
Antiviral Therapeutic Potential of Curcumin: An Update

 , , and
, , and
Abstract
:1. Introduction
2. Curcumin
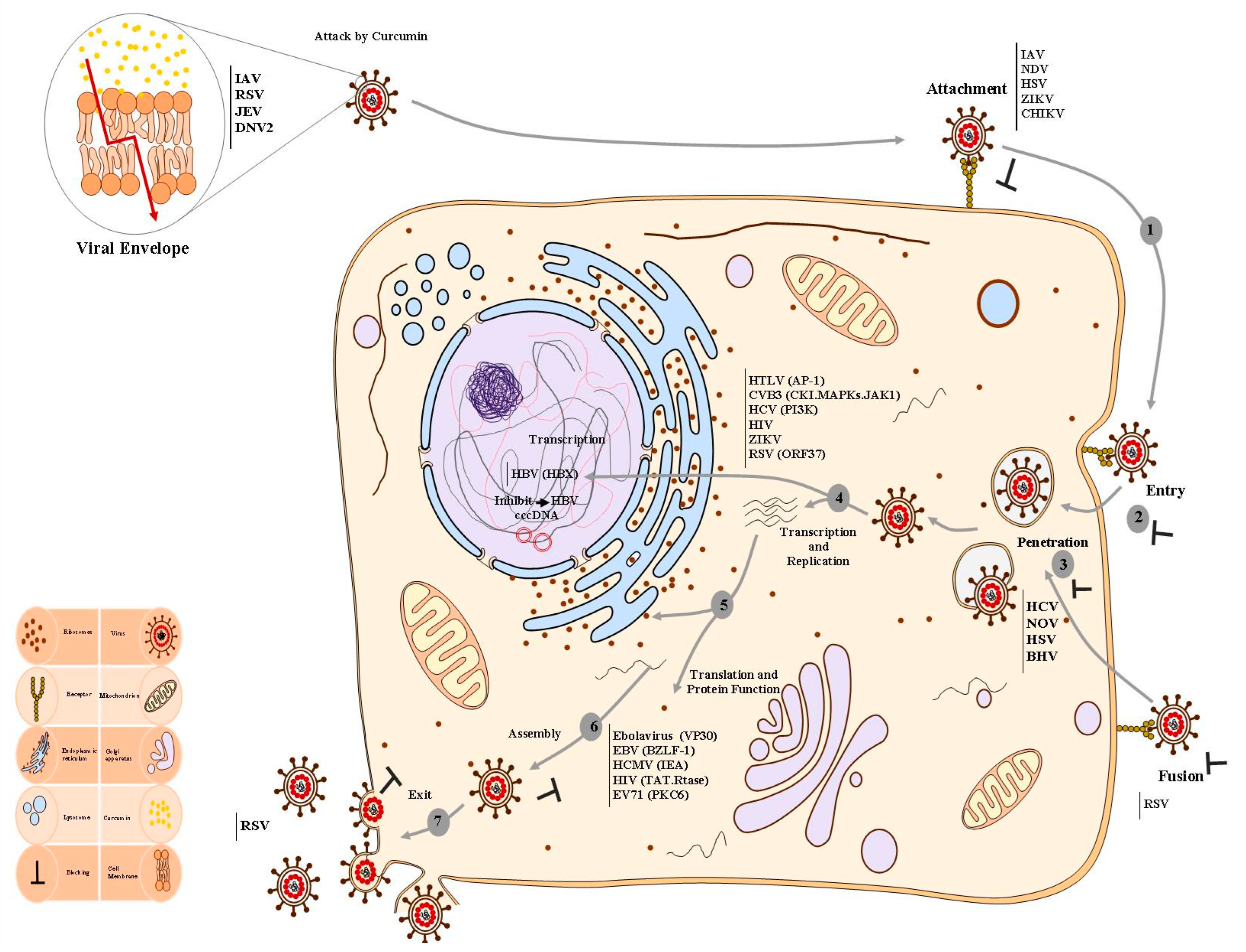
3. Selected Studies
4. Antiviral Activity of Curcumin against RNA Viruses
4.1. HCV
4.2. Zika Virus
4.3. Dengue Virus
4.4. JEV
4.5. RSV
4.6. EV71
4.7. IFVA
4.8. HIV
4.9. Coxsackievirus
4.10. VSV
4.11. Coronavirus
4.12. Norovirus
4.13. Human Parainfluenza Virus Type 3
5. Antiviral Activity of Curcumin against DNA Viruses
5.1. HBV
5.2. Adenovirus
5.3. HSV
5.4. HCMV
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Available online: http://www.emro.who.int/health-topics.html (accessed on 7 November 2021).
