
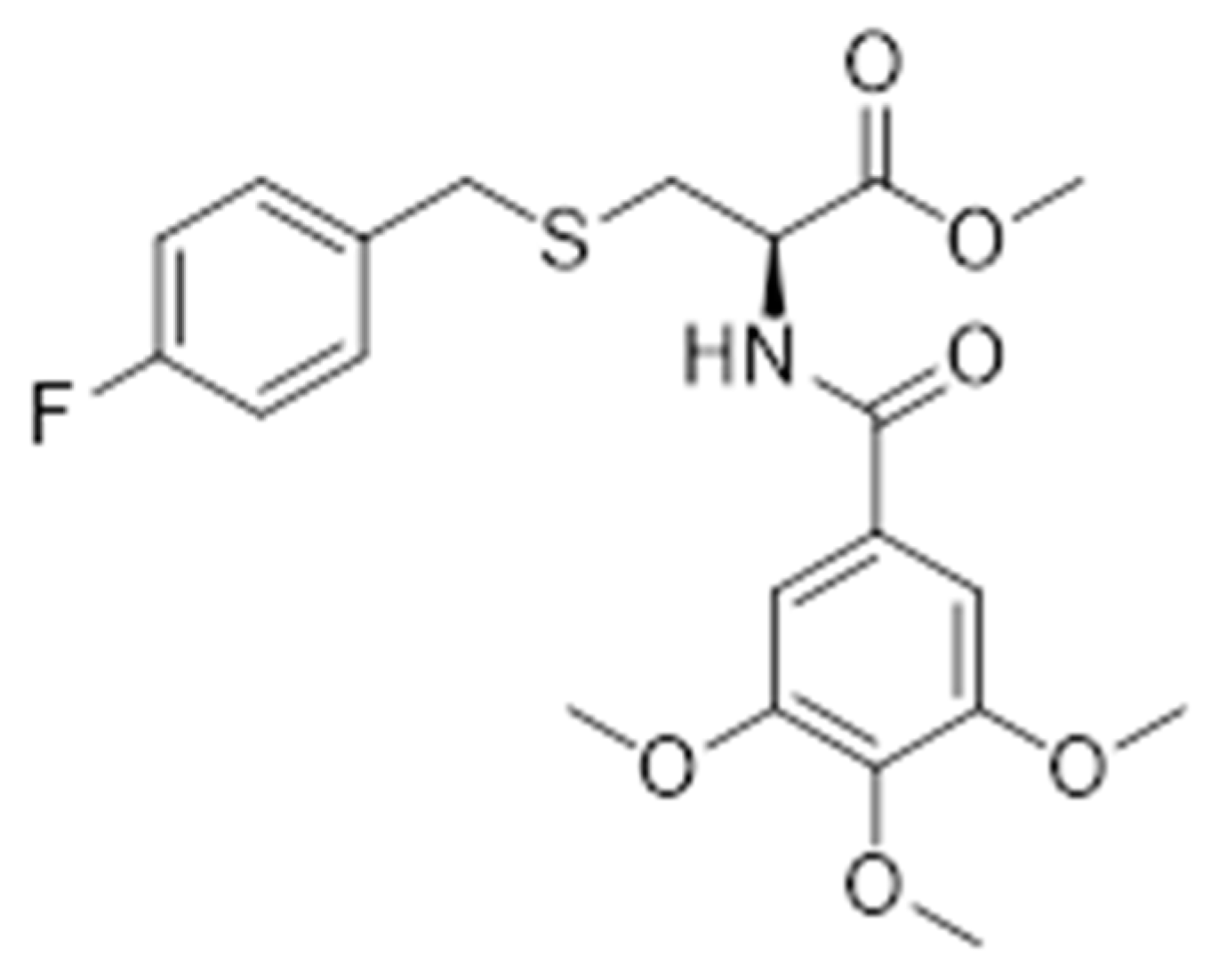
The Protective Effects of Hydrogen Sulfide New Donor Methyl S-(4-Fluorobenzyl)-N-(3,4,5-Trimethoxybenzoyl)-l-Cysteinate on the Ischemic Stroke

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
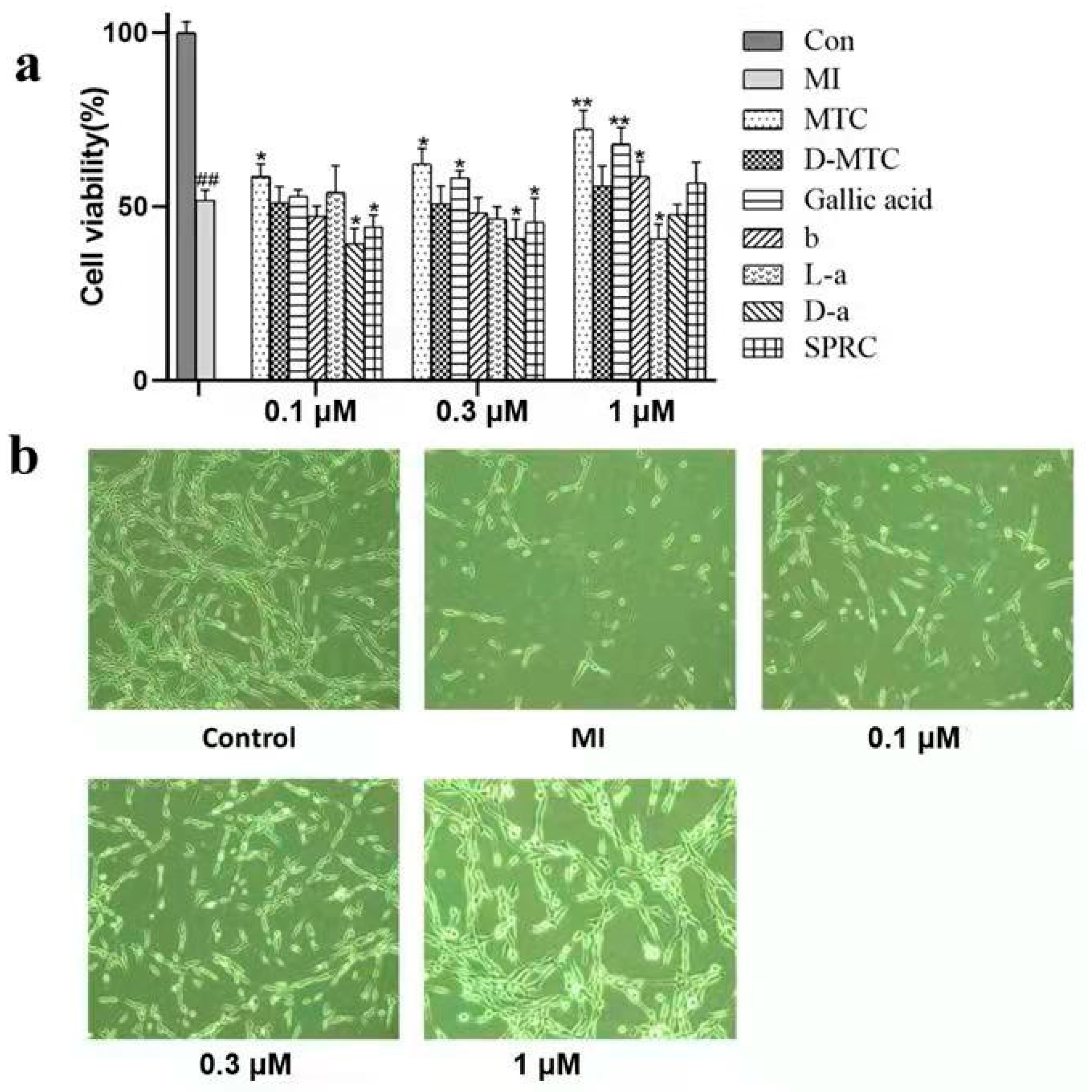
2.1. Effect of MTC and Multitarget Compounds on Cell Viability after Ischemia-Reperfusion Induced PC12 Cell Injury
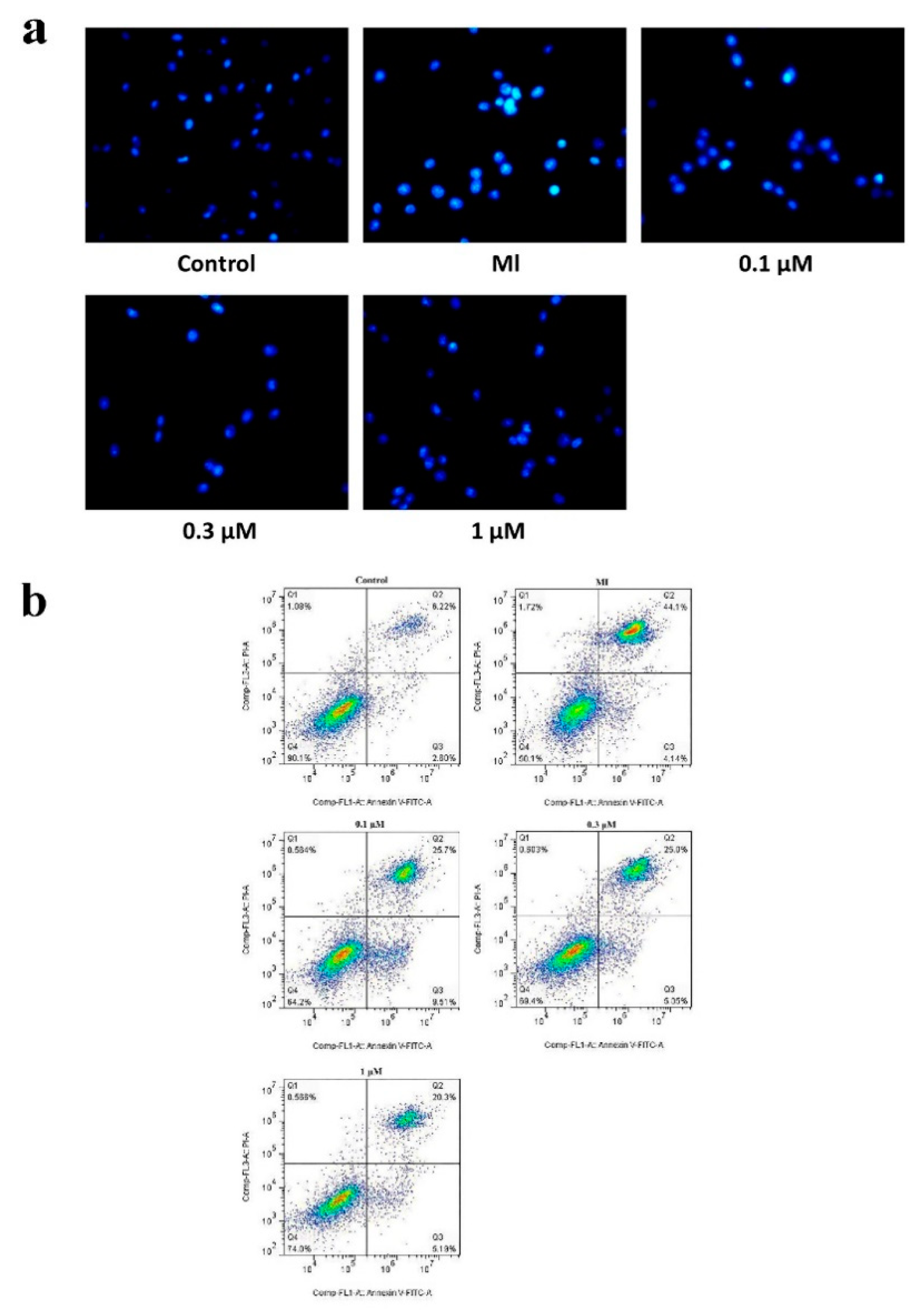
2.2. MTC Post-Treatment Reduces Apoptosis Induced by Ischemia-Reperfusion in PC12 Cells
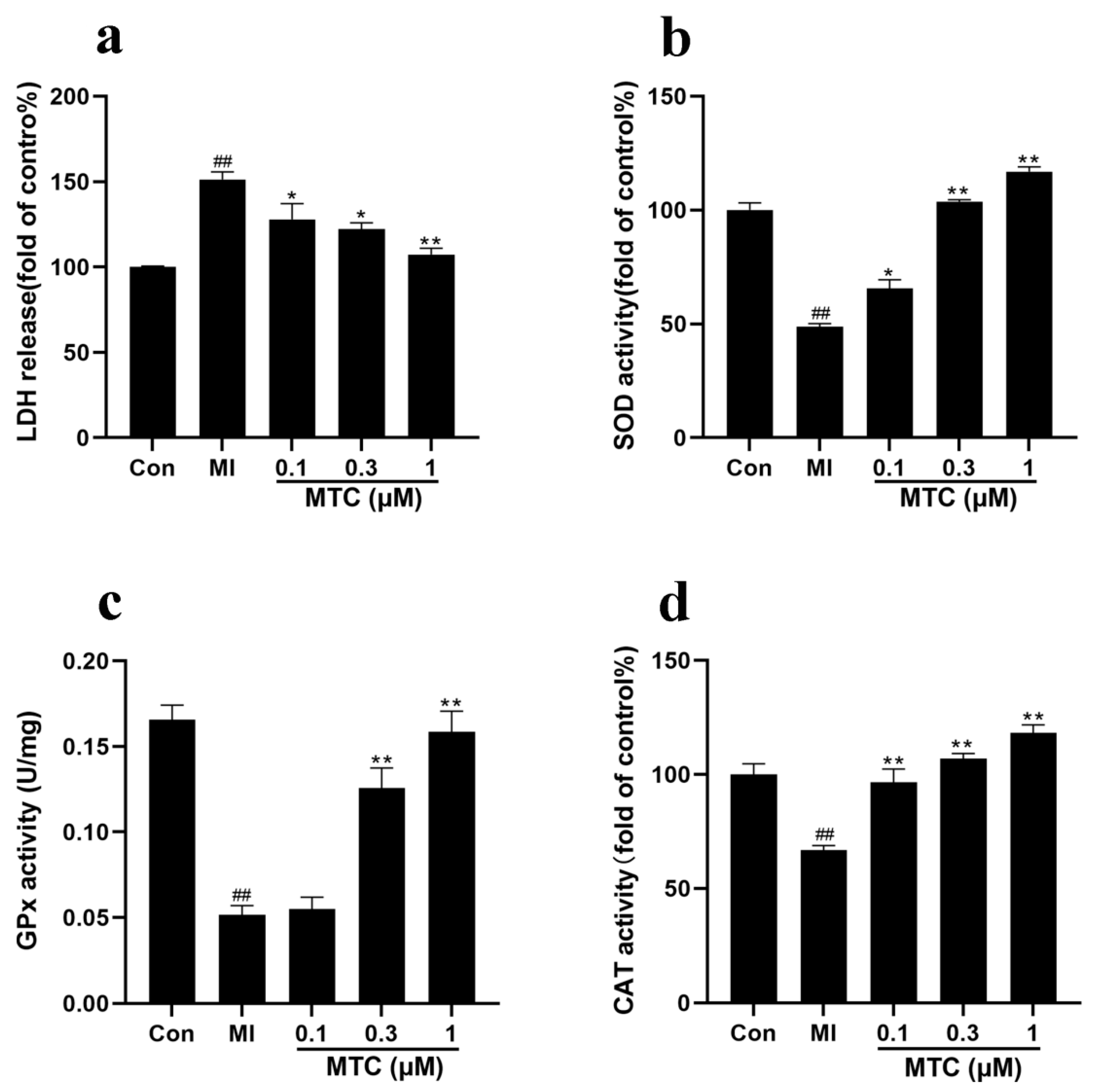
2.3. Effects of MTC on LDH, SOD, GPx and CAT after Ischemia-Reperfusion-Induced PC12 Cell Injury
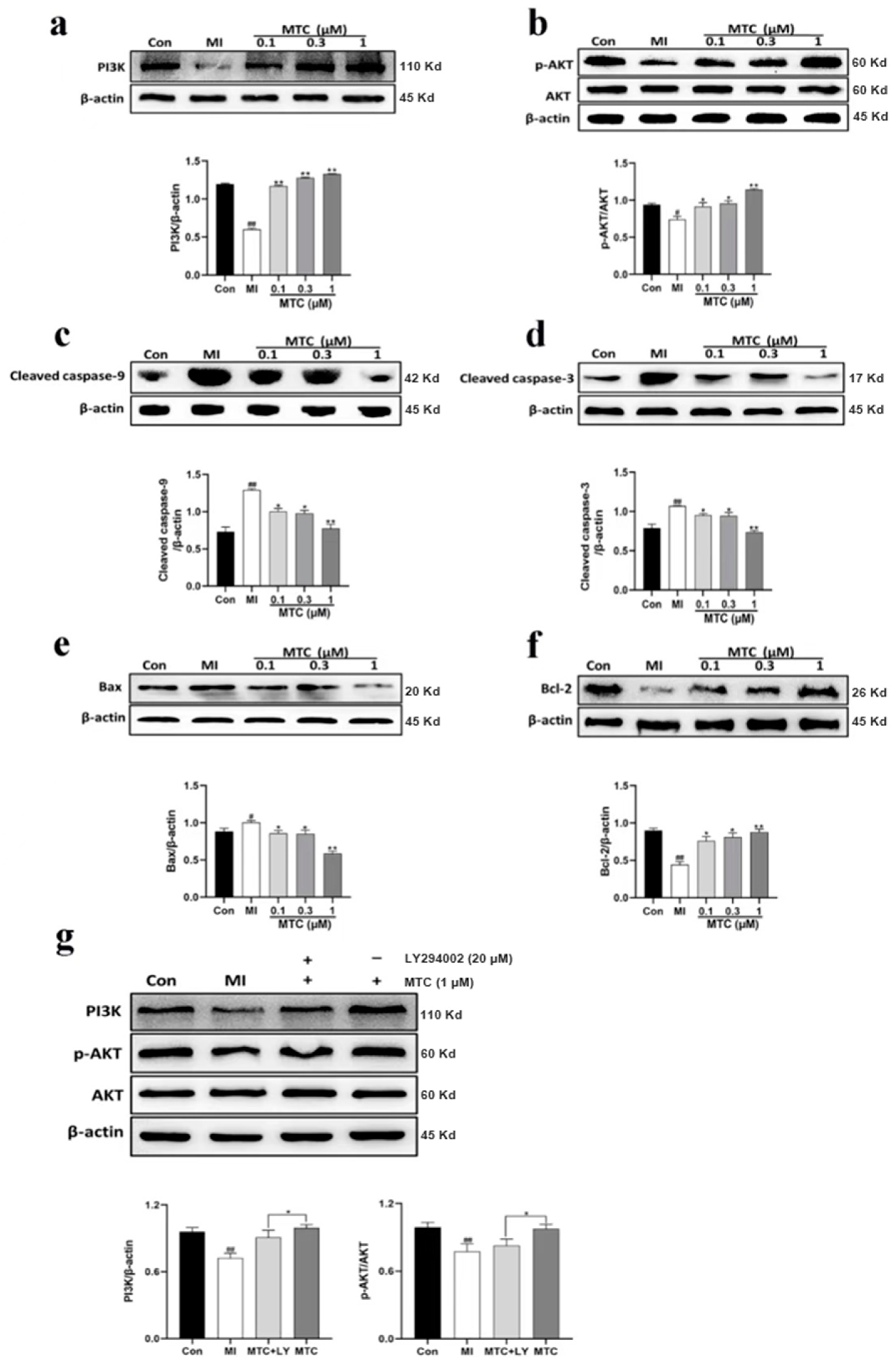
2.4. Effects of MTC on the Levels of PI3K, p-AKT, Cleaved Caspase-9, Cleaved Caspase-3, Bax and Bcl-2 after PC12 Cell Injury Induced by Ischemia-Reperfusion
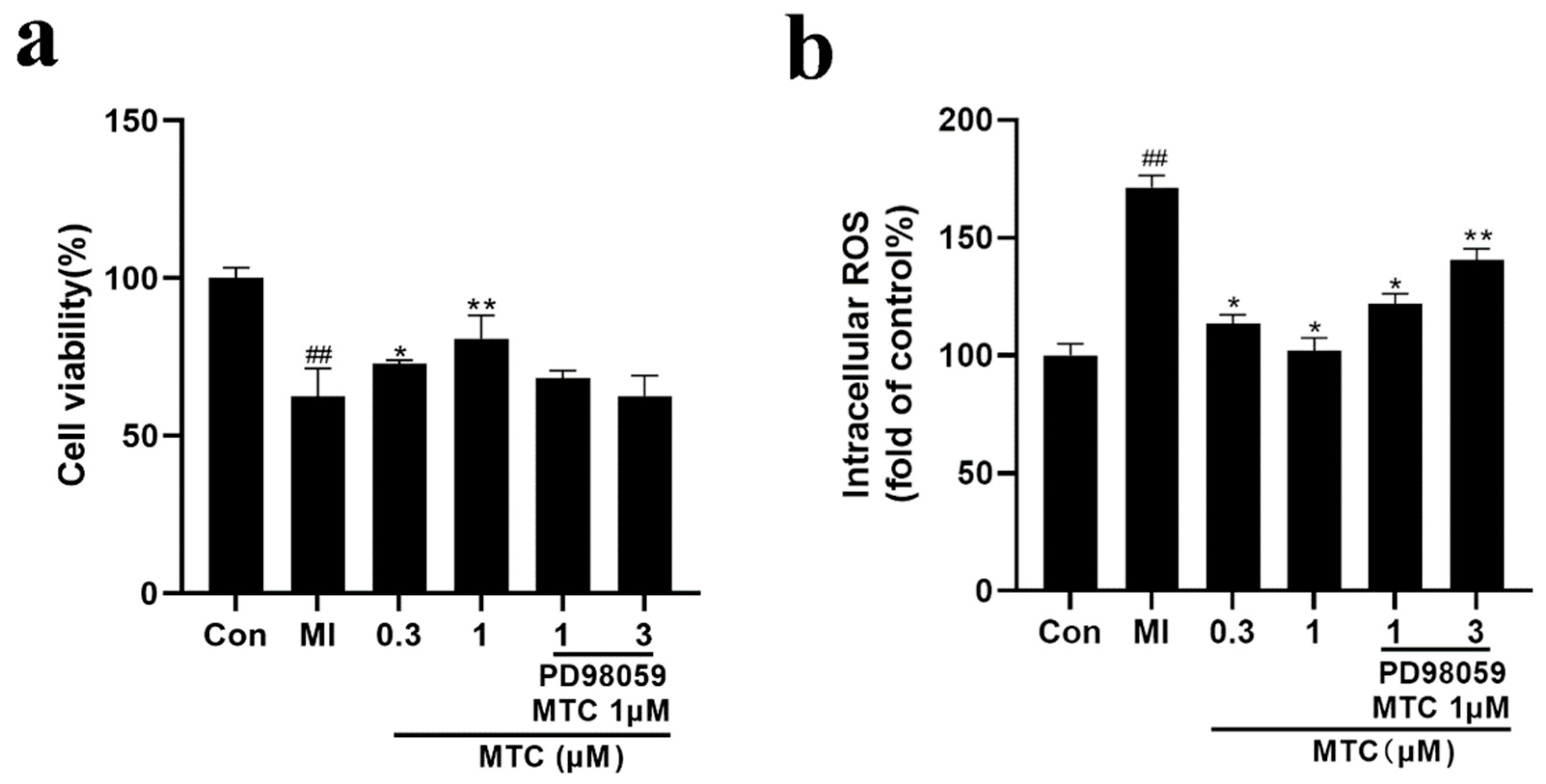
2.5. Effect of PD98059 Pretreatment on the Activity and ROS of MTC Treated PC12 Cells
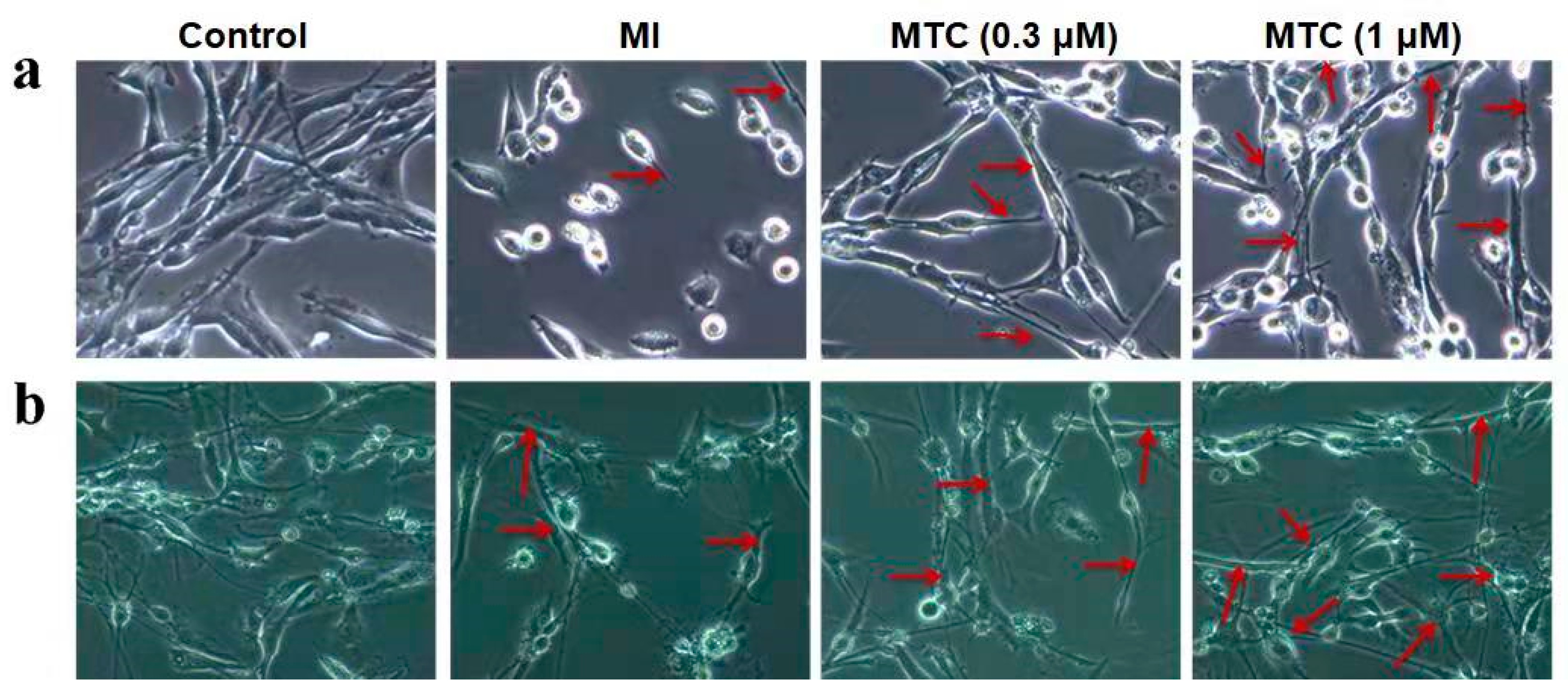
2.6. Effect of MTC on Axon Growth of PC12 Cells and Neurons Induced by Ischemia-Reperfusion Injury
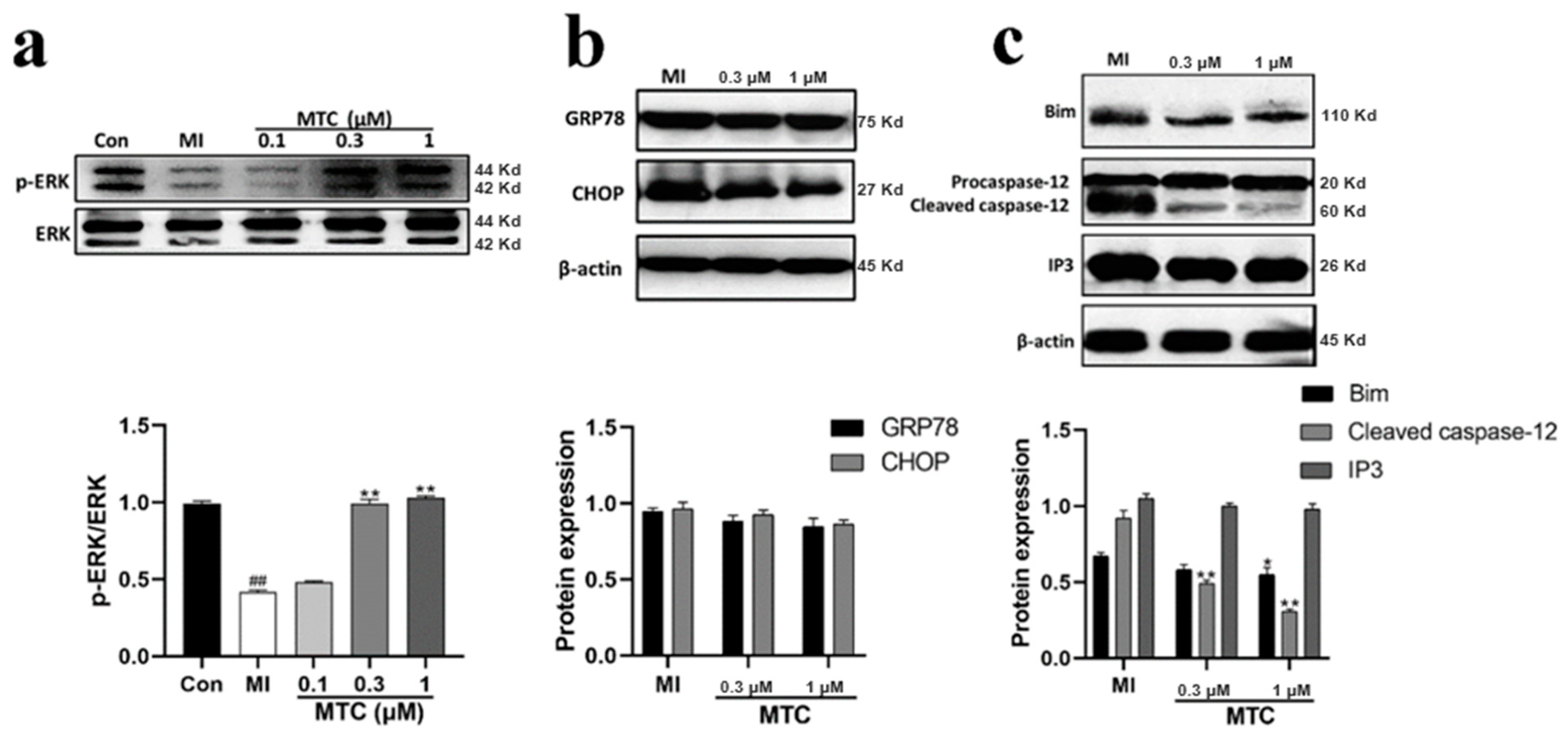
2.7. Effect of MTC on Expression of Endoplasmic Reticulum Stress Related Protein in PC12 Cells Induced by Ischemia-Reperfusion Injury
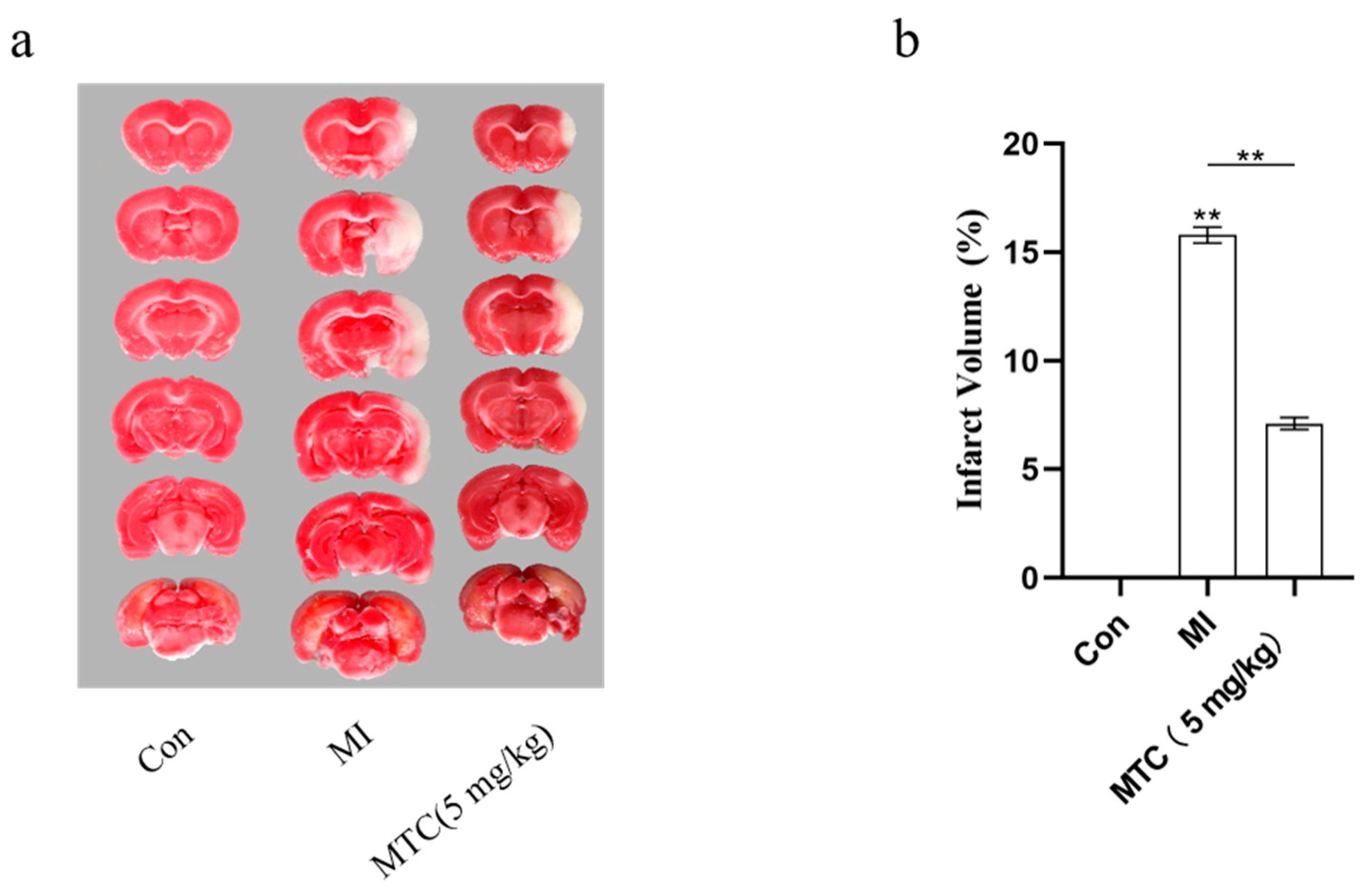
2.8. MTC on Cerebral Injury Induced by Ischemia-Reperfusion in Rats
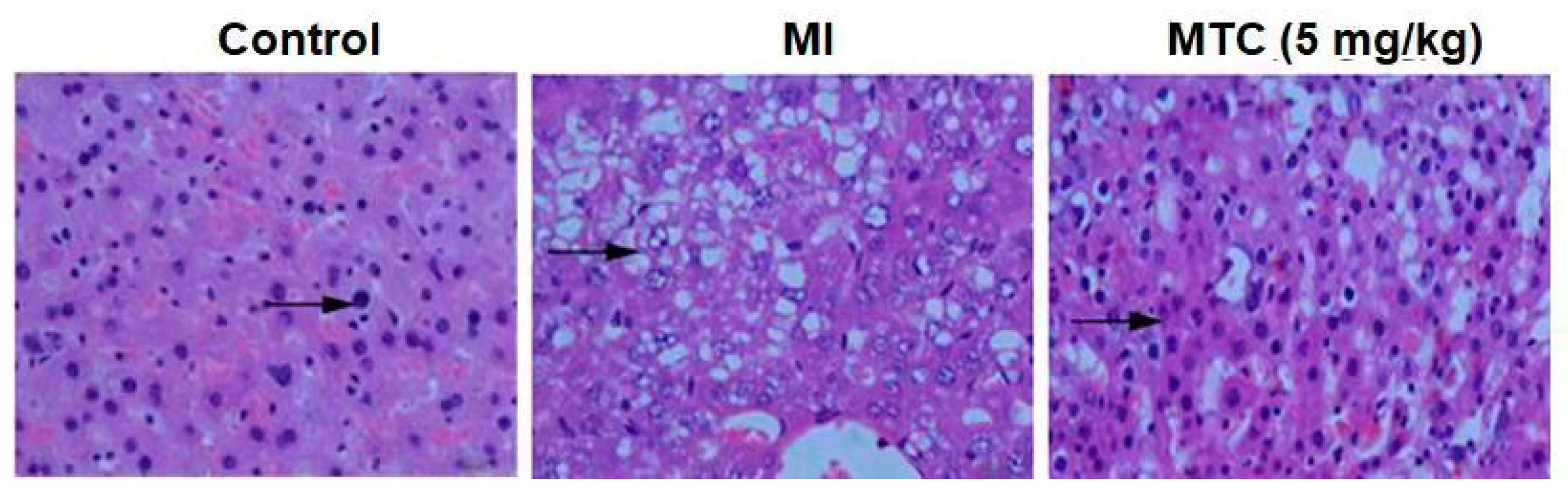
2.9. Pathological Observation of MTC on Hippocampus Injury Induced by Ischemia-Reperfusion in Rats
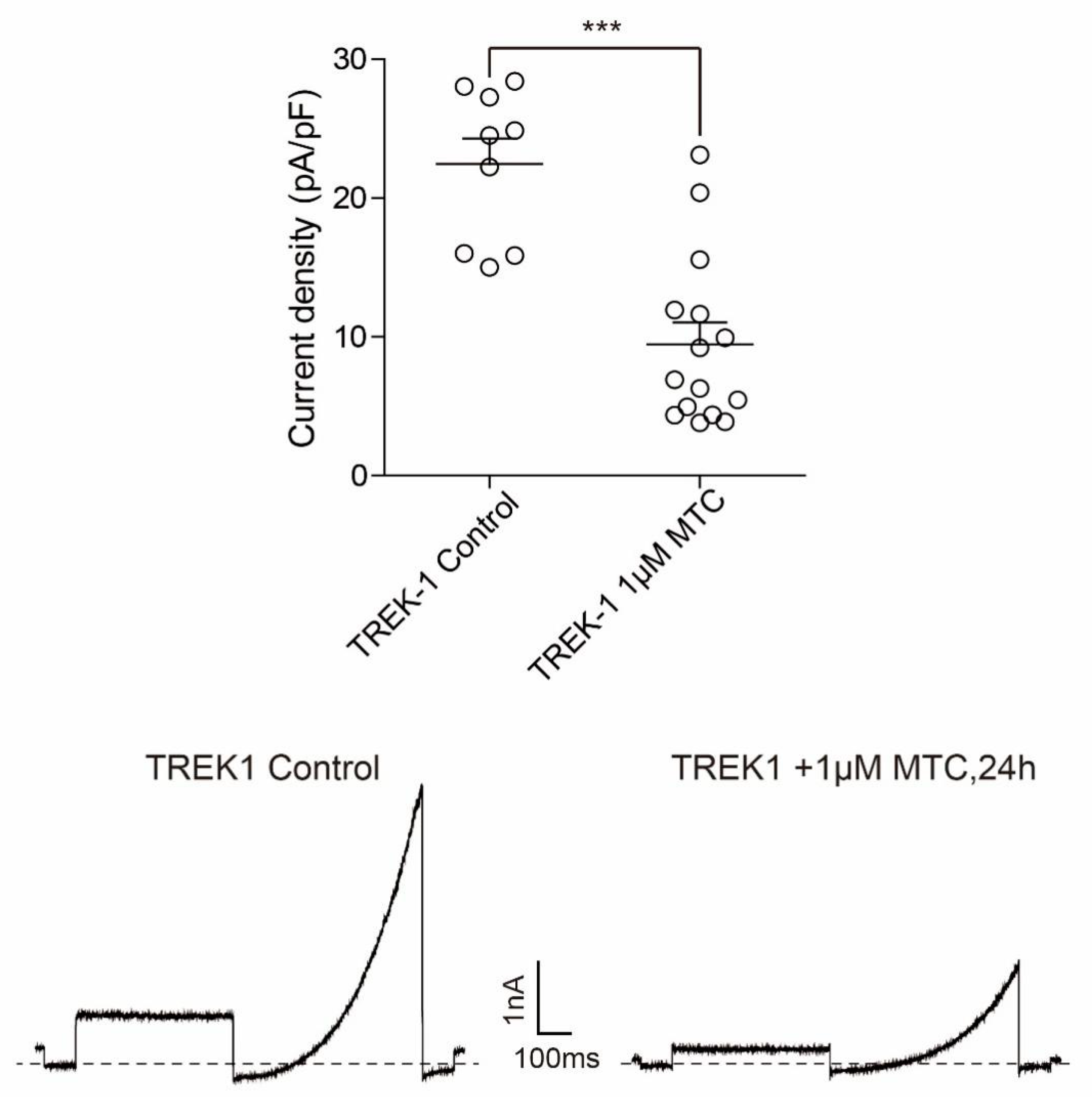
2.10. TREK1 Current Density and TREK1 Channel Protein Level Is Decreased by Prolonged MTC Stimulation in CHO Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cells and Materials
4.3. Chemical Synthesis
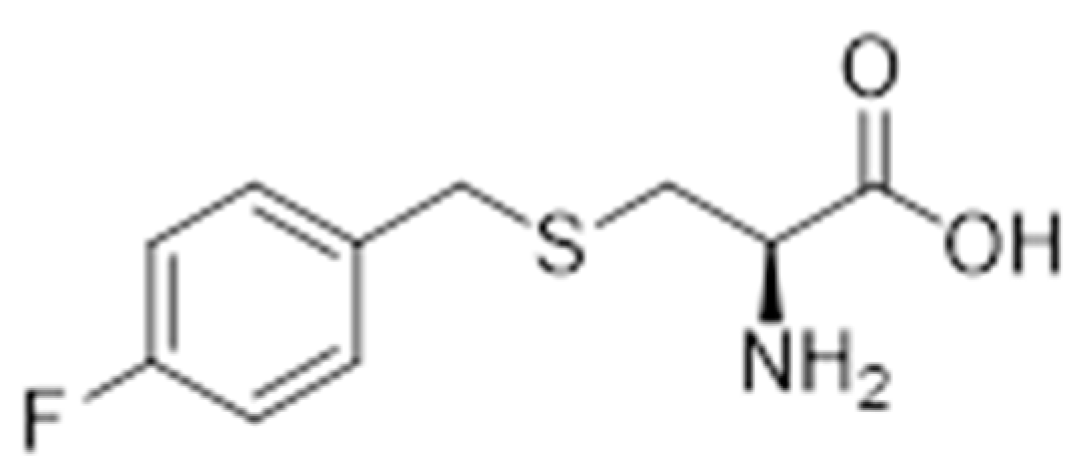
4.3.1. Synthesis of S-(4-Fluorobenzyl)-l-Cysteine
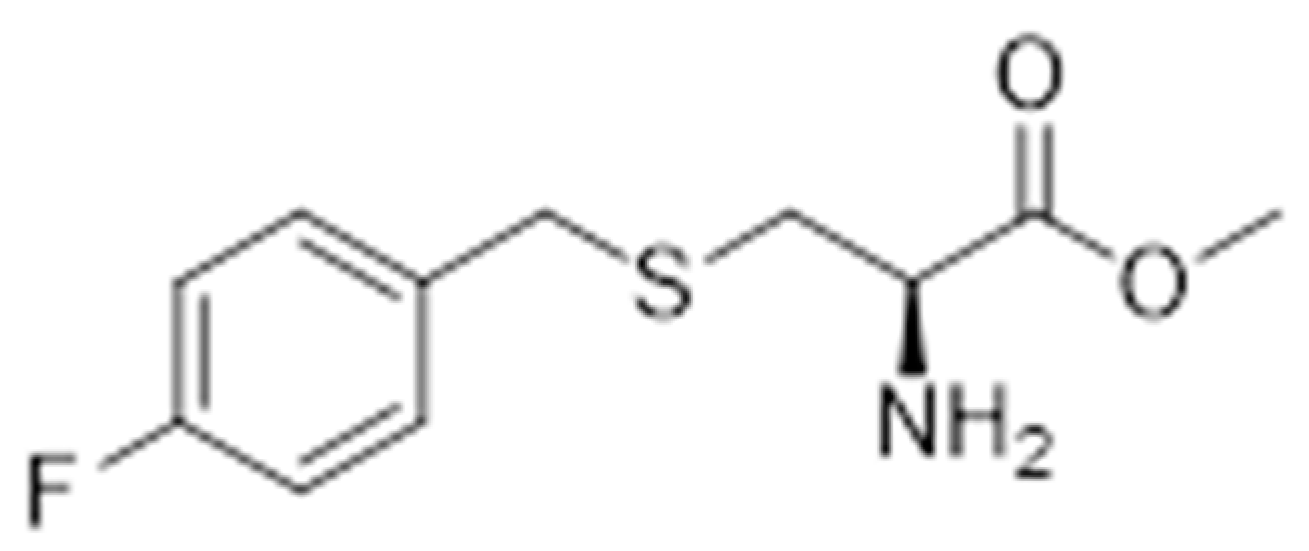
4.3.2. Synthesis of Methyl S-(4-Fluorobenzyl)-l-Cysteinate
4.3.3. Synthesis of Methyl S-(4-Fluorobenzyl)-N-(3,4,5-Trimethoxybenzoyl)-l-Cysteinate
- 1H-NMR (400 MHz, Chloroform-d) δ 7.28–7.23 (m, 2H), 7.04 (s, 2H), 6.97 (t, J = 7.8 Hz, 2H), 6.89 (d, J = 6.5 Hz, 1H), 5.02–4.95 (m, 1H), 3.91 (dd, J = 3.8, 2.1 Hz, 9H), 3.79 (d, J = 2.3 Hz, 3H), 3.72 (s, 2H), 3.06 (ddd, J = 14.0, 5.2, 2.0 Hz, 1H), 2.95 (ddd, J = 14.1, 5.9, 2.1 Hz, 1H).
