
Can Artemisia herba-alba Be Useful for Managing COVID-19 and Comorbidities?


 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Methods
3. Results and Discussion
3.1. Ethnobotanical Aspects of Artemisia herba-alba
3.2. Pharmacological Aspects of Artemisia herba-alba
3.3. Molecular Docking Studies of Artemisia herba-alba Phytochemicals with SARS-CoV-2 Mpro
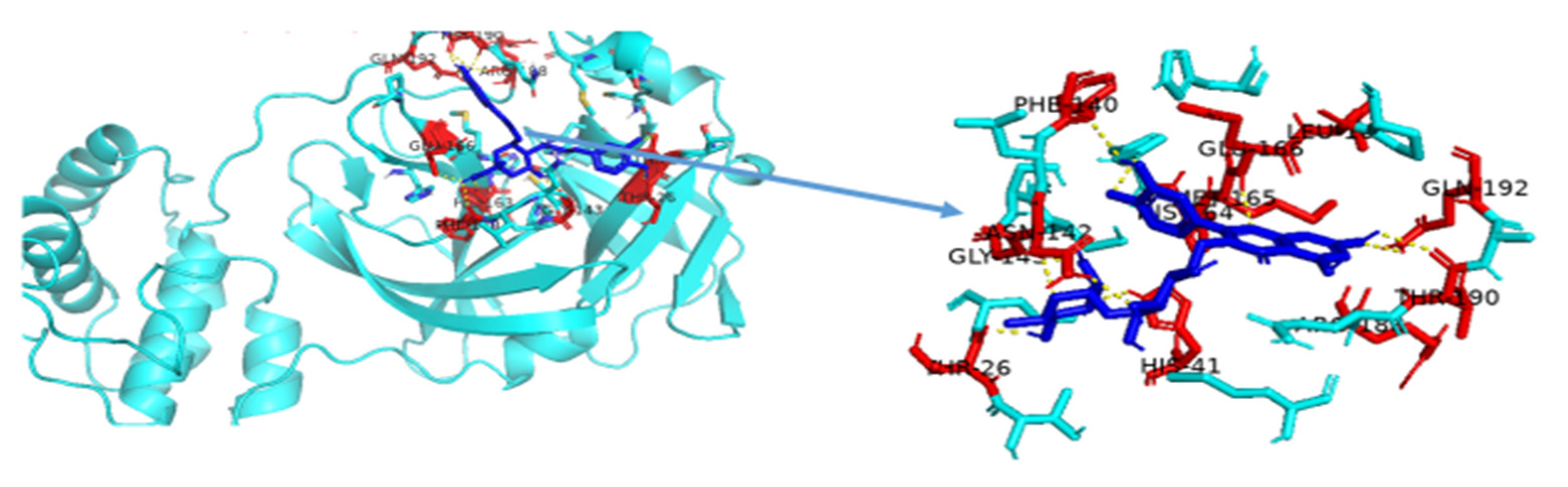
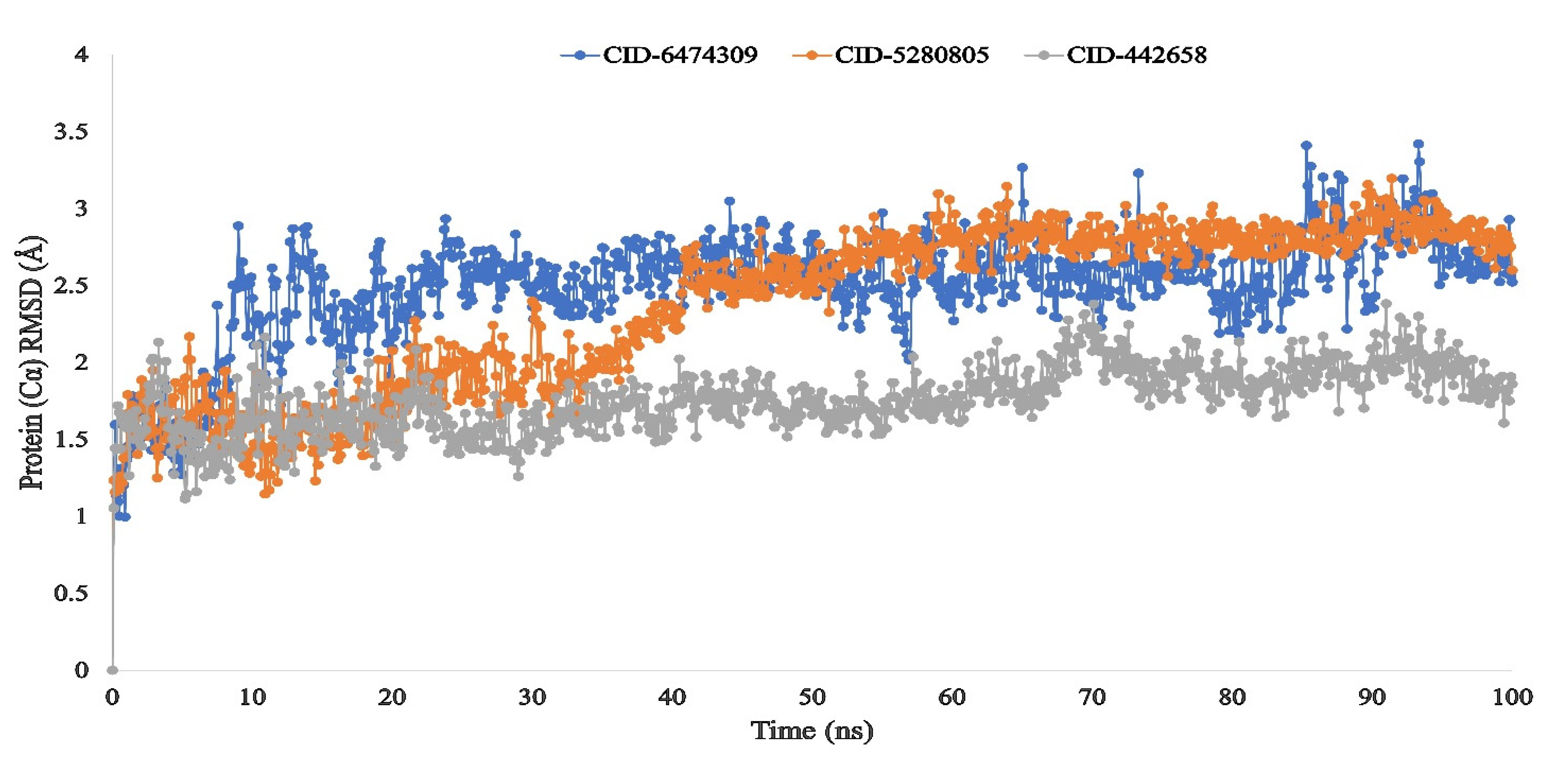
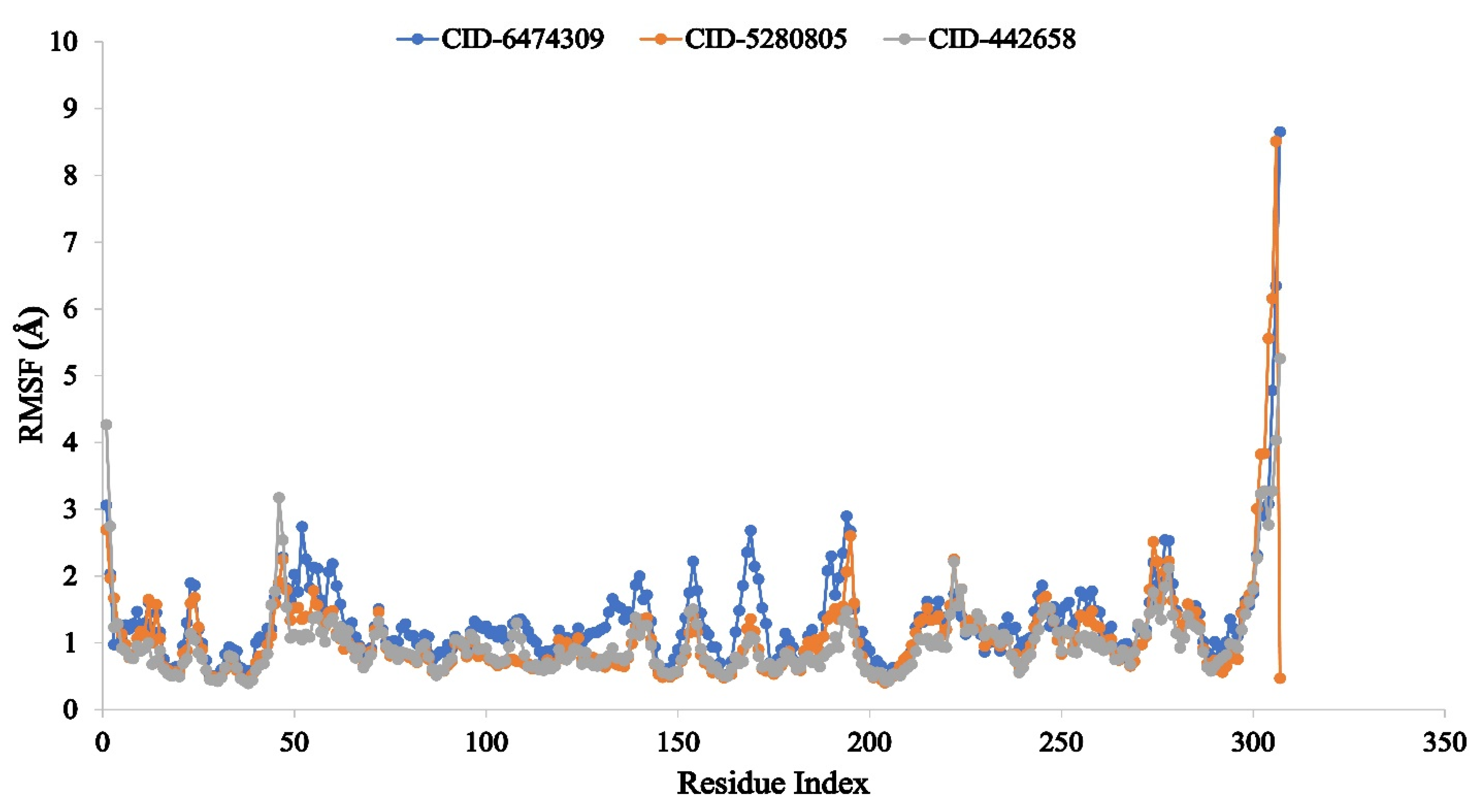
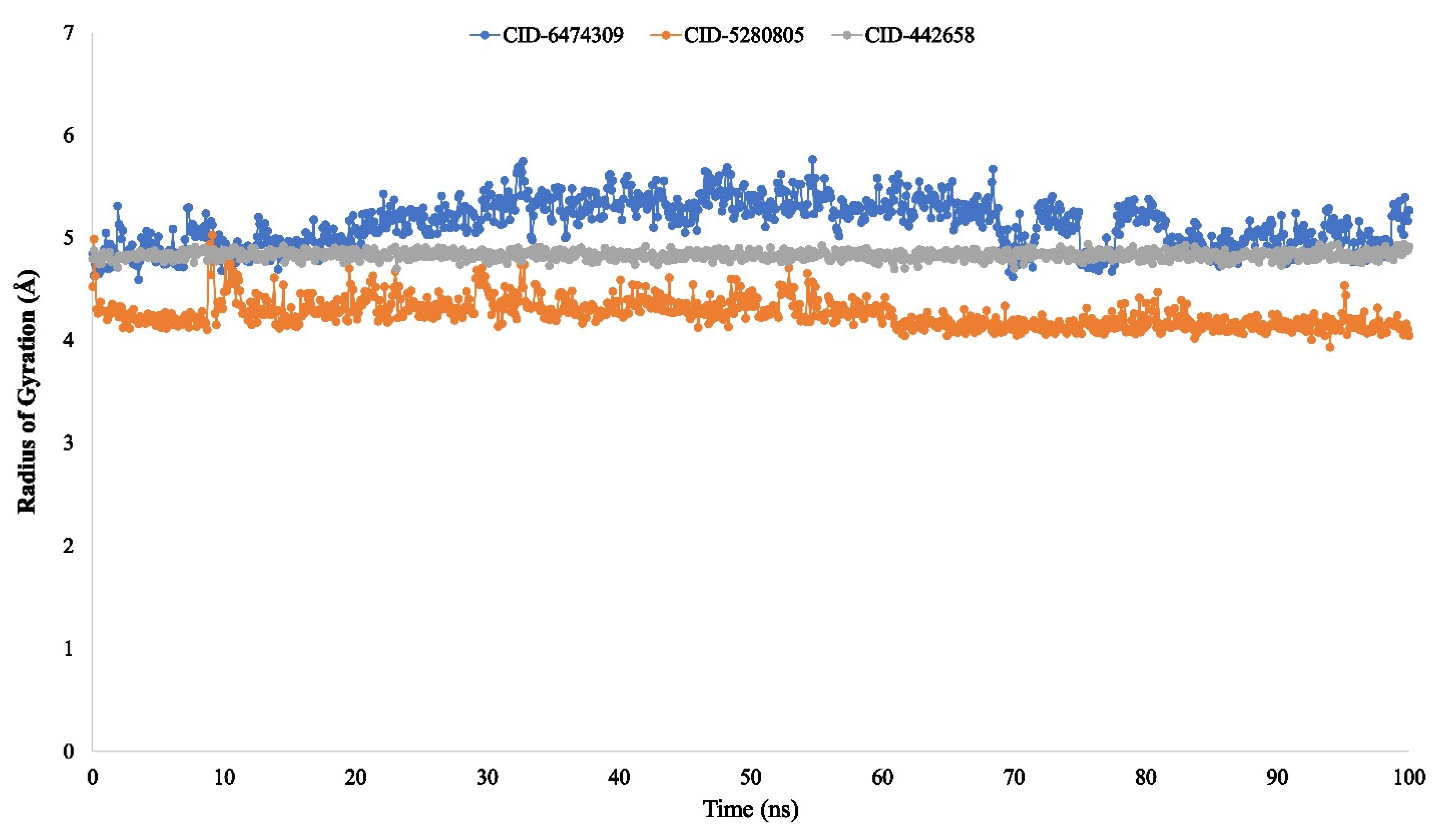
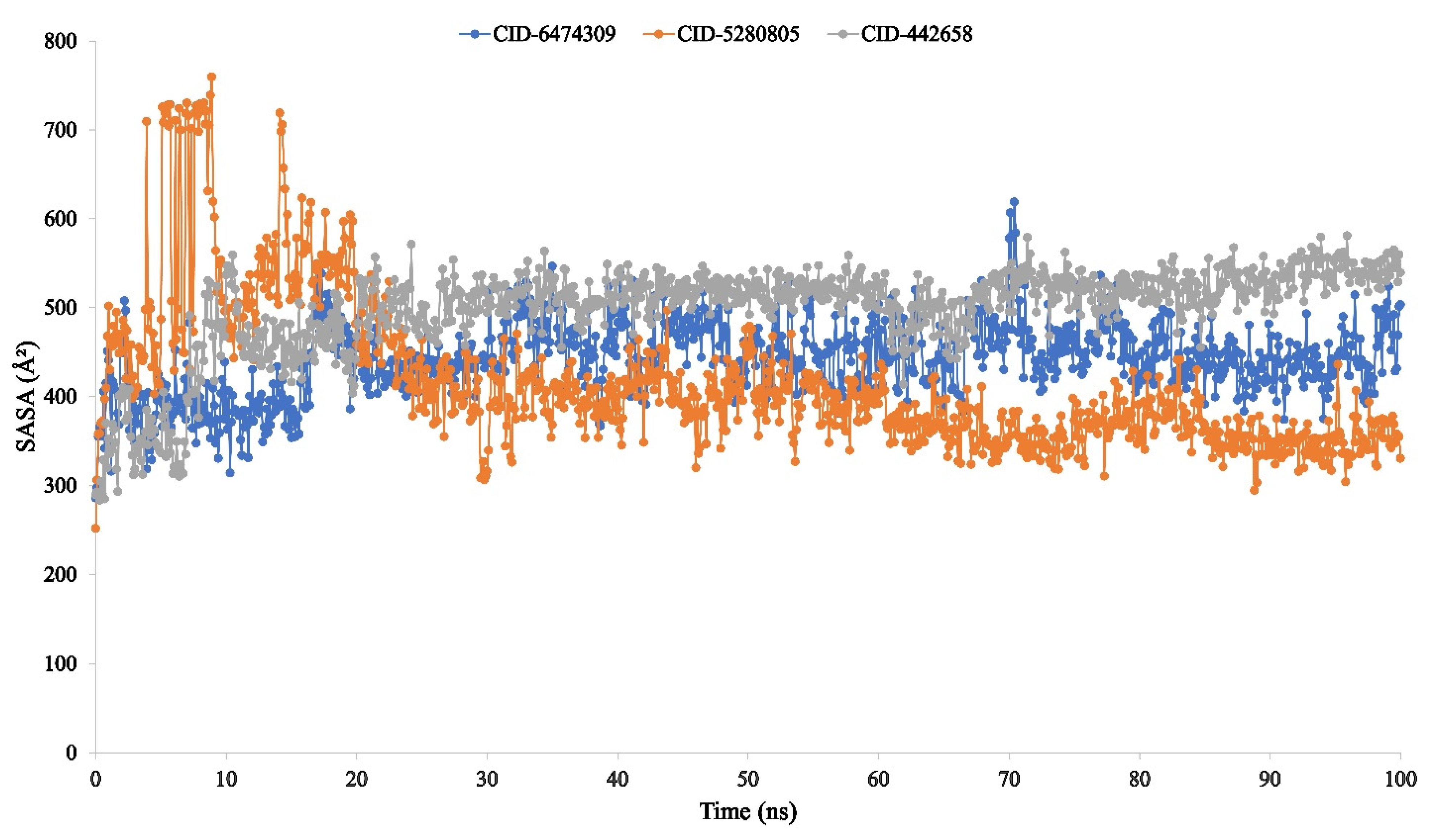
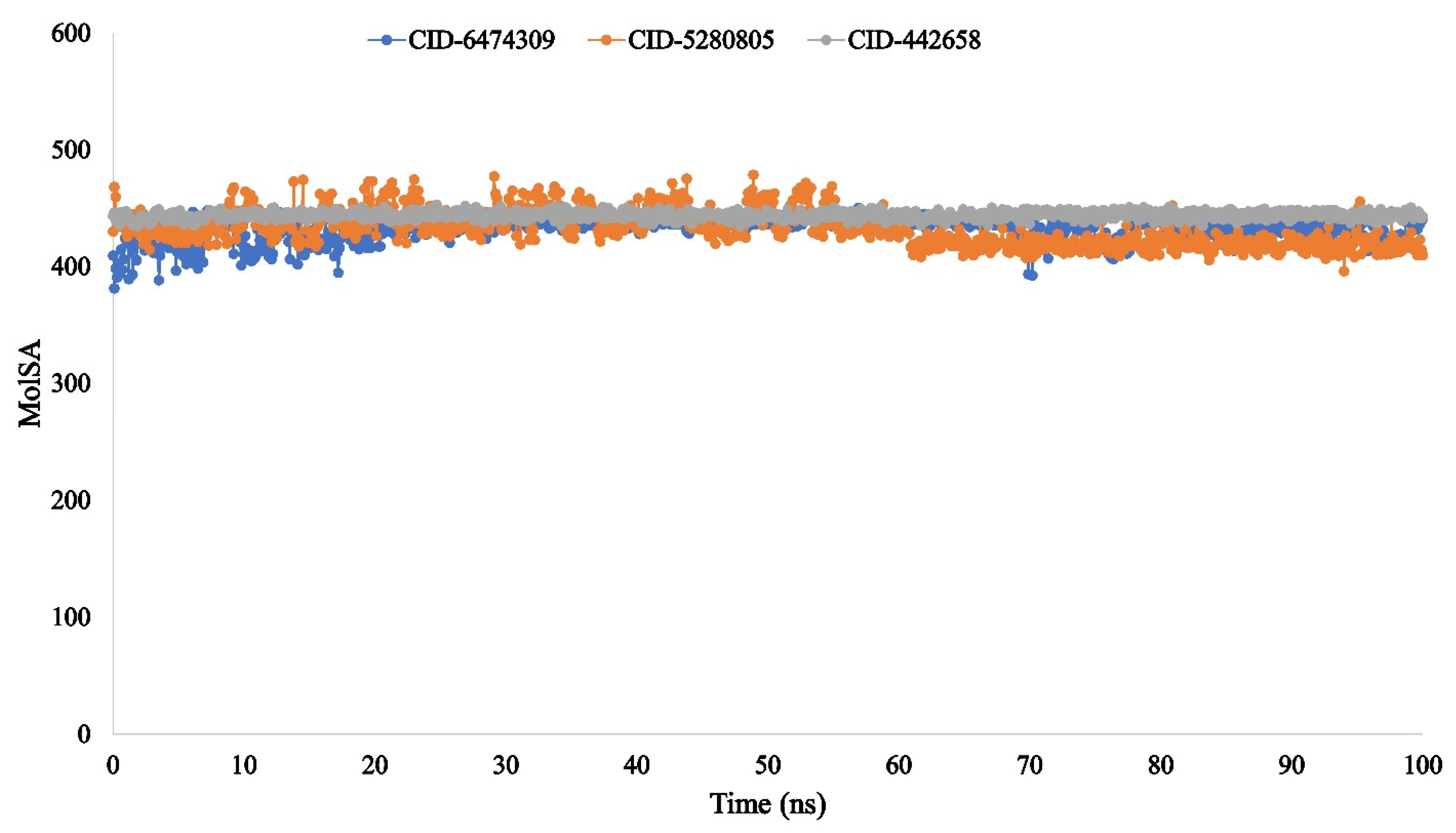
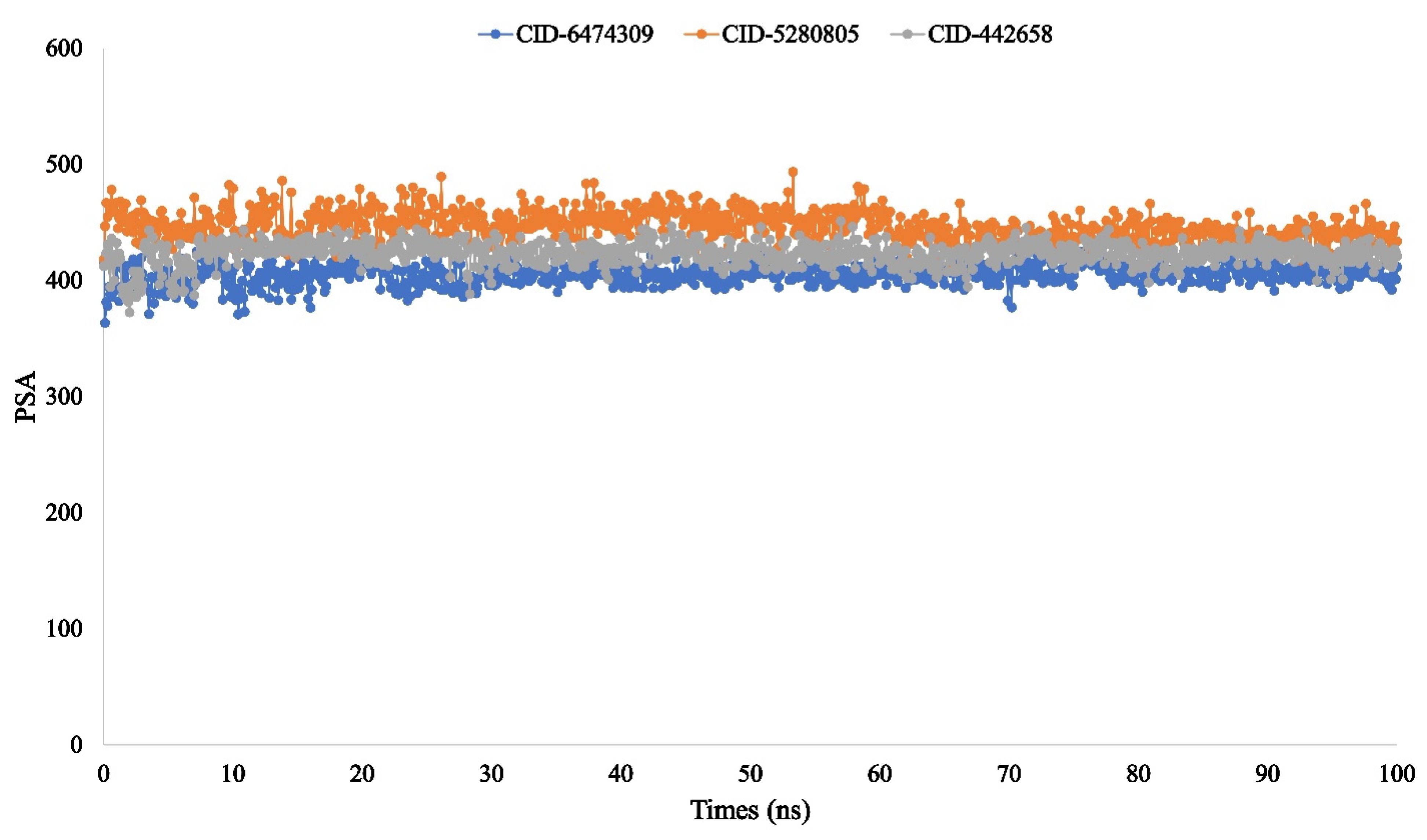
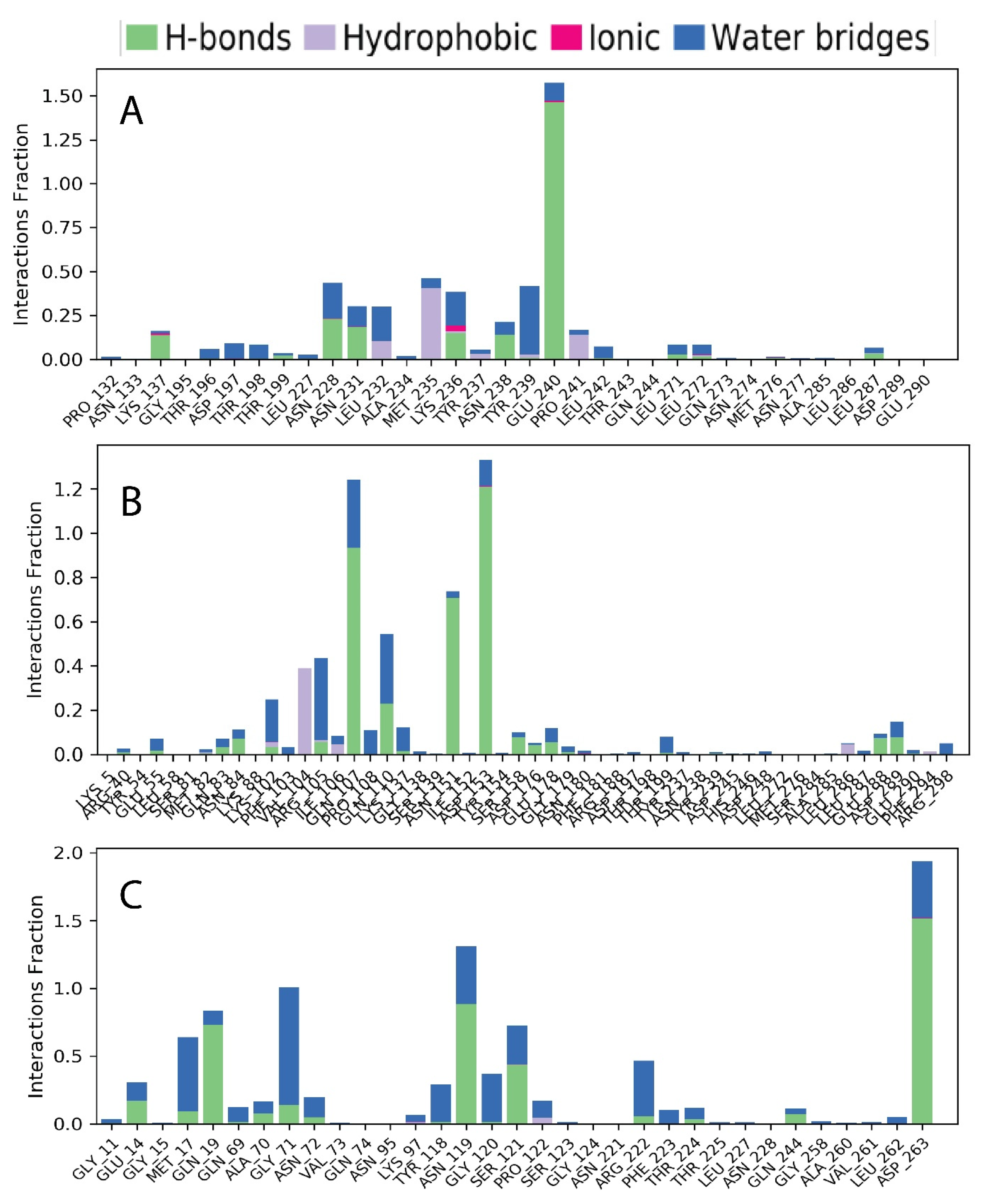
3.4. Molecular Dynamic Simulation of Artemisia herba-alba Phytochemicals with High Affinity Binding with SARS-CoV-2 Mpro
