
In Vivo Investigation of the Ameliorating Effect of Tempol against MIA-Induced Knee Osteoarthritis in Rats: Involvement of TGF-β1/SMAD3/NOX4 Cue

 ,
,
Abstract
:
1. Introduction
2. Results
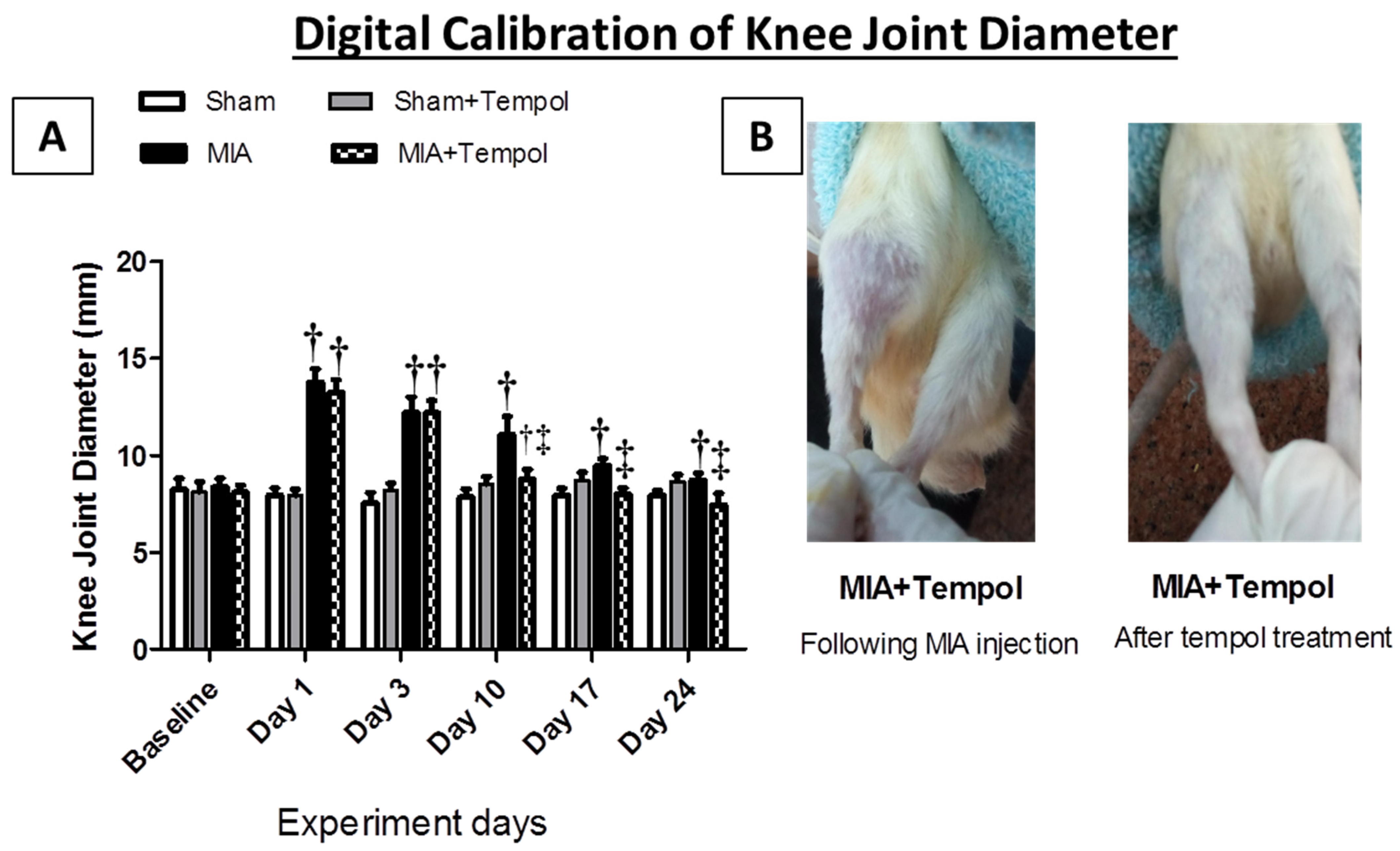
2.1. Effect of Tempol on Knee Joint Swelling
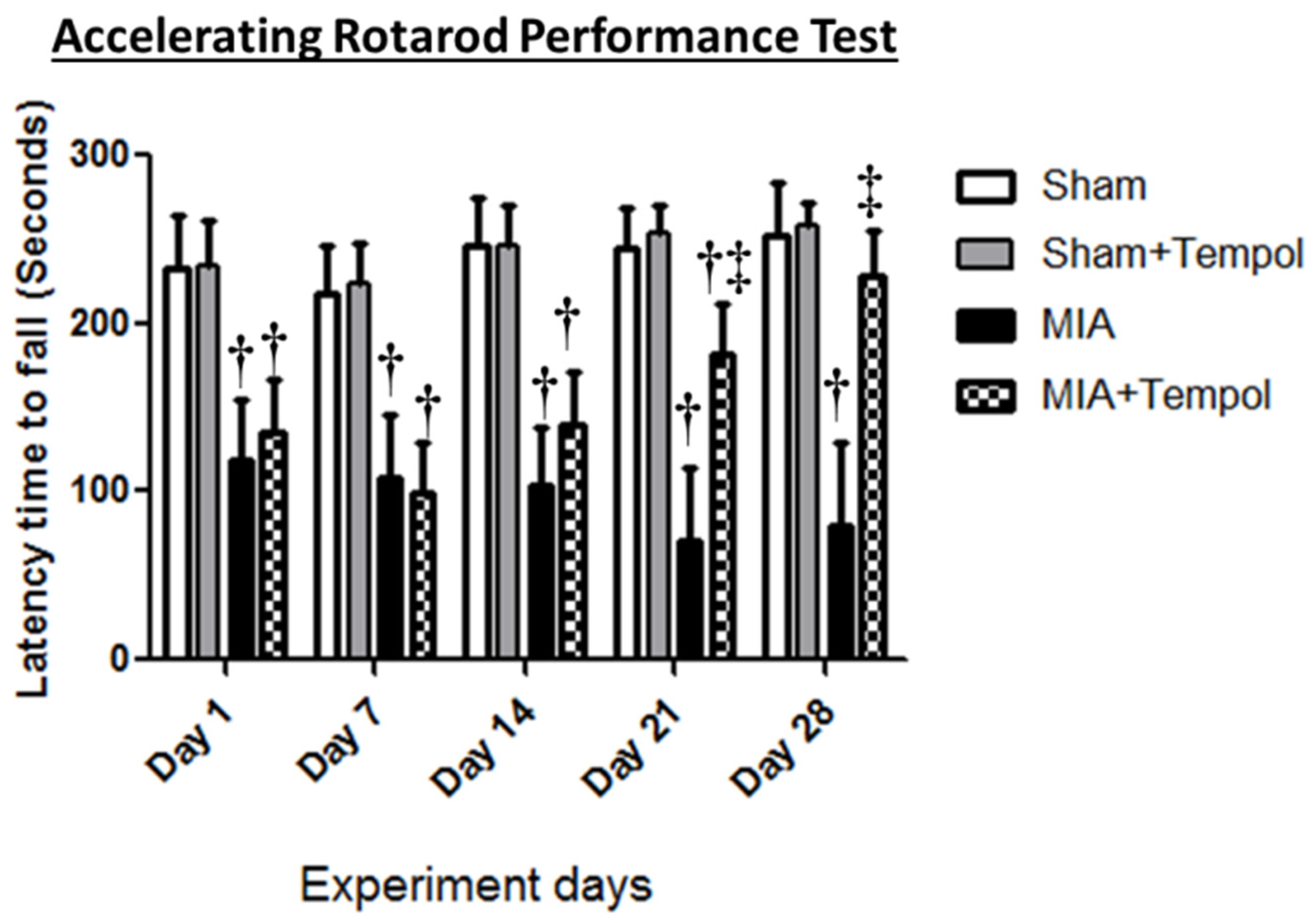