- 13C-NMR (101 MHz, CDCl3) δ 171.4, 166.6, 163.1, 160.7, 153.2, 141.4, 133.2, 133.2, 130.4, 130.3, 128.9, 115.5, 115.3, 104.6, 60.8, 56.3, 52.7, 52.1, 35.8, 33.3.
4.4. Cell Culture and Transfection
4.5. Establishment and Grouping of Ischemic Models
4.6. Cell Viability Assay
4.7. Hoechst 33258 Nuclear Staining
4.8. FITC-ANNEXinV/PI Dual Staining
4.9. LDH, SOD, CAT and GPx Analysis
4.10. Western Blot Analysis
4.11. Primary Neuron Culture
4.12. Focal Cerebral Ischemia-Reperfusion Model in Rats
4.13. TTC Staining
4.14. HE Staining
4.15. Whole-Cell Patch Clamp Recording
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Huang, H.-L.; Wang, N.; Zhou, H.; Yu, C.-Y. Study on influence of transient ischemic attack on subsequent cerebral infarction. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5164–5167. [Google Scholar] [PubMed]
- Fernandes, P.M.; Whiteley, W.; Hart, S.R.; Salman, R.A.-S. Strokes: Mimics and chameleons. Pr. Neurol. 2013, 13, 21–28. [Google Scholar] [CrossRef]
- Du, S.; Deng, Y.; Yuan, H.; Sun, Y. Safflower Yellow B Protects Brain against Cerebral Ischemia Reperfusion Injury through AMPK/NF-kB Pathway. Evid. Based Complement. Altern. Med. 2019, 2019, 7219740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-F.; Zhang, L.; Shi, L.-L.; Zhao, Z.-H.; Xu, H.; Liang, F.; Li, H.-B.; Zhao, Y.; Xu, X.; Yang, K.; et al. Parthenolide attenuates cerebral ischemia/reperfusion injury via Akt/GSK-3β pathway in PC12 cells. Biomed. Pharmacother. 2017, 89, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Tripathi, M.; Srivastava, M.V.P.; Vivekanandhan, S.; Prasad, K. Relationship between polymorphisms in beta -2 adrenergic receptor gene and ischemic stroke in North Indian Population: A hospital based case control study. BMC Res. Notes 2014, 7, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yang, J.; Zhang, C.; Jiang, X.; Zhou, H.; Liu, M. Calcium antagonists for acute ischemic stroke. Cochrane Database Syst. Rev. 2012, 16, CD001928. [Google Scholar] [CrossRef]
- Amarenco, P.B.J.; Callahan, A., 3rd; GoldsTein, L.B.; Hennerici, M.; Rudolph, A.E.; Sillesen, H.; Simunovic, L.; Szarek, M.; Welch, K.M.; Zivin, J.A. The Stroke Prevention by Aggressive Reduction in Cholesterol Levels (SPARCL) Investigators High-Dose Atorvastatin after Stroke or Transient Ischemic Attack. N. Engl. J. Med. 2006, 355, 549–559. [Google Scholar] [CrossRef]
