
Co-Production of Isoprene and Lactate by Engineered Escherichia coli in Microaerobic Conditions


Abstract
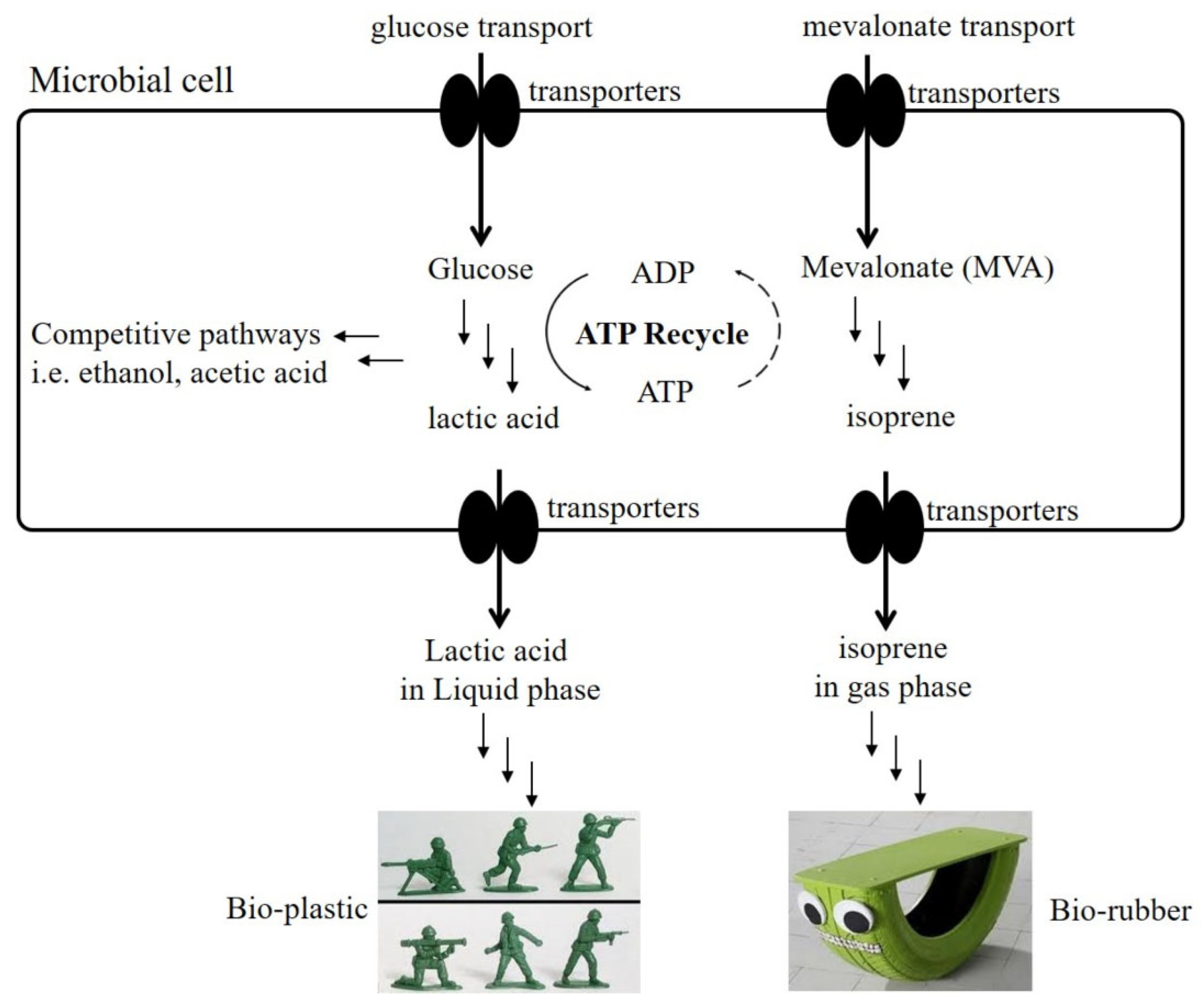
:1. Introduction
2. Results and Discussion
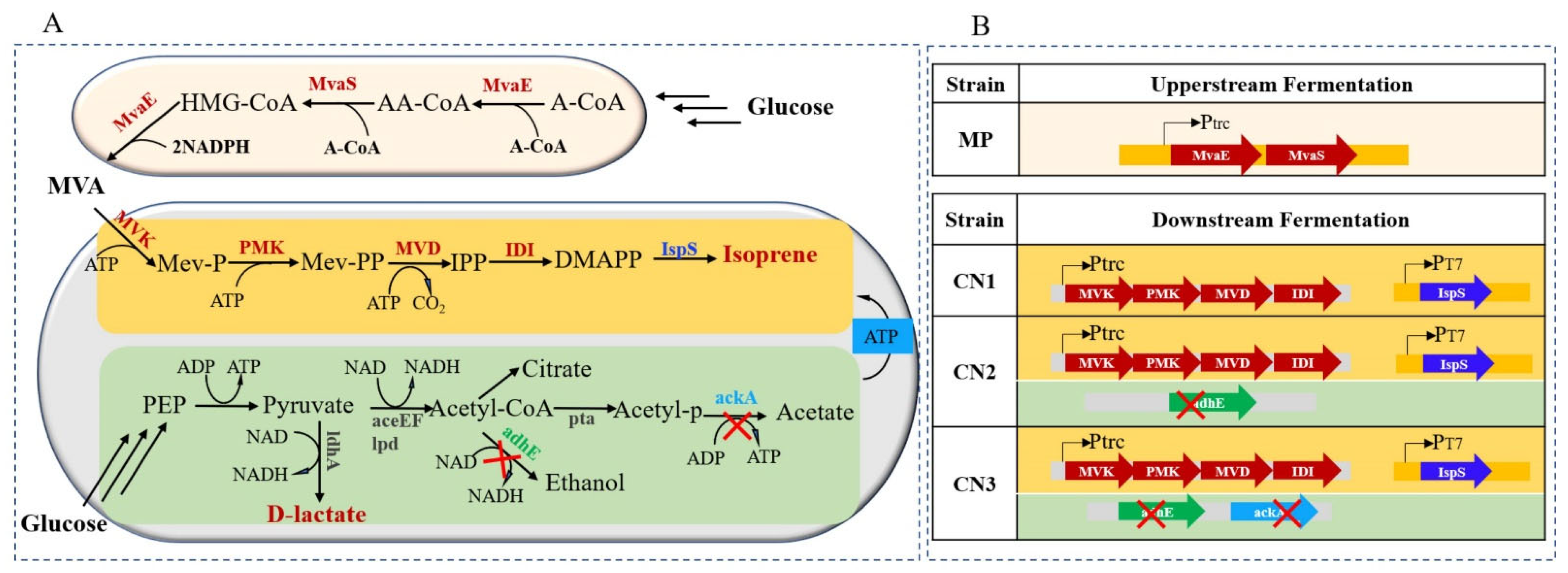
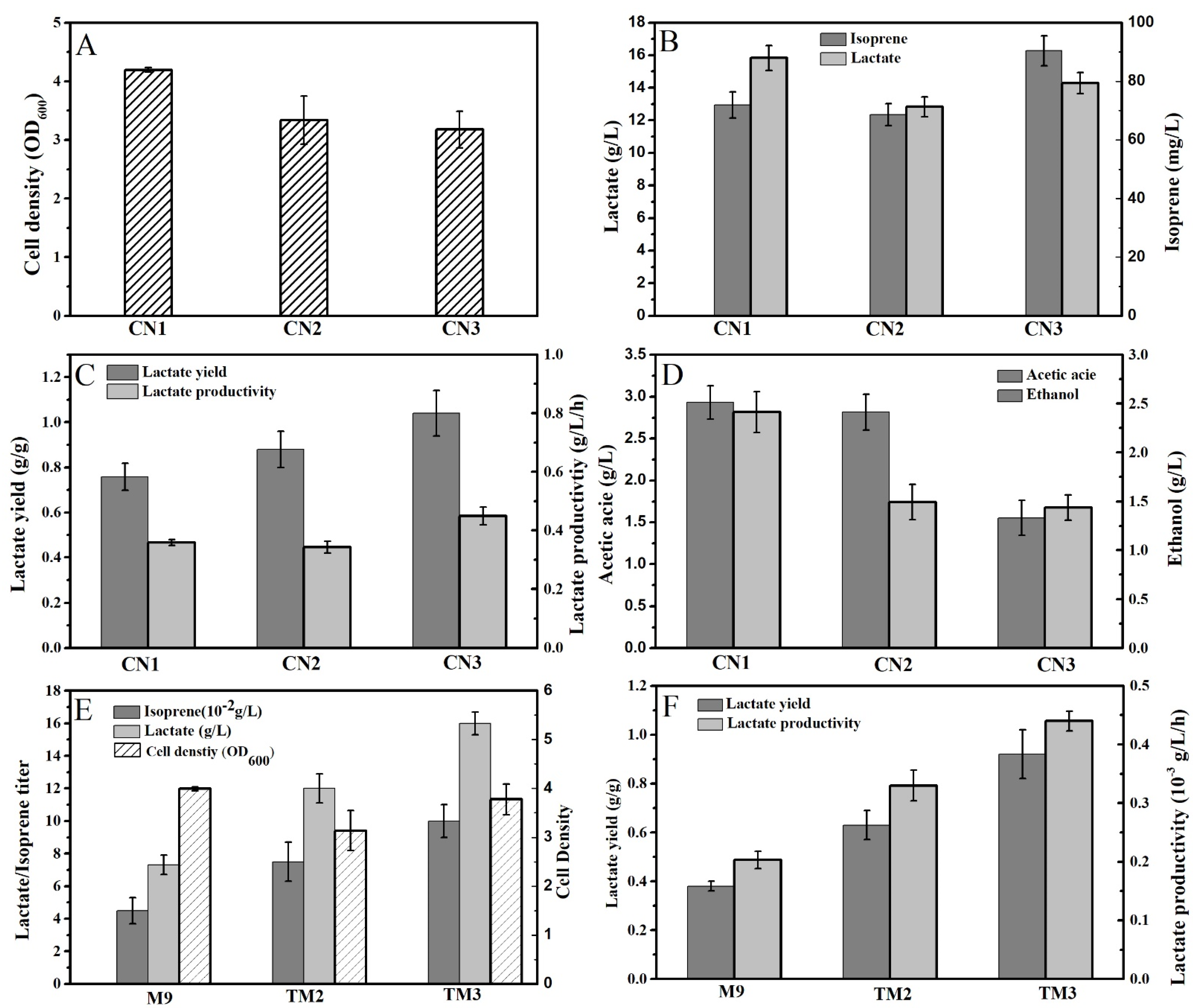
2.1. The Deletion of AdhE and AckA Improved the Lactate Production of Isoprene-Producing Strains
2.2. Medium Optimization Improved the Co-Production of Isoprene and D-Lactate by CN3
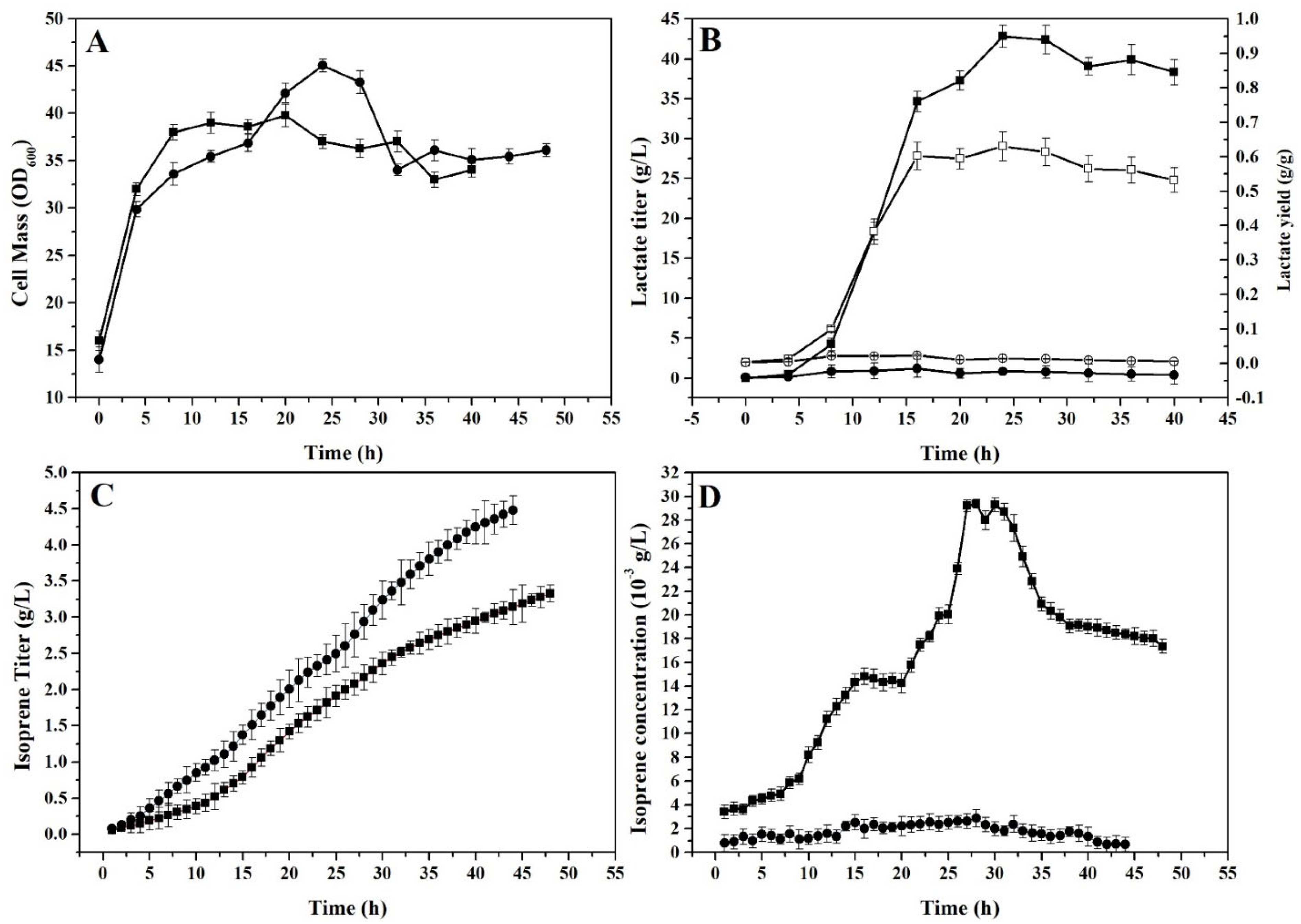
2.3. Fed-Batch Fermentation of Lactate and Isoprene under Aerobic or Microaerobic Conditions
3. Material and Methods
3.1. Strain Engineering
3.2. MVA Fermentation and Purification
3.3. Co-Fermentation of Isoprene and Lactic Acid at Flask Levels
3.4. Fed-Fermentation of Isoprene and Lactate
3.5. Analytical and Statistical Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kawaguchi, H.; Ogino, C.; Kondo, A. Microbial conversion of biomass into bio-based polymers. Bioresour. Technol. 2017, 245, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Whited, G.; Feher, F.; Benko, D.; Cervin, M.; Chotani, G.; McAuliffe, J.; LaDuca, R.; Ben-Shoshan, E.; Sanford, K. Development of a gas-phase bioprocess for isoprene-monomer production using metabolic pathway engineering. Ind. Biotechnol. 2010, 6, 152–163. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, T.W.; Kim, M.K.; Lee, S.Y.; Lim, S.C. Advanced bacterial polyhydroxyalkanoates: Towards a versatile and sustainable platform for unnatural tailor-made polyesters. Biotechnol. Adv. 2012, 30, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Nampoothiri, K.M.; Pandey, A. Fermentative production of lactic acid from biomass: An overview on process developments and future perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 524–534. [Google Scholar] [CrossRef]
- Zou, H.; Wu, Z.; Xian, M.; Liu, H.; Cheng, T.; Cao, Y. Not only osmoprotectant: Betaine increased lactate dehydrogenase activity and L-lactate production in lactobacilli. Bioresour. Technol. 2013, 148, 591–595. [Google Scholar] [CrossRef]
- Feng, X.J.; Ding, Y.M.; Xian, M.; Xu, X.; Zhang, R.B.; Zhao, G. Production of optically pure D-lactate from glycerol by engineered Klebsiella pneumoniae strain. Bioresour. Technol. 2014, 172, 269–275. [Google Scholar] [CrossRef]
- Zhou, S.D.; Shanmugam, K.T.; Ingram, L.O. Functional replacement of the Escherichia coli d-(-)-lactate dehydrogenase gene (ldhA) with the l-(+)-lactate dehydrogenase gene (ldhL) from Pediococcus acidilactici. Appl. Environ. Microb. 2003, 69, 2237–2244. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.L.; Zada, B.; Wei, G.Y.; Kim, S.W. Metabolic engineering and synthetic biology approaches driving isoprenoid production in Escherichia coli. Bioresour. Technol. 2017, 241, 430–438. [Google Scholar] [CrossRef]