2.2. Effect of Tempol on Rotarod Performance Test
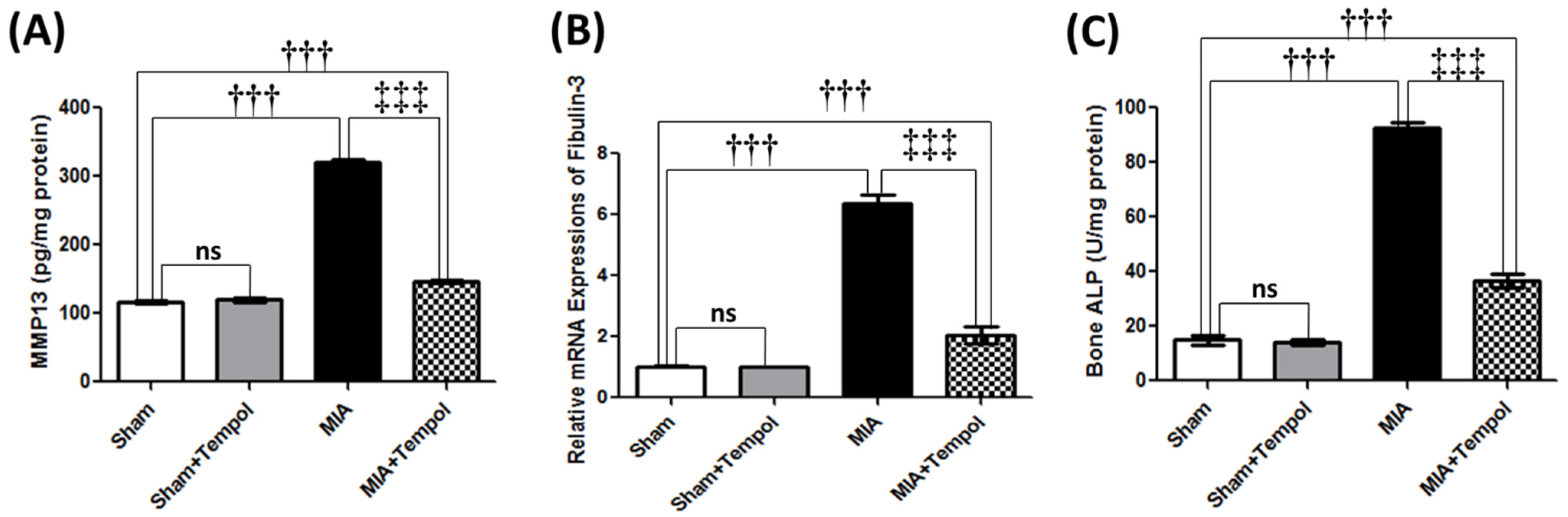
2.3. Effect of Tempol on Knee Joint Degradation-Related Biomarkers
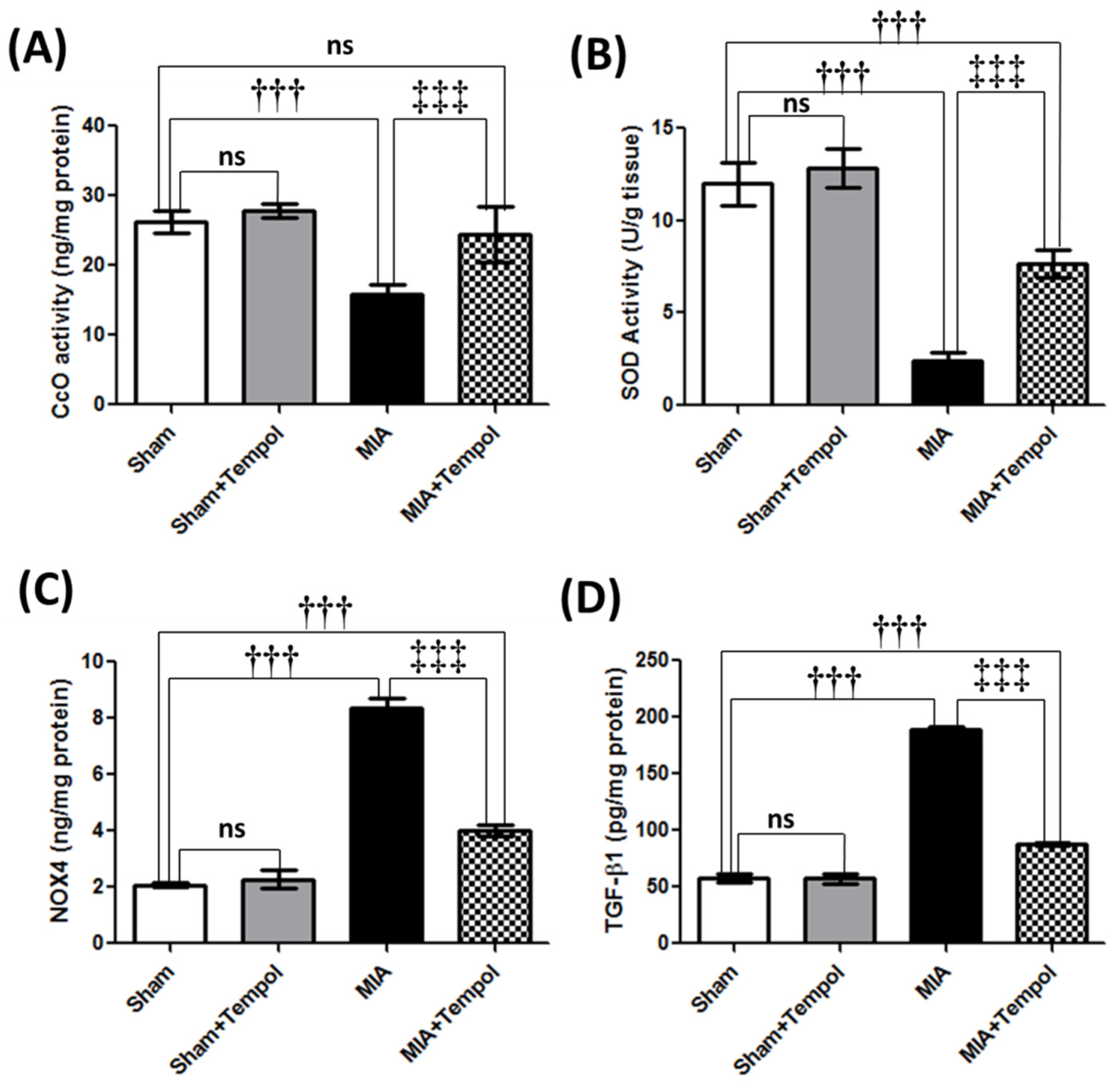
2.4. Effect of Tempol on Mitochondrial Complex IV Oxidase, Oxidative Stress Biomarkers, and TGF-β1
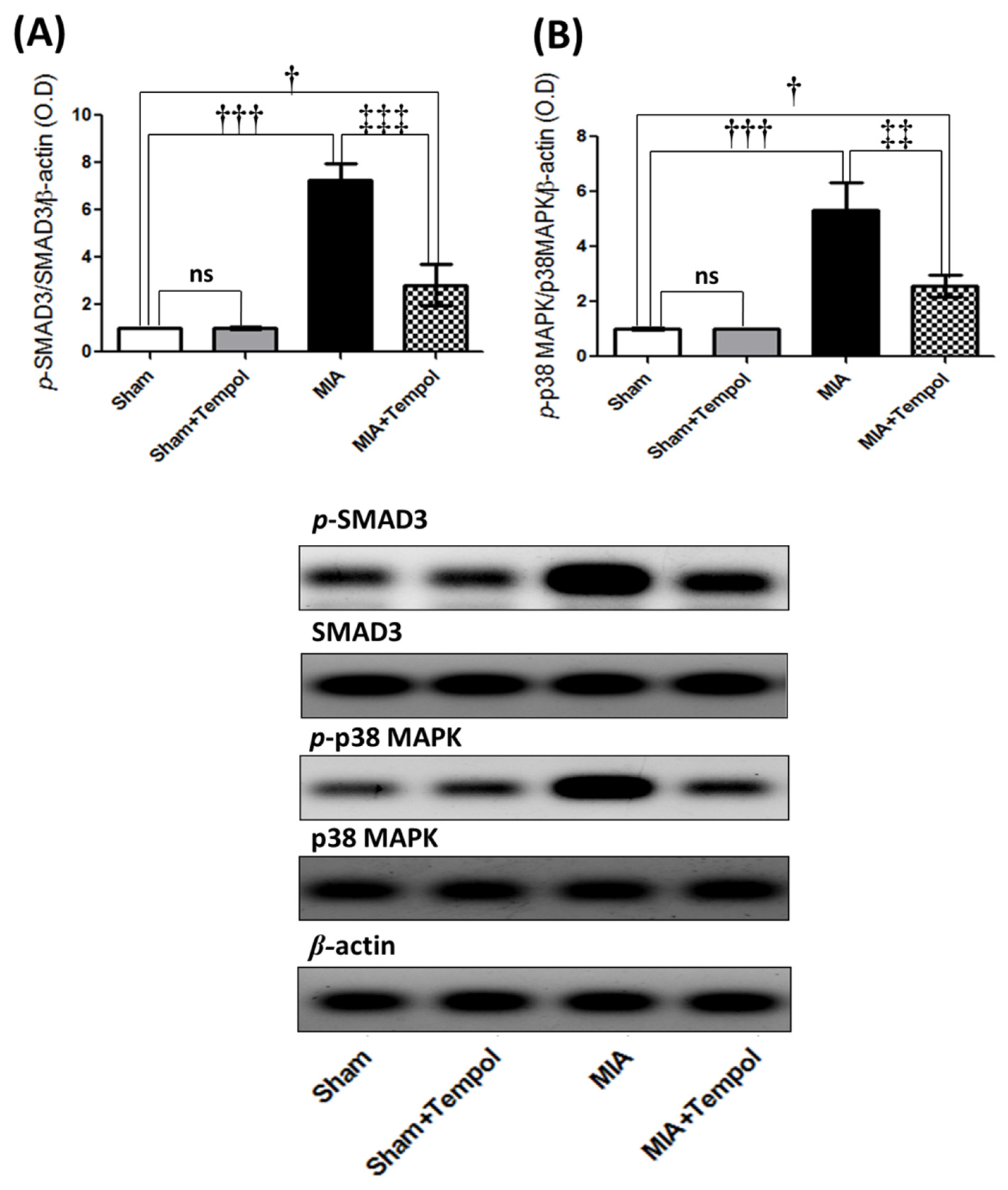
