
Lipid Profiles of the Heads of Four Shrimp Species by UPLC–Q–Exactive Orbitrap/MS and Their Cardiovascular Activities

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
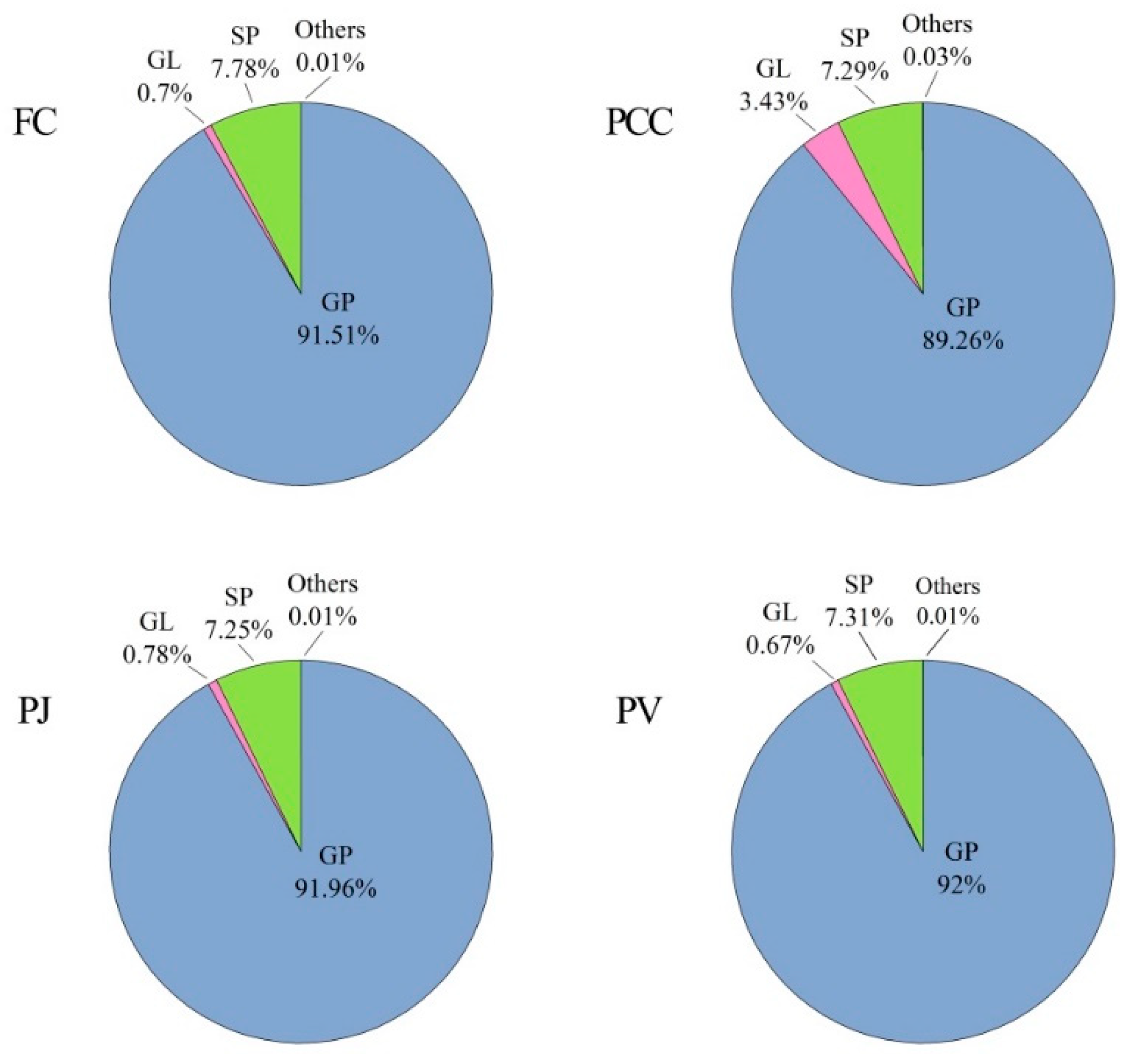
2.1. Lipid Fingerprint Identification
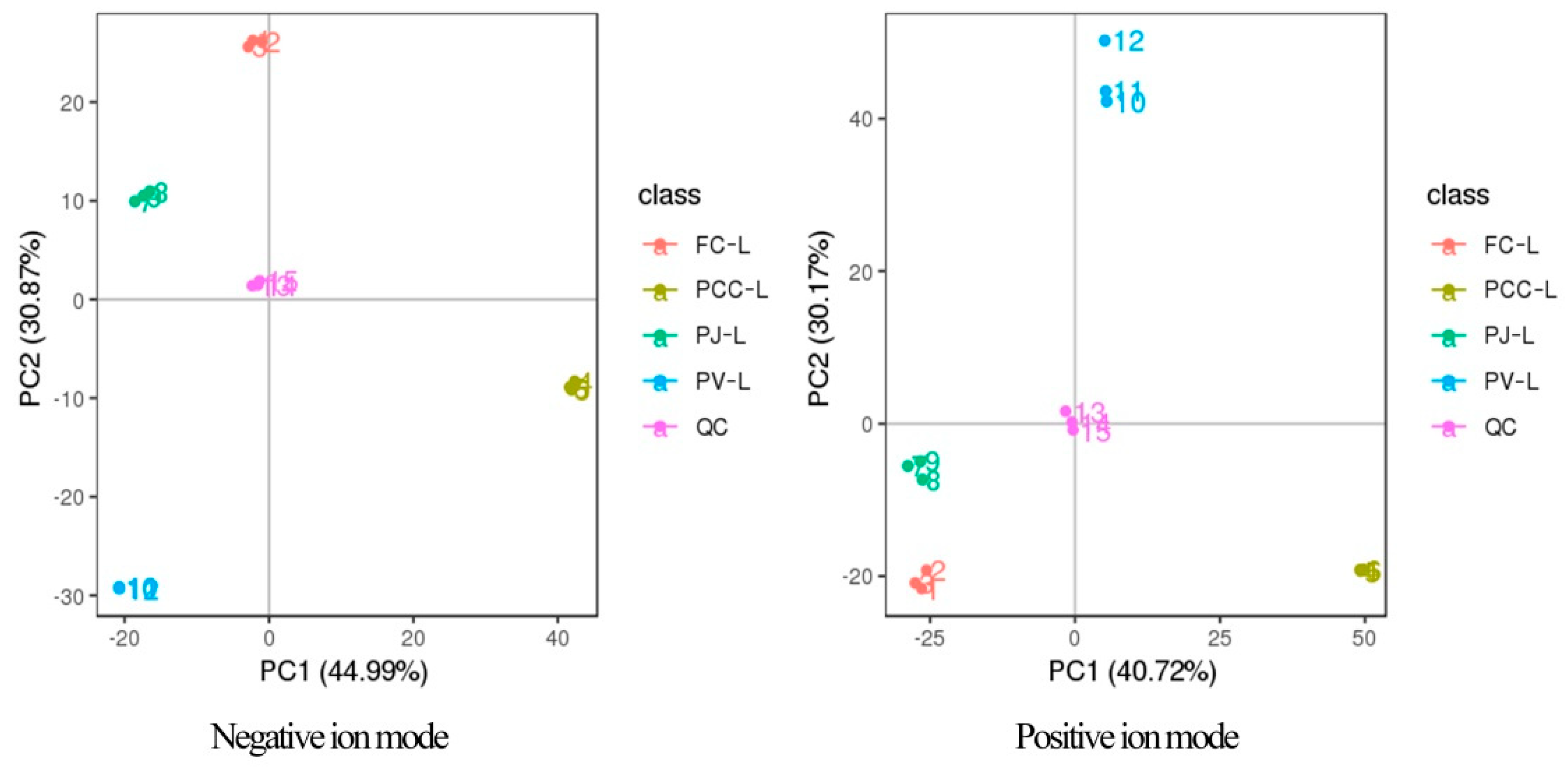
2.2. Multivariate Analysis
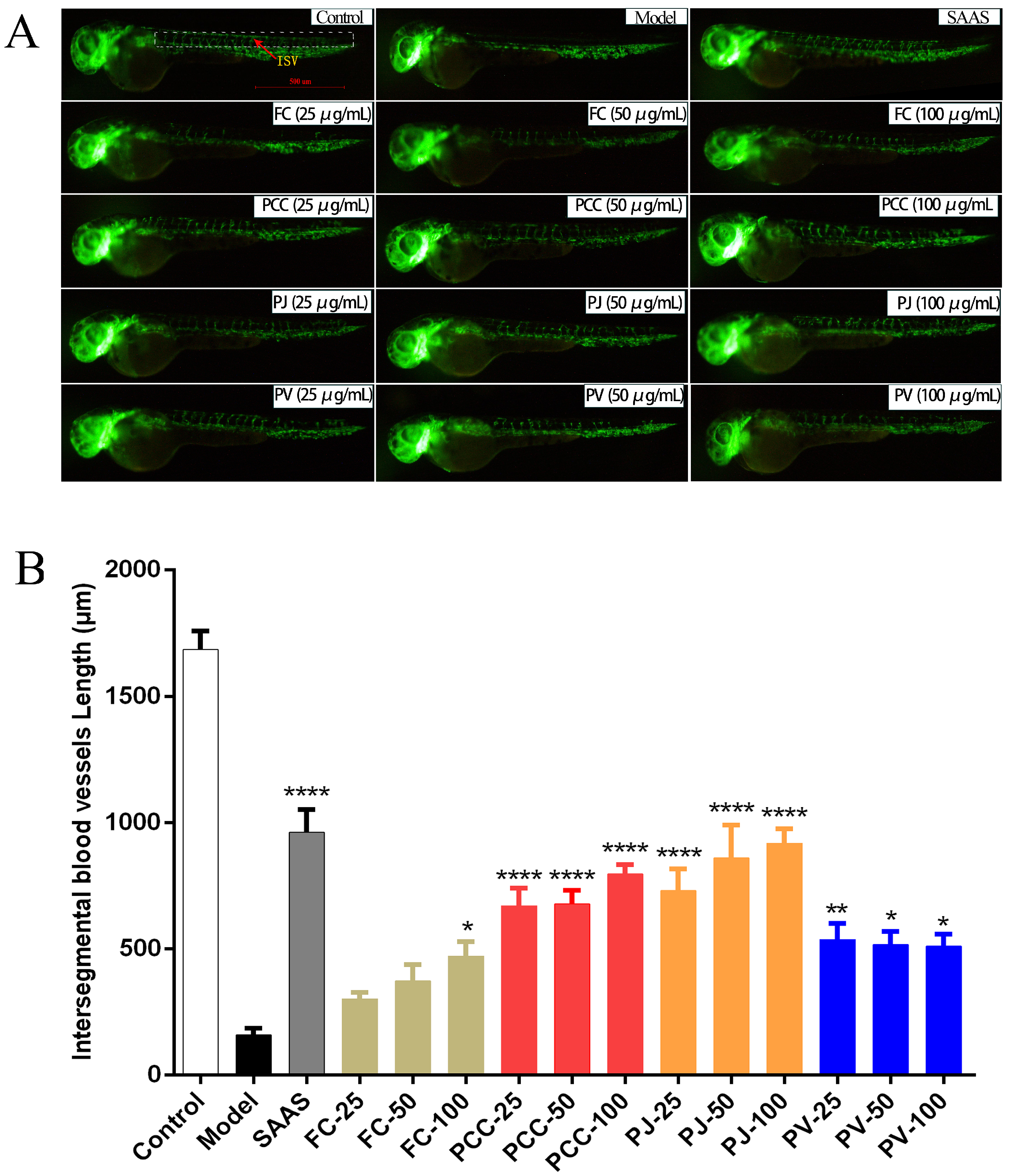
2.3. Pro-Angiogenic Activity
2.4. Anti-Inflammatory Activity
2.5. Antithrombotic Activity
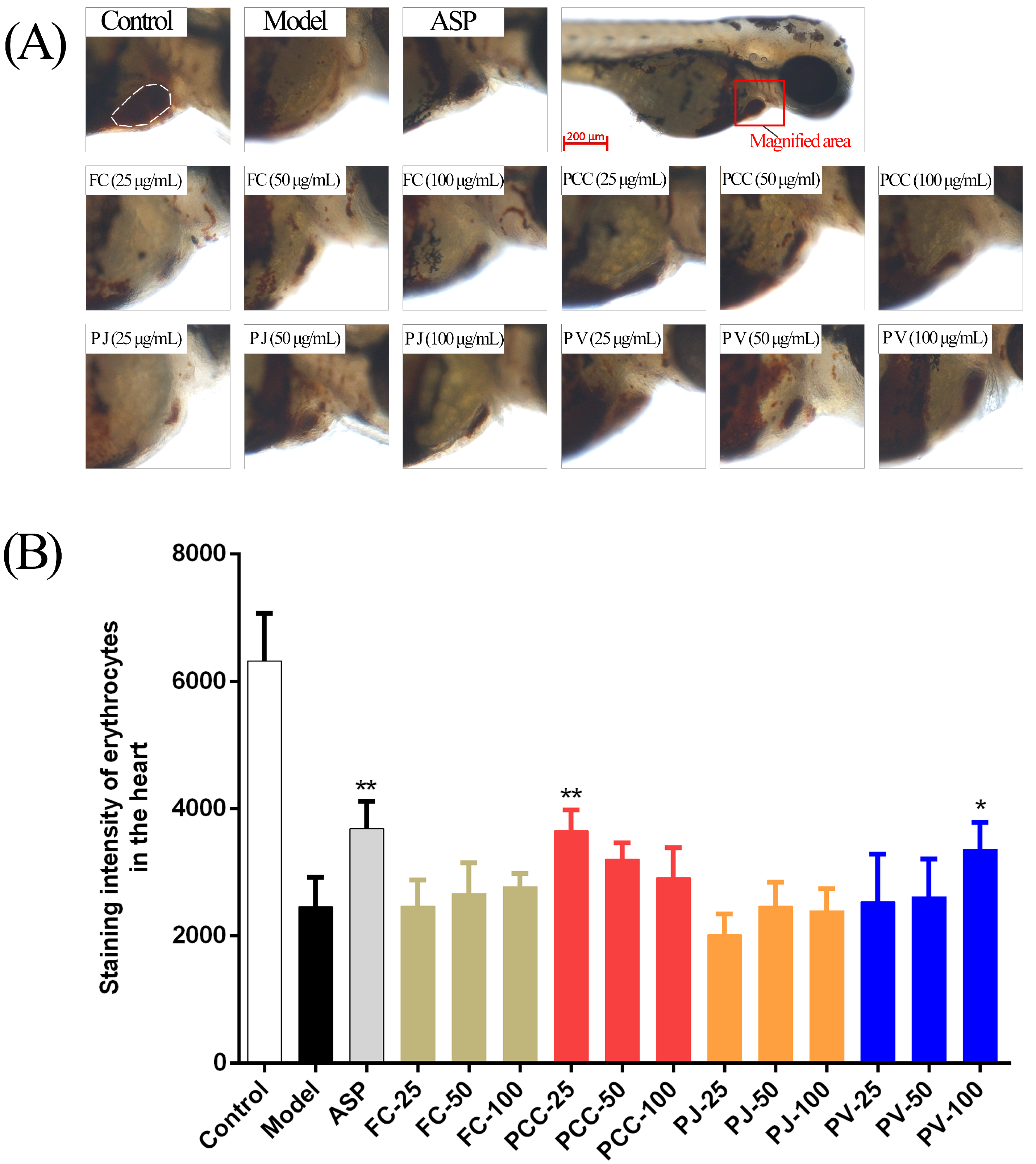
2.6. Cardiac Protective Activity
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Materials
4.3. Lipid Extraction
4.4. Sample Processing before Machine Analysis
4.5. UPLC–Q–Exactive Orbitrap/MS Analysis
4.6. Experimental Animals
4.7. Activity Assays of Zebrafish
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burdge, G.C.; Calder, P.C. Introduction to fatty acids and lipids. World Rev. Nutr. Diet. 2015, 112, 1–16. [Google Scholar] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid. Res. 2005, 46, 839–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuvillier, O. Sphingosine in apoptosis signaling. BBA—Mol.Cell Biol. L 2002, 1585, 153–162. [Google Scholar] [CrossRef]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Zu Heringdorf, D.M.; Jakobs, K.H. Lysophospholipid receptors: Signalling, pharmacology and regulation by lysophospholipid metabolism. BBA—Biomembranes 2007, 1768, 923–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, A.; Lee, H.; Goni, F.M.; Kolesnick, R.; Fernandez-Checa, J.C. Sphingolipids and cell death. Apoptosis 2007, 12, 923–939. [Google Scholar] [CrossRef]
