
New Eco-Sustainable Feed in Aquaculture: Influence of Insect-Based Diets on the Content of Potentially Toxic Elements in the Experimental Model Zebrafish (Danio rerio)

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Potentially Toxic Elements in Water Tanks
2.2. Potentially Toxic Elements in the Experimental Diets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Cd | Pb | Hg | As | Ni |
|---|---|---|---|---|---|
| Legal Limit for Fish Feed | 1.0 | 5.0 | 0.2 | 10 | - |
| Hi meal * | 0.20 ± 0.02 | 0.059 ± 0.007 | 0.053 ± 0.003 | 0.109 ± 0.003 | 1.06 ± 0.04 |
| CTRL | 0.38 ± 0.02 b | 0.63 ± 0.04 c | 0.033 ± 0.001 a | 0.21 ± 0.01 c | 1.72 ± 0.04 d |
| Hi25 | 0.31 ± 0.02 a | 0.71 ± 0.04 c | 0.035 ± 0.001 a | 0.14 ± 0.01 b | 1.04 ± 0.11 c |
| Hi50 | 0.45 ± 0.02 c | 0.42 ± 0.02 b | 0.037 ± 0.001 a | 0.12 ± 0.01 a | 0.20 ± 0.02 b |
| Hi75 | 0.51 ± 0.03 c | 0.34 ± 0.02 a | 0.032 ± 0.001 a | 0.12 ± 0.01a | 0.21 ± 0.03 b |
| Hi100 | 0.46 ± 0.04 c | 0.31 ± 0.02 a | 0.034 ± 0.002 a | 0.11 ± 0.01 a | 0.11 ± 0.01 a |
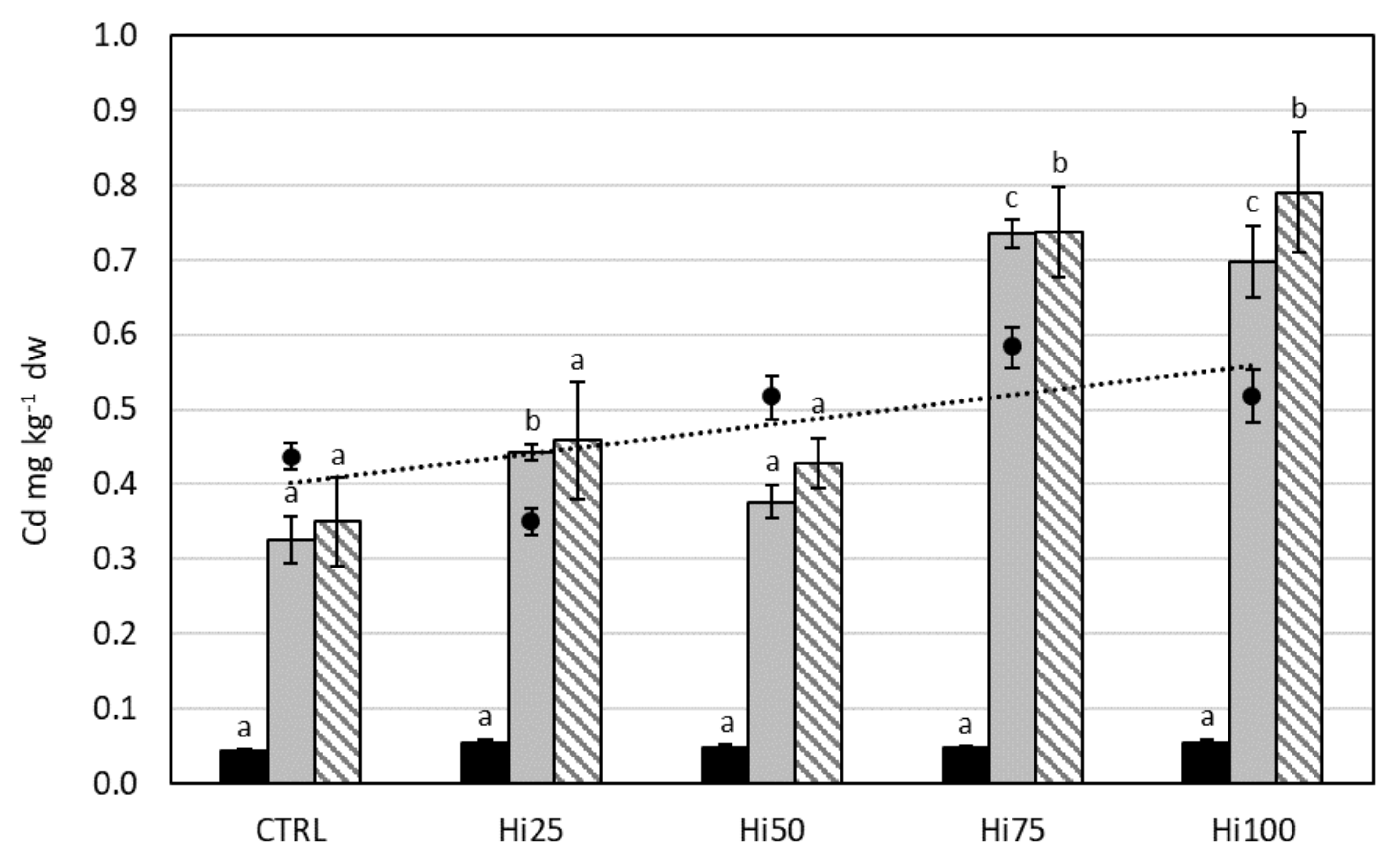
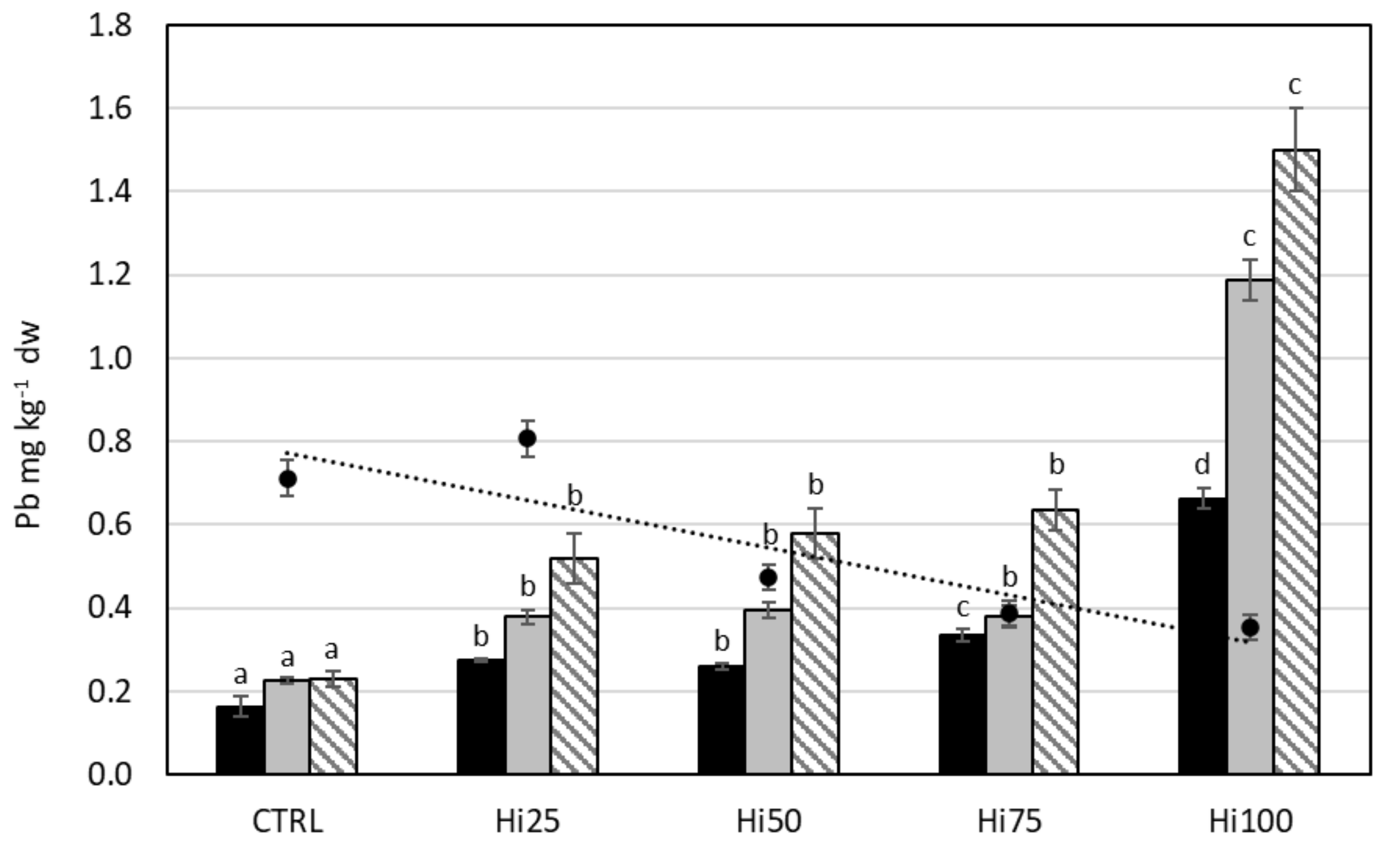
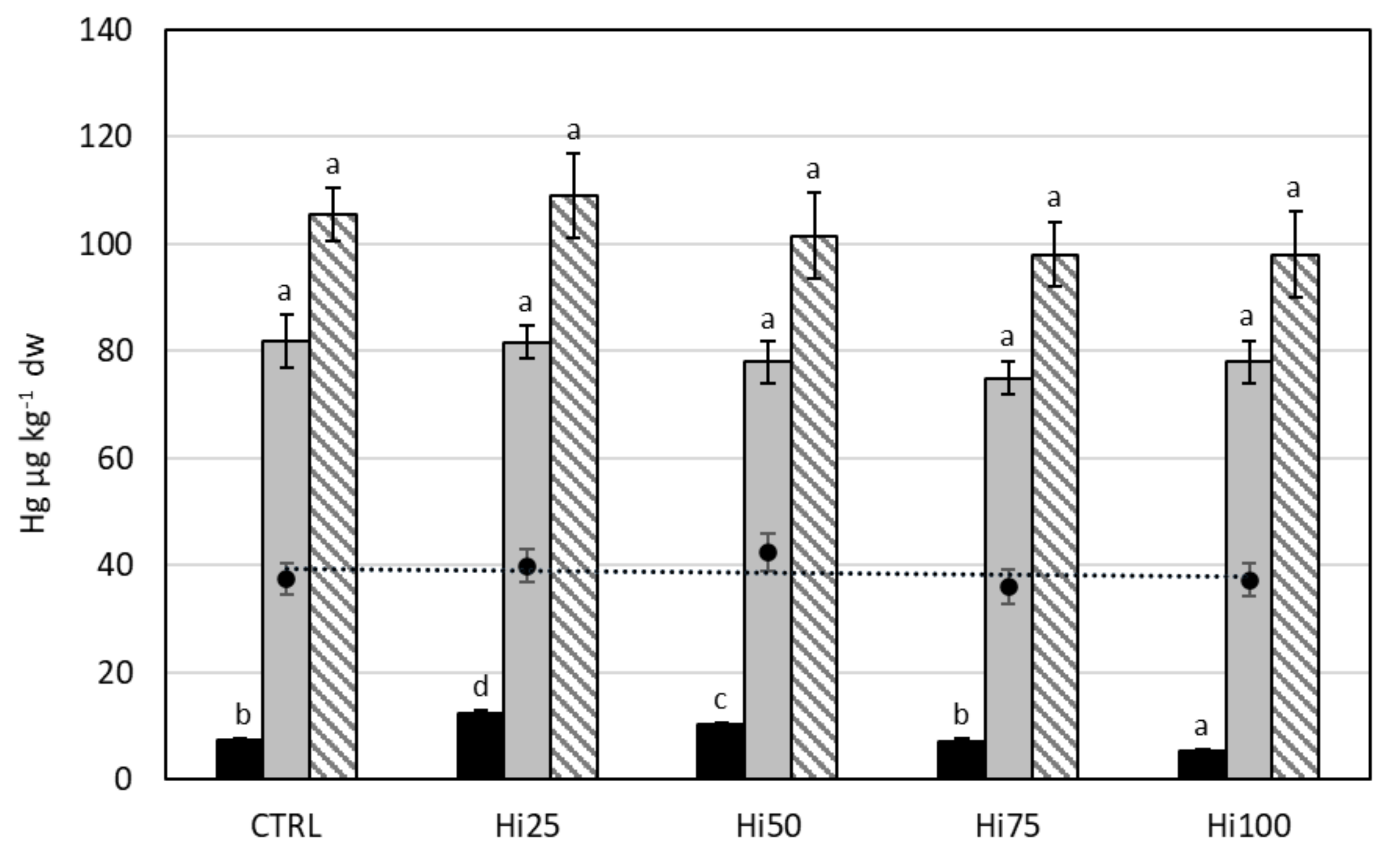
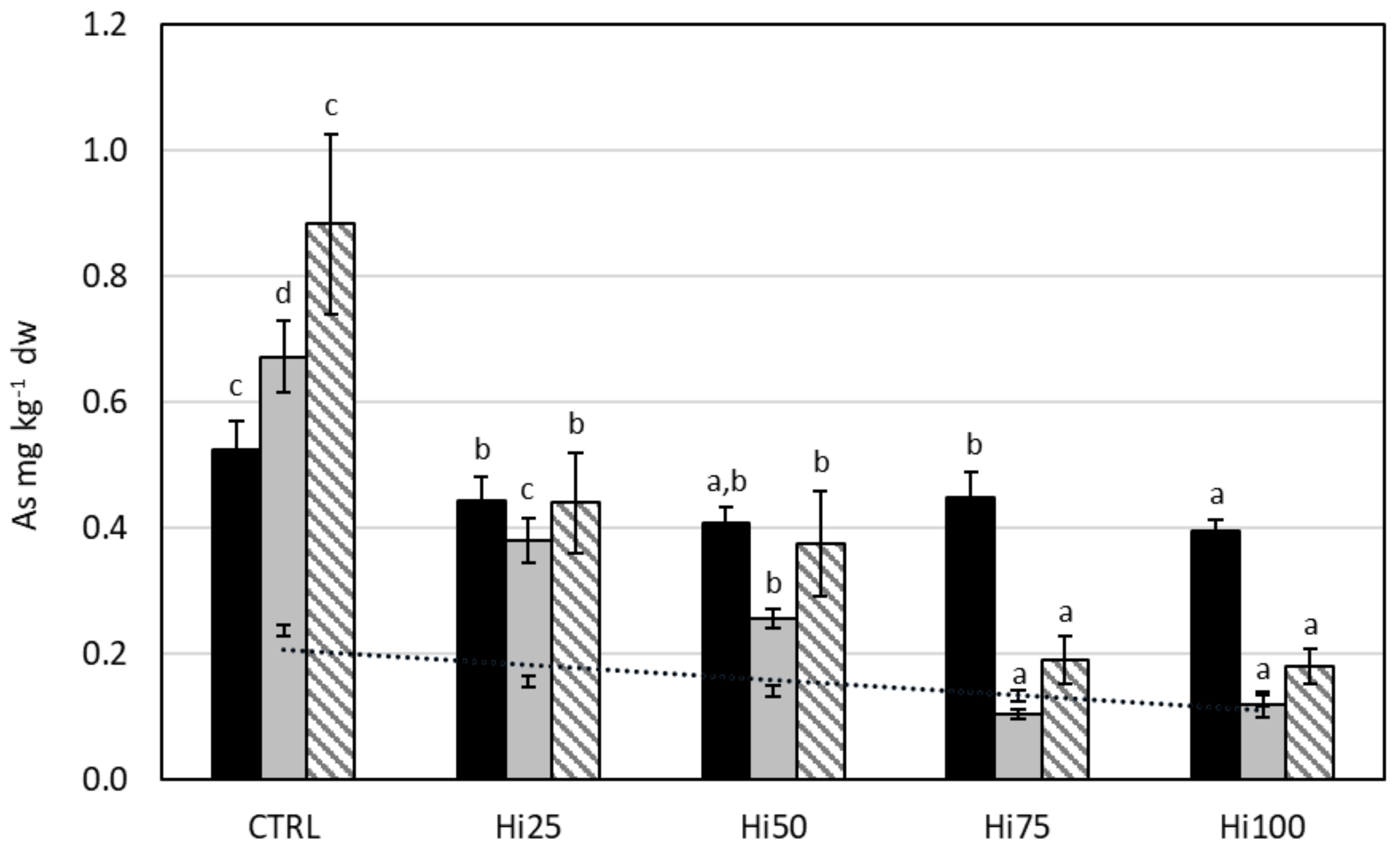
2.3. Potentially Toxic Elements in Danio rerio
2.3.1. Cadmium

2.3.2. Lead

2.3.3. Mercury

2.3.4. Arsenic

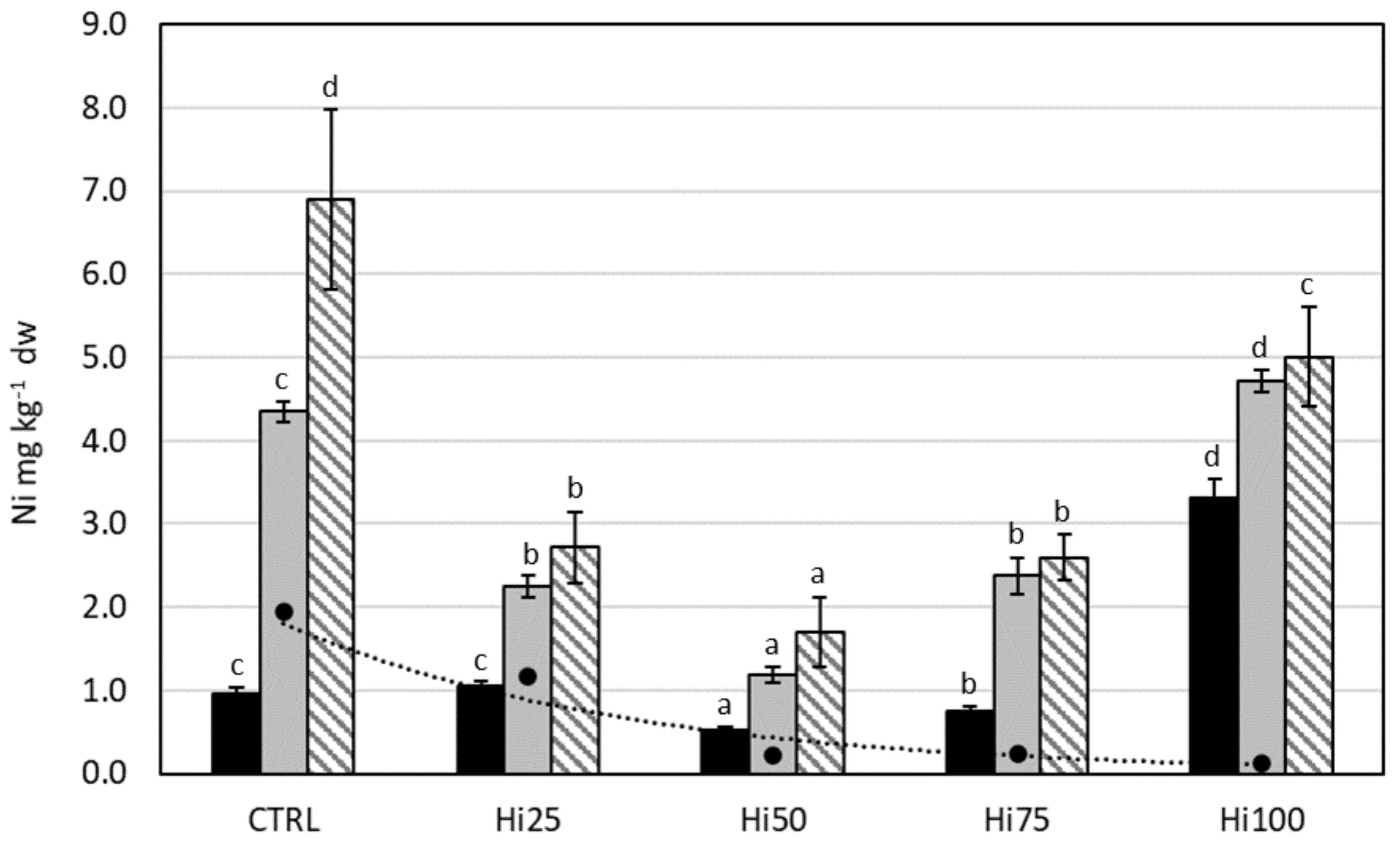
2.3.5. Nickel

3. Discussion
3.1. Potentially Toxic Elements in the Experimental Diets
3.2. Potentially Toxic Elements in Danio rerio
3.2.1. Cadmium
3.2.2. Lead
3.2.3. Mercury
