
Thermal Shift Assay as a Tool to Evaluate the Release of Breakdown Peptides from Cowpea β-Vignin during Seed Germination

 , , , , and
, , , , and 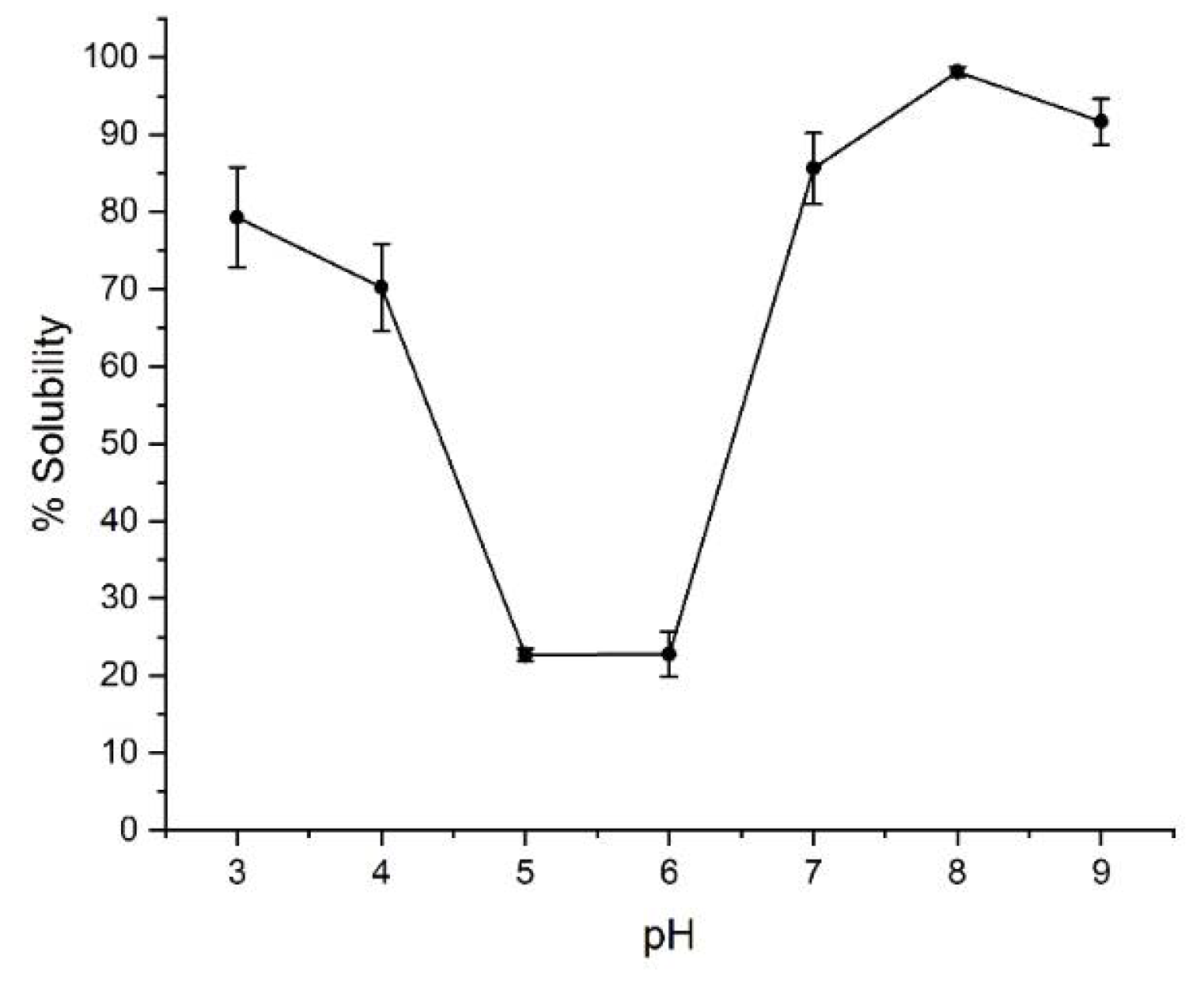{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Solubility
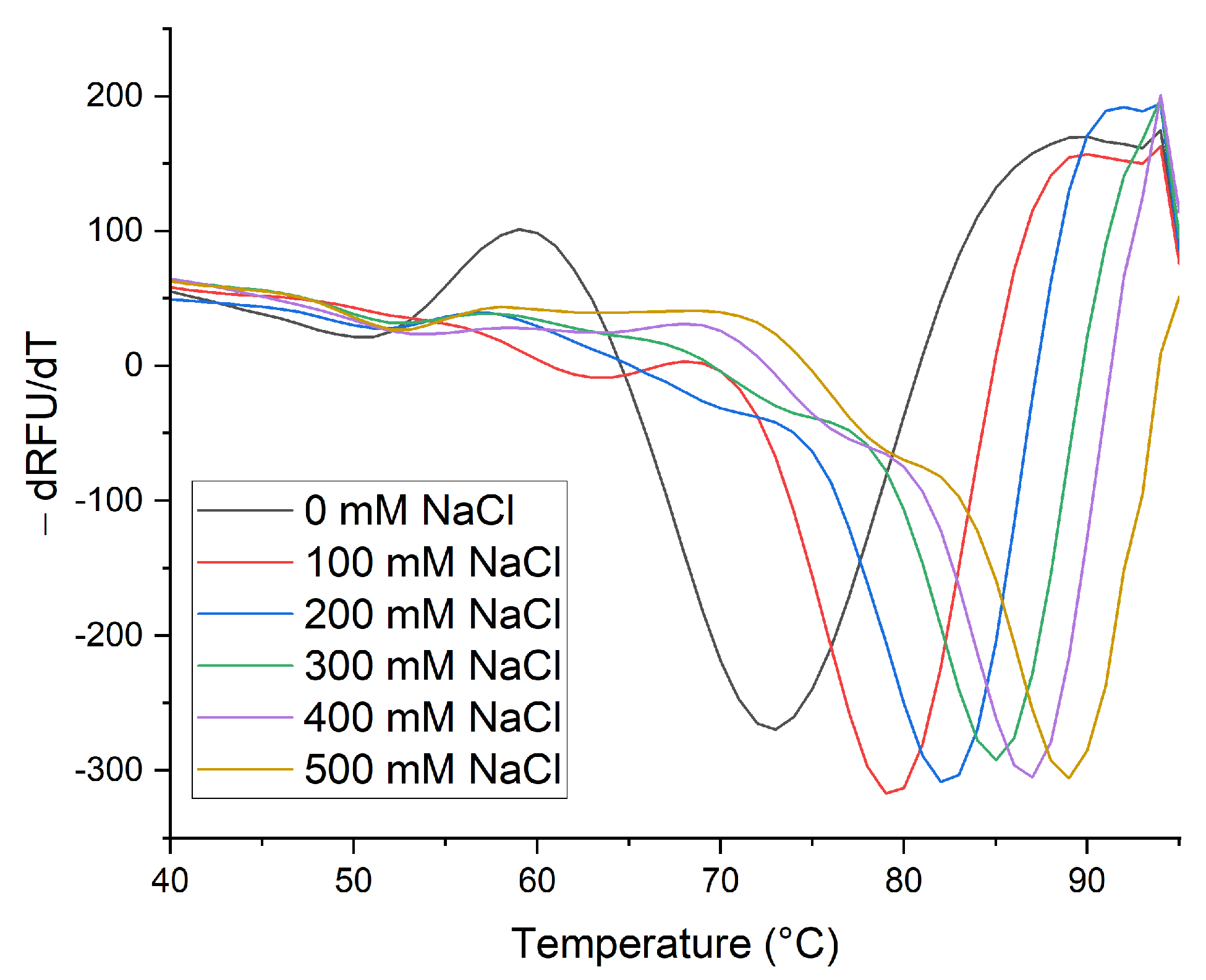
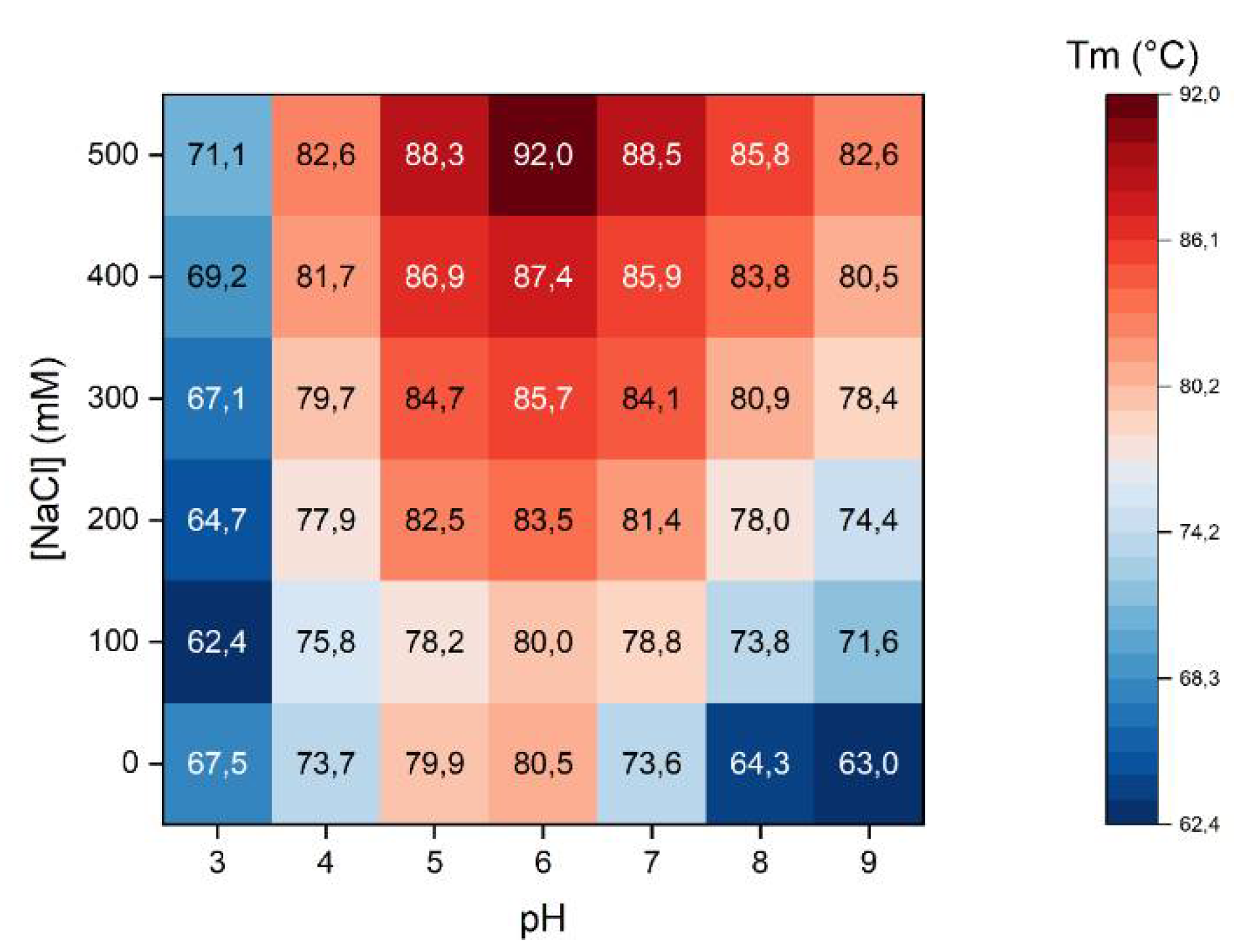
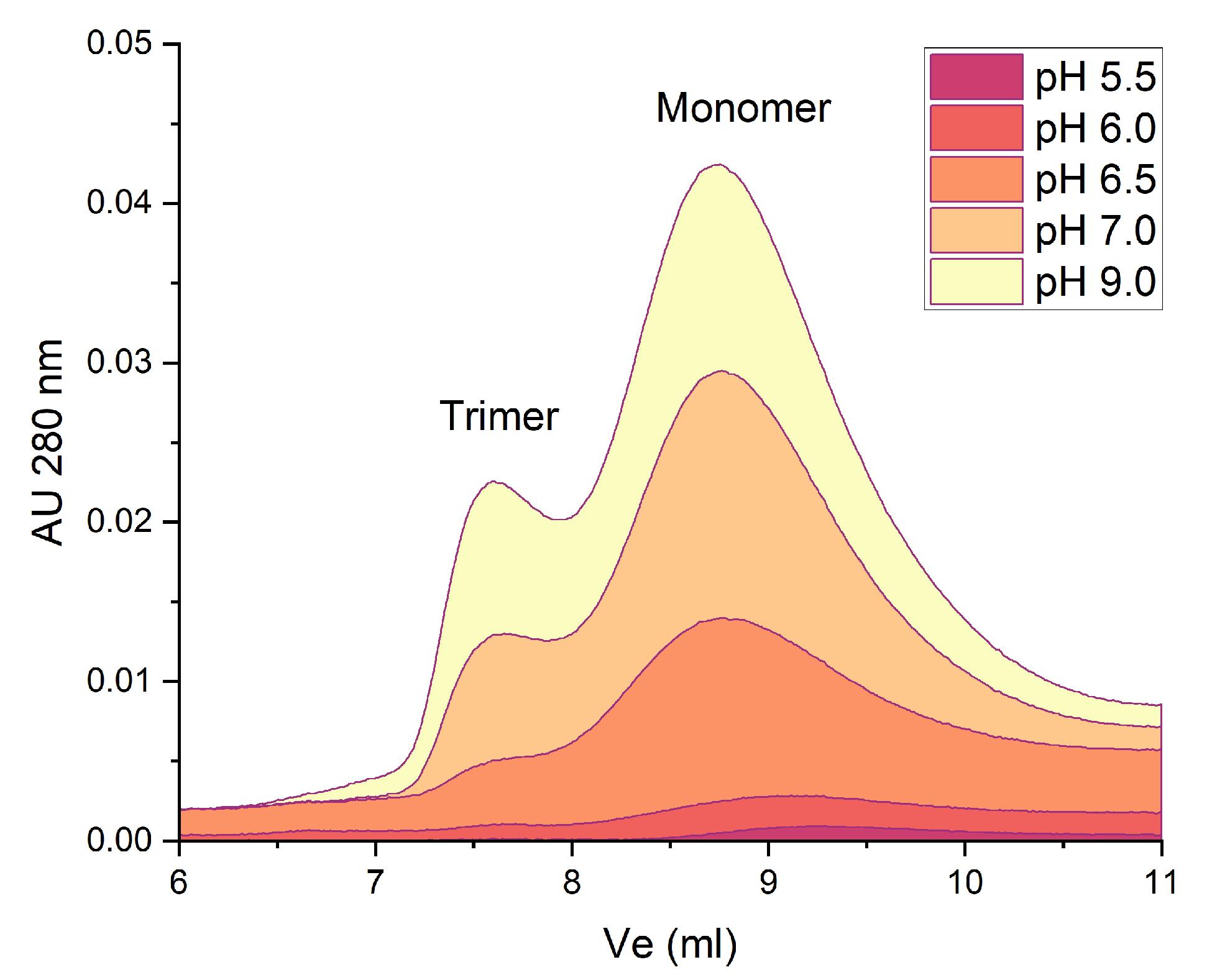
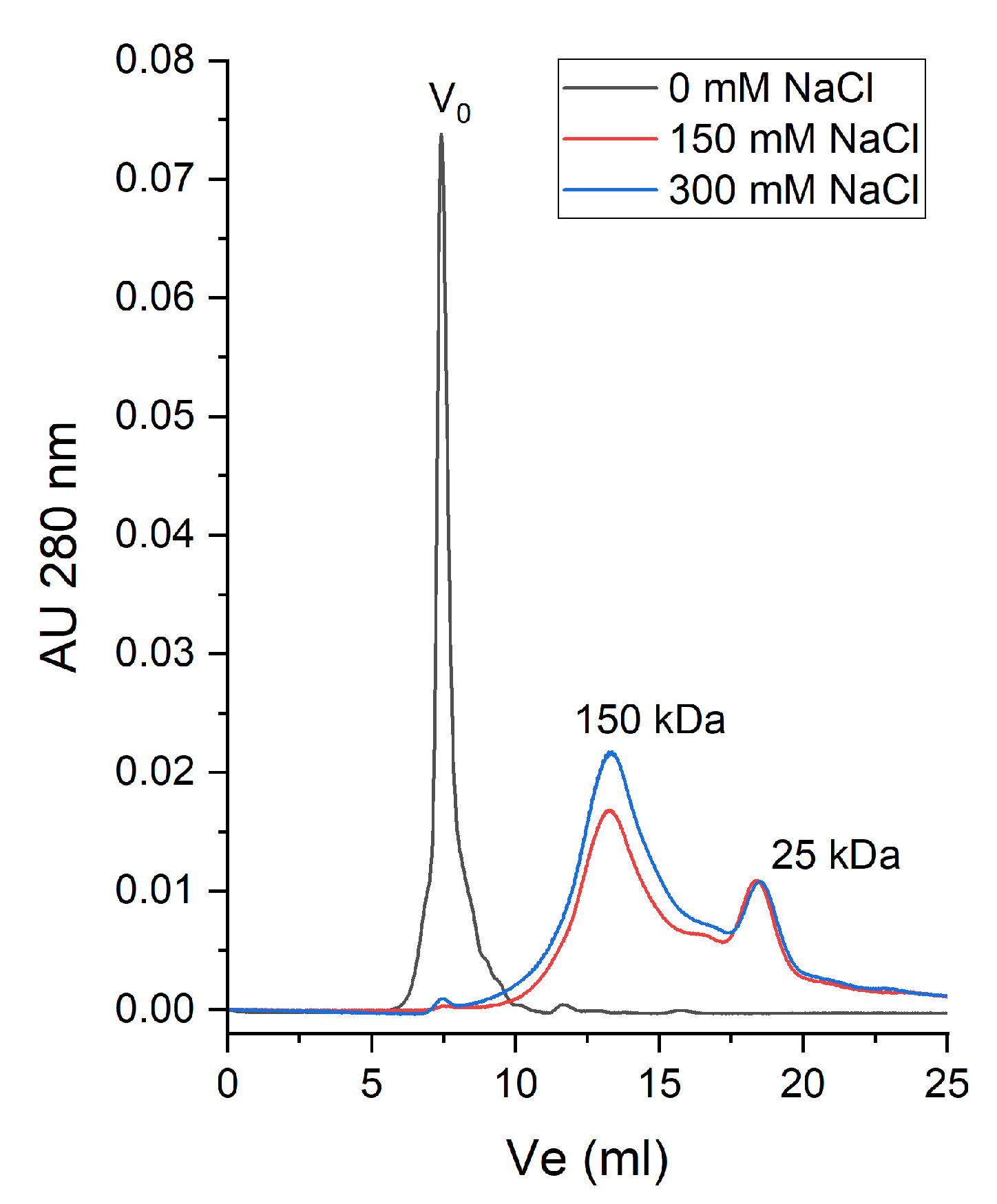
2.2. Thermal Stability of Native β-Vignin and Oligomerization
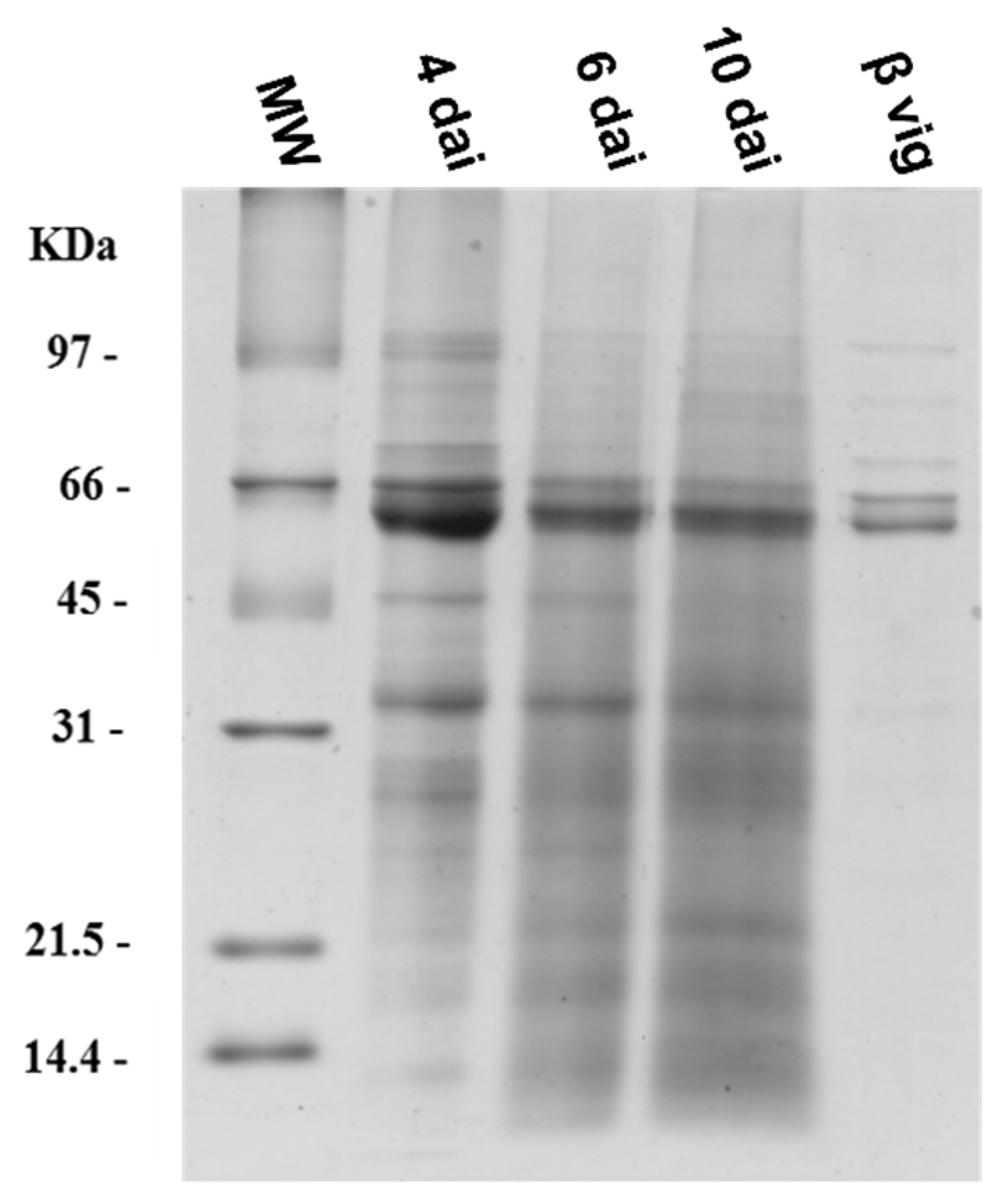
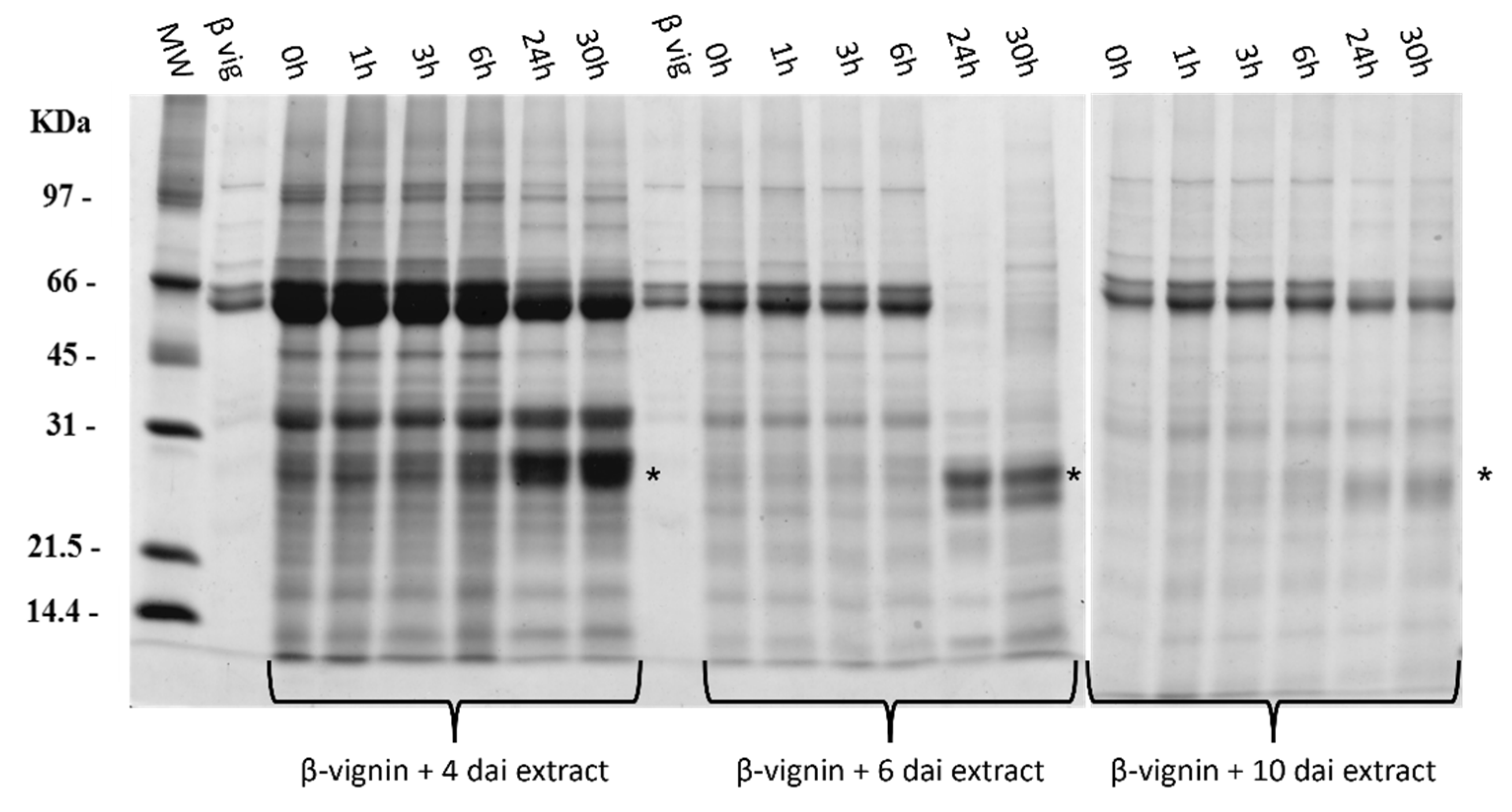
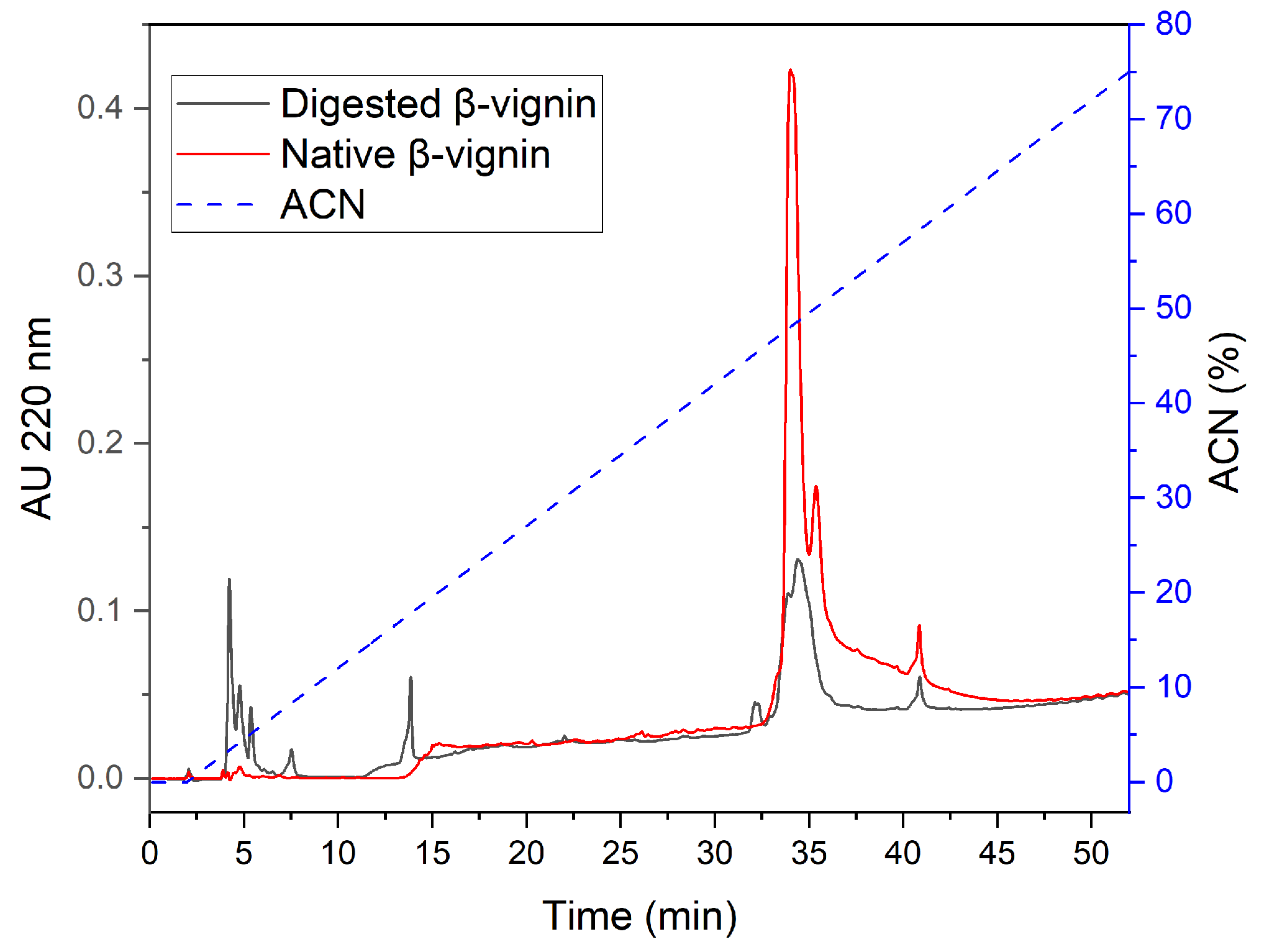
2.3. Proteases from Germinating Cowpea Seeds and β-Vignin Limited Proteolysis
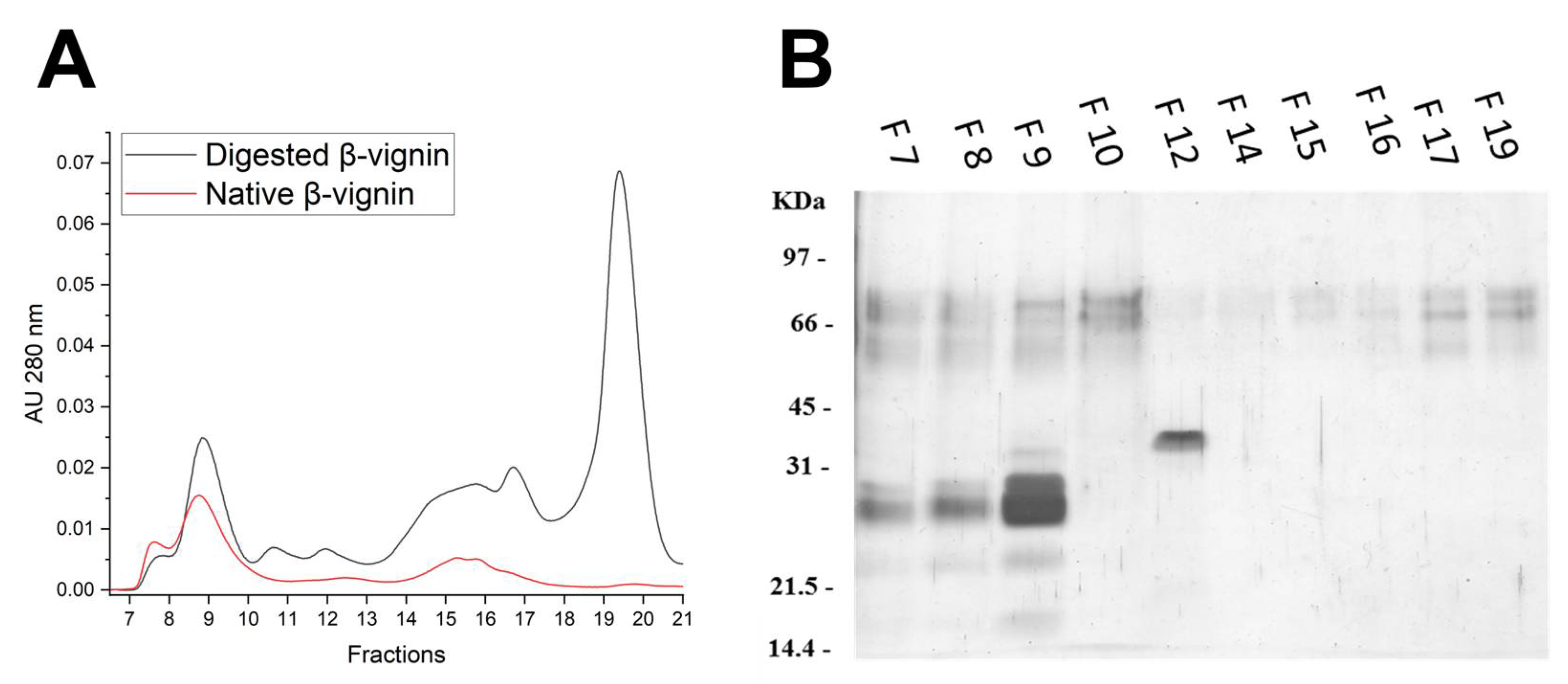
2.4. Structural Analysis of Proteolysis Main Products
2.5. Functional Biological Assays
2.5.1. Effects on Caco2 Cells
2.5.2. Effects on Fungal Conidial Germination
3. Materials and Methods
3.1. Cowpea Samples
3.2. Solubility
3.3. Chromatography
3.4. Proteases from Germinating Cowpea Seeds
3.5. β-Vignin Limited Proteolysis