- Yang, J.; Xian, M.; Su, S.; Zhao, G.; Nie, Q.; Jiang, X.; Zheng, Y.; Liu, W. Enhancing production of bio-isoprene using hybrid MVA pathway and isoprene synthase in E. coli. PLoS ONE 2012, 7, e33509. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Gao, X.; Jiang, Y.; Sun, B.; Gao, F.; Yang, S. Synergy between methylerythritol phosphate pathway and mevalonate pathway for isoprene production in Escherichia coli. Metab. Eng. 2016, 37, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Niu, D.D.; Tian, K.M.; Chen, X.Z.; Prior, B.A.; Shen, W.; Shi, G.Y.; Singh, S.; Wang, Z.X. Genetically switched D-lactate production in Escherichia coli. Metab. Eng. 2012, 14, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Grabar, T.B.; Shanmugam, K.T.; Ingram, L.O. Betaine tripled the volumetric productivity of d(-)-lactate by Escherichia coli strain SZ132 in mineral salts medium. Biotechnol. Lett. 2006, 28, 671–676. [Google Scholar] [CrossRef]
- Xu, J.; Xia, X.; Zhang, J.; Guo, Y.; Zhang, W. An overlooked effect of glycine betaine on fermentation: Prevents caramelization and increases the L-lysine production. J. Microbiol. Biotechnol. 2014, 24, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.B.; Chen, N.N.; Shi, M.X.; Xian, M.; Song, Y.M.; Liu, J.H. The metabolism and biotechnological application of betaine in microorganism. Appl. Microbiol. Biot. 2016, 100, 3865–3876. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, T.; Zou, H.B.; Zhang, H.B.; Xu, X.; Sun, C.; Aboulnaga, E.; Cheng, Z.K.; Zhao, G.; Xian, M. High titer mevalonate fermentation and its feeding as a building block for isoprenoids (isoprene and sabinene) production in engineered Escherichia coli. Process. Biochem. 2017, 62, 1–9. [Google Scholar] [CrossRef]