- Chrysant, S.G. The Pathophysiologic Role of the Brain Renin-Angiotensin System in Stroke Protection: Clinical Implications. J. Clin. Hypertens. 2007, 9, 454–459. [Google Scholar] [CrossRef]
- Gao, Y.; Yin, H.; Zhang, Y.; Dong, Y.; Yang, F.; Wu, X.; Liu, H. Dexmedetomidine protects hippocampal neurons against hypoxia/reoxygenation-induced apoptosis through activation HIF-1α/p53 signaling. Life Sci. 2019, 232, 116611. [Google Scholar] [CrossRef]
- Jiang, W.-W.; Huang, B.-S.; Han, Y.; Deng, L.-H.; Wu, L.-X. Sodium hydrosulfide attenuates cerebral ischemia/reperfusion injury by suppressing overactivated autophagy in rats. FEBS Open Bio 2017, 7, 1686–1695. [Google Scholar] [CrossRef]
- Li, X.; Zhang, K.-Y.; Zhang, P.; Chen, L.-X.; Wang, L.; Xie, M.; Wang, C.-Y.; Tang, X.-Q. Hydrogen Sulfide Inhibits Formaldehyde-Induced Endoplasmic Reticulum Stress in PC12 Cells by Upregulation of SIRT-1. PLoS ONE 2014, 9, e89856. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocer, B.; Guven, H.; Comoglu, S.S. Homocysteine Levels in Parkinson’s Disease: Is Entacapone Effective? BioMed Res. Int. 2016, 2016, 7563705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Yang, X.; Zhao, S.; Wei, C.; Yin, Y.; Liu, T.; Jiang, S.; Xie, J.; Wan, X.; Mao, M.; et al. Hydrogen sulfide prevents OGD/R-induced apoptosis via improving mitochondrial dysfunction and suppressing an ROS-mediated caspase-3 pathway in cortical neurons. Neurochem. Int. 2013, 63, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Sparatore, A.; Del Soldato, P.; Mcgeer, E.; McGeer, P.L. Hydrogen sulfide-releasing NSAIDs attenuate neuroinflammation induced by microglial and astrocytic activation. Glia 2010, 58, 103–113. [Google Scholar] [CrossRef]
- Hu, L.-F.; Lu, M.; Wu, Z.-Y.; Wong, P.T.-H.; Bian, J.-S. Hydrogen Sulfide Inhibits Rotenone-Induced Apoptosis via Preservation of Mitochondrial Function. Mol. Pharmacol. 2009, 75, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Niu, Y.-Y.; Jiang, W.-Z.; Tang, H.-L.; Zhang, C.; Xiao-Qing, T.; Tang, X.-Q. Neuroprotective effects of hydrogen sulfide and the underlying signaling pathways. Rev. Neurosci. 2015, 26, 129–142. [Google Scholar] [CrossRef]