- Bunea, R.; Farrah, K.E.; Deutsch, L. Evaluation of the effects of neptune krill oil on the clinical course of hyperlipidemia. Altern. Med. Rev. 2005, 9, 420–428. [Google Scholar]
- Hirafuji, M.; Machida, T.; Hamaue, N.; Minami, M. Cardiovascular protective effects of n-3 polyunsaturated fatty acids with special emphasis on docosahexaenoic acid. J. Pharmacol. Sci. 2003, 92, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Arshad, A.; Chung, W.Y.; Steward, W.; Metcalfe, M.S.; Dennison, A.R. Reduction in circulating pro-angiogenic and pro-inflammatory factors is related to improved outcomes in patients with advanced pancreatic cancer treated with gemcitabine and intravenous omega-3 fish oil. HPB 2013, 15, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Hsu, D.Z.; Chen, K.T.; Chien, S.P.; Li, Y.H.; Huang, B.M.; Chuang, Y.C.; Liu, M.Y. Sesame oil attenuates acute iron-induced lipid peroxidation-associated hepatic damage in mice. Shock 2006, 26, 625–630. [Google Scholar] [CrossRef]
- Deutsch, L. Evaluation of the effect of neptune krill oil on chronic inflammation and arthritic symptoms. J. Am. Coll. Nutr. 2007, 26, 39–48. [Google Scholar] [CrossRef]
- Li, Q.; Liang, X.; Zhao, L.; Zhang, Z.; Xue, X.; Wang, K.; Wu, L. Uplc-Q-exactive orbitrap/MS-based lipidomics approach to characterize lipid extracts from bee pollen and their in vitro anti-inflammatory properties. J. Agric. Food Chem. 2017, 65, 6848–6860. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrom. Rev. 2005, 24, 367–412. [Google Scholar] [CrossRef]