3.2.4. Arsenic
3.2.5. Nickel
| Life Stage | Organ | Exposure | PTEs Content (mg kg−1 ww or dw) | Ref |
|---|---|---|---|---|
| Cd | ||||
| larvae | whole | natural | 0.044–0.054 (dw) 0.0068–0.0082 (ww) | this study |
| juvenile | carcass | 0.32–0.73 (dw) 0.08–0.16 (ww) | this study | |
| adult | carcass | 0.35–0.79 (dw) 0.11–0.25 (ww) | this study | |
| larvae | carcass | natural | ~0.02 (ww) | [33] |
| adult | carcass | natural | 0.02 (dw) | [33] |
| adult | whole | natural | 1.5 (dw) | [10] |
| 10 ppt | ~8 (dw) | |||
| adult | brain | natural | 0.4 (ww) | [71] |
| 1 ppm (2 days) | 20.2 (ww) | |||
| adult | muscle | natural | 0.200 ± 0.03 (ww) | [75] |
| 1.9 0.6 ppb | 0.18 ± 0.04 (ww) | |||
| 9.6 ± 2.9 ppb | 0.14 ± 0.04 (ww) | |||
| embryos | whole | ~0.2 (dw) | [70] | |
| adult | liver and gills | river water | 4–27 (ww) | [68] |
| adult | carcass | 0.5 ppm | ~0.75 (ww) | [83] |
| Pb | ||||
| larvae | whole | natural | 0.16–0.66 (dw) 0.025–0.101 (ww) | this study |
| juvenile | carcass | 0.23–1.19 (dw) 0.055–0.26 (ww) | this study | |
| adult | carcass | 0.23–1.30 (dw) 0.072–0.42 (ww) | this study | |
| adult | liver and gills | river water | 0.2–38 (ww) | [68] |
| adult | carcass | 1 ppm | ~2 (ww) | [83] |
| Hg | ||||
| larvae | whole | natural | 0.009 ± 0.003 (dw) 0.0013 ± 0.0004 (ww) | this study |
| juvenile | carcass | 0.079 ± 0.003 (dw) 0.018 ± 0.001 (ww) | this study | |
| adult | carcass | 0.102 ± 0.005 (dw) 0.032 ± 0.002 (ww) | this study | |
| adult | muscle | natural | next to 0 (dw) | [76] |