- Clark, D.P. The Fermentation Pathways of Escherichia coli. FEMS Microbiol. Lett. 1989, 63, 223–234. [Google Scholar] [CrossRef]
- Partridge, J.D.; Sanguinetti, G.; Dibden, D.P.; Roberts, R.E.; Poole, R.K.; Green, J. Transition of Escherichia coli from aerobic to micro-aerobic conditions involves fast and slow reacting regulatory components. J. Biol. Chem. 2007, 282, 11230–11237. [Google Scholar] [CrossRef] [Green Version]
- Lange, J.; Takors, R.; Blombach, B. Zero-growth bioprocesses: A challenge for microbial production strains and bioprocess engineering. Eng. Life Sci. 2017, 17, 27–35. [Google Scholar] [CrossRef]
- Lopez, P.C. Towards Industry 4.0 in the Bioprocessing Industries: ‘Real-Time’ monitoring and Control of Lignocellulosic Ethanol Fermentations. Ph.D. Thesis, Technical University of Denmark, Copenhagen, Denmark, 2020; pp. 73–92. [Google Scholar]
- Cheng, T.; Zhao, G.; Xian, M.; Xie, C. Improved cis-Abienol production through increasing precursor supply in Escherichia coli. Sci. Rep. 2020, 10, 16791. [Google Scholar] [CrossRef]
- Yang, J.M.; Zhao, G.; Sun, Y.Z.; Zheng, Y.N.; Jiang, X.L.; Liu, W.; Xian, M. Bio-isoprene production using exogenous MVA pathway and isoprene synthase in Escherichia coli. Bioresour. Technol. 2012, 104, 642–647. [Google Scholar] [CrossRef]