- Tang, H.Y.; Wang, C.H.; Ho, H.Y.; Wu, P.T.; Hung, C.L.; Huang, C.Y.; Wu, P.R.; Yeh, Y.H.; Cheng, M.L. Lipidomics reveals accumulation of the oxidized cholesterol in erythrocytes of heart failure patients. Redox Biol. 2018, 14, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Blanksby, S.J.; Mitchell, T.W. Advances in mass spectrometry for lipidomics. Annu. Rev. Anal. Chem. 2010, 3, 433–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, X. Analytical challenges of shotgun lipidomics at different resolution of measurements. TrAC Trend Anal. Chem. 2019, 121, 115697. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chang, M.; Tao, G.; Wang, X.; Liu, Y.; Liu, R.; Jin, Q.; Wang, X. Analysis of phospholipids in Schizochytrium sp. S31 by using UPLC-Q-TOF-MS. Anal. Methods 2016, 8, 763–770. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Comprehensive analysis of lipids in biological systems by liquid chromatography-mass spectrometry. TrAC Trend Anal. Chem. 2014, 61, 192–206. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.A.; Ochoa, B.; Fresnedo, O.; Giralt, M.T.; Rodríguez-Puertas, R. Matrix-assisted laser desorption ionization imaging mass spectrometry in lipidomics. Anal. Bioanal. Chem. 2011, 401, 29–51. [Google Scholar] [CrossRef]
- Li, J.; Vosegaard, T.; Guo, Z. Applications of nuclear magnetic resonance in lipid analyses: An emerging powerful tool for lipidomics studies. Prog. Lipid Res. 2017, 68, 37–56. [Google Scholar] [CrossRef]
- Li, M.; Zhou, Z.; Nie, H.; Bai, Y.; Liu, H. Recent advances of chromatography and mass spectrometry in lipidomics. Anal. Bioanal. Chem. 2011, 399, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Dennis, E.A. High sensitivity quantitative lipidomics analysis of fatty acids in biological samples by gas chromatography-mass spectrometry. BBA—Mol.Cell Biol. L 2011, 1811, 