- Lorestani, N.; Moradi, A.; Teimoori, A.; Masodi, M.; Khanizadeh, S.; Hassanpour, M.; Javid, N.; Ardebili, A.; Tabarraei, A.; Nikoo, H.R. Molecular and serologic characterization of rotavirus from children with acute gastroenteritis in northern Iran, Gorgan. Bmc Gastroenterol. 2019, 19, 100. [Google Scholar] [CrossRef] [Green Version]
- Zorofchian Moghadamtousi, S.; Abdul Kadir, H.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. Biomed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef] [PubMed]
- Chesney, M.A. Factors affecting adherence to antiretroviral therapy. Clin. Infect. Dis. 2000, 30, S171–S176. [Google Scholar] [CrossRef] [PubMed]
- Bolken, T.C.; Hruby, D.E. Discovery and development of antiviral drugs for biodefense: Experience of a small biotechnology company. Antivir. Res. 2008, 77, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G. Treatment of antiretroviral-drug-resistant HIV-1 infection. Lancet 2003, 362, 2002–2011. [Google Scholar] [CrossRef]
- Parry, J. Use of antiviral drug in poultry is blamed for drug resistant strains of avian flu. BMJ 2005, 331, 10. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, P.; Koch, C.; Reichling, J. Susceptibility of drug-resistant clinical herpes simplex virus type 1 strains to essential oils of ginger, thyme, hyssop, and sandalwood. Antimicrob. Agents Chemother. 2007, 51, 1859–1862. [Google Scholar] [CrossRef] [Green Version]
- Meuss, A. Herbal medicine. Curr. Sci. 2000, 78, 35–39. [Google Scholar]
- Connell, B.J.; Chang, S.-Y.; Prakash, E.; Yousfi, R.; Mohan, V.; Posch, W.; Wilflingseder, D.; Moog, C.; Kodama, E.N.; Clayette, P. A cinnamon-derived procyanidin compound displays anti-hiv-1 activity by blocking heparan sulfate-and co-receptor-binding sites on gp120 and reverses T cell exhaustion via impeding tim-3 and PD-1 upregulation. PLoS ONE 2016, 11, e0165386. [Google Scholar] [CrossRef]
- Steinmann, J.; Buer, J.; Pietschmann, T.; Steinmann, E. Anti-infective properties of epigallocatechin-3-gallate (EGCG), a component of green tea. Br. J. Pharmacol. 2013, 168, 1059–1073. [Google Scholar] [CrossRef] [Green Version]
- Praditya, D.; Kirchhoff, L.; Brüning, J.; Rachmawati, H.; Steinmann, J.; Steinmann, E. Anti-infective properties of the golden spice curcumin. Front. Microbiol. 2019, 10, 912. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, I.; Biswas, K.; Bandyopadhyay, U.; Banerjee, R.K. Turmeric and curcumin: Biological actions and medicinal applications. Curr. Sci. 2004, 87, 44–53. [Google Scholar]
- Mathew, D.; Hsu, W.-L. Antiviral potential of curcumin. J. Funct. Foods 2018, 40, 692–699. [Google Scholar] [CrossRef]
- Lal, J. Turmeric, curcumin and our life:A review. Bull Env. Pharm. Life Sci 2012, 1, 11–17. [Google Scholar]
- Péret-Almeida, L.; Cherubino, A.; Alves, R.; Dufossé, L.; Glória, M. Separation and determination of the physico-chemical characteristics of curcumin, demethoxycurcumin and bisdemethoxycurcumin. Food Res. Int. 2005, 38, 1039–1044. [Google Scholar] [CrossRef]
- Qin, Y.; Lin, L.; Chen, Y.; Wu, S.; Si, X.; Wu, H.; Zhai, X.; Wang, Y.; Tong, L.; Pan, B. Curcumin inhibits the replication of enterovirus 71 in vitro. Acta Pharm. Sin. B 2014, 4, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Banerjea, A.C. Curcumin inhibits HIV-1 by promoting Tat protein degradation. Sci Rep 2016, 6, 27539. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.; Raghavan, K.; Weinstein, J.; Kohn, K.W.; Pommier, Y. Inhibition of human immunodeficiency virus type-1 integrase by curcumin. Biochem. Pharmacol. 1995, 49, 1165–1170. [Google Scholar] [CrossRef]
- Barthelemy, S.; Vergnes, L.; Moynier, M.; Guyot, D.; Labidalle, S.; Bahraoui, E. Curcumin and curcumin derivatives inhibit Tat-mediated transactivation of type 1 human immunodeficiency virus long terminal repeat. Res. Virol. 1998, 149, 43–52. [Google Scholar] [CrossRef]
- Tomita, M.; Kawakami, H.; Uchihara, J.-N.; Okudaira, T.; Masuda, M.; Takasu, N.; Matsuda, T.; Ohta, T.; Tanaka, Y.; Mori, N. RETRACTED: Curcumin Suppresses Constitutive Activation of AP-1 by Downregulation of JunD Protein in HTLV-1-Infected T-Cell Lines; Elsevier: New York, NY, USA, 2006. [Google Scholar]
- Si, X.; Wang, Y.; Wong, J.; Zhang, J.; McManus, B.M.; Luo, H. Dysregulation of the ubiquitin-proteasome system by curcumin suppresses coxsackievirus B3 replication. J. Virol. 2007, 81, 3142–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutluay, S.B.; Doroghazi, J.; Roemer, M.E.; Triezenberg, S.J. Curcumin inhibits herpes simplex virus immediate-early gene expression by a mechanism independent of p300/CBP histone acetyltransferase activity. Virology 2008, 373, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Yoo, H.S.; Kim, J.C.; Park, C.S.; Choi, M.S.; Kim, M.; Choi, H.; Min, J.S.; Kim, Y.S.; Yoon, S.W. Antiviral effect of Curcuma longa Linn extract against hepatitis B virus replication. J. Ethnopharmacol. 2009, 124, 189–196. [Google Scholar] [CrossRef]