- Edwards, R.A.; Keller, L.H.; Schifferli, D.M. Improved allelic exchange vectors and their use to analyze 987P fimbria gene expression. Gene 1998, 207, 149–157. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids/Prmers | Description | Source |
|---|---|---|
| strains | ||
| E.coli BL21(DE3) | F-ompT hsdSB(rB-mB-) gal dcm rne131(DE3) | Invitrogen |
| E.coli DH5 | Cloning host | Invitrogen |
| E.coli χ7213 | Host strain for pRE112, DAP auxotrophic strain | [20] |
| MP | BL21(DE3)/pYJM16 | [9] |
| CM1 | BL21(DE3)::ptrc-mvk-pmk-mvd-idi | [20] |
| CM2 | BL21(DE3)::ptrc-mvk-pmk-mvd-idi△adhE | This study |
| CM3 | BL21(DE3)::ptrc-mvk-pmk-mvd-idi△adhE△ackA | This study |
| CN1 | CM1/pYJM8 | This study |
| CN2 | CM2/pYJM8 | This study |
| CN3 | CM3/pYJM8 | This study |
| Plasmids | ||
| pRE112 | Suicide vector, R6K origin, chloramphenicol resistant | [22] |
| pRE112-ΔSU | pRE112 derivative carrying genes glmS, glmU | This study |
| pRE112-ΔSU-trc-low | pRE112 derivative carrying genes glmS, glmU, ERG8, ERG12, ERG19 and IDI1, Trc promoter | This study |
| pRE112-adhE | Suicide vector for construction of △adhE mutant | This study |
| pRE112-ackA | Suicide vector for construction of △ackA mutant | This study |
| pYJM8 | pACYCDuet-1 derivative carrying isoprene synthase gene ispS, T7 promoter, CmR | [9] |
| pYJM14 | pTrcHis2B derivative carrying phosphomevalonate kinase gene ERG8, mevalonate kinase gene ERG12, mevalonate pyrophosphate decarboxylase gene ERG19 and IPP isomerase gene IDI1, Trc promoter, ApR | [9] |