- Kimura, H. Signaling Molecules: Hydrogen Sulfide and Polysulfide. Antioxid. Redox Signal. 2015, 22, 362–376. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Liu, Y.; Tang, P.; Liu, P.; Hou, C.; Zhang, X.; Chen, L.; Zhang, L.; Gu, C. Hydrogen sulfide prevents OGD/R-induced apoptosis by suppressing the phosphorylation of p38 and secretion of IL-6 in PC12 cells. NeuroReport 2016, 27, 230–234. [Google Scholar] [CrossRef]
- Liu, C.; Guo, W.; Shi, X.; Kaium, M.; Gu, X.; Zhu, Y.Z. Leonurine-cysteine analog conjugates as a new class of multifunctional anti-myocardial ischemia agent. Eur. J. Med. Chem. 2011, 46, 3996–4009. [Google Scholar] [CrossRef]
- Ma, K.; Liu, Y.; Zhu, Q.; Liu, C.-H.; Duan, J.-L.; Tan, B.K.-H.; Zhu, Y.Z. H2S Donor, S-Propargyl-Cysteine, Increases CSE in SGC-7901 and Cancer-Induced Mice: Evidence for a Novel Anti-Cancer Effect of Endogenous H2S? PLoS ONE 2011, 6, e20525. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gu, X.; Zhu, Y.Z. Synthesis and biological evaluation of novel leonurine–SPRC conjugate as cardioprotective agents. Bioorganic Med. Chem. Lett. 2010, 20, 6942–6946. [Google Scholar] [CrossRef] [PubMed]
- Tsitsiou, E.; Sibley, C.P.; D’Souza, S.W.; Catanescu, O.; Jacobsen, D.W.; Glazier, J.D. Homocysteine transport by systems L, A and y+L across the microvillous plasma membrane of human placenta. J. Physiol. 2009, 587, 4001–4013. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate Sulfurtransferase Produces Hydrogen Sulfide and Bound Sulfane Sulfur in the Brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-D.; Wang, D.-Y.; Li, H.-M.; Guo, J.-C.; Duan, S.-F.; Ji, X.-Y. Hydrogen Sulfide as a Novel Regulatory Factor in Liver Health and Disease. Oxidative Med. Cell. Longev. 2019, 2019, 3831713. [Google Scholar] [CrossRef]
- Ka, H.; Yi, B.; Kim, M.; Lee, J. Evaluation of Antioxidant or Prooxidant Properties of Selected Amino Acids Using In Vitro Assays and in Oil-in-Water Emulsions Under Riboflavin Sensitization. J. Food Sci. 2016, 81, C1118–C1123. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Osama, N. Effect of Gallic acid in potentiating chemotherapeutic effect of Paclitaxel in HeLa cervical cancer cells. Cancer Cell Int. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing Gallic Acid for Its Broad Spectrum Applications. Mini Rev. Med. Chem. 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Akdemir, F.N.E.; Yildirim, S.; Kandemir, F.M.; Tanyeli, A.; Küçükler, S.; Dortbudak, M.B. Protective effects of gallic acid on doxorubicin-induced cardiotoxicity; an experimantal study. Arch. Physiol. Biochem. 2021, 127, 258–265. [Google Scholar] [CrossRef]