- Dutta, K.; Ghosh, D.; Basu, A. Curcumin protects neuronal cells from Japanese encephalitis virus-mediated cell death and also inhibits infective viral particle formation by dysregulation of ubiquitin–proteasome system. J. Neuroimmune Pharmacol. 2009, 4, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Rechtman, M.M.; Har-Noy, O.; Bar-Yishay, I.; Fishman, S.; Adamovich, Y.; Shaul, Y.; Halpern, Z.; Shlomai, A. Curcumin inhibits hepatitis B virus via down-regulation of the metabolic coactivator PGC-1α. Febs Lett. 2010, 584, 2485–2490. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.-Y.; Shien, J.-H.; Tiley, L.; Chiou, S.-S.; Wang, S.-Y.; Chang, T.-J.; Lee, Y.-J.; Chan, K.-W.; Hsu, W.-L. Curcumin inhibits influenza virus infection and haemagglutination activity. Food Chem. 2010, 119, 1346–1351. [Google Scholar] [CrossRef]
- Zandi, K.; Ramedani, E.; Mohammadi, K.; Tajbakhsh, S.; Deilami, I.; Rastian, Z.; Fouladvand, M.; Yousefi, F.; Farshadpour, F. Evaluation of antiviral activities of curcumin derivatives against HSV-1 in Vero cell line. Nat. Prod. Commun. 2010, 5, 1934578X1000501220. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Kim, K.H.; Kim, H.Y.; Cho, H.K.; Sakamoto, N.; Cheong, J. Curcumin inhibits hepatitis C virus replication via suppressing the Akt-SREBP-1 pathway. Febs Lett. 2010, 584, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Ruan, Z.; Sang, W.W. HDAC1/NFκB pathway is involved in curcumin inhibiting of Tat-mediated long terminal repeat transactivation. J. Cell. Physiol. 2011, 226, 3385–3391. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.-L.; Lan, A.-J.; Lan, Y.-Y.; Chao, L.; Ning, L.; Cheng, W.; Liu, L.-H. Activity of curcumin against human cytomegalovirus in vitro. Afr. J. Pharm. Pharmacol. 2012, 6, 30–35. [Google Scholar]
- Chen, M.-H.; Lee, M.-Y.; Chuang, J.-J.; Li, Y.-Z.; Ning, S.-T.; Chen, J.-C.; Liu, Y.-W. Curcumin inhibits HCV replication by induction of heme oxygenase-1 and suppression of AKT. Int. J. Mol. Med. 2012, 30, 1021–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanan, A.; Kehn-Hall, K.; Senina, S.; Lundberg, L.; Van Duyne, R.; Guendel, I.; Das, R.; Baer, A.; Bethel, L.; Turell, M. Curcumin inhibits Rift Valley fever virus replication in human cells. J. Biol. Chem. 2012, 287, 33198–33214. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-Y.; Chen, D.-Y.; Wen, H.-W.; Ou, J.-L.; Chiou, S.-S.; Chen, J.-M.; Wong, M.-L.; Hsu, W.-L. Inhibition of enveloped viruses infectivity by curcumin. PLoS ONE 2013, 8, e62482. [Google Scholar]
- Obata, K.; Kojima, T.; Masaki, T.; Okabayashi, T.; Yokota, S.; Hirakawa, S.; Nomura, K.; Takasawa, A.; Murata, M.; Tanaka, S. Curcumin prevents replication of respiratory syncytial virus and the epithelial responses to it in human nasal epithelial cells. PLoS ONE 2013, 8, e70225. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; An, Z.; Chen, H.; Wang, Z.; Liu, L. Mechanism of curcumin resistance to human cytomegalovirus in HELF cells. Bmc Complementary Altern. Med. 2014, 14, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Y.; Lei, N.; Wang, D.; An, Z.; Li, G.; Han, F.; Liu, H.; Liu, L. Protective effect of curcumin against cytomegalovirus infection in Balb/c mice. Environ. Toxicol. Pharmacol. 2014, 37, 1140–1147. [Google Scholar] [CrossRef]
- Zhu, L.; Ding, X.; Zhang, D.; Ch, Y.; Wang, J.; Ndegwa, E.; Zhu, G. Curcumin inhibits bovine herpesvirus type 1 entry into MDBK cells. Acta Virol. 2015, 59, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Colpitts, C.C.; Schang, L.M.; Rachmawati, H.; Frentzen, A.; Pfaender, S.; Behrendt, P.; Brown, R.J.; Bankwitz, D.; Steinmann, J.; Ott, M. Turmeric curcumin inhibits entry of all hepatitis C virus genotypes into human liver cells. Gut 2014, 63, 1137–1149. [Google Scholar]
- Ferreira, V.H.; Nazli, A.; Dizzell, S.E.; Mueller, K.; Kaushic, C. The anti-inflammatory activity of curcumin protects the genital mucosal epithelial barrier from disruption and blocks replication of HIV-1 and HSV-2. PLoS ONE 2015, 10, e0124903. [Google Scholar]
- Jeong, E.-H.; Vaidya, B.; Cho, S.-Y.; Park, M.-A.; Kaewintajuk, K.; Kim, S.R.; Oh, M.-J.; Choi, J.-S.; Kwon, J.; Kim, D. Identification of regulators of the early stage of viral hemorrhagic septicemia virus infection during curcumin treatment. Fish Shellfish Immunol. 2015, 45, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Kulkarni, A.A.; Lin, X.; McLean, C.; Ammosova, T.; Ivanov, A.; Hipolito, M.; Nekhai, S.; Nwulia, E. Inhibition of HIV-1 by curcumin A, a novel curcumin analog. Drug Des. Dev. Ther. 2015, 9, 5051. [Google Scholar]
- Flores, D.J.; Lee, L.H.; Adams, S.D. Inhibition of curcumin-treated Herpes Simplex virus 1 and 2 in vero cells. Adv. Microbiol. 2016, 6, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, W.; Aznar, R.; Sánchez, G. Curcumin-mediated photodynamic inactivation of norovirus surrogates. Food Environ. Virol. 2016, 8, 244–250. [Google Scholar] [CrossRef] [PubMed]