3.5. Reported Effect(s) of High Affinity Mpro Binding Phytochemicals of Artemisia herba-alba on Selected Comorbidities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Shanmugaraj, B.; Malla, A.; Phoolcharoen, W. Emergence of novel Coronavirus 2019-nCoV: Need for rapid vaccine and biologics development. Pathogens 2020, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayanand, P.; Wilkins, E.; Woodhead, M. Severe acute respiratory syndrome (SARS): A review. Clin. Med. 2004, 4, 152–160. [Google Scholar] [CrossRef]
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019; Comprehensive Tables (ST/ESA/SER.A/426); United Nations: New York, NY, USA, 2019; Volume 1. [Google Scholar]
- Aziz, A.B.; Raqib, R.; Khan, W.A.; Rahman, M.; Haque, R.; Alam, M.; Zaman, K.; Ross, A.G. Integrated control of COVID-19 in resource-poor countries. Int. J. Infect. Dis. 2020, 101, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Haidere, M.F.; Ratan, Z.A.; Nowroz, S.; Bin Zaman, S.; Jung, Y.-J.; Hosseinzadeh, H.; Cho, J.Y. COVID-19 Vaccine: Critical questions with complicated answers. Biomol. Ther. 2021, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Patidar, R.; Younis, K.; Desai, P.; Hosein, Z.; Padda, I.; Mangat, J.; Altaf, M. Comorbidity and its impact on patients with COVID-19. SN Compr. Clin. Med. 2020, 2, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Bajgain, K.T.; Badal, S.; Bajgain, B.B.; Santana, M.J. Prevalence of comorbidities among individuals with COVID-19: A rapid review of current literature. Am. J. Infect. Control 2021, 49, 238–246. [Google Scholar] [CrossRef]
- WHO. World Malaria Report; World Health Organization (WHO): Geneva, Switzerland, 2019; Available online: https://www.who.int/publications/i/item/9789241565721 (accessed on 20 June 2021).