648–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.; Yang, Q.; Cheung, H.Y. Hydrophilic interaction chromatography based solid-phase extraction and MALDI TOF mass spectrometry for revealing the influence of Pseudomonas fluorescens on phospholipids in salmon fillet. Anal. Bioanal. Chem. 2015, 407, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, P.; Wang, F.; Zhang, S.; Li, H.; Zhang, Y.; Wang, X.; Liu, K.; Li, X. Separation, identification and cardiovascular activities of phospholipid classes from the head of Penaeus vannamei by lipidomics and zebrafish models. Food Funct. 2021, 12, 2282–2291. [Google Scholar] [CrossRef]
- Cobos, M.A.; Garcia, L.E.; González, S.; Barcena, J.R.; Hernández, D.S.; Sato, M.P. The effect of shrimp shell waste on ruminal bacteria and performance of lambs. Anim. Feed Sci. Technol. 2002, 95, 179–187. [Google Scholar] [CrossRef]
- Coward-Kelly, G.; Agbogbo, F.K.; Holtzapple, M.T. Lime treatment of shrimp head waste for the generation of highly digestible animal feed. Bioresour. Technol. 2006, 97, 1515–1520. [Google Scholar] [CrossRef]
- Yang, X.; Zu, T.H.; Zheng, Q.W.; Zhang, Z.S. Supercritical carbon dioxide extraction of the fatty acids from pacific white shrimp waste (Litopenaeus vannamei). Adv. Mater. Res. 2013, 2450, 506–510. [Google Scholar] [CrossRef]
- Scurria, A.; Tixier, A.S.F.; Lino, C.; Pagliaro, M.; D’Agostino, F.; Avellone, G.; Chemat, F.; Ciriminna, R. High yields of shrimp oil rich in omega-3 and natural astaxanthin from shrimp waste. ACS Omega 2020, 5, 17500–17505. [Google Scholar] [CrossRef]