| 5 ppm (7 days) | ~3 (dw) | |||
| 5 ppm (21 days) | ~8 (dw) | |||
| 5 ppm (63 days) | ~15 (dw) | |||
| adult | muscle | 60 ± 10 ppb (diet) | ~0.6 (dw) | [72] |
| 11.6 ± 0.4 ppm | ~30 (dw) | |||
| As | ||||
| larvae | whole | natural | 0.39–0.52 (dw) 0.060–0.080 (ww) | this study |
| juvenile | carcass | 0.11–0.67 (dw) 0.026–0.16 (ww) | this study | |
| adult | carcass | 0.18–0.88 (dw) 0.058–0.27 (ww) | this study | |
| larvae | whole | 5 ppm | 0.4–0.5 (ww) | [73] |
| adult | muscle | 5 ppm | 0.4–0.5 (ww) | |
| adult | natural | 0.94 ± 0.08 (dw) | [77] | |
| adult | carcass | natural | 0.27–0.39 (dw) | [33] |
| adult | liver and gills | river water | 4–40 (ww) | [68] |
| Ni | ||||
| larvae | whole | natural | 0.52–3.3 (dw) 0.089–0.50 (ww) | this study |
| juvenile | carcass | 0.19–4.7 (dw) 0.27–1.03 (ww) | this study | |
| adult | carcass | 1.7–6.90 (dw) 0.54–2.1 (ww) | this study | |
| adult | muscle | natural | 0.79 ± 0.03 (ww) | [74] |
| carcass | natural | 2.19 ± 0.19 (ww) | ||
4. Materials and Methods
4.1. Ethics
4.2. Fish Diet Production
4.3. Experimental Design
4.4. Chemical Analyses and Quality Control
4.5. Bioaccumulation Factor
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Sample Availability
References
- FAO-GFCM. Fishery and Aquaculture Statistics. GFCM Capture Production 1970–2019; FAO Fisheries and Aquaculture Department: Rome, Italy, 2020; Available online: https://www.fao.org/gfcm/data/capture-production/en/ (accessed on 24 November 2021).