- Di Gennaro, F.; Marotta, C.; Locantore, P.; Pizzol, D.; Putoto, G. Malaria and COVID-19: Common and different findings. Trop. Med. Infect. Dis. 2020, 5, 141. [Google Scholar] [CrossRef]
- Torjesen, I. COVID-19: Omicron may be more transmissible than other variants and partly resistant to existing vaccines, scientists fear. BMJ 2021, 375, n2943. [Google Scholar] [CrossRef]
- Yamey, G. Rich countries should tithe their vaccines. Nature 2021, 590, 529. [Google Scholar] [CrossRef]
- Callaway, E.; Ledford, H. How bad is Omicron? What scientists know so far. Nature 2021, 600, 197–199. [Google Scholar] [CrossRef]
- Luo, H.; Yang, M.; Tang, Q.L.; Hu, X.Y.; Willcox, M.L.; Liu, J.P. Characteristics of registered clinical trials on traditional Chinese medicine for coronavirus disease 2019 (COVID-19): A scoping review. Eur. J. Integr. Med. 2021, 41, 101251. [Google Scholar] [CrossRef]
- Wadaa-Allah, A.; Emhamed, M.S.; Sadeq, M.A.; Dahman, N.B.H.; Ullah, I.; Farrag, N.S.; Negida, A. Efficacy of the current investigational drugs for the treatment of COVID-19: A scoping review. Ann. Med. 2021, 53, 318–334. [Google Scholar] [CrossRef]
- World Health Organization. Traditional Herbal Remedies for Primary Health Care; World Health Organization: Geneva, Switzerland, 2010; ISBN 978-92-9022-382-5. Available online: https://apps.who.int/iris/handle/10665/206024 (accessed on 16 November 2021).
- Udaykumar, P. Discovery of artemisinin: The Chinese wonder drug. Muller J. Med. Sci. Res. 2014, 5, 191. [Google Scholar] [CrossRef]
- Roy, P. Global pharma and local science: The untold tale of reserpine. Indian J. Psychiatry 2018, 60, S277–S283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-H.; Wu, K.-L.; Zhang, X.; Deng, S.-Q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Jahan, R.; Jannat, K.; Bondhon, T.A.; Hossan, S.; Mazumder, S.; Fatima, A.; Pereira, M.D.L.; Wiart, C.; Nissapatorn, V.; et al. Evaluation of Lens culinaris phytochemicals in binding to the 3C-like protease of SARS-CoV-2—A molecular docking approach. Indian J. Med. Sci. 2020, 72, 173–176. [Google Scholar] [CrossRef]
- Touafchia, A.; Bagheri, H.; Carrié, D.; Durrieu, G.; Sommet, A.; Chouchana, L.; Montastruc, F. Serious bradycardia and remdesivir for coronavirus 2019 (COVID-19): A new safety concerns. Clin. Microbiol. Infect. 2021, 27, 791.e5–791.e8. [Google Scholar] [CrossRef]
- Odeyemi, S.; Bradley, G. Medicinal plants used for the Traditional management of diabetes in the Eastern Cape, South Africa: Pharmacology and Toxicology. Molecules 2018, 23, 2759. [Google Scholar] [CrossRef] [Green Version]
- Mahomoodally, M.F. Traditional medicines in Africa: An Appraial of ten potent African Medicinal plants. Evid.-Based Complement. Altern. Med. 2013, 2013, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Revannavar, S.M.; Supriya, S.P.; Samaga, L.; Vineeth, V.K. COVID-19 triggering mucormycosis in a susceptible patient: A new phenomenon in the developing world? BMJ Case Rep. 2021, 14, e241663. [Google Scholar] [CrossRef]
- Waizel-Haiat, S.; Guerrero-Paz, J.A.; Sanchez-Hurtado, L.; Calleja-Alarcon, S.; Romero-Gutierrez, L. A case of fatal rhino-orbital mucormycosis associated with new Onsot diabetic ketoacidosis and COVID-19. Cureus 2021, 13, e13163. [Google Scholar] [CrossRef] [PubMed]
- Sipsas, N.V.; Gamaletsou, M.N.; Anastasopoulou, A.; Kontoyiannis, D.P. Therapy of Mucormycosis. J. Fungi 2018, 4, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewal, A.; Kataria, H.; Dhawan, I. Literature search for research planning and identification of research problem. Indian J. Anaesth. 2016, 60, 635–639. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, B.; Jin, Z.; Yang, H.; Rao, Z. The crystal structure of COVID-19 main protease in complex with an inhibitor N3. Protein Data Bank 2020. Available online: https://www.rcsb.org/structure/6LU7 (accessed on 16 November 2021). [CrossRef]
- Souhila, T.; Zohra, B.F.; Tahar, H.S. Identification and quantification of phenolic compounds of Artemisia herba-alba at three harvest time by HPLC-ESI-Q-TOF-MS. Int. J. Food Prop. 2019, 22, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Cheraif, K.; Bakchiche, B.; Gherib, A.; Bardaweel, S.K.; Ayvaz, M.; Flamini, G.; Ascrizzi, R.; Ghareeb, M.A. Chemical composition, antioxidant, anti-Tyrosinase, anti-Cholinesterase and cytotoxic activities of essential oils of six Algerian plants. Molecules 2020, 25, 1710. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.E.H.H.; El-Sayed, M.A.; Hegazy, M.E.; Helaly, S.E.; Esmail, A.M.; Mohamed, N.S. Chemical constituents and biological activities of Artemisia-herba-alba. Rec. Nat. Prod. 2010, 4, 1–25. [Google Scholar]
- Ihlenfeldt, W.D. PubChem. In Applied Chemoinformatics: Achievements and Future Opportunities; Wiley: Hoboken, NJ, USA, 2018; pp. 245–258. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Studio Design. Dassault Systemes BIOVIA, Discovery Studio Modelling Environment, Release 4.5; Accelrys Software Inc.: San Diego, CA, USA, 2015; pp. 98–104. Available online: https://www.addlink.es›whats-new-in-discovery-studio (accessed on 11 November 2021).