| pYJM16 | pACYCDuet-1 derivative carrying acetyl-CoA acetyltransferase/hydroxymethylglutaryl-CoA (HMG-CoA) reductase gene mvaE and HMG-CoA synthase gene mvaS, T7 promoter, CmR | [9] |
| primers | ||
| adhE_F_F′ | CGAGTACTCCACAGACAGGTTGGCTGTAAG | |
| adhE_F_R | GTAGGTATCCAGATCTTCGACGATACCCATGC | |
| adhE_R_F | TATCGTCGAAGATCTGGATACCTACTACGGTCGTG | |
| adhE_R_R | GCTCTAGAGATGAGATTCGTTCGGAACAT | |
| ackA_F_F | CTAGTACTGATAACAGAACGATTATCCG | |
| ackA_F_R | TATATACGGCAGAAATTGATGATCG | |
| ackA_R_F | CATAAAACGGATCGCATAACGC | |
| ackA_R_R | GCTCTAGACATAACGAAGACGATTTCCGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, T.; Liang, X.; Wang, Y.; Chen, N.; Feng, D.; Liang, F.; Xie, C.; Liu, T.; Zou, H. Co-Production of Isoprene and Lactate by Engineered Escherichia coli in Microaerobic Conditions. Molecules 2021, 26, 7173. https://doi.org/10.3390/molecules26237173
Cheng T, Liang X, Wang Y, Chen N, Feng D, Liang F, Xie C, Liu T, Zou H. Co-Production of Isoprene and Lactate by Engineered Escherichia coli in Microaerobic Conditions. Molecules. 2021; 26(23):7173. https://doi.org/10.3390/molecules26237173
Chicago/Turabian StyleCheng, Tao, Xiuhong Liang, Yaqun Wang, Ningning Chen, Dexin Feng, Fengbing Liang, Congxia Xie, Tao Liu, and Huibin Zou. 2021. "Co-Production of Isoprene and Lactate by Engineered Escherichia coli in Microaerobic Conditions" Molecules 26, no. 23: 7173. https://doi.org/10.3390/molecules26237173