analysis. GC–MS analysis identified 29 chemicals from the extract, four phytosterol derivatives, cholesterol margarate, 7-dehydrodiosgenin, stigmastan-3,5-diene, and γ-sitosterol, which were shown to be non-toxic, non-carcinogenic, and non-mutagenic. The extent of molecular interaction with breast cancer targets is also revealed by molecular docking studies. This research reveals that phytosterols derived from Lagerstroemia speciosa’s ethanolic seed extract could be a potential anti-breast cancer alternative. | [13] | |
| 4 | β-sitosterol | Male albino wistar rats | In vivo | Anti-inflammatory activity | In high fat diet- and sucrose-induced type-2 diabetic rats, β-sitosterol treatment normalises raised serum levels of proinflammatory cytokines, such as leptin, resistin, tumour necrosis factor-(TNF-α), and interleukin-6 (IL-6) and increases anti-inflammatory adipocytokines, such as adiponectin. In diabetic rats’ adipocytes, β-sitosterol lowers sterol regulatory element binding protein-1c (SREBP-1c) and increases Peroxisome Proliferator–Activated Receptor-γ (PPAR-γ) gene expression. In β-sitosterol-treated groups, c-Jun-N-terminal kinase-1 (JNK1), inhibitor of nuclear factor kappa-B kinase subunit beta (IKK), and nuclear factor kappa B (NF-kB) gene and protein expression were likewise considerably reduced. This study reveals that SIT prevents obesity-induced insulin resistance by reducing inflammatory events in the adipose tissue via inhibiting the IKK/NF-kB and c-Jun-N-terminal kinase (JNK) signalling pathways. | [14] |
| 5 | β-sitosterol | Male albino wistar rats | In vivo | Antidiabetic activity | Treatment with β-sitosterol restored the altered levels of blood glucose, serum insulin, testosterone, lipid profile, oxidative stress indicators, antioxidant enzymes, insulin receptor (IR), and glucose transporter 4 (GLUT4) proteins in a high-fat diet and sucrose-induced diabetic rats. This research shows that β-sitosterol improves glycemic control in high-fat and sucrose-induced type-2 diabetic rats through activating IR and GLUT4. In addition, the results of in silico analysis match those of in vivo testing. | [15] |
| 6 | β-sitosterol | Male albino wistar rats | In vivo | Antidiabetic activity | When compared to high-fat diet and sucrose induced type-2 diabetic rats, β-sitosterol increased the mRNA expression of IR and post-receptor insulin signalling molecules such as IRS-1, β-arrestin-2, Akt, AS160, and GLUT4, as well as the levels of IRS-1, p-IRS1-1Tyr632, Akt, p-AktSer473/Thr308, AS160, and p-AS160Thr642. In this study, in silico analysis revealed that β-sitosterol has a higher binding affinity for β-arrestin-2, c-Src, and IRS-1, as well as Akt proteins, and has been shown to reduce insulin resistance, as evidenced by in vivo data. According to the findings, β-sitosterol reduces the potential effects of a high-fat diet on adipose tissue. | [16] |
| 7 | β-sitosterol | Male albino wistar rats | In vivo | Antidiabetic activity | In this study, high-fat diet and sucrose-induced diabetic rats showed reduced glucose and insulin tolerances, as well as insulin signalling molecules (IR and GLUT4) and glycogen levels. Serum insulin, lipid profile, LPO, H2O2, and OH* levels were shown to be higher in diabetic rats. The β-sitosterol therapy brought blood glucose, insulin, lipid profile, oxidative stress indicators, IR, and GLUT4 protein levels back to normal. This research suggests that IR and GLUT4 activation by β-sitosterol improves glycemic control in the gastrocnemius muscle of HFD-fed and sucrose-induced type 2 diabetic rats. | [17] |