- von Rhein, C.; Weidner, T.; Henß, L.; Martin, J.; Weber, C.; Sliva, K.; Schnierle, B.S. Curcumin and Boswellia serrata gum resin extract inhibit chikungunya and vesicular stomatitis virus infections in vitro. Antivir. Res. 2016, 125, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Naseri, S.; Darroudi, M.; Aryan, E.; Gholoobi, A.; Rahimi, H.R.; Ketabi, K.; Movaqar, A.; Abdoli, M.; Gouklani, H.; Teimourpour, R. The antiviral effects of curcumin nanomicelles on the attachment and entry of hepatitis C virus. Iran. J. Virol. 2017, 11, 29–35. [Google Scholar]
- Ahmed, J.; Tan, Y.; Ambegaokar, S. Effects of Curcumin on Vesicular Stomatitis Virus (VSV) Infection and Dicer-1 Expression. Faseb J. 2017, 31, 622-11. [Google Scholar]
- Mounce, B.C.; Cesaro, T.; Carrau, L.; Vallet, T.; Vignuzzi, M. Curcumin inhibits Zika and chikungunya virus infection by inhibiting cell binding. Antivir. Res. 2017, 142, 148–157. [Google Scholar] [CrossRef]
- Wei, Z.-Q.; Zhang, Y.-H.; Ke, C.-Z.; Chen, H.-X.; Ren, P.; He, Y.-L.; Hu, P.; Ma, D.-Q.; Luo, J.; Meng, Z.-J. Curcumin inhibits hepatitis B virus infection by down-regulating cccDNA-bound histone acetylation. World, J. Gastroenterol. 2017, 23, 6252. [Google Scholar] [CrossRef]
- Sharma, R.K.; Cwiklinski, K.; Aalinkeel, R.; Reynolds, J.L.; Sykes, D.E.; Quaye, E.; Oh, J.; Mahajan, S.D.; Schwartz, S.A. Immunomodulatory activities of curcumin-stabilized silver nanoparticles:Efficacy as an antiretroviral therapeutic. Immunol. Investig. 2017, 46, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-I.; Chio, C.-C.; Lin, J.-Y. Inhibition of EV71 by curcumin in intestinal epithelial cells. PLoS ONE 2018, 13, e0191617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poursina, Z.; Mohammadi, A.; Yazdi, S.Z.; Humpson, I.; Vakili, V.; Boostani, R.; Rafatpanah, H. Curcumin increased the expression of c-FLIP in HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) patients. J. Cell. Biochem. 2019, 120, 15740–15745. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Pilankatta, R.; Teramoto, T.; Sajith, A.M.; Nwulia, E.; Kulkarni, A.; Padmanabhan, R. Inhibition of dengue virus by curcuminoids. Antivir. Res. 2019, 162, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhong, C.; Wang, Q.; Chen, W.; Yuan, Y. Curcumin is an APE1 redox inhibitor and exhibits an antiviral activity against KSHV replication and pathogenesis. Antivir. Res. 2019, 167, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Yadav, A.; Gupta, N.; Sharma, S.; Kakkar, R.; Cwiklinski, K.; Quaye, E.; Mahajan, S.D.; Schwartz, S.A.; Sharma, R.K. Multifunctional mesoporous curcumin encapsulated iron-phenanthroline nanocluster: A new Anti-HIV agent. Colloids Surf. B Biointerfaces 2019, 180, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Nabila, N.; Suada, N.K.; Denis, D.; Yohan, B.; Adi, A.C.; Veterini, A.S.; Anindya, A.L.; Sasmono, R.T.; Rachmawati, H. Antiviral action of curcumin encapsulated in nanoemulsion against four serotypes of dengue virus. Pharm. Nanotechnol. 2020, 8, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Liu, Y.; Luo, X.; Lei, W.; Xie, L. Antiviral and virucidal effects of curcumin on transmissible gastroenteritis virus in vitro. J. Gen. Virol. 2020, 101, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, K.; Zang, G.; Chen, T.; Lu, N.; Wang, S.; Zhang, G. Curcumin Inhibits Replication of Human Parainfluenza Virus Type 3 by Affecting Viral Inclusion Body Formation. Biomed Res. Int. 2021, 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Thongsri, P.; Pewkliang, Y.; Borwornpinyo, S.; Wongkajornsilp, A.; Hongeng, S.; Sa-Ngiamsuntorn, K. Curcumin inhibited hepatitis B viral entry through NTCP binding. Sci. Rep. 2021, 11, 19125. [Google Scholar] [CrossRef] [PubMed]
- Halim, J.A.N.; Halim, S.N.; Denis, D.; Haryanto, S.; Dharmana, E.; Hapsari, R.; Sasmono, R.T.; Yohan, B. Antiviral activities of curcumin and 6-gingerol against infection of four dengue virus serotypes in A549 human cell line in vitro. Indones. J. Biotechnol. 2021, 26, 41–47. [Google Scholar] [CrossRef]
- Wahyuni, T.S.; Permatasari, A.A.; Widiandani, T.; Fuad, A.; Widyawaruyanti, A.; Aoki-Utsubo, C.; Hotta, H. Antiviral activities of curcuma genus against hepatitis C virus. Nat. Prod. Commun. 2018, 13, 1934578X1801301204. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Tai, W.; Wang, N.; Li, X.; Jiang, S.; Debnath, A.K.; Du, L.; Chen, S. Identification of novel natural products as effective and broad-spectrum anti-Zika virus inhibitors. Viruses 2019, 11, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key gaps in the knowledge of the porcine respiratory reproductive syndrome virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, F.; Dudziak, D.; Dirmeier, U.; Hobom, G.; Riedel, A.; Schlee, M.; Staudt, L.M.; Rosenwald, A.; Behrends, U.; Bornkamm, G.W. Active NF-κB signalling is a prerequisite for influenza virus infection. J. Gen. Virol. 2004, 85, 2347–2356. [Google Scholar] [CrossRef]