- Hoffman, J.M.; Margolis, K.G. Building community in the gut: A role for mucosal serotonin. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 6–8. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Dubey, A.; Yadava, U.; Mishra, S.K.; Kang, S.G.; Dwivedi, V.D. Exploration of natural compounds with anti-SARS-CoV-2 activity via inhibition of SARS-CoV-2 Mpro. Brief. Bioinform. 2021, 22, 1361–1377. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Z.; Ren, J.; Wei, Z.; Li, J. Potential inhibitors for the novel coronavirus (SARS-CoV-2). Brief. Bioinform. 2020, 22, 1225–1231. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Zumla, A.; Petersen, E.; Nyirenda, T.; Chakaya, J. Tackling the tuberculosis epidemic in sub-Saharan Africa—Unique opportunities arising from the second European developing countries clinical trials partnership (EDCTP) programme 2015–2024. Int. J. Infect. Dis. 2015, 32, 46–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Maggar, E.M.B. Artemisia herba alba & Artemisia monosperma: The Discovery of the first potential Egyptian plant sources for the Pharmaceutical Commercial Production of Artemisinin and Some of Its Related Analogues. J. Appl. Pharm. Sci. 2012. [Google Scholar] [CrossRef] [Green Version]
- Ziyyat, A.; Legssyer, A.; Mekhfi, H.; Dassouli, A.; Serhrouchni, M.; Benjelloun, W. Phytotherapy of hypertension and diabetes in oriental Morocco. J. Ethnopharmacol. 1997, 58, 45–54. [Google Scholar] [CrossRef]
- Tahraoui, A.; EL Hilaly, J.; Israili, Z.H.; Lyoussi, B. Ethnopharmacological survey of plants used in the traditional treatment of hypertension and diabetes in south-eastern Morocco (Errachidia province). J. Ethnopharmacol. 2007, 110, 105–117. [Google Scholar] [CrossRef]
- Friedman, J.; Yaniv, Z.; Dafni, A.; Palewitch, D. A preliminary classification of the healing potential of medicinal plants, based on a rational analysis of an ethnopharmacological field survey among Bedouins in the Negev Desert, Israel. J. Ethnopharmacol. 1986, 16, 275–287. [Google Scholar] [CrossRef]
- Hamamouch, N. Use of Ethnomedicinal Plants by the People Living in the Middle Atlas Mountains in Morocco. Med. Aromat. Plants 2020, 9, 349. [Google Scholar] [CrossRef]
- Sarri, M.; Mouyet, F.Z.; Benziane, M.; Cheriet, A. Traditional use of medicinal plants in a city at steppic character (M’sila, Algeria). J. Pharm. Pharmacogn. Res. 2014, 2, 31–35. [Google Scholar]
- Amel, B. Traditional treatment of high blood pressure and diabetes in Souk Ahras District. J. Pharmacogn. Phytother. 2013, 5, 12–20. [Google Scholar] [CrossRef]
- Darwish, R.M.; Aburjai, T.A. Effect of ethnomedicinal plants used in folklore medicine in Jordan as antibiotic resistant in-hibitors on Escherichia coli. BMC Complement. Alternat. Med. 2010, 10, 9. Available online: http://www.biomedcentral.com/1472-6882/10/9 (accessed on 16 November 2021). [CrossRef] [Green Version]
- Najem, M.; Harouak, H.; Ibijbijen, J.; Nassiri, L. Oral disorders and ethnobotanical treatments: A field study in the central Middle Atlas (Morocco). Heliyon 2020, 6, e04707. [Google Scholar] [CrossRef] [PubMed]
- Benarba, B. Medicinal plants used by traditional healers from South-west Algeria: An ethnobotanical study. J. Intercult. Ethnopharmacol. 2016, 5, 320–330. [Google Scholar] [CrossRef]
- Zeb, S.; Ali, A.; Zaman, W.; Zeb, S.; Ali, S.; Ullah, F.; Shakoor, A. Pharmacology, taxonomy and phytochemistry of the genus Artemisia specifically from Pakistan: A comprehensive review. Pharm. Biomed. Res. 2018, 4, 1–12. [Google Scholar] [CrossRef]
- Žarković, N.; Orehovec, B.; Milković, L.; Baršić, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataić, A.; Jakovčević, A.; et al. Preliminary findings on the association of the lipid peroxidation product 4-hydroxynonenal with the lethal outcome of aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef] [PubMed]
- Moufid, A.; Eddouks, M. Artemisia herba alba: A popular plant with potential medicinal properties. Pak. J. Biol. Sci. 2012, 15, 1152–1159. [Google Scholar] [CrossRef]
- Allam, N.G.; Hefnawy, M.A.; Aldamanhoury, M.A. Antibacterial activity of six Egyptian medicinal plants against some pathogenic bacteria. Egypt. J. Exp. Biol. 2015, 11, 71–75. [Google Scholar]
- Al-Waili, N.S.D. Treatment of diabetes mellitus by Artemisia Herba-Alba extract: Preliminary study. Clin. Exp. Pharmacol. Physiol. 1986, 13, 569–574. [Google Scholar] [CrossRef]
- Awad, N.E.; Seida, A.A.; Shaffie, Z.E.K.N.; El Aziz, A.M.A. Hypoglycemic activity of Artemisia herba-alba (Asso.) used in Egyptian traditional medicine as hypoglycemic remedy. J. Appl. Pharm. Sci. 2012, 2, 30–39. [Google Scholar] [CrossRef]