2.5. Effect of Tempol on p-SMAD3, and p-p38MAPK Signaling Proteins
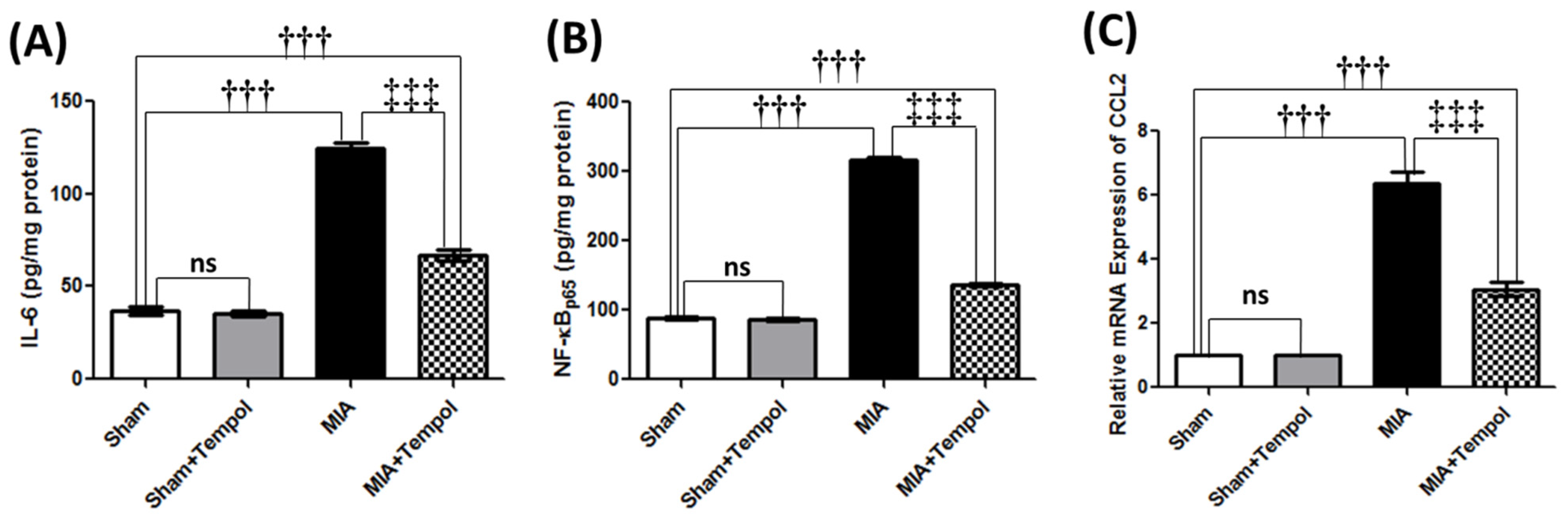
2.6. Effect of Tempol on Inflammatory and Pain Mediators
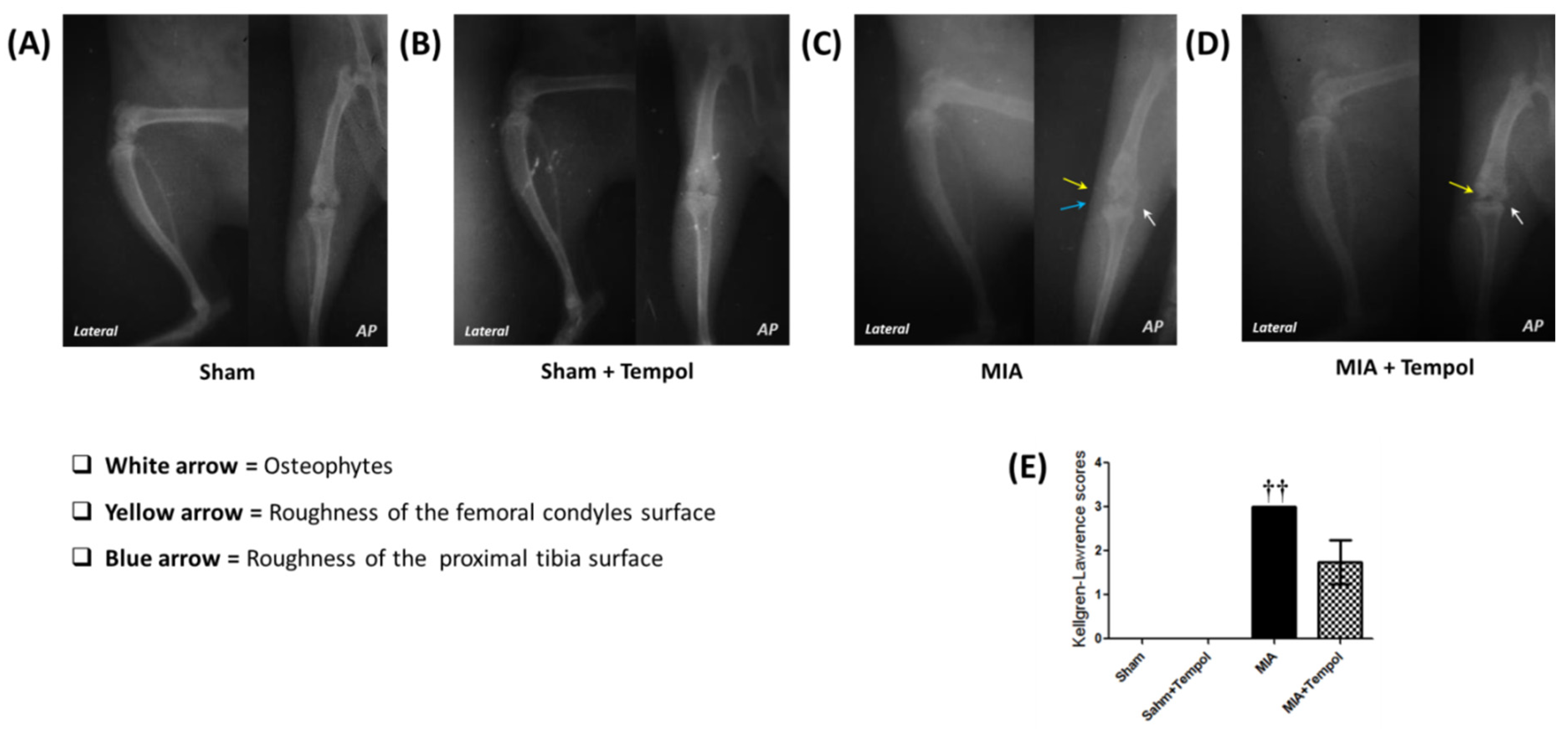
2.7. Effect of Tempol on the Radiographic Changes
2.8. Effect of Tempol on the Histopathological Alterations
2.9. Effect of Tempol on Histochemical Changes