3.6. SDS-PAGE
3.7. Thermal Shift Assay (TSA)
3.8. Effects on Fungal Germination
3.9. Caco-2 Cells Viability
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gupta, P.; Singh, R.; Malhotra, S.; Boora, K.S.; Singal, H.R. Characterization of Seed Storage Proteins in High Protein Genotypes of Cowpea [Vigna unguiculata (L.) Walp.]. Physiol. Mol. Biol. Plants Int. J. Funct. Plant Biol. 2010, 16, 53. [Google Scholar] [CrossRef] [Green Version]
- Gondwe, T.M.; Alamu, E.O.; Mdziniso, P.; Maziya-Dixon, B. Cowpea (Vigna unguiculata (L.) Walp) for Food Security: An Evaluation of End-User Traits of Improved Varieties in Swaziland. Sci. Rep. 2019, 9, 15991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, R.L.; Teixeira, A.R.; Ferreira, R.B. Characterization of the Proteins from Vigna Unguiculata Seeds. J. Agric. Food Chem. 2004, 52, 1682–1687. [Google Scholar] [CrossRef] [PubMed]
- Müntz, K. Deposition of Storage Proteins. Plant Mol. Biol. 1998, 38, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed Storage Proteins: Structures and Biosynthesis. Plant Cell 1995, 7, 945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza Ferreira, E.; Capraro, J.; Sessa, F.; Magni, C.; Demonte, A.; Consonni, A.; Augusto Neves, V.; Maffud Cilli, E.; Duranti, M.; Scarafoni, A. New Molecular Features of Cowpea Bean (Vigna unguiculata, l. Walp) β-Vignin. Biosci. Biotechnol. Biochem. 2018, 82, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Wakasa, Y.; Takaiwa, F. Seed Storage Proteins. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 346–348. ISBN 978-0-08-096156-9. [Google Scholar]
- Dall, E.; Brandstetter, H. Structure and Function of Legumain in Health and Disease. Biochimie 2016, 122, 126–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrispeels, M.J.; Boulter, D. Control of Storage Protein Metabolism in the Cotyledons of Germinating Mung Beans: Role of Endopeptidase. Plant Physiol. 1975, 55, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, J.P.; Green, J.L.; Goulter, K.C.; Manners, J.M. A Family of Antimicrobial Peptides Is Produced by Processing of a 7S Globulin Protein in Macadamia Integrifolia Kernels. Plant J. Cell Mol. Biol. 1999, 19, 699–710. [Google Scholar] [CrossRef]