- Bolcato, G.; Bissaro, M.; Pavan, M.; Sturlese, M.; Moro, S. Targeting the coronavirus SARS-CoV-2: Computational insights into the mechanism of action of the protease inhibitors lopinavir, ritonavir and nelfinavir. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Li, S.-Y.; Chen, C.; Zhang, H.-Q.; Guo, H.-Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.-N.; Yu, J.; Xiao, P.-G.; et al. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Niwayama, S.; Hayashi, T.; Nago, R.; Ochiai, H.; Morita, N. In vitro and in vivo antiviral activity of scopadulcic acid B from Scoparia dulcis, Scrophulariaceae, against herpes simplex virus type 1. Antivir. Res. 1988, 9, 345–354. [Google Scholar] [CrossRef]
- Li, Y.; Ooi, L.S.M.; Wang, H.; But, P.P.H.; Ooi, V.E.C. Antiviral activities of medicinal herbs traditionally used in southern mainland China. Phytother. Res. 2004, 18, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Lelešius, R.; Karpovaitė, A.; Mickienė, R.; Drevinskas, T.; Tiso, N.; Ragažinskienė, O.; Kubilienė, L.; Maruška, A.; Šalomskas, A. In vitro antiviral activity of fifteen plant extracts against avian infectious bronchitis virus. BMC Vet. Res. 2019, 15, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, V.K. Rutin—A Potential Phyto-Therapeutic Agent against COVID-19 Protease Based on in Silico Studies. In Proceedings of the International e-Conference Recent Trends in Drug Discovery, Diagnostics and Therapeutics: A Special Emphasis on COVID-19, 2–4 July 2020; Available online: https://www.researchgate.net/publication/343685061_Rutin_-A_Potential_Phyto-Therapeutic_Agent_Against_COVID-19_Protease_Based_on_In_Silico_Studies (accessed on 11 November 2021).
- Mouhajir, F.; Hudson, J.B.; Rejdali, M.; Towers, G.H.N. Multiple Antiviral activities of endemic medicinal Plpnts used by Berber Pepples of Morocco. Pharm. Biol. 2001, 39, 364–374. [Google Scholar] [CrossRef]
- Grum-Tokars, V.; Ratia, K.; Begaye, A.; Baker, S.C.; Mesecar, A.D. Evaluating the 3C-like protease activity of SARS-Coronavirus: Recommendations for standardized assays for drug discovery. Virus Res. 2008, 133, 63–73. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.-H. An overview of severe acute respiratory syndrome–Coronavirus (SARS-CoV) 3CL protease inhibitors: Peptidomimetics and small molecule chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef]
- Ul Qamar, M.T.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef]
- Rehman, T.; Al Ajmi, M.F.; Hussain, A. Natural compounds as inhibitors of SARS-CoV-2 main protease (3CLpro): A molecular docking and simulation approach to combat COVID-19. Curr. Pharm. Des. 2021, 27, 3577–3589. [Google Scholar] [CrossRef]
- Al-Zahrani, A.A. Rutin as a Promising inhibitor of main protease and other protein targets of COVID-19: In silico study. Nat. Prod. Commun. 2020, 15. [Google Scholar] [CrossRef]
- Kamboj, A.; Saluja, A.K.; Kumar, M.; Atri, P. Antiviral activity of plant polyphenols. J. Pharm. Res. 2012, 5, 2402–2412. [Google Scholar]
- Coelho, G.R.; Figueiredo, C.A.; Negri, G.; Fernandes-Silva, C.C.; Villar, K.D.S.; Badari, J.C.; De Oliveira, M.I.; Barbosa, T.F.; Taniwaki, N.N.; Namiyama, G.M.; et al. Antiviral activity of Geopropolis extract from Scaptotrigona Aff. Postica against Rubella virus. J. Food Res. 2018, 7, 91–106. [Google Scholar] [CrossRef]
- Sarwar, M.W.; Riaz, A.; Dilshad, S.M.R.; Al-Qahtani, A.; Nawaz-Ul-Rehman, M.S.; Mubin, M. Structure activity relationship (SAR) and quantitative structure activity relationship (QSAR) studies showed plant flavonoids as potential inhibitors of dengue NS2B-NS3 protease. BMC Struct. Biol. 2018, 18, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venu, L.N.; Austin, A. Antiviral efficacy of medicinal plants against respiratory viruses: Respiratory Syscytial virus (RSV) and Coronavirus (COV)/COVID 19. J. Phytopharm. 2020, 9, 281–290. [Google Scholar] [CrossRef]
- Corrêa, W.R.; Serain, A.F.; Netto, L.A.; Marinho, J.V.N.; Arena, A.C.; Aquino, D.F.D.S.; Kuraoka-Oliveira, M.; Júnior, A.J.; Bernal, L.P.T.; Kassuya, C.A.L.; et al. Anti-inflammatory and antioxidant properties of the extract, tiliroside, and patuletin 3-O-β-D-glucopyranoside from Pfaffia townsendii (Amaranthaceae). Evid.-Based Complement. Altern. Med. 2018, 2018, 6057579. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, G.V.; Kuzhiumparambil, U.; Shree, A.B.R.; Balachandran, I. HPLC estimation of berberine in Tinospora cordifolia and Tinospora sinensis. Indian J. Pharm. Sci. 2008, 70, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Xing, H.; Ye, J. Efficacy of berberine in patients with type 2 diabetes mellitus. Metabolism 2008, 57, 712–717. [Google Scholar] [CrossRef] [Green Version]