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; Food and Agricultural Organization: Rome, Italy, 2018. [Google Scholar]
- EFSA Scientific Committee. Risk Profile Related to Production and Consumption of Insects as Food and Feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-Art on Use of Insects as Animal Feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G.A.B. The Environmental Sustainability of Insects as Food and Feed. A Review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of Black Soldier Fly (Diptera: Stratiomyidae) Larvae to Recycle Food Waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Fowles, T.M.; Nansen, C. Insect-Based Bioconversion: Value from Food Waste. In Food Waste Management: Solving the Wicked Problem; Springer International Publishing: Cham, Switzerland, 2019; pp. 321–346. ISBN 9783030205614. [Google Scholar]
- Olsen, R.L.; Hasan, M.R. A Limited Supply of Fishmeal: Impact on Future Increases in Global Aquaculture Production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the Use of Insects in the Diet of Farmed Fish: Past and Future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Måge, A.; Julshamn, K.; Hemre, G.I.; Lunestad, B.T. Norwegian Surveillance Program on Feed for Fish and Other Aquatic Organisms; Annual Report 2007; National Institute of Nutrition and Seafood Research: Bergen, Norway, 2008; www.mattilsynet.no; www.nifes.no; (accessed on 24 November 2021). (In Norwegian) [Google Scholar]
- Amlund, H.; Berntssen, M.H.G.; Lunestad, B.T.; Lundebye, A.K. Aquaculture Feed Contamination by Persistent Organic Pollutants, Heavy Metals, Additives and Drug Residues. In Animal Feed Contamination: Effects on Livestock and Food Safety; Elsevier: Amsterdam, The Netherlands, 2012; pp. 205–229. ISBN 9781845697259. [Google Scholar]
- Do’rea, J.G. Fish Meal in Animal Feed and Human Exposure to Persistent Bioaccumulative and Toxic Substances. J. Food Prot. 2006, 69, 2777–2785. [Google Scholar] [CrossRef]
- Sissener, N.H.; Julshamn, K.; Espe, M.; Lunestad, B.T.; Hemre, G.I.; Waagbø, R.; Måge, A. Surveillance of Selected Nutrients, Additives and Undesirables in Commercial Norwegian Fish Feeds in the Years 2000–2010. Aquac. Nutr. 2013, 19, 555–572. [Google Scholar] [CrossRef]
- Berntssen, M.H.G.; Julshamn, K.; Lundebye, A.K. Chemical Contaminants in Aquafeeds and Atlantic Salmon (Salmo salar) Following the Use of Traditional- versus Alternative Feed Ingredients. Chemosphere 2010, 78, 637–646. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Bioaccumulation of Heavy Metals in the Black Soldier Fly, Hermetia illucens and Effects on Its Life Cycle. J. Insects Food Feed 2015, 1, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Truzzi, C.; Annibaldi, A.; Girolametti, F.; Giovannini, L.; Riolo, P.; Ruschioni, S.; Olivotto, I.; Illuminati, S. A Chemically Safe Way to Produce Insect Biomass for Possible Application in Feed and Food Production. Int. J. Environ. Res. Public Health 2020, 17, 2121. [Google Scholar] [CrossRef] [Green Version]
- Proc, K.; Bulak, P.; Wiącek, D.; Bieganowski, A. Hermetia Illucens Exhibits Bioaccumulative Potential for 15 Different Elements—Implications for Feed and Food Production. Sci. Total Environ. 2020, 723, 138125. [Google Scholar] [CrossRef]
- Zanoli, P.; Rossi, T.; Vaccari, G.; Giacobazzi, A.; Truzzi, C.; Avallone, R.; Ventura, A. Effetti Di Deficit Ed Accumulo Di Metalli Pesanti Sulla Funzionalità Del Sistema Nervoso Centrale. In Nutrizione e Cervello; Pythagora Press: Milan, Italy, 1992. [Google Scholar]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. EXS 2012, 101, 133–164. [Google Scholar] [CrossRef] [Green Version]
- Illuminati, S.; Annibaldi, A.; Truzzi, C.; Tercier-Waeber, M.L.; Nöel, S.; Braungardt, C.B.; Achterberg, E.P.; Howell, K.A.; Turner, D.; Marini, M.; et al. In-Situ Trace Metal (Cd, Pb, Cu) Speciation along the Po River Plume (Northern Adriatic Sea) Using Submersible Systems. Mar. Chem. 2019, 212, 47–63. [Google Scholar] [CrossRef]
- Sethy, K.; Pati, S.; Jena, D.; Mishra, C.K. Heavy Metal Toxicity in Animals: A Review. Pharma Innov. J. 2020, 9, 134–137. [Google Scholar]
- Jin, M.; Yuan, H.; Liu, B.; Peng, J.; Xu, L.; Yang, D. Review of the Distribution and Detection Methods of Heavy Metals in the Environment. Anal. Methods 2020, 12, 5747–5766. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Agents Classified by the IARC Monographs, Volumes 1–130; International Agency for Research on Cancer: Lyon, France, 2021. [Google Scholar]
- Godt, J.; Scheidig, F.; Grosse-Siestrup, C.; Esche, V.; Brandenburg, P.; Reich, A.; Groneberg, D.A. The Toxicity of Cadmium and Resulting Hazards for Human Health. J. Occup. Med. Toxicol. 2006, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Goyer, R.A. Lead Toxicity: From Overt to Subclinical to Subtle Health Effects. Environ. Health Perspect. 1990, 86, 177–181. [Google Scholar] [CrossRef]
- Zahir, F.; Rizwi, S.J.; Haq, S.K.; Khan, R.H. Low Dose Mercury Toxicity and Human Health. Environ. Toxicol. Pharmacol. 2005, 20, 351–360. [Google Scholar] [CrossRef]
- Bibi, S.; Kamran, M.A.; Sultana, J.; Farooqi, A. Occurrence and Methods to Remove Arsenic and Fluoride Contamination in Water. Environ. Chem. Lett. 2017, 15, 125–149. [Google Scholar] [CrossRef]
- Cempel, M.; Nikel, G. Nickel: A Review of Its Sources and Environmental Toxicology. Pol. J. Environ. Stud. 2006, 15, 375–382. [Google Scholar]
- Wood, C.M.; Farrell, A.P.; Brauner, C.J. Homeostasis and Toxicology of Essential Metals; Academic Press: Cambridge, MA, USA, 2012; Volume 31A. [Google Scholar]
- Wood, C.M.; Farrell, A.P.; Brauner, C.J. Homeostasis and Toxicology of Non-Essential Metals; Academic Press: Cambridge, MA, USA, 2012; Volume 31B. [Google Scholar]
- Luoma, S.N.; Rainbow, P.S. Why Is Metal Bioaccumulation so Variable? Biodynamics as a Unifying Concept. Environ. Sci. Technol. 2005, 39, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Boyle, D.; Hogstrand, C.; Bury, N.R. Physiological Response to a Metal-Contaminated Invertebrate Diet in Zebrafish: Importance of Metal Speciation and Regulation of Metal Transport Pathways. Aquat. Toxicol. 2011, 105, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Boyle, D.; Brix, K.V.; Amlund, H.; Lundebye, A.K.; Hogstrand, C.; Bury, N.R. Natural Arsenic Contaminated Diets Perturb Reproduction in Fish. Environ. Sci. Technol. 2008, 42, 5354–5360. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef] [Green Version]
- DeForest, D.K.; Brix, K.V.; Adams, W.J. Assessing Metal Bioaccumulation in Aquatic Environments: The Inverse Relationship between Bioaccumulation Factors, Trophic Transfer Factors and Exposure Concentration. Aquat. Toxicol. 2007, 84, 236–246. [Google Scholar] [CrossRef]
- Zhuang, P.; Li, Z.; McBride, M.B.; Zou, B.; Wang, G. Health Risk Assessment for Consumption of Fish Originating from Ponds near Dabaoshan Mine, South China. Environ. Sci. Pollut. Res. 2013, 20, 5844–5854. [Google Scholar] [CrossRef]