- Daglia, M.; Di Lorenzo, A.; Nabavi, S.F.; Talas, Z.S. Polyphenols: Well Beyond The Antioxidant Capacity: Gallic Acid and Related Compounds as Neuroprotective Agents: You are What You Eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef]
- Cheng, Y.; Tse, H.-F.; Li, X.; Han, Y.; Rong, J. Gallic acid- l -leucine (GAL) conjugate enhances macrophage phagocytosis via inducing leukotriene B4 12-hydroxydehydrogenase (LTB4DH) expression. Mol. Immunol. 2016, 74, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Li, X.; Tse, H.-F.; Rong, J. Gallic Acid-L-Leucine Conjugate Protects Mice against LPS-Induced Inflammation and Sepsis via Correcting Proinflammatory Lipid Mediator Profiles and Oxidative Stress. Oxidative Med. Cell. Longev. 2018, 2018, 1081287. [Google Scholar] [CrossRef]
- Li, L.; Chang, K.-C.; Zhou, Y.; Shieh, B.; Ponder, J.; Abraham, A.D.; Ali, H.; Snow, A.; Petrash, J.M.; LaBarbera, D.V. Design of an Amide N-Glycoside Derivative of β-Glucogallin: A Stable, Potent, and Specific Inhibitor of Aldose Reductase. J. Med. Chem. 2014, 57, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesi, G.; Chen, Q.; Sestito, S.; Digiacomo, M.; Yang, X.; Wang, S.; Pi, R.; Rapposelli, S. Nature-based molecules combined with rivastigmine: A symbiotic approach for the synthesis of new agents against Alzheimer’s disease. Eur. J. Med. Chem. 2017, 141, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Wang, W.; Du, J.; Chen, K.; Cheng, K. Structure-activity relationship study and biological evaluation of SAC-Garlic acid conjugates as novel anti-inflammatory agents. Eur. J. Med. Chem. 2019, 179, 233–245. [Google Scholar] [CrossRef]
- Kweon, J.H.; Kim, S.; Lee, S.B. The cellular basis of dendrite pathology in neurodegenerative diseases. BMB Rep. 2017, 50, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.M.; Baig, H.S.; Hammarlund, M. Mitochondria Localize to Injured Axons to Support Regeneration. Neuron 2016, 92, 1308–1323. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.; Peers, C.; Kemp, P.J. Polymodal regulation of hTREK1 by pH, arachidonic acid, and hypoxia: Physiological impact in acidosis and alkalosis. Am. J. Physiol. Physiol. 2004, 286, C272–C282. [Google Scholar] [CrossRef]
- Göb, E.; Bittner, S.; Bobak, N.; Kraft, P.; Göbel, K.; Langhauser, F.; Homola, G.A.; Brede, M.; Budde, T.; Meuth, S.G.; et al. The two-pore domain potassium channel KCNK5 deteriorates outcome in ischemic neurodegeneration. Pflügers Arch. Eur. J. Physiol. 2014, 467, 973–987. [Google Scholar] [CrossRef]
- White, B.C.; I Grossman, L.; O’Neil, B.J.; DeGracia, D.J.; Neumar, R.W.; A Rafols, J.; Krause, G.S. Global Brain Ischemia and Reperfusion. Ann. Emerg. Med. 1996, 27, 588–594. [Google Scholar] [CrossRef]
- Shimoji, M.; Hara, H.; Kamiya, T.; Okuda, K.; Adachi, T. Hydrogen sulfide ameliorates zinc-induced cell death in neuroblastoma SH-SY5Y cells. Free Radic. Res. 2017, 51, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Yu, Q.; Zhao, P.; Yang, Q.; Wang, B.; Yang, Y.; Kuai, J.; Ding, Q. Preconditioning with hydrogen sulfide ameliorates cerebral ischemia/reperfusion injury in a mouse model of transient middle cerebral artery occlusion. Chem. Interact. 2019, 310, 108738. [Google Scholar] [CrossRef]
- Ren, C.; Du, A.; Li, D.; Sui, J.; Mayhan, W.G.; Zhao, H. Dynamic change of hydrogen sulfide during global cerebral ischemia–reperfusion and its effect in rats. Brain Res. 2010, 1345, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Warenycia, M.W.; Goodwin, L.R.; Benishin, C.G.; Reiffenstein, R.J.; Francom, D.M.; Taylor, J.D.; Dieken, F.P. Acute hydrogen sulfide poisoning: Demonstration of selective uptake of sulfide by the brainstem by measurement of brain sulfide levels. Biochem. Pharmacol. 1989, 38, 973–981. [Google Scholar] [CrossRef]
- Bullock, R.; Zauner, A.; Woodward, J.J.; Myseros, J.; Choi, S.C.; Ward, J.D.; Marmarou, A.; Young, H.F. Factors affecting excitatory amino acid release following severe human head injury. J. Neurosurg. 1998, 89, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Fan, G.-C.; Ren, X.; Waggoner, J.; Gregory, K.N.; Chen, G.; Jones, W.K.; Kranias, E.G. Overexpression of histidine-rich Ca-binding protein protects against ischemia/reperfusion-induced cardiac injury. Cardiovasc. Res. 2007, 75, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.L.; Bayraktutan, U. Oxidative Stress and Its Role in the Pathogenesis of Ischaemic Stroke. Int. J. Stroke 2009, 4, 461–470. [Google Scholar] [CrossRef]
- Jassem, W.; Fuggle, S.V.; Rela, M.; Koo, D.D.; Heaton, N.D. The role of mitochondria in ischemia/reperfusion injury. Transplant. 2002, 73, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.-I.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, L.; Yu, X. Protective effect of hydrogen sulfide on rats with myocardial ischemia/reperfusion injury and its mechanism. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi = Chin. J. Cell. Mol. Immunol. 2015, 31, 316–320. [Google Scholar]
- Wen, Z.; Hou, W.; Wu, W.; Zhao, Y.; Dong, X.; Bai, X.; Peng, L.; Song, L. 6′-O-Galloylpaeoniflorin Attenuates Cerebral Ischemia Reperfusion-Induced Neuroinflammation and Oxidative Stress via PI3K/Akt/Nrf2 Activation. Oxidative Med. Cell. Longev. 2018, 2018, 8678267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Hu, G.; Zhu, S.; Li, J.; Liu, G. Propofol pretreatment attenuates lipopolysaccharide-induced acute lung injury in rats by activating the phosphoinositide-3-kinase/Akt pathway. Braz. J. Med. Biol. Res. 2014, 47, 1062–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Zhao, J.; Zhou, X.; Yu, L.; Liu, Z.; Chang, Y. Propofol can suppress renal ischemia-reperfusion injury through the activation of PI3K/AKT/mTOR signal pathway. Gene 2019, 708, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-F.; Jiang, H.; Chen, J.; Guo, X.; Li, Y.; Hu, Q.; Yang, S. Nobiletin ameliorates myocardial ischemia and reperfusion injury by attenuating endoplasmic reticulum stress-associated apoptosis through regulation of the PI3K/AKT signal pathway. Int. Immunopharmacol. 2019, 73, 98–107. [Google Scholar] [CrossRef]
- Lee, S.; Youn, K.; Kim, D.H.; Ahn, M.-R.; Yoon, E.; Kim, O.-Y.; Jun, M. Anti-Neuroinflammatory Property of Phlorotannins from Ecklonia cava on Aβ25-35-Induced Damage in PC12 Cells. Mar. Drugs 2018, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; An, R.; Luo, Y.; Li, M.; Xu, L.; Dong, Z. Tamibarotene Improves Hippocampus Injury Induced by Focal Cerebral Ischemia-Reperfusion via Modulating PI3K/Akt Pathway in Rats. J. Stroke Cerebrovasc. Dis. 2019, 28, 1832–1840. [Google Scholar] [CrossRef]
- Mohamed, S.K.; Ahmed, A.A.; Elmorsy, E.M.; Nofal, S. ERK activation by zeranol has neuroprotective effect in cerebral ischemia reperfusion. Life Sci. 2019, 227, 137–144. [Google Scholar] [CrossRef]