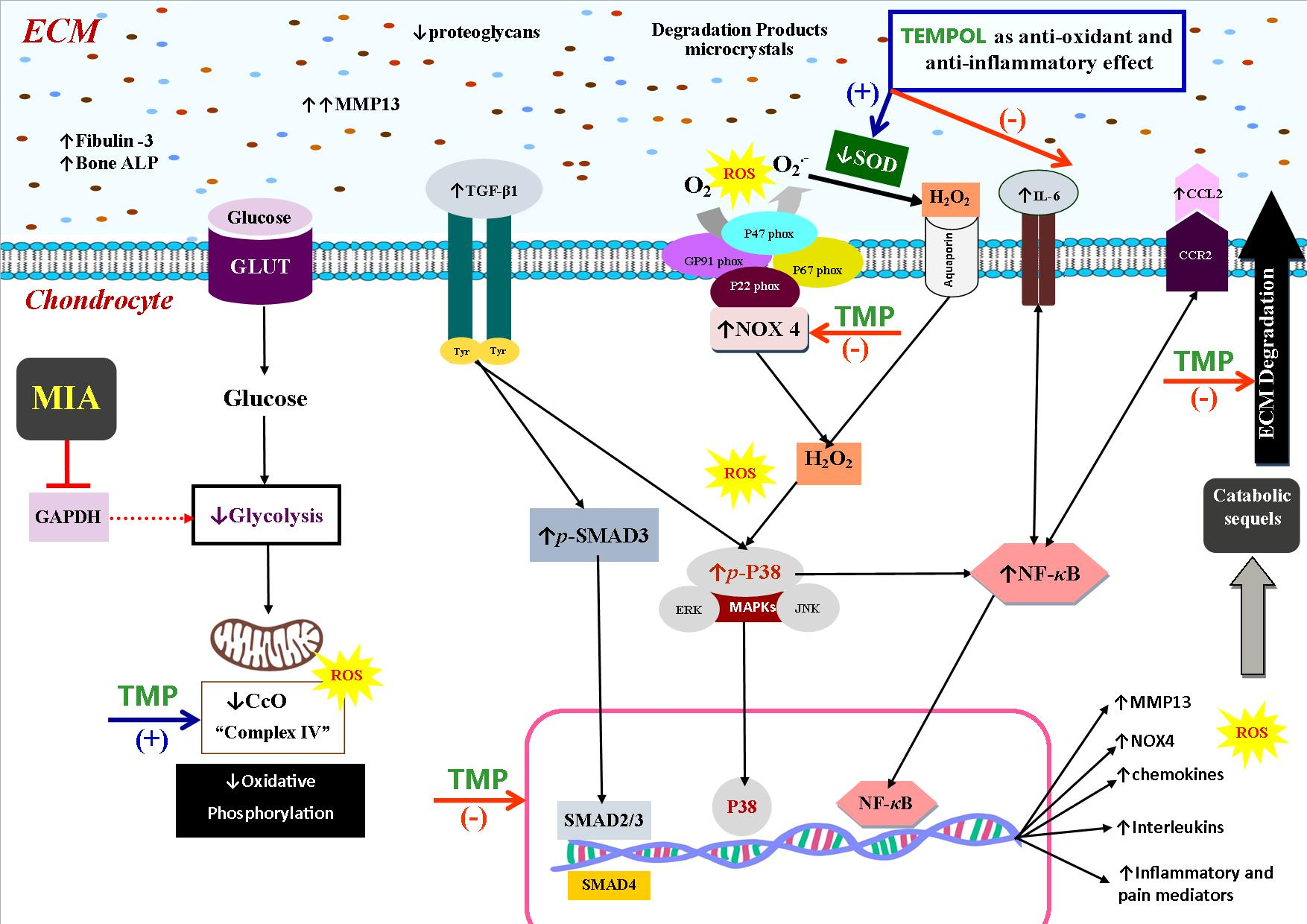
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Experimental Animals
4.3. Induction of Osteoarthritis
4.4. Drugs
4.5. Experimental Design
4.6. Assessment of Knee Joint Edema
4.7. Evaluation of Motor Performance Using Accelerating Rotarod Test
4.8. Radiographical Examination