- Asnani, A.; Peterson, R.T. The zebrafish as a tool to identify novel therapies for human cardiovascular disease. Dis. Model. Mech. 2014, 7, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Xin, S.C.; Zhao, Y.Q.; Li, S.; Lin, S.; Zhong, H.B. Application of zebrafish models in drug screening. Genetics 2012, 34, 1144–1152. [Google Scholar]
- Bournele, D.; Beis, D. Zebrafish models of cardiovascular disease. Heart Fail. Rev. 2016, 21, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. MetaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [Green Version]
- Tal, T.L.; Mccollum, C.W.; Harris, P.S.; Olin, J.; Kleinstreuer, N.; Wood, C.E.; Hans, C.; Shah, S.; Merchant, F.A.; Bondesson, M. Immediate and long-term consequences of vascular toxicity during zebrafish development. Reprod. Toxicol. 2014, 48, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.H.; Jiao, W.H.; Zhou, M.; Zhang, Y.; Zeng, D.Q.; Zhu, H.R.; Liu, K.C.; Sun, F.; Chen, H.F.; Lin, H.W. Septosones A–C, in vivo anti-inflammatory meroterpenoids with rearranged carbon skeletons from the marine sponge dysidea septosa. Org. Lett. 2019, 21, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Seuter, F.; Busse, W.D. Arachidonic acid-induced mortality in animals—An appropriate model for the evaluation of antithrombotic drugs? Agents Actions Suppl. 1979, 95, 175–183. [Google Scholar]
- Gyojeong, G.; Yirang, N.; Hyewon, C.; Hyeok, S.S.; Hae, Y. Zebrafish larvae model of dilated cardiomyopathy induced by terfenadine. Korean Circ. J. 2017, 47, 960–969. [Google Scholar]
- Zhang, X.G.; Zhou, A.M.; Lin, X.X.; Chen, Y.Q.; Yang, G.M. Comparative study of chemical compositions of white shrimp head and shell. Mod. Food Sci. Technol. 2009, 25, 224–227. [Google Scholar]