- Rahman, M.S.; Molla, A.H.; Saha, N.; Rahman, A. Study on Heavy Metals Levels and Its Risk Assessment in Some Edible Fishes from Bangshi River, Savar, Dhaka, Bangladesh. Food Chem. 2012, 134, 1847–1854. [Google Scholar] [CrossRef]
- Girolametti, F.; Annibaldi, A.; Carnevali, O.; Pignalosa, P.; Illuminati, S.; Truzzi, C. Potential Toxic Elements (PTEs) in Wild and Farmed Atlantic Bluefin Tuna (Thunnus thynnus) from Mediterranean Sea: Risks and Benefits for Human Consumption. Food Control 2021, 125, 108012. [Google Scholar] [CrossRef]
- Fakhri, Y.; Mohseni-Bandpei, A.; Oliveri Conti, G.; Ferrante, M.; Cristaldi, A.; Jeihooni, A.K.; Karimi Dehkordi, M.; Alinejad, A.; Rasoulzadeh, H.; Mohseni, S.M.; et al. Systematic Review and Health Risk Assessment of Arsenic and Lead in the Fished Shrimps from the Persian Gulf. Food Chem. Toxicol. 2018, 113, 278–286. [Google Scholar] [CrossRef]
- Rahmani, J.; Fakhri, Y.; Shahsavani, A.; Bahmani, Z.; Urbina, M.A.; Chirumbolo, S.; Keramati, H.; Moradi, B.; Bay, A.; Bjørklund, G. A Systematic Review and Meta-Analysis of Metal Concentrations in Canned Tuna Fish in Iran and Human Health Risk Assessment. Food Chem. Toxicol. 2018, 118, 753–765. [Google Scholar] [CrossRef]
- Annibaldi, A.; Truzzi, C.; Carnevali, O.; Pignalosa, P.; Api, M.; Scarponi, G.; Illuminati, S. Determination of Hg in Farmed and Wild Atlantic Bluefin Tuna (Thunnus thynnus L.) Muscle. Molecules 2019, 24, 1273. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.; Khan, E. Bioaccumulation of Non-Essential Hazardous Heavy Metals and Metalloids in Freshwater Fish. Risk to Human Health. Environ. Chem. Lett. 2018, 16, 903–917. [Google Scholar] [CrossRef]
- Decreto Legislativo 2 Febbraio 2001, n. 31. Attuazione Della Direttiva 98/83/CE Relativa Alla Qualita’ Delle Acque Destinate al Consumo Umano. Pubblicato Nella Gazzetta Ufficiale n. 52 Del 3 Marzo 2001—Supplemento Ordinario n. 41. Available online: https://www.camera.it/parlam/leggi/deleghe/01031dl.htm (accessed on 20 October 2020).
- European Parliament; European Council. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy. Off. J. Eur. Communities 2000, L327, 1–72. [Google Scholar]
- European Parliament; European Council. Directive 2002/32/EC of 7 May 2002 on Undesirable Substances in Animal Feed. Off. J. Eur. Communities 2002, L140, 10–21. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Union 2006, L364, 5–24. [Google Scholar]
- European Parliament; European Council. Commission Regulation (EU) No 420/2011 29 April 2011 Amending Regulation (EC) No 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Communities 2011, L111, 3–6. [Google Scholar]
- European Parliament; European Council. Commission Regulation (EU) No 1006/2011 25 June 2015 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Inorganic Arsenic in Foodstuffs. Off. J. Eur. Communities 2015, L161, 14–16. [Google Scholar]
- Van der Fels-Klerx, H.J.; Camenzuli, L.; van der Lee, M.K.; Oonincx, D.G.A.B. Uptake of Cadmium, Lead and Arsenic by Tenebrio molitor and Hermetia illucens from Contaminated Substrates. PLoS ONE 2016, 11, e0166186. [Google Scholar] [CrossRef]
- Truzzi, C.; Illuminati, S.; Girolametti, F.; Antonucci, M.; Scarponi, G.; Ruschioni, S.; Riolo, P.; Annibaldi, A. Influence of Feeding Substrates on the Presence of Toxic Metals (Cd, Pb, Ni, as, Hg) in Larvae of Tenebrio molitor: Risk Assessment for Human Consumption. Int. J. Environ. Res. Public Health 2019, 16, 4815. [Google Scholar] [CrossRef] [Green Version]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The Potential of Various Insect Species for Use as Food for Fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Faccenda, F.; Cerri, R.; Tibaldi, E.; Olivotto, I. Physiological Response of Rainbow Trout (Oncorhynchus mykiss) to Graded Levels of Hermetia illucens or Poultry by-Product Meals as Single or Combined Substitute Ingredients to Dietary Plant Proteins. Aquaculture 2021, 538, 736550. [Google Scholar] [CrossRef]
- Lock, E.-J.; Biancarosa, I.; Gasco, L. Insects as Raw Materials in Compound Feed for Aquaculture. In Edible Insects in Sustainable Food Systems; Springer International Publishing: Cham, Switzerland, 2018; pp. 263–276. [Google Scholar]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional Value of the Black Soldier Fly (Hermetia illucens L.) and Its Suitability as Animal Feed—A Review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Borgogno, M.; Dinnella, C.; Iaconisi, V.; Fusi, R.; Scarpaleggia, C.; Schiavone, A.; Monteleone, E.; Gasco, L.; Parisi, G. Inclusion of Hermetia Illucens Larvae Meal on Rainbow Trout (Oncorhynchus mykiss) Feed: Effect on Sensory Profile According to Static and Dynamic Evaluations. J. Sci. Food Agric. 2017, 97, 3402–3411. [Google Scholar] [CrossRef]
- Zhou, J.S.; Liu, S.S.; Ji, H.; Yu, H.B. Effect of Replacing Dietary Fish Meal with Black Soldier Fly Larvae Meal on Growth and Fatty Acid Composition of Jian Carp (Cyprinus carpio Var. Jian). Aquac. Nutr. 2018, 24, 424–433. [Google Scholar] [CrossRef]
- Biancarosa, I.; Sele, V.; Belghit, I.; Ørnsrud, R.; Lock, E.J.; Amlund, H. Replacing Fish Meal with Insect Meal in the Diet of Atlantic Salmon (Salmo Salar) Does Not Impact the Amount of Contaminants in the Feed and It Lowers Accumulation of Arsenic in the Fillet. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2019, 36, 1191–1205. [Google Scholar] [CrossRef] [Green Version]
- Truzzi, C.; Giorgini, E.; Annibaldi, A.; Antonucci, M.; Illuminati, S.; Scarponi, G.; Riolo, P.; Isidoro, N.; Conti, C.; Zarantoniello, M.; et al. Fatty Acids Profile of Black Soldier Fly (Hermetia illucens): Influence of Feeding Substrate Based on Coffee-Waste Silverskin Enriched with Microalgae. Anim. Feed Sci. Technol. 2020, 259, 114309. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Cardinaletti, G.; Truzzi, C.; Chemello, G.; Riolo, P.; Olivotto, I. Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase. Animals 2021, 11, 751. [Google Scholar] [CrossRef]
- Dahm, R.; Geisler, R. Learning from Small Fry: The Zebrafish as a Genetic Model Organism for Aquaculture Fish Species. Mar. Biotechnol. 2006, 8, 329–345. [Google Scholar] [CrossRef]
- Hoo, J.Y.; Kumari, Y.; Shaikh, M.F.; Hue, S.M.; Goh, B.H. Zebrafish: A Versatile Animal Model for Fertility Research. BioMed Res. Int. 2016, 2016, 9732780. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, D.J.; Eisen, J.S. Headwaters of the Zebrafish—Emergence of a New Model Vertebrate. Nat. Rev. Genet. 2002, 3, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Piccinetti, C.C.; Migliarini, B.; Olivotto, I.; Simoniello, M.P.; Giorgini, E.; Carnevali, O. Melatonin and Peripheral Circuitries: Insights on Appetite and Metabolism in Danio rerio. Zebrafish 2013, 10, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccinetti, C.C.; Montis, C.; Bonini, M.; Laurà, R.; Guerrera, M.C.; Radaelli, G.; Vianello, F.; Santinelli, V.; Maradonna, F.; Nozzi, V.; et al. Transfer of Silica-Coated Magnetic (Fe3O4) Nanoparticles through Food: A Molecular and Morphological Study in Zebrafish. Zebrafish 2014, 11, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Alsop, D.; Wood, C.M. Metal Uptake and Acute Toxicity in Zebrafish: Common Mechanisms across Multiple Metals. Aquat. Toxicol. 2011, 105, 385–393. [Google Scholar] [CrossRef]
- Dai, Y.J.; Jia, Y.F.; Chen, N.; Bian, W.P.; Li, Q.K.; Ma, Y.B.; Chen, Y.L.; Pei, D.S. Zebrafish as a Model System to Study Toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef]
- Bhuvaneshwari, R.; Rajendran, B. Histopathological Alterations in Muscle, Liver and Gill Tissues of Zebra Fish Danio rerio Due to Environmentally Relevant Concentrations of Organochlo rine Pesticides (OCPs) and Heavy Metals. Int. J. Environ. Res 2015, 9, 1365–1372. [Google Scholar]
- Hu, J.; Liu, J.; Li, J.; Lv, X.; Yu, L.; Wu, K.; Yang, Y. Metal Contamination, Bioaccumulation, ROS Generation, and Epigenotoxicity Influences on Zebrafish Exposed to River Water Polluted by Mining Activities. J. Hazard. Mater. 2021, 405, 124150. [Google Scholar] [CrossRef]
- Lacave, J.M.; Bilbao, E.; Gilliland, D.; Mura, F.; Dini, L.; Cajaraville, M.P.; Orbea, A. Bioaccumulation, Cellular and Molecular Effects in Adult Zebrafish after Exposure to Cadmium Sulphide Nanoparticles and to Ionic Cadmium. Chemosphere 2020, 238, 124588. [Google Scholar] [CrossRef]
- Hallare, A.V.; Schirling, M.; Luckenbach, T.; Köhler, H.R.; Triebskorn, R. Combined Effects of Temperature and Cadmium on Developmental Parameters and Biomarker Responses in Zebrafish (Danio rerio) Embryos. J. Therm. Biol. 2005, 30, 7–17. [Google Scholar] [CrossRef]
- Favorito, R.; Chiarelli, G.; Grimaldi, M.C.; de Bonis, S.; Lancieri, M.; Ferrandino, I. Bioaccumulation of Cadmium and Its Cytotoxic Effect on Zebrafish Brain. Chem. Ecol. 2011, 27, 39–46. [Google Scholar] [CrossRef]
- Feng, C.; Pedrero, Z.; Gentès, S.; Barre, J.; Renedo, M.; Tessier, E.; Berail, S.; Maury-Brachet, R.; Mesmer-Dudons, N.; Baudrimont, M.; et al. Specific Pathways of Dietary Methylmercury and Inorganic Mercury Determined by Mercury Speciation and Isotopic Composition in Zebrafish (Danio rerio). Environ. Sci. Technol. 2015, 49, 12984–12993. [Google Scholar] [CrossRef]
- Hamdi, M.; Sanchez, M.A.; Beene, L.C.; Liu, Q.; Landfear, S.M.; Rosen, B.P.; Liu, Z. Arsenic Transport by Zebrafish Aquaglyceroporins. BMC Mol. Biol. 2009, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Alsop, D.; Lall, S.P.; Wood, C.M. Reproductive Impacts and Physiological Adaptations of Zebrafish to Elevated Dietary Nickel. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 165, 67–75. [Google Scholar] [CrossRef]
- Gonzalez, P.; Baudrimont, M.; Boudou, A.; Bourdineaud, J.-P. Comparative Effects of Direct Cadmium Contamination on Gene Expression in Gills, Liver, Skeletal Muscles and Brain of the Zebrafish (Danio rerio). BioMetals 2006, 19, 225–235. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Wang, X.; Ren, J.; Jia, P.; Fan, W. Influence of Humic Acid on Arsenic Bioaccumulation and Biotransformation to Zebrafish: A Comparative Study between As(III) and As(V) Exposure. Environ. Pollut. 2020, 256, 113459. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black Soldier Fly (Hermetia illucens) Reared on Roasted Coffee by-Product and Schizochytrium Sp. as a Sustainable Terrestrial Ingredient for Aquafeeds Production. Aquaculture 2020, 518, 734659. [Google Scholar] [CrossRef]
- Qiu, Y.W.; Lin, D.; Liu, J.Q.; Zeng, E.Y. Bioaccumulation of Trace Metals in Farmed Fish from South China and Potential Risk Assessment. Ecotoxicol. Environ. Saf. 2011, 74, 284–293. [Google Scholar] [CrossRef]
- Mount, D.R.; Barth, A.K.; Garrison, T.D.; Barten, K.A.; Hockett, J.R. Dietary and Waterborne Exposure of Rainbow Trout (Oncorhynchus mykiss) to Copper, Cadmium, Lead and Zinc Using a Live Diet. Environ. Toxicol. Chem. Int. J. 1994, 13, 2031–2041. [Google Scholar] [CrossRef]
- Monge-Ortiz, R.; Martínez-Llorens, S.; Lemos-Neto, M.J.; Falcó-Giaccaglia, S.L.; Pagán, M.J.; Godoy-Olmos, S.; Jover-Cerdá, M.; Tomás-Vidal, A. Growth, Sensory and Chemical Characterization of Mediterranean Yellowtail (Seriola dumerili) Fed Diets with Partial Replacement of Fish Meal by Other Protein Sources. Aquac. Rep. 2020, 18, 100466. [Google Scholar] [CrossRef]
- Chemello, G.; Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Cardinaletti, G.; Riolo, P.; Olivotto, I. Effects of Black Soldier Fly (Hermetia illucens) Enriched with Schizochytrium Sp. on Zebrafish (Danio Rerio) Reproductive Performances. Aquaculture 2022, 550, 737853. [Google Scholar] [CrossRef]
- Annabi, A.; Said, K.; Messaoudi, I. Cadmium: Bioaccumulation, Histopathology and Detoxifying Mechanisms in Fish. Am. J. Res. Commun. 2013, 1, 62–79. [Google Scholar]
- Zhang, Y.; Feng, J.; Gao, Y.; Liu, X.; Qu, L.; Zhu, L. Physiologically Based Toxicokinetic and Toxicodynamic (PBTK-TD) Modelling of Cd and Pb Exposure in Adult Zebrafish Danio rerio: Accumulation and Toxicity. Environ. Pollut. 2019, 249, 959–968. [Google Scholar] [CrossRef]
- Romeo, M.; Siau, Y.; Sidoumou, Z.; Gnassia-Barelli, M. Heavy Metal Distribution in Different Fish Species from the Mauritania Coast. Sci. Total Environ. 1999, 232, 169–175. [Google Scholar] [CrossRef]
- Francis, P.C.; Birge, W.J.; Bwxt, J.A. Effects of Cadmium-Enriched Sediment on Fish and Amphibian Embryo-Larval Stages. Ecotoxicol. Environ. Saf. 1984, 8, 378–387. [Google Scholar] [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms Controlling Mitochondrial Biogenesis and Respiration through the Thermogenic Coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.E.; Klaverkamp, J.F. Uptake, elimination and tissue distribution of dietary and aqueous cadmium by rainbow trout (Salmo gairdneri richardson) and lake whitefish (Coregonus clupeaformis mitchill). Environ. Toxicol. Chem. 1989, 8, 87–97. [Google Scholar] [CrossRef]
- Rehwoldt, R.; Karimian-Teherani, D. Uptake and Effect of Cadmium on Zebrafish. Bull. Environ. Contam. Toxicol. 1976, 15, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Illuminati, S.; Truzzi, C.; Annibaldi, A.; Migliarini, B.; Carnevali, O.; Scarponi, G. Cadmium Bioaccumulation and Metallothionein Induction in the Liver of the Antarctic Teleost Trematomus bernacchii during an On-Site Short-Term Exposure to the Metal via Seawater. Toxicol. Environ. Chem. 2010, 92, 617–640. [Google Scholar] [CrossRef]
- Adhikary, J.; Maity, A.; Kumar Das, B.; Ghosh, S.; Jayati Adhikary, C.; Pal, P. Accumulation of Cadmium (Cd) and Lead (Pb) in Tissues of Rohu Fish (Labeo rohita) Collected from the Sewage-Fed Pond of Kolkata. J. Entomol. Zool. Stud. 2019, 7, 146–150. [Google Scholar]
- Hu, S.; Han, J.; Yang, L.; Li, S.; Guo, Y.; Zhou, B.; Wu, H. Bioconcentration, Depuration and Toxicity of Pb in the Presence of Titanium Dioxide Nanoparticles in Zebrafish Larvae. Aquat. Toxicol. 2019, 214, 105257. [Google Scholar] [CrossRef]
- International Lead Association (ILA). Lead in Aquatic Environments. 2019. Available online: https://www.ila-lead.org/userfiles/file/aquatic_environment_report_v6_hi%20res.pdf (accessed on 20 September 2021).