4.9. Tissue Sampling
4.10. Biochemical ELISA Measurements
4.11. Quantitative Real-Time PCR (qRT-PCR) Measurements for Gene Expression
4.12. Western Blot Analysis
4.13. Histopathological Examinations
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Holyoak, D.T.; Tian, Y.F.; Van Der Meulen, M.C.H.; Singh, A. Osteoarthritis: Pathology, Mouse Models, and Nanoparticle Injectable Systems for Targeted Treatment. Ann. Biomed. Eng. 2016, 44, 2062–2075. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Chen, D.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Tong, Osteoarthritis Pathogenesis: A Review of Molecular Mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, I.; Matsuzaki, T.; Hoso, M. Long-term histopathological developments in knee-joint components in a rat model of osteoarthritis induced by monosodium iodoacetate. J. Phys. Ther. Sci. 2017, 29, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-G.; Park, S.-Y.; Chung, W.-S.; Park, J.-H.; Hwang, E.; Mavlonov, G.T.; Kim, I.-H.; Kim, K.-Y.; Yi, T.-H. Fucoidan Prevents the Progression of Osteoarthritis in Rats. J. Med. Food 2015, 18, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Cui, F.; Du, X.; Shang, G.; Xiao, W.; Yang, X.; Cui, Q. Antioxidative nanofullerol inhibits macrophage activation and development of osteoarthritis in rats. