
Heartwood of Dalbergia cochinchinensis: 4,7,2′-Trihydroxy-4′-methoxyisoflavanol and 6,4′-Dihydroxy-7-methoxyflavane Reduce Cytokine and Chemokine Expression In Vitro


 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Extraction of Compounds
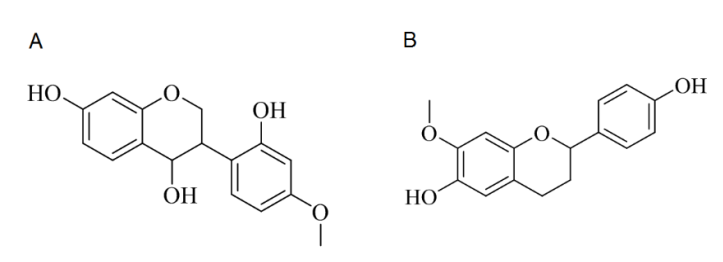
2.2. Isolation of 4,7,2′-Trihydroxy-4′-methoxyisoflavanol (472T4MIF)
2.3. Isolation of 6,4′-Dihydroxy-7-methoxyflavane (64D7MF)
2.4. Murine RAW 264.7 Cells, Primary Macrophages, and Human Gingival Fibroblasts
2.5. Cell Stimulation
2.6. RT-PCR and Immunoassay
2.7. Western Blot
2.8. Statistical Analysis
3. Results
3.1. Isolation of 472T4MIF and 64D7MF
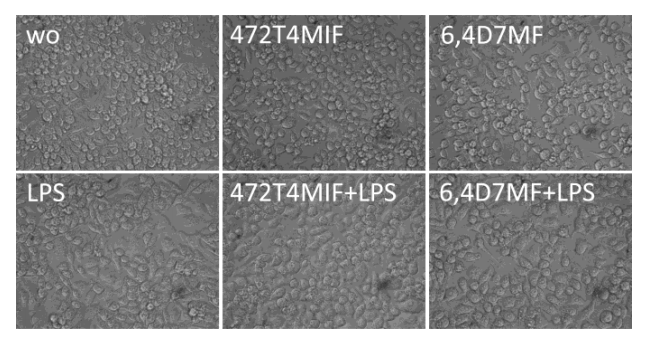
3.2. 472T4MIF and 64D7MF Do Not Affect Viability of RAW 264.7 Macrophages
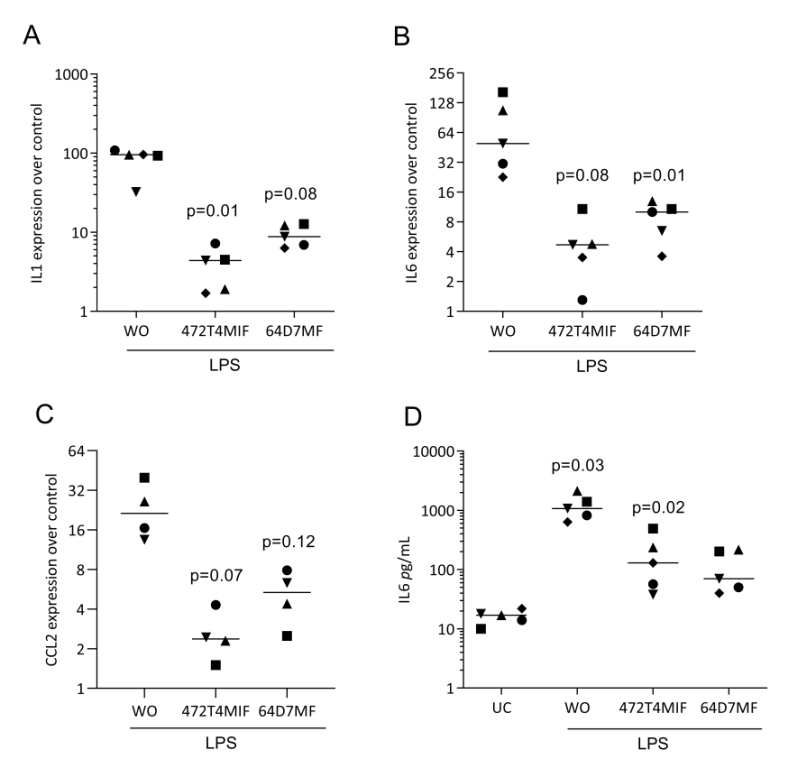
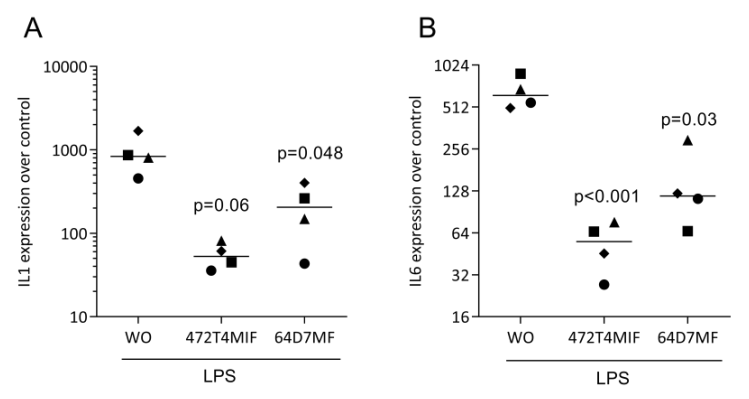
3.3. 472T4MIF and 64D7MF Have an Activity in Murine Macrophages
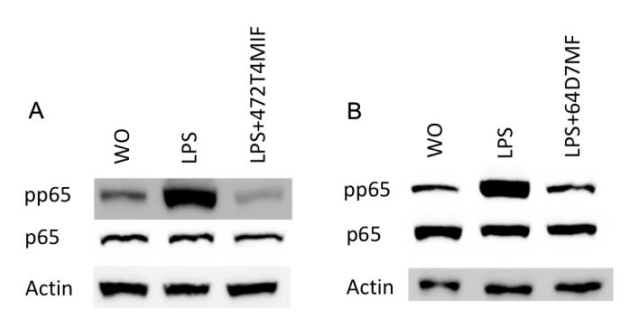
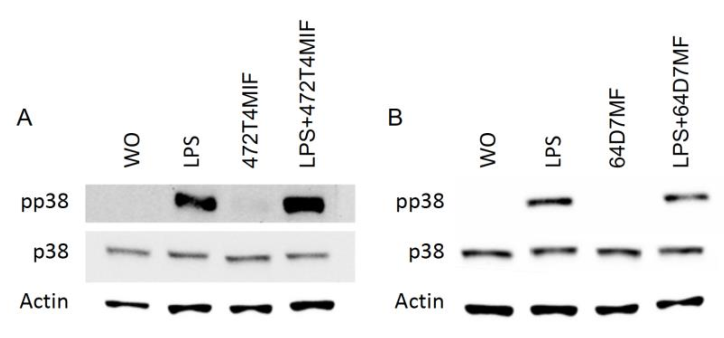
3.4. 472T4MIF and 64D7MF Diminished the p65 and p38 Phosphorylation in Macrophages
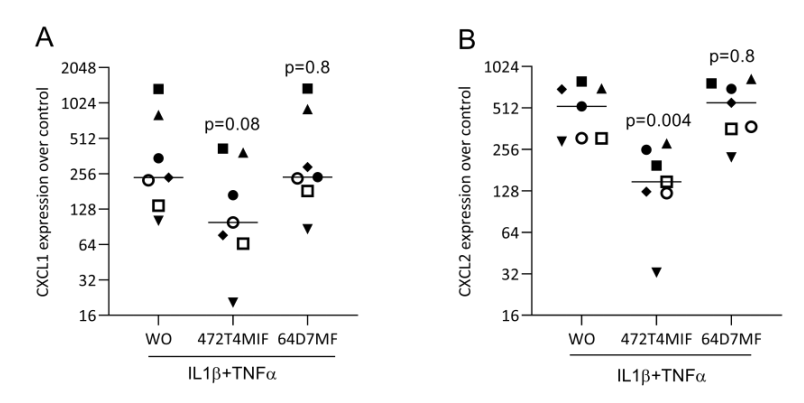
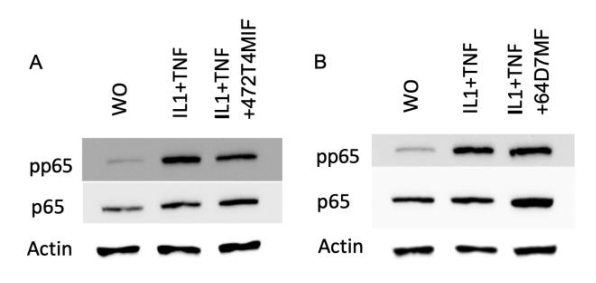
3.5. 472T4MIF and 64D7MF Do Not Reduce the Response of Fibroblasts to IL1β and TNFa
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Hung, T.H.; Gooda, R.; Rizzuto, G.; So, T.; Thammavong, B.; Tran, H.T.; Jalonen, R.; Boshier, D.H.; MacKay, J.J. Physiological responses of rosewoods Dalbergia cochinchinensis and D. oliveri under drought and heat stresses. Ecol. Evol. 2020, 10, 10872–10885. [Google Scholar] [CrossRef] [PubMed]
- Hirata, M.; Mitsuyuki, C.; Moritsuka, E.; Chhang, P.; Tagane, S.; Toyama, H.; Sokh, H.; Rueangruea, S.; Suddee, S.; Suyama, Y.; et al. Evaluating the genetic diversity in two tropical leguminous trees, Dalbergia cochinchinensis and D. nigrescens, in lowland forests in Cambodia and Thailand using MIG-seq. Genes Genet. Syst. 2021, 96, 41–53. [Google Scholar] [CrossRef]
- Hung, T.H.; So, T.; Sreng, S.; Thammavong, B.; Boounithiphonh, C.; Boshier, D.H.; MacKay, J.J. Reference transcriptomes and comparative analyses of six species in the threatened rosewood genus Dalbergia. Sci. Rep. 2020, 10, 17749. [Google Scholar] [CrossRef] [PubMed]
- Hartvig, I.; So, T.; Changtragoon, S.; Tran, H.T.; Bouamanivong, S.; Theilade, I.; Kjaer, E.D.; Nielsen, L.R. Population genetic structure of the endemic rosewoods Dalbergia cochinchinensis and D. oliveri at a regional scale reflects the Indochinese landscape and life-history traits. Ecol. Evol. 2018, 8, 530–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tor-Ngern, P.; Leksungnoen, N. Investigating carbon dioxide absorption by urban trees in a new park of Bangkok, Thailand. BMC Ecol. 2020, 20, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Z.; Chen, X.; Zhao, Z.; Xiao, X.; Guo, P.; Song, H.; Yang, X.; Huang, M. Analysis of volatile components in Dalbergia cochinchinensis Pierre by a comprehensive two-dimensional gas chromatography with mass spectrometry method using a solid-state modulator. J. Sep. Sci. 2018, 41, 4315–4322. [Google Scholar] [CrossRef]
- Liu, R.H.; Wen, X.C.; Li, Y.Y.; Zhang, P.Z.; Shao, F.; Huang, H.L.; Tang, F.R. Chemical Constituents from Dalbergia cochinchinensis. Zhong Yao Cai=Zhongyaocai=J. Chin. Med. Mater. 2015, 38, 1868–1871. [Google Scholar]
- Shirota, O.; Pathak, V.; Sekita, S.; Satake, M.; Nagashima, Y.; Hirayama, Y.; Hakamata, Y.; Hayashi, T. Phenolic constituents from Dalbergia cochinchinensis. J. Nat. Prod. 2003, 66, 1128–1131. [Google Scholar] [CrossRef]
- Pathak, V.; Shirota, O.; Sekita, S.; Hirayama, Y.; Hakamata, Y.; Hayashi, T.; Yanagawa, T.; Satake, M. Antiandrogenic phenolic constituents from Dalbergia cochinchinensis. Phytochemistry 1997, 46, 1219–1223. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Z.; Lu, X.; Zhu, X.; Huang, Q.; Liang, J.; Xu, J. Occupational toxic epidermal necrolysis associated with dalbergia cochinchinensis: A retrospective comparative study of eight cases in China. Int. J. Dermatol. 2015, 54, 1435–1441. [Google Scholar] [CrossRef]
- Sun, K.; Su, C.; Li, W.; Gong, Z.; Sha, C.; Liu, R. Quality markers based on phytochemical analysis and anti-inflammatory screening: An integrated strategy for the quality control of Dalbergia odorifera by UHPLC-Q-Orbitrap HRMS. Phytomedicine 2021, 84, 153511. [Google Scholar] [CrossRef]
- Lee, C.; Lee, J.W.; Jin, Q.; Jang, D.S.; Lee, S.J.; Lee, D.; Hong, J.T.; Kim, Y.; Lee, M.K.; Hwang, B.Y. Inhibitory constituents of the heartwood of Dalbergia odorifera on nitric oxide production in RAW 264.7 macrophages. Bioorg. Med. Chem. Lett. 2013, 23, 4263–4266. [Google Scholar] [CrossRef]
- Lai, X.X.; Zhang, N.; Chen, L.Y.; Luo, Y.Y.; Shou, B.Y.; Xie, X.X.; Liu, R.H. Latifolin protects against myocardial infarction by alleviating myocardial inflammatory via the HIF-1alpha/NF-kappaB/IL-6 pathway. Pharm. Biol. 2020, 58, 1156–1166. [Google Scholar] [CrossRef]
- Zhang, N.; Shou, B.; Chen, L.; Lai, X.; Luo, Y.; Meng, X.; Liu, R. Cardioprotective Effects of Latifolin Against Doxorubicin-Induced Cardiotoxicity by Macrophage Polarization in Mice. J. Cardiovasc. Pharmacol. 2020, 75, 564–572. [Google Scholar] [CrossRef]
- Kim, E.N.; Kim, Y.G.; Lee, J.H.; Min, B.S.; Jeong, G.S. 6,7,4′-Trihydroxyflavone inhibits osteoclast formation and bone resorption in vitro and in vivo. Phytother. Res. 2019, 33, 2948–2959. [Google Scholar] [CrossRef]
- Choi, H.S.; Park, J.A.; Hwang, J.S.; Ham, S.A.; Yoo, T.; Lee, W.J.; Paek, K.S.; Shin, H.C.; Lee, C.H.; Seo, H.G. A Dalbergia odorifera extract improves the survival of endotoxemia model mice by inhibiting HMGB1 release. BMC Complement. Altern. Med. 2017, 17, 212. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kim, J.Y.; Cheon, Y.H.; Kwak, S.C.; Baek, J.M.; Kim, Y.C.; Yoon, K.H.; Oh, J.; Lee, M.S. 9-Hydroxy-6,7-dimethoxydalbergiquinol inhibits osteoclast differentiation through down-regulation of Akt, c-Fos and NFATc1. Int. Immunopharmacol. 2014, 20, 213–220. [Google Scholar] [CrossRef]
- Im, N.K.; Choi, J.Y.; Oh, H.; Kim, Y.C.; Jeong, G.S. 6,4′-Dihydroxy-7-methoxyflavanone inhibits osteoclast differentiation and function. Biol. Pharm. Bull. 2013, 36, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jiang, W.; Wang, Y. Anti-inflammation of flavonoid compounds from Dalbergia odorifera T. Chen in lipopolysaccharide stimulated RAW264.7 macrophages. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi=Chin. J. Cell. Mol. Immunol. 2013, 29, 681–684. [Google Scholar]
- Panahipour, L.; Tabatabaei, A.A.; Gruber, R. Hypoallergenic infant formula lacks transforming growth factor beta activity and has a lower anti-inflammatory activity than regular infant formula. J. Dairy Sci. 2020, 103, 6771–6781. [Google Scholar] [CrossRef]
- Jia, L.; Han, N.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Pathogenesis of Important Virulence Factors of Porphyromonas gingivalis via Toll-Like Receptors. Front. Cell Infect. Microbiol. 2019, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.D.; Cvikl, B.; Lussi, A.; Gruber, R. Chemokine expression of oral fibroblasts and epithelial cells in response to artificial saliva. Clin. Oral. Investig. 2016, 20, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyabe, Y.; Lian, J.; Miyabe, C.; Luster, A.D. Chemokines in rheumatic diseases: Pathogenic role and therapeutic implications. Nat. Rev. Rheumatol. 2019, 15, 731–746. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.-H.; Wen, X.-C.; Shao, F.; Zhang, P.-Z.; Huang, H.-L.; Zhang, S. Flavonoids from Heartwood of Dalbergia cochinchinensis. Chin. Herb. Med. 2016, 8, 89–93. [Google Scholar] [CrossRef]
- Liu, R.-h.; Wen, X.-C.; Zhang, P.-Z.; Shao, F.; Yang, Z.-H.; Huang, H.-L.; Wang, Y. Chemical constituents of isoflavonoids from Dalbergia cochinchinensis. Chin. Tradit. Herb. Drugs 2015, 46, 2851–2855. [Google Scholar] [CrossRef]
- Bojase, G.; Wanjala, C.C.W.; Majinda, R.R.T. Two new isoflavanoids from bolusanthus speciosus. Bull Chem. Soc. Ethiop. 2001, 15, 131–136. [Google Scholar] [CrossRef]
- Panahipour, L.; Kochergina, E.; Laggner, M.; Zimmermann, M.; Mildner, M.; Ankersmit, H.J.; Gruber, R. Role for Lipids Secreted by Irradiated Peripheral Blood Mononuclear Cells in Inflammatory Resolution in Vitro. Int. J. Mol. Sci. 2020, 21, 4694. [Google Scholar] [CrossRef]
- Panahipour, L.; Kochergina, E.; Kreissl, A.; Haiden, N.; Gruber, R. Milk modulates macrophage polarization in vitro. Cytokine X 2019, 1, 100009. [Google Scholar] [CrossRef]
- Tang, P.M.; Nikolic-Paterson, D.J.; Lan, H.Y. Macrophages: Versatile players in renal inflammation and fibrosis. Nat. Rev. Nephrol. 2019, 15, 144–158. [Google Scholar] [CrossRef]
- Na, Y.R.; Stakenborg, M.; Seok, S.H.; Matteoli, G. Macrophages in intestinal inflammation and resolution: A potential therapeutic target in IBD. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 531–543. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, C.; Meng, H.; Yu, Z.; Yang, M.; Wei, J. Dalbergia odorifera: A review of its traditional uses, phytochemistry, pharmacology, and quality control. J. Ethnopharmacol. 2020, 248, 112328. [Google Scholar] [CrossRef]
- The, S.N. A Review on the Medicinal Plant Dalbergia odorifera Species: Phytochemistry and Biological Activity. Evid.-Based Complement. Alternat. Med. 2017, 2017, 7142370. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Dai, C.; Wang, J.; Liu, R.; Li, L.; Yin, Z.S. Therapeutic effect of neohesperidin on TNF-alpha-stimulated human rheumatoid arthritis fibroblast-like synoviocytes. Chin. J. Nat. Med. 2021, 19, 741–749. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Liu, M.; Zhou, B.; Yang, G. Diosmetin exhibits anti-proliferative and anti-inflammatory effects on TNF-alpha-stimulated human rheumatoid arthritis fibroblast-like synoviocytes through regulating the Akt and NF-kappaB signaling pathways. Phytother. Res. 2020, 34, 1310–1319. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Q. Kaempferitrin inhibits proliferation, induces apoptosis, and ameliorates inflammation in human rheumatoid arthritis fibroblast-like synoviocytes. Phytother. Res. 2019, 33, 1726–1735. [Google Scholar] [CrossRef]
- Xiong, G.; Ji, W.; Wang, F.; Zhang, F.; Xue, P.; Cheng, M.; Sun, Y.; Wang, X.; Zhang, T. Quercetin Inhibits Inflammatory Response Induced by LPS from Porphyromonas gingivalis in Human Gingival Fibroblasts via Suppressing NF-kappaB Signaling Pathway. Biomed Res. Int. 2019, 2019, 6282635. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Venegas, G.; Contreras-Sanchez, A. Luteolin and fisetin inhibit the effects of lipopolysaccharide obtained from Porphyromonas gingivalis in human gingival fibroblasts. Mol. Biol. Rep. 2013, 40, 477–485. [Google Scholar] [CrossRef]
- Lin, P.; Niimi, H.; Ohsugi, Y.; Tsuchiya, Y.; Shimohira, T.; Komatsu, K.; Liu, A.; Shiba, T.; Aoki, A.; Iwata, T.; et al. Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. Int. J. Mol. Sci. 2021, 22, 8900. [Google Scholar] [CrossRef]
- Lee, D.S.; Li, B.; Im, N.K.; Kim, Y.C.; Jeong, G.S. 4,2′,5′-trihydroxy-4′-methoxychalcone from Dalbergia odorifera exhibits anti-inflammatory properties by inducing heme oxygenase-1 in murine macrophages. Int. Immunopharmacol. 2013, 16, 114–121. [Google Scholar] [CrossRef]
- Lee, D.S.; Kim, K.S.; Ko, W.; Li, B.; Keo, S.; Jeong, G.S.; Oh, H.; Kim, Y.C. The neoflavonoid latifolin isolated from MeOH extract of Dalbergia odorifera attenuates inflammatory responses by inhibiting NF-kappaB activation via Nrf2-mediated heme oxygenase-1 expression. Phytother. Res. 2014, 28, 1216–1223. [Google Scholar] [CrossRef]
- Domblides, C.; Lartigue, L.; Faustin, B. Metabolic Stress in the Immune Function of T Cells, Macrophages and Dendritic Cells. Cells 2018, 7, 68. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mIL1β | AAGGGCTGCTTCCAAACCTTTGAC | ATACTGCCTGCCTGAAGCTCTTGT |
| mIL6 | GCTACCAAACTGGATATAATCAGGA | CCAGGTAGCTATGGTACTCCAGAA |
| mCCL2 | GCTACAAGAGGATCACCAGCAG | GTCTGGACCCATTCCTTCTTGG |
| mGAPDH | AACTTTGGCATTGTCGAACG | GGATGCAGGGATGATGTTCT |
| hCXCL1 | TCCTGCATCCCCCATAGTTA | CTTCAGGAACAGCCACCAGT |
| hCXCL2 | CCCATGGTTAAGAAAATCATCG | CTTCAGGAACAGCCACCAAT |
| hGAPDH | AAGCCACATCGCTC AGACAC | GCCCAATACGACCAAATCC |
Publisher′s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, F.; Panahipour, L.; Sordi, M.B.; Tang, F.; Liu, R.; Gruber, R. Heartwood of Dalbergia cochinchinensis: 4,7,2′-Trihydroxy-4′-methoxyisoflavanol and 6,4′-Dihydroxy-7-methoxyflavane Reduce Cytokine and Chemokine Expression In Vitro. Molecules 2022, 27, 1321. https://doi.org/10.3390/molecules27041321
Shao F, Panahipour L, Sordi MB, Tang F, Liu R, Gruber R. Heartwood of Dalbergia cochinchinensis: 4,7,2′-Trihydroxy-4′-methoxyisoflavanol and 6,4′-Dihydroxy-7-methoxyflavane Reduce Cytokine and Chemokine Expression In Vitro. Molecules. 2022; 27(4):1321. https://doi.org/10.3390/molecules27041321
Chicago/Turabian StyleShao, Feng, Layla Panahipour, Mariane Beatriz Sordi, Fangrui Tang, Ronghua Liu, and Reinhard Gruber. 2022. "Heartwood of Dalbergia cochinchinensis: 4,7,2′-Trihydroxy-4′-methoxyisoflavanol and 6,4′-Dihydroxy-7-methoxyflavane Reduce Cytokine and Chemokine Expression In Vitro" Molecules 27, no. 4: 1321. https://doi.org/10.3390/molecules27041321