- Nøstbakken, O.J.; Hove, H.T.; Duinker, A.; Lundebye, A.K.; Berntssen, M.H.G.; Hannisdal, R.; Lunestad, B.T.; Maage, A.; Madsen, L.; Torstensen, B.E.; et al. Contaminant Levels in Norwegian Farmed Atlantic Salmon (Salmo salar) in the 13-Year Period from 1999 to 2011. Environ. Int. 2015, 74, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Hannisdal, R.; Nøstbakken, O.-J.; Hove, H.T.; Lunestad, B.T.; Frøyland, L.; Madsen, L. Monitoring Program for Pharmaceuticals, Illegal Substances, and Contaminants in Farmed Fish. 2013. Available online: https://nifes.hi.no/wp-content/uploads/2013/01/9623rapportfor2012130801.pdf (accessed on 20 October 2021).
- Nandi, S.; Srivastava, R.C.; Agarwal, K.M. Lead and Cadmium Accumulation in Fresh Water Fishes Labeo rohita and Catla catla. J. Environ. Res. Dev. 2012, 6, 748–752. [Google Scholar]
- Gonzalez, P.; Dominique, Y.; Massabuau, J.C.; Boudou, A.; Bourdineaud, J.P. Comparative Effects of Dietary Methylmercury on Gene Expression in Liver, Skeletal Muscle, and Brain of the Zebrafish (Danio rerio). Environ. Sci. Technol. 2005, 39, 3972–3980. [Google Scholar] [CrossRef]
- Kelly, B.C.; Ikonomou, M.G.; Higgs, D.A.; Oakes, J.; Dubetz, C.; Canada, O. Mercury and other trace elements in farmed and wild salmon from British Columbia, Canada. Environ. Toxicol. Chem. 2008, 27, 1361–1370. [Google Scholar] [CrossRef]
- Wiener, J.G.; Krabbenhoft, D.P.; Heinz, G.H.; Scheuhammer, M. Ecotoxicology of Mercury. In Handbook of Ecotoxicology; CRC Press: Boca Raton, FL, USA, 2003; pp. 409–443. [Google Scholar]
- Korbas, M.; MacDonald, T.C.; Pickering, I.J.; George, G.N.; Krone, P.H. Chemical Form Matters: Differential Accumulation of Mercury Following Inorganic and Organic Mercury Exposures in Zebrafish Larvae. ACS Chem. Biol. 2012, 7, 411–420. [Google Scholar] [CrossRef]
- Gentès, S.; Maury-Brachet, R.; Feng, C.; Pedrero, Z.; Tessier, E.; Legeay, A.; Mesmer-Dudons, N.; Baudrimont, M.; Maurice, L.; Amouroux, D.; et al. Specific Effects of Dietary Methylmercury and Inorganic Mercury in Zebrafish (Danio rerio) Determined by Genetic, Histological, and Metallothionein Responses. Environ. Sci. Technol. 2015, 49, 14560–14569. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) Physiological and Behavioural Responses to Insect-Based Diets: A Multidisciplinary Approach. Sci. Rep. 2020, 10, 10648. [Google Scholar] [CrossRef]
- Balshaw, S.; Edwards, J.W.; Ross, K.E.; Daughtry, B.J. Mercury Distribution in the Muscular Tissue of Farmed Southern Bluefin Tuna (Thunnus maccoyii) Is Inversely Related to the Lipid Content of Tissues. Food Chem. 2008, 111, 616–621. [Google Scholar] [CrossRef]
- Meador, J.P. Comparative Toxicokinetics of Tributyltin in Five Marine Species and Its Utility in Predicting Bioaccumulation and Acute Toxicity. Aquat. Toxicol. 1997, 37, 307–326. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, L.; Wang, W.X. Arsenic Bioaccumulation in a Marine Juvenile Fish Terapon jarbua. Aquat. Toxicol. 2011, 105, 582–588. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, L.; Wang, W.X. Biotransformation and Detoxification of Inorganic Arsenic in a Marine Juvenile Fish Terapon jarbua after Waterborne and Dietborne Exposure. J. Hazard. Mater. 2012, 221–222, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, J.S.; Shibata, Y.; Francesconi, K.A.; Rippingale, R.J.; Morita, M. Arsenic Transformations in Short Marine Food Chains Studied by HPLC-ICP MS. Appl. Organometal. Chem 1997, 11, 281–287. [Google Scholar] [CrossRef]
- Azizur Rahman, M.; Hasegawa, H.; Peter Lim, R. Bioaccumulation, Biotransformation and Trophic Transfer of Arsenic in the Aquatic Food Chain. Environ. Res. 2012, 116, 118–135. [Google Scholar] [CrossRef] [Green Version]
- Jankong, P.; Chalhoub, C.; Kienzl, N.; Goessler, W.; Francesconi, K.A.; Visoottiviseth, P. Arsenic Accumulation and Speciation in Freshwater Fish Living in Arsenic-Contaminated Waters. Environ. Chem. 2007, 4, 11–17. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The Arsenic Accumulation and Its Effect on Oxidative Stress Responses in Juvenile Rockfish, Sebastes schlegelii, Exposed to Waterborne Arsenic (As3+). Environ. Toxicol. Pharmacol. 2015, 39, 668–676. [Google Scholar] [CrossRef]
- Pei, J.; Zuo, J.; Wang, X.; Yin, J.; Liu, L.; Fan, W. The Bioaccumulation and Tissue Distribution of Arsenic Species in Tilapia. Int. J. Environ. Res. Public Health 2019, 16, 757. [Google Scholar] [CrossRef] [Green Version]
- Bordajandi, L.R.; Gómez, G.; Fernández, M.A.; Abad, E.; Rivera, J.; González, M.J. Study on PCBs, PCDD/Fs, Organochlorine Pesticides, Heavy Metals and Arsenic Content in Freshwater Fish Species from the River Turia (Spain). Chemosphere 2003, 53, 163–171. [Google Scholar] [CrossRef]
- Komjarova, I.; Blust, R. Multimetal Interactions between Cd, Cu, Ni, Pb, and Zn Uptake from Water in the Zebrafish Danio rerio. Environ. Sci. Technol. 2009, 43, 7225–7229. [Google Scholar] [CrossRef]
- Ptashynski, M.D.; Klaverkamp, J.F. Accumulation and Distribution of Dietary Nickel in Lake Whitefish (Coregonus clupeaformis). Aquat. Toxicol. 2002, 58, 249–264. [Google Scholar] [CrossRef]
- Lawrence, C.; Adatto, I.; Best, J.; James, A.; Maloney, K. Generation Time of Zebrafish (Danio rerio) and Medakas (Oryzias latipes) Housed in the Same Aquaculture Facility. Lab. Anim. 2012, 41, 158–165. [Google Scholar] [CrossRef]
- Illuminati, S.; Annibaldi, A.; Truzzi, C.; Scarponi, G. Recent Temporal Variations of Trace Metal Content in an Italian White Wine. Food Chem. 2014, 159, 493–497. [Google Scholar] [CrossRef]
- Morgano, M.A.; Milani, R.F.; Perrone, A.A.M. Determination of Total Mercury in Sushi Samples Employing Direct Mercury Analyzer. Food Anal. Methods 2015, 8, 2301–2307. [Google Scholar] [CrossRef]
- Daniel, W.W.; Cross, C.L. Biostatistics: A Foundation for Analysis in the Health Sciences, 10th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- STATGRAPHICS Centurion 19 Software, Manugistics Inc.: Rockville, MD, USA, 2019.