- Lai, Y.; Yan, Y.; Liao, S.; Li, Y.; Ye, Y.; Liu, N.; Zhao, F.; Xu, P. 3D-quantitative structure–activity relationship and antiviral effects of curcumin derivatives as potent inhibitors of influenza H1N1 neuraminidase. Arch. Pharmacal Res. 2020, 43, 489–502. [Google Scholar] [CrossRef]
- Song, Y.; Ge, W.; Cai, H.; Zhang, H. Curcumin protects mice from coxsackievirus B3-induced myocarditis by inhibiting the phosphatidylinositol 3 kinase/Akt/nuclear factor-κB pathway. J. Cardiovasc. Pharmacol. Ther. 2013, 18, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Sobhanimonfared, F.; Bamdad, T.; Sadigh, Z.; Nikoo, H.; Choobin, H. Combination of virotherapy with VSV and tumor vaccination significantly enhances the efficacy of antitumor therapy. Acta Virol. 2018, 62, 394–400. [Google Scholar] [CrossRef]
- Zahedipour, F.; Hosseini, S.A.; Sathyapalan, T.; Majeed, M.; Jamialahmadi, T.; Al-Rasadi, K.; Banach, M.; Sahebkar, A. Potential effects of curcumin in the treatment of COVID-19 infection. Phytother. Res. 2020, 34, 2911–2920. [Google Scholar] [CrossRef]
- Taghizadeh, P.; Salehi, S.; Heshmati, A.; Houshmand, S.M.; InanlooRahatloo, K.; Mahjoubi, F.; Sanati, M.H.; Yari, H.; Alavi, A.; Jamehdar, S.A. Study on SARS-CoV-2 strains in Iran reveals potential contribution of co-infection with and recombination between different strains to the emergence of new strains. Virology 2021, 526, 63–73. [Google Scholar] [CrossRef]
- Maurya, V.K.; Kumar, S.; Prasad, A.K.; Bhatt, M.L.; Saxena, S.K. Structure-based drug designing for potential antiviral activity of selected natural products from Ayurveda against SARS-CoV-2 spike glycoprotein and its cellular receptor. Virusdisease 2020, 31, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kashyap, P.; Chowdhury, S.; Kumar, S.; Panwar, A.; Kumar, A. Identification of phytochemicals as potential therapeutic agents that binds to Nsp15 protein target of coronavirus (SARS-CoV-2) that are capable of inhibiting virus replication. Phytomedicine 2021, 85, 153317. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, K.; Varakumar, P.; Baliwada, A.; Byran, G. Activity of phytochemical constituents of Curcuma longa (turmeric) and Andrographis paniculata against coronavirus (COVID-19): An in silico approach. Future J. Pharm. Sci. 2020, 6, 104. [Google Scholar] [CrossRef]
- Yang, M.; Lee, G.; Si, J.; Lee, S.-J.; You, H.J.; Ko, G. Curcumin shows antiviral properties against norovirus. Molecules 2016, 21, 1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, A. Functions of adenovirus E1A. Cancer Surv. 1986, 5, 367–387. [Google Scholar] [PubMed]
- Jennings, M.R.; Parks, R.J. Antiviral Effects of Curcumin on Adenovirus Replication. Microorganisms 2020, 8, 1524. [Google Scholar] [CrossRef]
- Vitali, D.; Bagri, P.; Wessels, J.M.; Arora, M.; Ganugula, R.; Parikh, A.; Mandur, T.; Felker, A.; Garg, S.; Kumar, M. Curcumin can decrease tissue inflammation and the severity of HSV-2 infection in the female reproductive mucosa. Int. J. Mol. Sci. 2020, 21, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingolfsson, H.I.; Koeppe, R.E.; Andersen, O.S. Curcumin is a modulator of bilayer material properties. Biochemistry 2007, 46, 10384–10391. [Google Scholar] [CrossRef]
- Wu, J.; Hou, W.; Cao, B.; Zuo, T.; Xue, C.; Leung, A.W.; Xu, C.; Tang, Q.-J. Virucidal efficacy of treatment with photodynamically activated curcumin on murine norovirus bio-accumulated in oysters. Photodiagnosis Photodyn. Ther. 2015, 12, 385–392. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
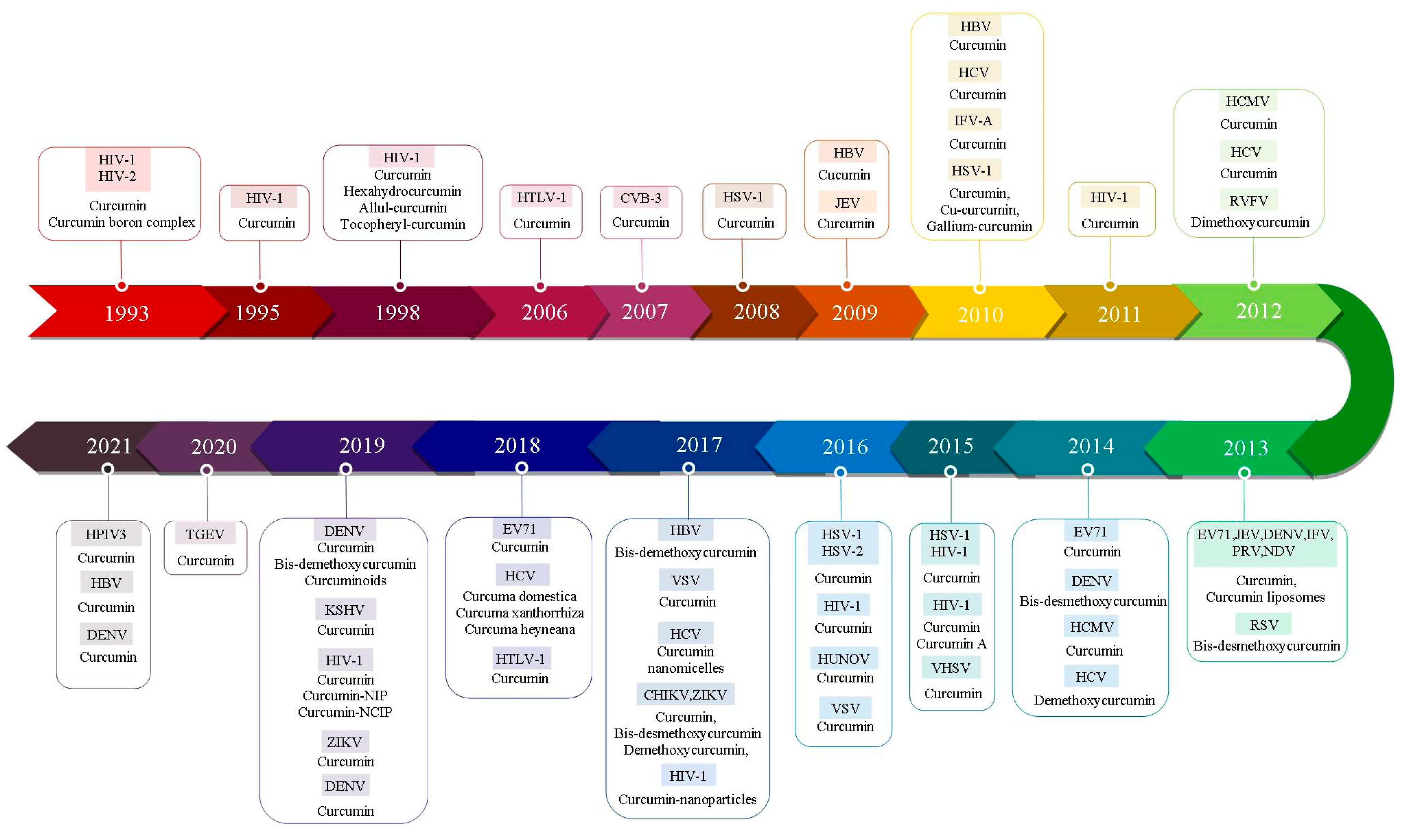
| Year | Virus | Study Scope | Substance | CC50 1 | IC50 2 | SI 3 | Method | Mechanism of Action | Ref |
|---|---|---|---|---|---|---|---|---|---|
| 1993 | Human immunodeficiency virus 1 and 2 (HIV-1 and -2) | Evaluation of curcumin and curcumin boron complexes on the HIV-1 and HIV-2 proteases | Curcumin, Curcumin boron complex | 1–500 μM | HIV1 (100 μM), HIV2 (250 μM) | ND 4 | HPLC 5, SDS-PAGE 6, Fluorescent test | Inhibition of viral proteases | [18] |
| 1995 | Human immunodeficiency virus 1 (HIV-1) | Evaluation of curcumin on HIV-1 integrase | Curcumin | 10–100 μM | 40 μM | ND | PAGE | Inhibition of HIV-1 integrase | [19] |
| 1998 | Human immunodeficiency virus 1 (HIV-1) | Determination of curcumin and curcumin derivatives activity on HIV-1 Tat protein | Curcumin, Hexahydrocurcumin, Allyl-curcumin, Tocopheryl-curcumin | 1 μM, 5 μM, 100 μM | ND | ND | Trypanblue, Oշ-assay | Reduction of Tat-mediated HIV transcription, leading to inhibition of replication | [20] |