- Li, X.; Li, C.; Zhu, Y.; Shi, Y.; Zhang, X.; Zhang, S.; Wang, L.; Lin, H.W.; Hou, H.; Hsiao, C.D.; et al. Lipids fingerprinting of different materials sources by UPLC-Q-exactive orbitrap/MS approach and their zebrafish-based activities comparison. J. Agric. Food Chem. 2020, 68, 2007–2015. [Google Scholar] [CrossRef]
- Li, X.; He, Q.; Hou, H.; Zhang, S.; Zhang, X.; Zhang, Y.; Wang, X.; Han, L.; Liu, K. Targeted lipidomics profiling of marine phospholipids from different resources by UPLC-Q-exactive orbitrap/MS approach. J. Chromatogr. B 2018, 1096, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Xie, D.; Lu, M.; Li, P.; Lv, H.; Yang, C.; Peng, Q.; Zhu, Y.; Guo, L.; Zhang, Y.; et al. Characterization of white tea metabolome: Comparison against green and black tea by a nontargeted metabolomics approach. Food Res. Int. 2017, 96, 40–45. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Phospholipid Species | VIP Score | p-Value |
|---|---|---|---|
| FC vs. PCC | |||
| 1 | PC (18:2_18:3) | 3.154 | 1.02 × 10−3 |
| 2 | PC (19:0_16:1) | 3.037 | 2.01 × 10−2 |
| 3 | PC (24:1_22:6) | 3.021 | 1.64 × 10−4 |
| 4 | PE (18:1_24:1) | 2.790 | 1.40 × 10−5 |
| 5 | PC (14:0e_20:4) | 2.749 | 1.98 × 10−4 |
| FC vs. PJ | |||
| 1 | TG (12:0_15:1_18:3) | 3.217 | 1.20 × 10−4 |
| 2 | PC (22:6e_4:0) | 3.111 | 3.46 × 10−4 |
| 3 | TG (18:3_18:3_18:3) | 2.973 | 2.47 × 10−3 |
| 4 | PC (22:6e_2:0) | 2.942 | 1.52 × 10−4 |
| 5 | SM (d29:2_12:1) | 2.702 | 6.30 × 10−3 |
| FC vs. PV | |||
| 1 | PC (14:0e_18:2) | 3.766 | 1.39 × 10−2 |
| 2 | PC (19:0_18:1) | 3.607 | 2.91 × 10−2 |
| 3 | PC (16:0e_3:0) | 3.566 | 7.88 × 10−5 |
| 4 | PC (21:2_20:3) | 3.456 | 4.03 × 10−2 |
| 5 | PC (18:2_18:3) | 3.422 | 4.44 × 10−4 |
| PJ vs. PCC | |||