| 8 | Phormidium autumnale (P. autumnale) cyanobacteria | In vitro | Neuroprotective properties | Compressed fluid technologies were used to produce phytosterol-rich extracts from P. autumnale cyanobacteria in order to investigate their potential neuroprotective capabilities in this work. The optimised compressed fluid extract demonstrated stronger in vitro neuroprotective efficacy than the non-enriched extract in bioactivity tests, such as acetylcholinesterase inhibition, lipoxygenase inhibition, and antioxidant capacity. The specificity of sterol interaction with acetylcholinesterase active sites was demonstrated using in-silico molecular docking investigations. These findings support future investigation of P. autumnale as a source of bioactive phytosterols, highlighting the utility of compressed fluid methods in obtaining phytosterol-rich extracts. | [18] | |
| 9 | Soybean oil contains stigmasterol | KK-Ay mice | In vivo | Anti-diabetic activity | In L6 cells, stigmasterol greatly boosted GLUT4 translocation and glucose absorption. Furthermore, this in vivo research revealed that after 4 weeks of stigmasterol therapy, the hyperglycemic phenotype of diabetic KK-Ay mice was dramatically relieved, with significantly lower fasting glucose, indicating a considerable hypoglycemic impact against T2DM. In this investigation, stigmasterol appeared to offer potential benefits in the treatment of type 2 diabetes, with the likely method of targeting the GLUT4 glucose transporter, including increased GLUT4 translocation and expression. | [19] |
| 10 | Lophenol (Lo) and cycloartanol (Cy), minor phytosterols of Aloe vera gel | Zucker diabetic fatty (ZDF) rats | In vivo | Anti-diabetic activity | After 35 days of treatment, minor phytosterols from Aloe vera gel, lophenol (Lo) and cycloartanol (Cy), decreased hyperglycemia and random blood glucose levels in Zucker diabetic fatty (ZDF) rats, an obese animal model of type II diabetes. Apart from total cholesterol, continued treatment of Lo and Cy lowered blood free fatty acid (FFA) and triglyceride (TG) levels (T-Cho). Furthermore, with Lo (27.7%) and Cy (26.3%) treatment, the weights of total abdominal fat tissues in ZDF rats were considerably lower than in the control group. These findings imply that phytosterols obtained from Aloe vera may help to minimise visceral fat buildup and ameliorate hyperlipidemia and hyperglycemia. | [20] |
| 11 | Nuts | Human study | In vivo | Anti-diabetic activity | Randomised controlled studies of type 2 diabetes patients corroborated the positive benefits of nuts on blood lipids, which were also found in non-diabetic people, but no change in A1C or other glycated proteins was identified in the trials. However, acute feeding studies have shown that nuts can lower postprandial glycemia when consumed with a carbohydrate (bread). Additionally, nut intake was related to lower postprandial oxidative stress. Nuts have a favourable nutritional profile, being abundant in monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA), as well as being good sources of vegetable protein. Nuts may so increase the diet’s overall nutritious quality. | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, M.; Jayaraman, S.; Eladl, M.A.; El-Sherbiny, M.; Abdelrahman, M.A.E.; Veeraraghavan, V.P.; Vengadassalapathy, S.; Umapathy, V.R.; Jaffer Hussain, S.F.; Krishnamoorthy, K.; et al. A Comprehensive Review on Therapeutic Perspectives of Phytosterols in Insulin Resistance: A Mechanistic Approach. Molecules 2022, 27, 1595. https://doi.org/10.3390/molecules27051595
Prasad M, Jayaraman S, Eladl MA, El-Sherbiny M, Abdelrahman MAE, Veeraraghavan VP, Vengadassalapathy S, Umapathy VR, Jaffer Hussain SF, Krishnamoorthy K, et al. A Comprehensive Review on Therapeutic Perspectives of Phytosterols in Insulin Resistance: A Mechanistic Approach. Molecules. 2022; 27(5):1595. https://doi.org/10.3390/molecules27051595
Chicago/Turabian StylePrasad, Monisha, Selvaraj Jayaraman, Mohamed Ahmed Eladl, Mohamed El-Sherbiny, Mosaab Abdella Ebrahim Abdelrahman, Vishnu Priya Veeraraghavan, Srinivasan Vengadassalapathy, Vidhya Rekha Umapathy, Shazia Fathima Jaffer Hussain, Kalaiselvi Krishnamoorthy, and et al. 2022. "A Comprehensive Review on Therapeutic Perspectives of Phytosterols in Insulin Resistance: A Mechanistic Approach" Molecules 27, no. 5: 1595. https://doi.org/10.3390/molecules27051595