- Warowicka, A.; Nawrot, R.; Goździcka-Józefiak, A. Antiviral activity of berberine. Arch. Virol. 2020, 165, 1935–1945. [Google Scholar] [CrossRef]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of natural products as potential inhibitors of COVID-19 main protease (Mpro): In-silico evidences. Nat. Prod. Bioprospecting 2020, 10, 297–306. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diogo, C.V.; Machado, N.G.; Barbosa, I.A.; Serafim, T.L.; Burgeiro, A.; Oliveira, P.J. Berberine as a promising safe anti-cancer agent—Is there a role for mitochondria? Curr. Drug Targets 2011, 12, 850–859. [Google Scholar] [CrossRef]
- Aljahdali, M.O.; Molla, M.H.R.; Ahammad, F. Compounds identified from marine Mangrove plant (Avicennia alba) as po-tential antiviral drug candidates against WDSV, an in-silico approach. Mar. Drugs 2021, 19, 253. [Google Scholar] [CrossRef]
- Ahammad, F.; Alam, R.; Mahmud, R.; Akhtar, S.; Kabir, E.U.; Tonmoy, A.M.; Fahim, S.; Alghamdi, K.; Samad, A.; Qadri, I. Pharmacoinformatics and molecular dynamics simulation-based phytochemical screening of Neem plant (Azadirachta indica) against human cancer by targeting MCM7 protein. Brief. Bioinform. 2021, 22, bbab098. [Google Scholar] [CrossRef] [PubMed]
- Krupanidhi, S.; Abraham Peele, K.; Venkateswarulu, T.; Ayyagari, V.S.; Nazneen Bobby, M.; John Babu, D.; Venkata Narayana, A.; Aishwarya, G. Screening of phytochemical compounds of Tinospora cordifolia for their inhibitory activity on SARS-CoV-2: An in silico study. J. Biomol. Struct. Dyn. 2021, 39, 5799–5803. [Google Scholar] [CrossRef]
- Baildya, N.; Khan, A.A.; Ghosh, N.N.; Dutta, T.; Chattopadhyay, A.P. Screening of potential drug from Azadirachta Indica (Neem) extracts for SARS-CoV-2: An insight from molecular docking and MD-simulation studies. J. Mol. Struct. 2021, 1227, 129390. [Google Scholar] [CrossRef]
- Elebeedy, D.; Elkhatib, W.F.; Kandeil, A.; Ghanem, A.; Kutkat, O.; Alnajjar, R.; Saleh, M.A.; El Maksoud, A.I.A.; Badawy, I.; Al-Karmalawy, A.A. Anti-SARS-CoV-2 activities of tanshinone IIA, carnosic acid, rosmarinic acid, salvianolic acid, baicalein, and glycyrrhetinic acid between computational and in vitro insights. RSC Adv. 2021, 11, 29267–29286. [Google Scholar] [CrossRef]
- Mahmud, S.; Rahman, E.; Nain, Z.; Billah, M.; Karmakar, S.; Mohanto, S.C.; Paul, G.K.; Amin, A.; Acharjee, U.K.; Saleh, A. Computational discovery of plant-based inhibitors against human carbonic anhydrase IX and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2020, 39, 2754–2770. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.; Mostafa, A.; Al-Karmalawy, A.A.; Zidan, A.; Abulkhair, H.S.; Mahmoud, S.H.; Shehata, M.; Elhefnawi, M.M.; Ali, M.A. Telaprevir is a potential drug for repurposing against SARS-CoV-2: Computational and in vitro studies. Heliyon 2021, 7, e07962. [Google Scholar] [CrossRef]
- Balunas, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef]
- Yang, Y.; Islam, M.S.; Wang, J.; Li, Y.; Chen, X. Traditional Chinese medicine in the treatment of patients infected with 2019-new Coronavirus (SARS-CoV-2): A Review and Perspective. Int. J. Biol. Sci. 2020, 16, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Jannat, K.; Paul, A.K.; Bondhon, T.A.; Hasan, A.; Nawaz, M.; Jahan, R.; Mahboob, T.; Nissapatorn, V.; Wilairatana, P.; Pereira, M.D.L.; et al. Nanotechnology applications of flavonoids for viral diseases. Pharmaceutics 2021, 13, 1895. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, R.; Saravanan, M.; Ovais, M.; Padmavathy, S.; Shinwari, Z.K.; Prakash, P. Antimicrobial efficacy of drug blended biosynthesized colloidal gold nanoparticles from Justicia glauca against oral pathogens: A nanoantibiotic approach. Microb. Pathog. 2017, 113, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.S.; Othman, H.H.; Hammadi, N.I.; Yeap, S.K.; Amin, K.M.; Samad, N.A.; Alitheen, N.B. Novel drug delivery systems for loading of natural plant extracts and their biomedical applications. Int. J. Nanomed. 2020, 15, 2439–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
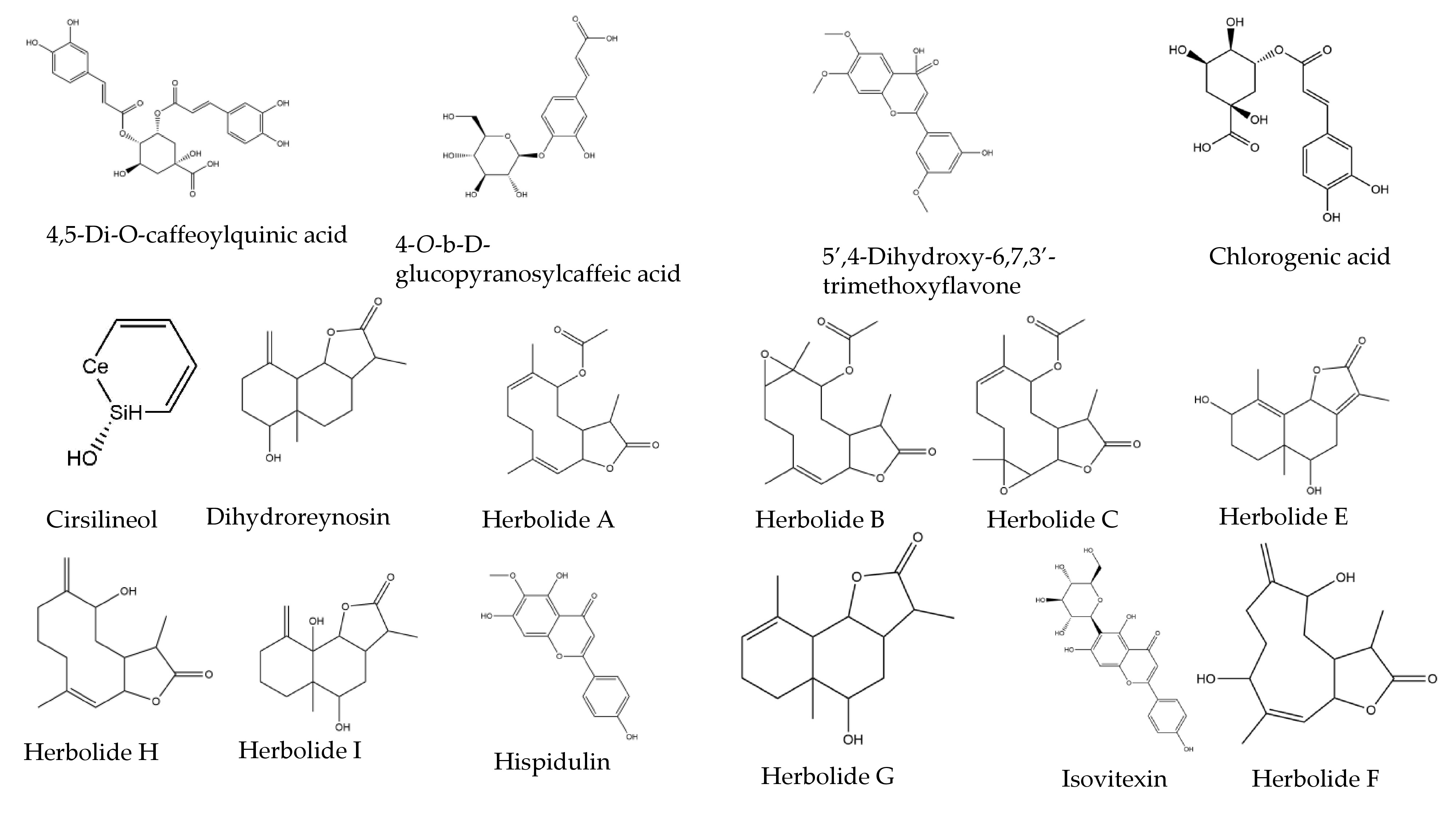
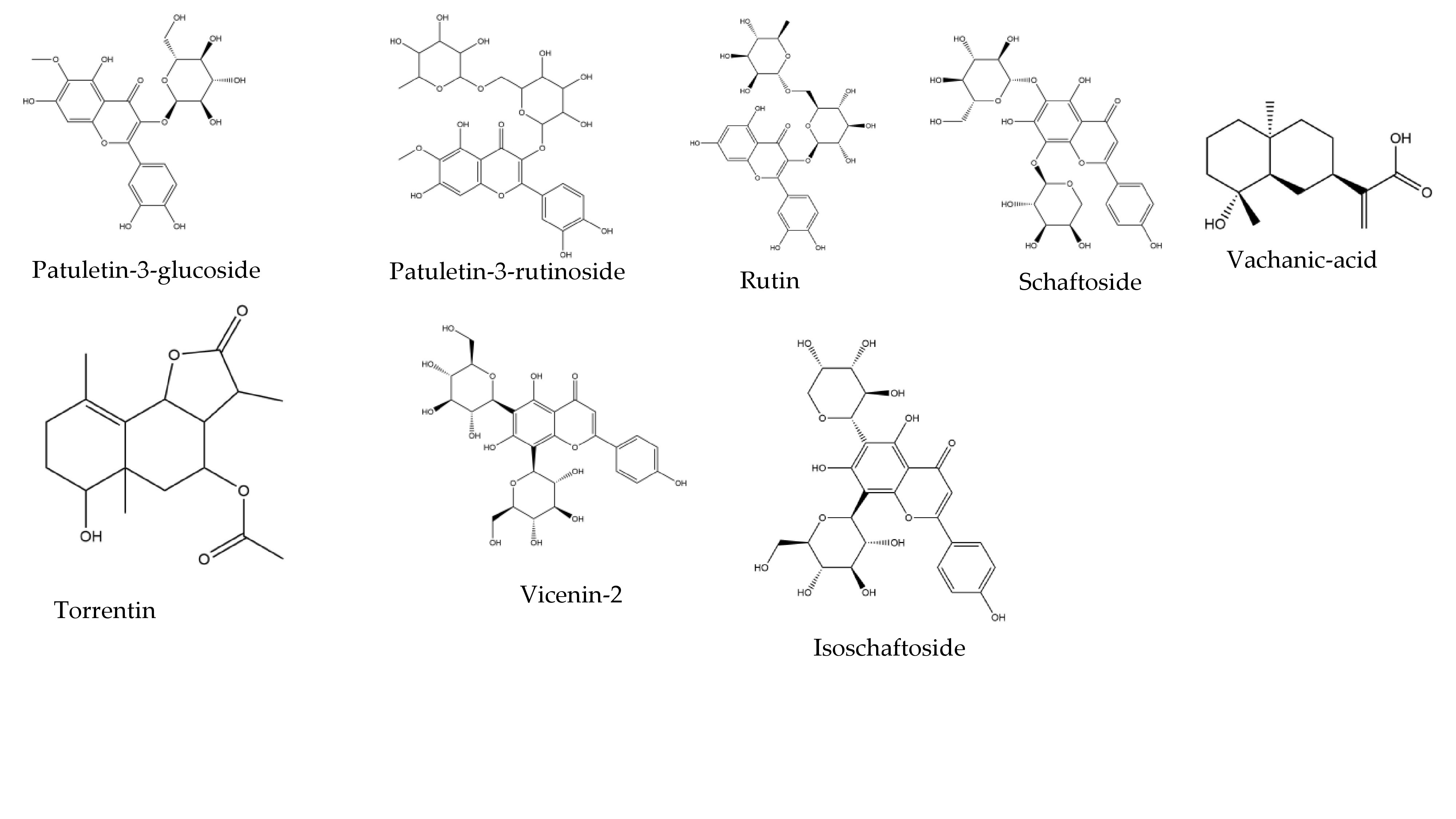
| Phytochemical | Binding Energy (ΔG = kcal/mol) to Mpro |
|---|---|
| 4,5-di-O-Caffeoylquinic acid | −8.5 |
| 4-O-b-D-glucopyranosylcaffeic acid | −6.8 |