- Li, Z.; Yang, L.; Zhen, J.; Zhao, Y.; Lu, Z. Nobiletin protects PC12 cells from ERS-induced apoptosis in OGD/R injury via activation of the PI3K/AKT pathway. Exp. Ther. Med. 2018, 16, 1470–1476. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-C.; Kim, Y.H.; Lee, T.-K.; Kim, I.H.; Cho, J.H.; Cho, G.-S.; Shin, B.-N.; Park, J.H.; Ahn, J.H.; Shin, M.C.; et al. Effects of ischemic preconditioning on PDGF-BB expression in the gerbil hippocampal CA1 region following transient cerebral ischemia. Mol. Med. Rep. 2017, 16, 1627–1634. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Qi, D.; Sun, Y.; Huang, X.; Zhang, F.; Wu, J.; Fu, Y.; Ma, K.; Du, Y.; Dong, H.; et al. H2S attenuates cognitive deficits through Akt1/JNK3 signaling pathway in ischemic stroke. Behav. Brain Res. 2014, 269, 6–14. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Q.; Chen, X.; Jing, L.; Wang, W.; Yu, Z.; Zhang, G.; Xie, M. Linolenic Acid Provides Multi-cellular Protective Effects After Photothrombotic Cerebral Ischemia in Rats. Neurochem. Res. 2014, 39, 1797–1808. [Google Scholar] [CrossRef] [PubMed]
- Wakeno-Takahashi, M.; Otani, H.; Nakao, S.; Imamura, H.; Shingu, K. Isoflurane induces second window of preconditioning through upregulation of inducible nitric oxide synthase in rat heart. Am. J. Physiol. Circ. Physiol. 2005, 289, H2585–H2591. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, J.; Wang, Y.; Tan, R. Anti-Neuroinflammatory Effect of Alantolactone through the Suppression of the NF-κB and MAPK Signaling Pathways. Cells 2019, 8, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Du, J.; Zhang, Z.; Shi, W.; Hu, B.; Hu, J.; Xue, Y.; Li, H.; Ji, W.; Zhuang, J.; et al. The Protective Effects of Hydrogen Sulfide New Donor Methyl S-(4-Fluorobenzyl)-N-(3,4,5-Trimethoxybenzoyl)-l-Cysteinate on the Ischemic Stroke. Molecules 2022, 27, 1554. https://doi.org/10.3390/molecules27051554
Fan J, Du J, Zhang Z, Shi W, Hu B, Hu J, Xue Y, Li H, Ji W, Zhuang J, et al. The Protective Effects of Hydrogen Sulfide New Donor Methyl S-(4-Fluorobenzyl)-N-(3,4,5-Trimethoxybenzoyl)-l-Cysteinate on the Ischemic Stroke. Molecules. 2022; 27(5):1554. https://doi.org/10.3390/molecules27051554
Chicago/Turabian StyleFan, Jing, Junxi Du, Zhongwei Zhang, Wenjing Shi, Binyan Hu, Jiaqin Hu, Yan Xue, Haipeng Li, Wenjin Ji, Jian Zhuang, and et al. 2022. "The Protective Effects of Hydrogen Sulfide New Donor Methyl S-(4-Fluorobenzyl)-N-(3,4,5-Trimethoxybenzoyl)-l-Cysteinate on the Ischemic Stroke" Molecules 27, no. 5: 1554. https://doi.org/10.3390/molecules27051554