- Jimenez-Lopez, J.C.; Melser, S.; De Boer, K.; Thatcher, L.F.; Kamphuis, L.G.; Foley, R.C.; Singh, K.B. Narrow-Leafed Lupin (Lupinus angustifolius) Β1- and Β6-Conglutin Proteins Exhibit Antifungal Activity, Protecting Plants against Necrotrophic Pathogen Induced Damage from Sclerotinia sclerotiorum and Phytophthora nicotianae. Front. Plant Sci. 2016, 7, 1856. [Google Scholar] [CrossRef] [Green Version]
- Fotso, M.; Azanza, J.-L.; Pasquet, R.; Raymond, J. Molecular Heterogeneity of Cowpea (Vigna unguiculata Fabaceae) Seed Storage Proteins. Plant Syst. Evol. 1994, 191, 39–56. [Google Scholar] [CrossRef]
- Huynh, K.; Partch, C.L. Analysis of Protein Stability and Ligand Interactions by Thermal Shift Assay. Curr. Protoc. Protein Sci. 2015, 79, 28.9.1–28.9.14. [Google Scholar] [CrossRef] [PubMed]
- Scarafoni, A.; Ronchi, A.; Prinsi, B.; Espen, L.; Assante, G.; Venturini, G.; Duranti, M. The Proteome of Exudates from Germinating Lupinus Albus Seeds Is Secreted through a Selective Dual-Step Process and Contains Proteins Involved in Plant Defence. FEBS J. 2013, 280, 1443–1459. [Google Scholar] [CrossRef]
- Capraro, J.; De Benedetti, S.; Di Dio, M.; Bona, E.; Abate, A.; Corsetto, P.A.; Scarafoni, A. Characterization of Chenopodin Isoforms from Quinoa Seeds and Assessment of Their Potential Anti-Inflammatory Activity in Caco-2 Cells. Biomolecules 2020, 10, 795. [Google Scholar] [CrossRef] [PubMed]
- Capraro, J.; Galanti, E.; Marengo, M.; Duranti, M.; Scarafoni, A. Cysteine-Containing Peptides Are Produced by Sequential Clipping, but Not Released, from Lupin 11S Storage Globulin during Early Germination. Peptidomics 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, S.D.; Galanti, E.; Capraro, J.; Magni, C.; Scarafoni, A. Lupinus Albus γ-Conglutin, a Protein Structurally Related to GH12 Xyloglucan-Specific Endo-Glucanase Inhibitor Proteins (XEGIPs), Shows Inhibitory Activity against GH2 β-Mannosidase. Int. J. Mol. Sci. 2020, 21, 7305. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-59259-890-8. [Google Scholar]
- Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Osman, A.; Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules 2019, 24, 1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capraro, J.; Spotti, P.; Magni, C.; Scarafoni, A.; Duranti, M. Spectroscopic Studies on the PH-Dependent Structural Dynamics of γ-Conglutin, the Blood Glucose-Lowering Protein of Lupin Seeds. Int. J. Biol. Macromol. 2010, 47, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Magni, C.; Scarafoni, A.; Herndl, A.; Sessa, F.; Prinsi, B.; Espen, L.; Duranti, M. Combined 2D Electrophoretic Approaches for the Study of White Lupin Mature Seed Storage Proteome. Phytochemistry 2007, 68, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Devi, C.B.; Kushwaha, A.; Kumar, A. Sprouting Characteristics and Associated Changes in Nutritional Composition of Cowpea (Vigna unguiculata). J. Food Sci. Technol. 2015, 52, 6821–6827. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Merril, C.R.; Goldman, D.; Sedman, S.A.; Ebert, M.H. Ultrasensitive Stain for Proteins in Polyacrylamide Gels Shows Regional Variation in Cerebrospinal Fluid Proteins. Science 1981, 211, 1437–1438. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.M.; Kunova, A.; Gardana, C.; Pizzatti, C.; Simonetti, P.; Cortesi, P.; Saracchi, M.; Pasquali, M. Investigating Useful Properties of Four Streptomyces Strains Active against Fusarium Graminearum Growth and Deoxynivalenol Production on Wheat Grains by QPCR. Toxins 2020, 12, 560. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Spanu, F.; Scherm, B.; Balmas, V.; Hoffmann, L.; Hammond-Kosack, K.E.; Beyer, M.; Migheli, Q. FcStuA from Fusarium Culmorum Controls Wheat Foot and Root Rot in a Toxin Dispensable Manner. PLoS ONE 2013, 8, e57429. [Google Scholar] [CrossRef] [Green Version]
- Colombo, E.M.; Pizzatti, C.; Kunova, A.; Gardana, C.; Saracchi, M.; Cortesi, P.; Pasquali, M. Evaluation of In-Vitro Methods to Select Effective Streptomycetes against Toxigenic Fusaria. PeerJ 2019, 7, e6905. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedetti, S.; Leogrande, C.; Castagna, F.; Heinzl, G.C.; Pasquali, M.; Heinzl, A.L.; Lupi, D.; Scarafoni, A. Thermal Shift Assay as a Tool to Evaluate the Release of Breakdown Peptides from Cowpea β-Vignin during Seed Germination. Molecules 2022, 27, 277. https://doi.org/10.3390/molecules27010277
De Benedetti S, Leogrande C, Castagna F, Heinzl GC, Pasquali M, Heinzl AL, Lupi D, Scarafoni A. Thermal Shift Assay as a Tool to Evaluate the Release of Breakdown Peptides from Cowpea β-Vignin during Seed Germination. Molecules. 2022; 27(1):277. https://doi.org/10.3390/molecules27010277
Chicago/Turabian StyleDe Benedetti, Stefano, Camilla Leogrande, Francesco Castagna, Giuditta C. Heinzl, Matias Pasquali, Alessandro L. Heinzl, Daniela Lupi, and Alessio Scarafoni. 2022. "Thermal Shift Assay as a Tool to Evaluate the Release of Breakdown Peptides from Cowpea β-Vignin during Seed Germination" Molecules 27, no. 1: 277. https://doi.org/10.3390/molecules27010277