| 5′4-Dihydroxy-6,7,3′-trimethoxyflavone | −7.2 |
| Chlorogenic acid | −7.5 |
| Cirsilineol | −7.3 |
| Dihydroreynosin | −6.7 |
| Herbolide A | −7.0 |
| Herbolide B | −6.2 |
| Herbolide C | −6.5 |
| Herbolide E | −6.5 |
| Herbolide F | −6.4 |
| Herbolide G | −7.1 |
| Herbolide H | −7.2 |
| Herbolide I | −6.9 |
| Hispidulin | −7.8 |
| Isoschaftoside | −7.3 |
| Isovitexin | −7.8 |
| Patuletin-3-glucoside | −8.0 |
| Patuletin-3-rutinoside | −7.9 |
| Rutin | −8.8 |
| Schaftoside | −8.3 |
| Torrentin | −6.3 |
| Vachanic_acid | −6.9 |
| Vicenin-2 | −7.9 |
| Lopinavir | −8.2 |
| Nelfinavir | −8.1 |
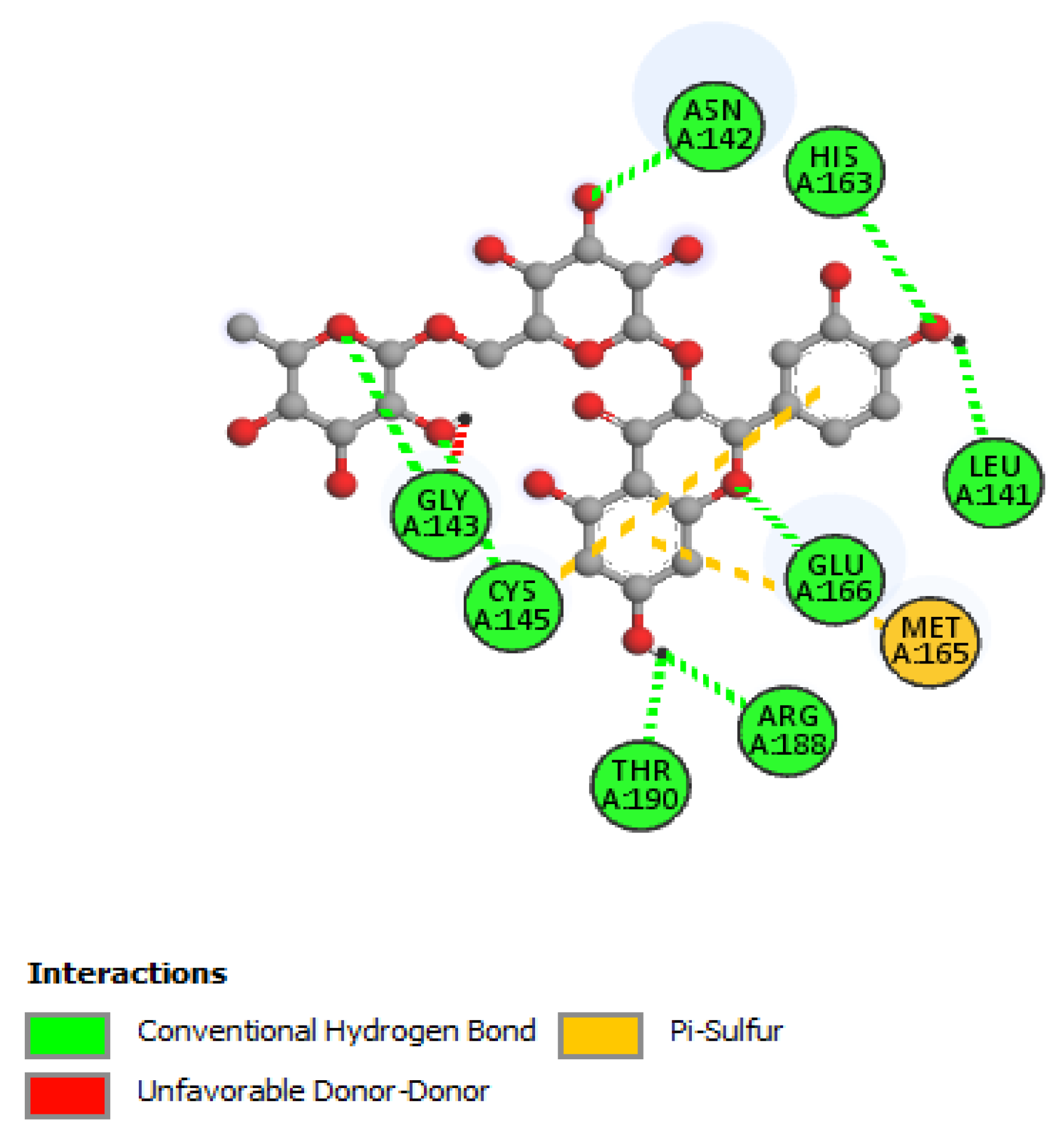
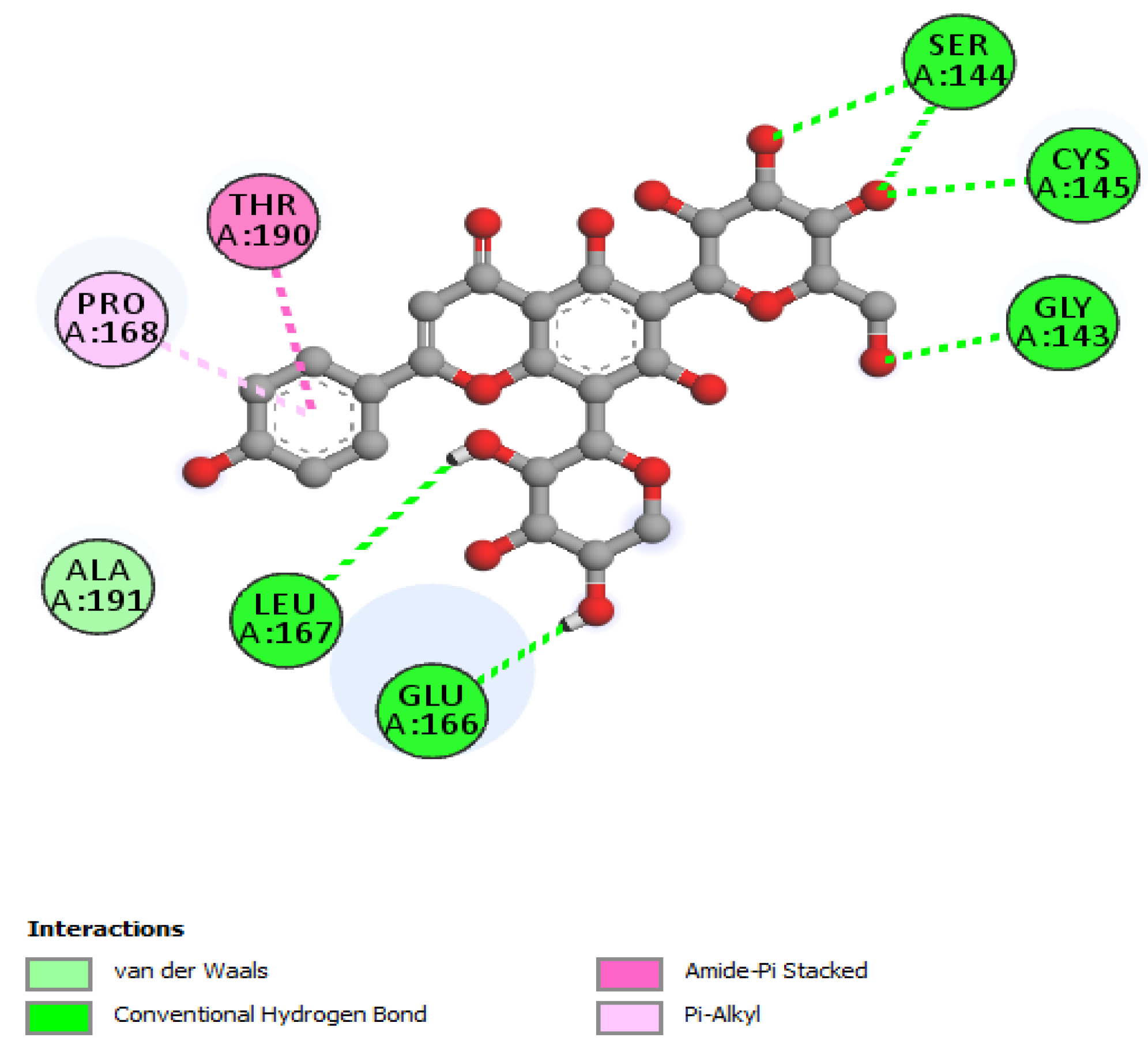
| Rutin | |||
|---|---|---|---|
| Interacting Residue | Distance | Category | Type |
| His41 | 2.33 | Hydrogen Bond | CH |
| Asn142 | 2.28 | Hydrogen Bond | CH |
| Gly143 | 2.27 | Hydrogen Bond | CH |
| Cys145 | 2.99 | Hydrogen Bond | CH |
| Glu166 | 2.27 | Hydrogen Bond | CH |
| Phe140 | 2.85 | Hydrogen Bond | CH |
| Thr190 | 2.60 | Hydrogen Bond | CH |
| Thr26 | 1.90 | Hydrogen Bond | CH |
| Cys145 | 4.97 | Other | Pi-Sulfur |
| Met165 | 5.12 | Other | Pi-Sulfur |
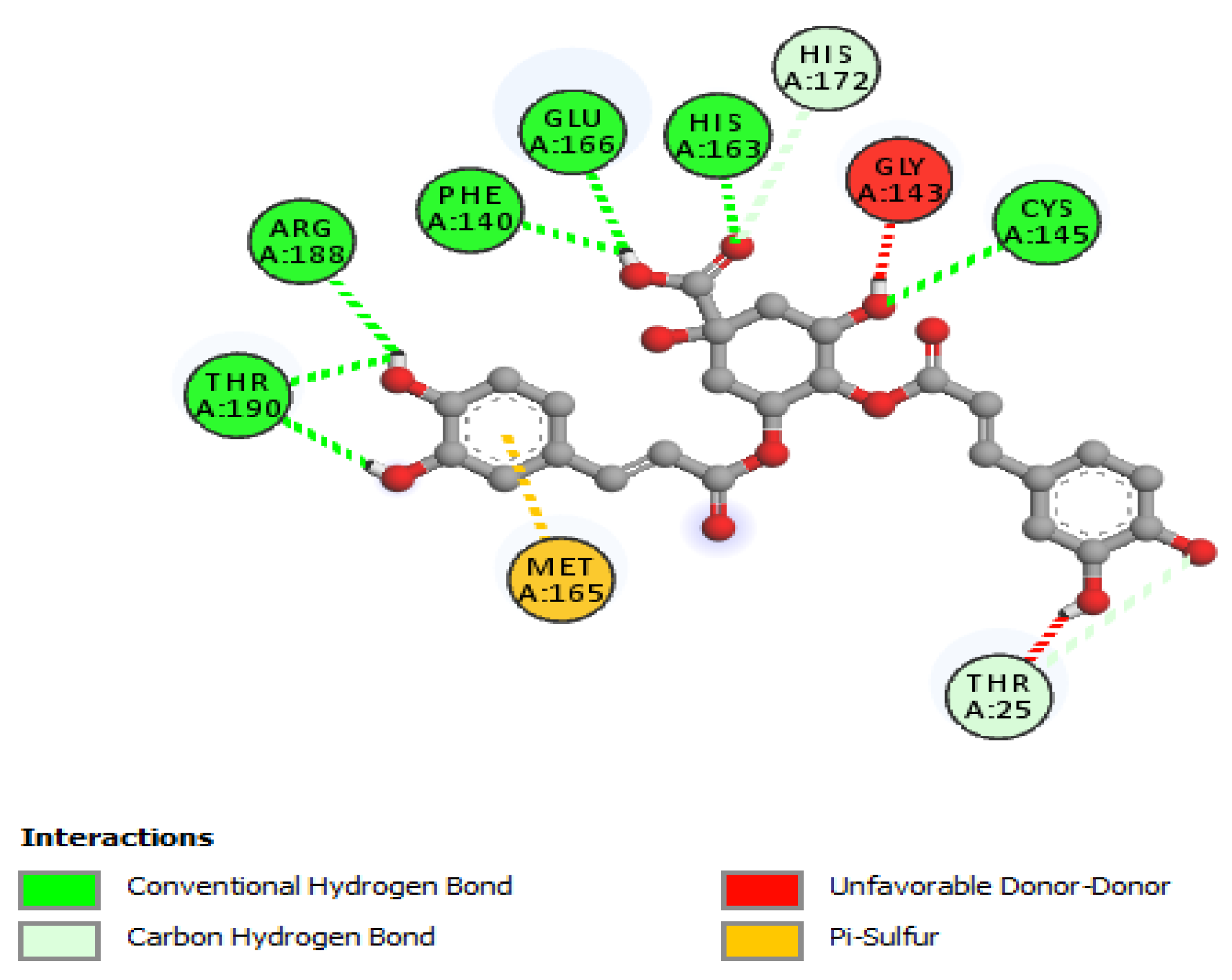
| 4,5-di-O-Caffeoylquinic acid | |||
| Cys145 | 2.75 | Hydrogen Bond | CH |
| His163 | 2.01 | Hydrogen Bond | CH |
| Thr190 | 2.02 | Hydrogen Bond | CH |
| Arg188 | 2.43 | Hydrogen Bond | CH |
| Thr190 | 2.32 | Hydrogen Bond | CH |
| Phe140 | 2.38 | Hydrogen Bond | CH |
| Glu166 | 2.25 | Hydrogen Bond | CH |
| Thr25 | 3.30 | Hydrogen Bond | C |
| His172 | 3.54 | Hydrogen Bond | C |
| Met165 | 4.88 | Other | Pi-Sulfur |
| Schaftoside | |||
| Gly143 | 2.18 | Hydrogen Bond | CH |
| Ser144 | 2.54 | Hydrogen Bond | CH |
| Ser144 | 2.71 | Hydrogen Bond | CH |
| Cys145 | 2.73 | Hydrogen Bond | CH |
| Glu166 | 2.270 | Hydrogen Bond | CH |
| Leu167 | 3.00 | Hydrogen Bond | CH |
| Thr190; Ala191 | 3.99 | Hydrophobic | Amide-Pi Stacked |
| Pro168 | 5.08 | Hydrophobic | Pi-Alkyl |
| Lopinavir | |||
| Gln110 | 2.12 | Hydrogen Bond | Conventional Hydrogen Bond |
| Asp153 | 2.42 | Hydrogen Bond | Conventional Hydrogen Bond |
| Ile249 | 3.44 | Hydrophobic | Pi-Sigma |
| Phe294 | 3.73 | Hydrophobic | Pi-Pi Stacked |
| Pro252 | 3.96 | Hydrophobic | Alkyl |
| Val104 | 3.78 | Hydrophobic | Alkyl |
| Phe294 | 5.19 | Hydrophobic | Pi-Alkyl |
| Pro252 | 5.39 | Hydrophobic | Pi-Alkyl |
| Val297 | 5.10 | Hydrophobic | Pi-Alkyl |
| Val202 | 5.14 | Hydrophobic | Pi-Alkyl |
| Nelfinavir | |||
| Asn142 | 2.25 | Hydrogen Bond | Conventional Hydrogen Bond |
| Leu141 | 2.88 | Hydrogen Bond | Conventional Hydrogen Bond |
| Cys145 | 5.90 | Other | Pi-Sulfur |
| Leu27 | 5.04 | Hydrophobic | Alkyl |
| Cys145 | 4.98 | Hydrophobic | Alkyl |
| Cys145 | 4.51 | Hydrophobic | Alkyl |
| Compound Name | Molecular Weight | Number of H-Bond Acceptors | Number of H-Bond Donors | Log P | Molar Refractivity | Number of Violation |
|---|---|---|---|---|---|---|
| 4,5-di-O-Caffeoylquinic acid | 516.45 | 12 | 7 | 1.25 | 126.90 | 3 |
| Chlorogenic acid | 354.31 | 9 | 6 | 0.96 | 83.50 | 1 |
| Hispidulin | 300.26 | 6 | 3 | 2.27 | 80.48 | 0 |
| Isoschaftoside | 564.49 | 14 | 10 | 1.41 | 133.26 | 3 |
| Isovitexin | 432.38 | 10 | 7 | 1.94 | 106.51 | 1 |
| Patuletin-3-glucoside | 494.40 | 13 | 8 | 2.45 | 116.65 | 2 |
| Patuletin-3-rutinoside | 640.50 | 17 | 10 | 1.61 | 147.87 | 3 |
| Rutin | 610.52 | 16 | 10 | 2.43 | 141.38 | 3 |
| Schaftoside | 564.49 | 14 | 10 | 1.63 | 133.26 | 3 |
| Vicenin-2 | 594.52 | 15 | 11 | 1.27 | 139.23 | 3 |
| Lopinavir | 628.80 | 5 | 4 | 3.44 | 187.92 | 2 |
| Nelfinavir | 567.78 | 5 | 4 | 3.87 | 166.17 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, A.; Biswas, P.; Bondhon, T.A.; Jannat, K.; Paul, T.K.; Paul, A.K.; Jahan, R.; Nissapatorn, V.; Mahboob, T.; Wilairatana, P.; et al. Can Artemisia herba-alba Be Useful for Managing COVID-19 and Comorbidities? Molecules 2022, 27, 492. https://doi.org/10.3390/molecules27020492
Hasan A, Biswas P, Bondhon TA, Jannat K, Paul TK, Paul AK, Jahan R, Nissapatorn V, Mahboob T, Wilairatana P, et al. Can Artemisia herba-alba Be Useful for Managing COVID-19 and Comorbidities? Molecules. 2022; 27(2):492. https://doi.org/10.3390/molecules27020492
Chicago/Turabian StyleHasan, Anamul, Partha Biswas, Tohmina Afroze Bondhon, Khoshnur Jannat, Tridib K. Paul, Alok K. Paul, Rownak Jahan, Veeranoot Nissapatorn, Tooba Mahboob, Polrat Wilairatana, and et al. 2022. "Can Artemisia herba-alba Be Useful for Managing COVID-19 and Comorbidities?" Molecules 27, no. 2: 492. https://doi.org/10.3390/molecules27020492