| 1 | TG (18:3_18:3_18:3) | 3.287 | 1.60 × 10−4 |
| 2 | PC (20:5e_16:4) | 3.127 | 5.37 × 10−6 |
| 3 | PC (17:2_16:3) | 2.984 | 1.50 × 10−7 |
| 4 | PC (18:3_18:2) | 2.911 | 7.21 × 10−3 |
| 5 | PC (19:0_16:1) | 2.521 | 2.48 × 10−2 |
| PJ vs. PV | |||
| 1 | PC (14:0e_18:2) | 3.536 | 1.22 × 10−2 |
| 2 | PC (18:0e_14:0) | 3.503 | 1.04 × 10−6 |
| 3 | PC (18:4e_16:4) | 3.437 | 1.99 × 10−3 |
| 4 | PC (19:0_18:1) | 3.290 | 3.58 × 10−2 |
| 5 | PC (14:0e_18:1) | 3.174 | 7.09 × 10−3 |
| PV vs. PCC | |||
| 1 | PC (21:1_21:1) | 3.554 | 1.75 × 10−6 |
| 2 | PC (24:0_18:1) | 3.318 | 3.94 × 10−4 |
| 3 | PC (21:0_22:1) | 3.047 | 1.53 × 10−4 |
| 4 | TG (18:3_18:3_18:3) | 3.042 | 3.89 × 10−6 |
| 5 | PC (18:1_24:1) | 2.849 | 1.25 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Li, P.; Meng, R.; Li, X.; Qiu, Y.; Wang, L.; Zhang, S.; Zhang, X.; Lin, H.; Zhai, H.; et al. Lipid Profiles of the Heads of Four Shrimp Species by UPLC–Q–Exactive Orbitrap/MS and Their Cardiovascular Activities. Molecules 2022, 27, 350. https://doi.org/10.3390/molecules27020350
Zhu Y, Li P, Meng R, Li X, Qiu Y, Wang L, Zhang S, Zhang X, Lin H, Zhai H, et al. Lipid Profiles of the Heads of Four Shrimp Species by UPLC–Q–Exactive Orbitrap/MS and Their Cardiovascular Activities. Molecules. 2022; 27(2):350. https://doi.org/10.3390/molecules27020350
Chicago/Turabian StyleZhu, Yongqiang, Peihai Li, Ronghua Meng, Xiaobin Li, Yuezi Qiu, Lizheng Wang, Shanshan Zhang, Xuanming Zhang, Houwen Lin, Hongbin Zhai, and et al. 2022. "Lipid Profiles of the Heads of Four Shrimp Species by UPLC–Q–Exactive Orbitrap/MS and Their Cardiovascular Activities" Molecules 27, no. 2: 350. https://doi.org/10.3390/molecules27020350