| 2006 | Human T lymphotropic virus 1 (HTLV-1) | Determination of curcumin effect on AP-1 in HTLV-1 | Curcumin | 50 μM | ND | ND | Western blot | Inhibition of the constitutive AP-1 7 activity and viral transcription by downregulation of JunD protein | [21] |
| 2007 | Coxackivirus B3 (CVB3) | Evaluation of curcumin on replication oxsackievirus B3 | Curcumin | 30 μM | ND | ND | Plaque assay, Western blot | Reduction of CVB3 replication by inhibition of intracellular signaling pathways, including MAPKs 8, CKII, and Jab1 9 | [22] |
| 2008 | Herpes simplex virus-1 (HSV-1) | Evaluation of curcumin on herpes simplex virus immediate-early gene expression | Curcumin | 20 μM | 25 μM | ND | Plaque assay, PCR 10 Western blot, Real-time PCR | Interference of VP-16 mediated recruitment of RNA polymerase II to immediate-early gene promoters | [23] |
| 2009 | Hepatitis B virus (HBV) | Determination of antiviral effect of Curcuma longa Linn extract on hepatitis B virus replication | Curcumin | 200 μg/L, 500 μM | ND | ND | MTT 11, Southern blot, RT-PCR 12, Western blot | Enhancing the cellular accumulation of p53 protein, and repression of the HBx gene replication and transcription process | [24] |
| 2009 | Japanese encephalitis virus (JEV) | Determination of curcumin activity on Japanese encephalitis virus infectivity | Curcumin | 5 μM, 10 μM | Dose-dependent | Dose-dependent | MTS 13, Plaque assay | Decreases ubiquitin proteasome system, causing the reduction of infective viral particle production | [25] |
| 2010 | Hepatitis B virus (HBV) | Evaluation of curcumin on hepatitis B virus replication | Curcumin | 50–150 μM | Dose-dependent | Dose-dependent | Western blot | Suppression of HBV expression in a PGC-1a 14 dependent manner | [26] |
| 2010 | Influenza virus (IFV-A) | Evaluation of curcumin on influenza virus infection and hemagglutination | Curcumin | 43 μM | 0.47 μM | 92.5 μM | Plaque assay, Western blot, HI 15 | Interruption of virus attachment | [27] |
| 2010 | Herpes simplex virus-1 (HSV-1) | Evaluation of antiviral activities of curcumin derivatives against HSV-1 | Curcumin, Gallium-curcumin, Cu-curcumin | 484.2 μM, 255.8 μM, 326.6 μM | 33.0 μM, 13.9 μM, 23.1 μM | 14.6 μM, 18.4 μM, 14.1 μM | TCID50 16 Trypanblue | Antiviral effects on HSV-1 in cell culture. | [28] |
| 2010 | Hepatitis C virus (HCV) | Evaluation of curcumin activity on the replication of hepatitis C virus | Curcumin | 5–15 μM | Dose-dependent | Dose-dependent | MTT assay RT-PCR | Inhibition of HCV replication via the PI3K 17/Akt and SREBP-1-pathway, not NF-kB 18 pathway | [29] |
| 2011 | Human immunodeficiency virus 1 (HIV-1) | Determination of curcumin effect on HDAC1/NFκB in HTLV-1 | Curcumin | ND | ND | ND | MTT | Inhibition of Tat-regulated transcription, by targeting cellular factors such as AMPK/HDAC1/NFκB. | [30] |
| 2012 | Human cytomegalovirus (HCMV or HHV5) | Evaluation of in vitro activity of curcumin on HCMV | Curcumin | 25 μM, 10 μM, 1 μM | 10 | ND | MTT, TCID50 | Decreases viral DNA and apoptosis in the infected cells | [31] |
| 2012 | Hepatitis C virus (HCV) | Evaluation of curcumin on HCV replication | Curcumin | 5–25 μM | Does–dependent | ND | MTT, Real-time PCR, Western blot | Inhibition of viral replication by induction of the HO-1 expression and the inhibition of the PI3K-AKT signaling pathway | [32] |
| 2012 | Rift Valley fever virus (RVFV) | Evaluation of curcumin on Rift Valley fever virus replication | Demethoxycurcumin | 10 μM | Time-dependent | ND | Western blot, RT-PCR, Plaque assay | Inhibition of NF-κB transcription factor | [33] |
| 2013 | Enterovirus 71 (EV71), Japanese encephalitis virus (JEV), Dengue virus (DENV), Influenza virus (IFV), Pseudorabies viruses (PRV), Newcastle disease viruses (NDV) | Evaluation of curcumin on enveloped viruses’ infectivity | Curcumin, Curcumin liposomes | 30 μM, 62.5 μM | 4 μM | ND | Plaque assay, HI, MTT | Disruption of the integrity of the viral membrane envelopes and liposomes. | [34] |
| 2013 | Respiratory syncytial virus (RSV) | Evaluation of curcumin on replication of RSV | Bis-desmethoxycurcumin | 5 μM | ND | ND | MTT, Real-time PCR, Western blot, RT-PCR, ELISA 19 | Prevention of viral replication, budding process, and reduction of cell pro-inflammatory responses. Inhibition of NF-κB transcription factor and eIF-2a | [35] |
| 2014 | Enterovirus-71 (EV71) | Evaluation of curcumin on the replication of enterovirus 71 | Curcumin | 40 μM | ND (6 log decrease) | ND | TCID50, RT-PCR, Western blot | Inhibition of viral replication by downregulation of the GBF1 20 and PI4KB 21 in EV71- infected cells. Curcumin suppressed UPS 22 and apoptosis in EV71-infected cells. | [17] |
| 2014 | Human cytomegalovirus (HCMV or HHV5) | Evaluation of curcumin on the cytomegalovirus replication | Curcumin | 0/2–0/8 μg | ND | ND | ELISA, Flow cytometry, Real-time PCR, IF 23, Western blot | Downregulation of the gene expression of HCMV immediate early and UL83 genes by curcumin, causing the reduction of infective viral particle production. | [36] |
| 2014 | Human cytomegalovirus (HCMV or HHV5) | Determination of curcumin antiviral activity against cytomegalovirus infection | Curcumin | 12/5 μM, 25 μM, 50 Μm | 10⁴, 10³ | ND | TCID50, PCR | Decreases the serum levels of AST 24, ALT 25, CK 26, and LDH 27 in the model mice, and liver protection in HCMV-infected mice. | [37] |
| 2014 | Dengue virus (DENV) | Antiviral effects of curcumin on dengue virus type 2-infected cells | Bis-desmethoxycurcumin | 29.5 μM | 11.51 μM | 2.56 | MTT, Western blot | Decreases viral particles by suppression of the ubiquitin-proteasome system. | [38] |
| 2014 | Hepatitis C virus (HCV) | Evaluation of turmeric curcumin on entry of the hepatitis C virus | Demethoxycurcumin | 5–25 μM | Does- dependent | Does-depended | MTT, RT-PCR, TCID50 | Inhibition of viral entry into both hepatoma cell lines and cell-to-cell spread between neighboring cells. Curcumin also did not affect viral assembly/release of both genotypes. | [39] |
| 2015 | Herpes simplex virus 2 (HSV 2), Human immunodeficiency virus 1 (HIV-) | Determination of anti-inflammatory activity of curcumin on HIV-1 and HSV-2 | Curcumin | 5 μM, 50 μM | ND | ND | Trypanblue | Anti-inflammatory properties. Decreases HIV-1 and HSV-2 replication in chronically infected T-cells and primary GECs 28, respectively. | [40] |
| 2015 | Viral hemorrhagic septicemia virus (VHSV) | Antiviral effect of curcumin on VHSV | Curcumin | 15–240 μM | ND | ND | TCID50, CCK-8, Real-time PCR, Western blot | Reduction of infective particle production. Curcumin inhibits entry of viral particles into cells by downregulating FN1 or upregulating F-actin. Curcumin inhibits viral replication by downregulation of HSC71. | [41] |
| 2015 | Human immunodeficiency virus 1 (HIV-1) | Determination of curcumin activity on HIV-1 | Curcumin, Curcumin A | 2 μM | 35 μM, 22 μM, 0.7 μM, 0.8 μM | ND | RT-PCR, Trypanblue | Curcumin and curcumin A might affect an earlier stage of HIV-1 infection and thus indirectly reduce the subsequent HIV-1 transcription step. Both curcumin and curcumin A inhibited early LTR similarly or better than the established HIV-1 inhibitor, AZT. | [42] |
| 2016 | Human immunodeficiency virus 1 (HIV-1) | Determination of curcumin activity on HIV-1 | Curcumin | 20–120 μM | Dose- dependent, Time- dependent | ND | Western blot, RT-PCR | Reduction of Tat protein in infected cells, leading to inhibition of viral replication. | [18] |
| 2016 | Herpes simplex virus1 and 2 (HSV-1 and 2) | Evaluation of curcumin activity on Herpes simplex virus 1 and 2 in | Curcumin | 10–100 μM | 1/8 × 10 7 2/1 × 10 7 | ND | WST-1 assay 29 Plaque assay | Prevention of viral entry into vero cells. Curcumin also did not affect penetration. | [43] |
| 2016 | Human Norovirus (HuNoV) | Antiviral properties of curcumin against Norovirus | Curcumin | 0/25–2 mg/mL | Dose- dependent | ND | WST-1 assay, Plaque assay, Neutralization, Real-time PCR, | Inhibition of entry or other life cycle stages rather than the replication of viral RNA. | [44] |
| 2016 | Vesicular Stomatitis virus (VSV) | Determination of curcumin effects on vesicular stomatitis virus infections | Curcumin | 25–60–100 μmol | ND | ND | Plaque assay, IF | Inhibition of viral entry | [45] |
| 2017 | Hepatitis C virus (HCV) | The antiviral effects of curcumin nanomicelles on Hepatitis C virus | Curcumin, Nanomicelles | 0.256 μM (highest concentration) | 0.1647 mg/mL | ND | MTT, Real-time PCR | Decreases the gene expression of HCV via suppression of the Akt-SREBP-1 activation, not by NF-kB pathway. Curcumin has anti-cancer effects against anti-hepatocellular carcinoma. Inhibition of the attachment and entry of hepatitis C. | [46] |
| 2017 | Vesicular Stomatitis virus (VSV) | Determination of curcumin effects on vesicular stomatitis virus Dicer-1 Expression | Curcumin | 25–60–100 μmol | ND | ND | MTT, Western blot | Antioxidant properties. | [47] |
| 2017 | Chikungunya virus (CHIKV), Zika virus (ZIKV) | Antiviral activity of curcumin against Zika and chikungunya virus | Curcumin, Bisdesmethoxycurcumin Demethoxycurcumin | 11.6 μM, 16.0 μM, 13.2 μM | CHIKV: 3.89 μM 4.84 μM 0.89 μM ZIKV: 1.90 μM 3.61 μM 5.91 μM | ND | Real-time PCR, Plaque assay, Western blot | Disruption of the integrity of the viral membrane envelopes and reduction of the infectivity of viruses in a dose dependent manner. | [48] |
| 2017 | Hepatitis B virus (HBV) | Evaluation of curcumin on the hepatitis B virus replication | Bisdemethoxycurcumin | 5–30 μM | Does-and time- dependent | ND | CCK8 7, Western blot | Inhibition of viral replication via downregulation of cccDNA-bound histone acetylation. | [49] |
| 2017 | Human immunodeficiency virus 1 (HIV-1) | Immunomodulatory activities of curcumin-stabilized silver nanoparticles on HIV-1 | Curcumin- nanoparticles | ND | ND | ND | Inhibition of NF-κB nuclear translocation and the downstream expression of the pro-inflammatory cytokines IL-1β, TNF-α, and IL-6. | [50] | |
| 2018 | Enterovirus 71 (EV71) | Antiviral effects of curcumin on EV71 | Curcumin | 5–50 μM | ND | ND | Western blot Real-time PCR Plaque assay MTT | Inhibition of viral translation and increase of host cell viability. Decreases the phosphorylation of PKCδ 30 and suppression viral translation. | [51] |
| 2018 | Hepatitis C virus (HCV) | Antiviral activities of curcuma genus against Hepatitis C virus | Curcuma domestica Curcuma xanthorrhiza Curcuma heyneana | >100 μM >100 μM >100 μM | 1.68 μM 4.93 μM 5.49 μM | >59.5 >20.3 >18.2 | MTT, Docking | Inhibition of viral entry and interaction with viral proteins. | [41] |
| 2018 | Human T lymphotropic virus 1 (HTLV-1) | Determination of curcumin on the expression of c-FLIP in HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) patients | Curcumin | 80 mg | ND | ND | Real-time PCR | Induction of apoptosis in HTLV-1 infected cells in patients with HAM/TSP 31. | [52] |
| 2019 | Dengue virus (DENV) | The effects of curcuminoids on dengue virus | Curcumin (CC1) Bisdemethoxycurcumin (CC2) Curcuminoids CC4, CC5, [46] | 49.01 μM (CC1) 43.37 μM (CC2) 32.34 μM (CC3) 87.40 μM (CC4) 25.50 μM (CC5) | 66.01 μM (CC1) 36.23 μM (CC2) 39.17 μM (CC3) 43.84 μM (CC4) 60.98 μM (CC5) | 3.21 6.68 12.06 16.27 10.89 | CCK-8 Plaque assay Real-time PCR | Inhibition of viral protease, resulting in suppression of DENV infectivity. | [53] |
| 2019 | Kaposi’s sarcoma-associated herpesvirus (KSHV or HHV8) | Antiviral activity of curcumin against KSHV replication and pathogenesis | Curcumin | 23.56 μM | 8.76 μM | 2.69 | EMSA 8 | Inhibition of APE1, resulting in reduce of the transcription activity of AP-1 and NF-κB. | [54] |
| 2019 | Human immunodeficiency virus 1 (HIV-1) | Multifunctional mesoporous curcumin encapsulated iron phenanthroline nanocluster on HIV-1 | Curcumin Curcumin-NIP 9 Curcumin-NCIP | 1 mg 5 mg/mL 8 mg/mL | ND | ND | CCK-8, IF, Real time-PCR Flow cytometry, MTT | Inhibition of the release of numerous cytokines such as IL1β, IL8, TNFα 32, MCP1 33 and MIP1α 34 in response to viral infection. Anti-inflammatory, anti-oxidative, and anti-HIV effects. Decreased the expression of HIV-p24 by 41%. | [55] |
| 2019 | Zika Virus (ZIKV) | Inhibitory effects of novel natural products against Zika virus | Curcumin | 52.86 μM | 13.67 μM | ND | ELISA, plaque assay | Disruption of the integrity of the viral membrane envelopes and reduce infectivity of viruses in a dose dependent manner. | [46] |
| 2019 | Dengue Virus (DENV) | Antiviral activity of curcumin encapsulated in nanoemulsion against Dengue virus serotypes | Curcumin (Nanocurcumin), | 52.97 μM | ND | ND | MTT | Inhibition of A549 cell proliferation by inducing apoptosis | [56] |
| 2020 | Coronavirus (TGEV) | Antiviral effects of curcumin on transmissible gastroenteritis virus | Curcumin | 78 μM | 8.5 μM | 9 μM | MTT TCID50 Western blot | Inhibitory effects on the adsorption of TGEV and virucidal activity. | [57] |
| 2021 | Human Parainfluenza Virus Type 3 (HPIV3) | Evaluation of Curcumin on Replication of Human Parainfluenza Virus Type 3 | Curcumin | 30 μM | Dose-dependent | ND | CCK-8 Plaque assay Real-time PCR Western blot | Inhibition of viral inclusion body (IB) formation, and virus replication by downregulate cellular PI4KB. | [58] |
| 2021 | Hepatitis B virus (HBV) | Evaluation of Curcumin on viral entry of Hepatitis B | Curcumin | 30 μM | Dose-dependent | ND | MTT Real-time PCR ELISA IF | Interruption of viral entry and suppression of HBV re-infection | [59] |
| 2021 | Dengue Virus (DENV) | Antiviral activity of curcumin against Dengue virus serotypes | Curcumin | 108 μM | Serotypes- dependent | ND | MTT Plaque assay | Inhibition of viral replication | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ardebili, A.; Pouriayevali, M.H.; Aleshikh, S.; Zahani, M.; Ajorloo, M.; Izanloo, A.; Siyadatpanah, A.; Razavi Nikoo, H.; Wilairatana, P.; Coutinho, H.D.M. Antiviral Therapeutic Potential of Curcumin: An Update. Molecules 2021, 26, 6994. https://doi.org/10.3390/molecules26226994
Ardebili A, Pouriayevali MH, Aleshikh S, Zahani M, Ajorloo M, Izanloo A, Siyadatpanah A, Razavi Nikoo H, Wilairatana P, Coutinho HDM. Antiviral Therapeutic Potential of Curcumin: An Update. Molecules. 2021; 26(22):6994. https://doi.org/10.3390/molecules26226994
Chicago/Turabian StyleArdebili, Abdollah, Mohammad Hassan Pouriayevali, Sahar Aleshikh, Marziyeh Zahani, Mehdi Ajorloo, Ahdieh Izanloo, Abolghasem Siyadatpanah, Hadi Razavi Nikoo, Polrat Wilairatana, and Henrique Douglas Melo Coutinho. 2021. "Antiviral Therapeutic Potential of Curcumin: An Update" Molecules 26, no. 22: 6994. https://doi.org/10.3390/molecules26226994