| Element | Determined Value (µg L−1) | Legal Limit (µg L−1) |
|---|---|---|
| Hg | <0.0006 | 1 |
| Pb | <0.030 | 25 |
| Cd | 0.008 ± 0.001 | 5 |
| As | 0.51 ± 0.04 | 10 |
| Ni | 3.3 ± 0.5 | 20 |
| Sample * | Cd | Pb | Hg | As | Ni |
|---|---|---|---|---|---|
| LCTRL | 0.10 ± 0.01 | 0.23 ± 0.2 | 0.20 ± 0.01 | 2.0 ± 0.2 | 0.49 ± 0.04 |
| LHi25 | 0.16 ± 0.01 | 0.34 ± 0.02 | 0.31 ± 0.02 | 2.8 ± 0.4 | 0.9 ± 0.1 |
| LHi50 | 0.09 ± 0.01 | 0.55 ± 0.03 | 0.24 ± 0.01 | 2.9 ± 0.2 | 2.3 ± 0.3 |
| LHi75 | 0.08 ± 0.01 | 0.87 ± 0.04 | 0.20 ± 0.01 | 3.4 ± 0.1 | 3.1 ± 0.5 |
| LHi100 | 0.11 ± 0.01 | 1.9 ± 0.1 | 0.15 ± 0.01 | 3.2 ± 0.4 | 27 ± 3 |
| JCTRL | 0.7 ± 0.1 | 0.32 ± 0.02 | 2.2 ± 0.2 | 2.8 ± 0.3 | 2.2 ± 0.1 |
| JHi25 | 1.3 ± 0.1 | 0.47 ± 0.03 | 2.0 ± 0.1 | 2.4 ± 0.3 | 1.9 ± 0.2 |
| JHi50 | 0.7 ± 0.1 | 0.84 ± 0.05 | 1.8 ± 0.2 | 1.8 ± 0.2 | 5.2 ± 0.8 |
| JHi75 | 1.3 ± 0.1 | 0.98 ± 0.07 | 2.1 ± 0.3 | 0.8 ± 0.1 | 10 ± 2 |
| JHi100 | 1.5 ± 0.1 | 3.4 ± 0.1 | 2.1 ± 0.2 | 0.9 ± 0.2 | 39 ± 3 |
| ACTRL | 0.8 ± 0.1 | 0.32 ± 0.05 | 2.8 ± 0.2 | 3.7 ± 0.6 | 3.5 ± 0.6 |
| AHi25 | 1.3 ± 0.2 | 0.65 ± 0.14 | 2.7 ± 0.2 | 2.8 ± 0.6 | 2.3 ± 0.4 |
| AHi50 | 0.8 ± 0.1 | 0.91 ± 0.14 | 2.4 ± 0.2 | 2.7 ± 0.6 | 7.4 ± 2.0 |
| AHi75 | 1.3 ± 0.1 | 1.2 ± 0.1 | 2.7 ± 0.2 | 1.4 ± 0.3 | 11 ± 2 |
| AHi100 | 1.5 ± 0.2 | 3.7 ± 0.4 | 2.6 ± 0.3 | 1.4 ± 0.2 | 41 ± 6 |
| Element | Analytical Method | Analytical Result (n = 9) | Certified Value | Δ (%) |
|---|---|---|---|---|
| Cd | GF-AAS | 0.041 ± 0.005 | 0.043 ± 0.008 | −4.7 |
| Pb | GF-AAS | 0.067 ± 0.003 | 0.065 ± 0.007 | +3.1 |
| As | GF-AAS | 17.6 ± 0.5 | 18 ± 1.1 | −2.2 |
| Ni | GF-AAS | 18.8 ± 0.9 | 19.4 ± 3.1 | −3.1 |
| Hg | DMA-1 | 4.30 ± 0.2 | 4.64 ± 0.2 | −6.1 |
| Instrumental Parameter | Hg | Pb | Cd | As |
|---|---|---|---|---|
| Lamp current (mA) | 30/0 | 80/40 | 60/30 | 60/30 |
| PMT (Volts) | 275 | 285 | 280 | 320 |
| Carrier gas (mL min−1) | 300 | 300 | 500 | 300 |
| Shield gas (mL min−1) | 1000 | 800 | 800 | 800 |
| Reading time (s) | 15 | 15 | 15 | 15 |
| Delay time (s) | 0.5 | 0.5 | 0.5 | 0.5 |
| Blank judgement Value (if) | 5 | 10 | 10 | 10 |
| Torch height (mm) | 10 | 8 | 8 | 8 |
| IFS Step (s × rpm, a = analyte; c = carrier; r = reading) | (a) 10 × 100 (c) (r) 16 × 120 | (a) 10 × 100 (c) (r) 16 × 120 | (a) 10 × 100 (c) (r) 16 × 120 | (a) 10 × 100 (c) (r) 18 × 120 |
| Element | Instrument | LOD (µg·L−1) | LOQ (µg·L−1) | CRM | ||
|---|---|---|---|---|---|---|
| Name | Certified Value (µg L−1) | Measured Value (µg L−1) | ||||
| Hg | AFS | 0.0006 | 0.006 | DORM-2 | 4.64 ± 0.26 a | 4.84 ± 0.22 |
| Pb | AFS | 0.03 | 3 | DORM-2 | 0.065 ± 0.007 a | 0.060 ± 0.008 |
| Cd | AFS | 0.0005 | 0.005 | NASS-6 | 0.0311 ± 0.0019 | 0.031 ± 0.008 |
| As | AFS | 0.01 | 0.13 | SLEW-3 | 1.36 ± 0.09 | 1.47 ± 0.27 |
| Ni | GFAAS | 0.144 | 1.440 | SLEW-3 | 1.23 ± 0.07 | 1220 ± 390 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truzzi, C.; Girolametti, F.; Giovannini, L.; Olivotto, I.; Zarantoniello, M.; Scarponi, G.; Annibaldi, A.; Illuminati, S. New Eco-Sustainable Feed in Aquaculture: Influence of Insect-Based Diets on the Content of Potentially Toxic Elements in the Experimental Model Zebrafish (Danio rerio). Molecules 2022, 27, 818. https://doi.org/10.3390/molecules27030818
Truzzi C, Girolametti F, Giovannini L, Olivotto I, Zarantoniello M, Scarponi G, Annibaldi A, Illuminati S. New Eco-Sustainable Feed in Aquaculture: Influence of Insect-Based Diets on the Content of Potentially Toxic Elements in the Experimental Model Zebrafish (Danio rerio). Molecules. 2022; 27(3):818. https://doi.org/10.3390/molecules27030818
Chicago/Turabian StyleTruzzi, Cristina, Federico Girolametti, Leonardo Giovannini, Ike Olivotto, Matteo Zarantoniello, Giuseppe Scarponi, Anna Annibaldi, and Silvia Illuminati. 2022. "New Eco-Sustainable Feed in Aquaculture: Influence of Insect-Based Diets on the Content of Potentially Toxic Elements in the Experimental Model Zebrafish (Danio rerio)" Molecules 27, no. 3: 818. https://doi.org/10.3390/molecules27030818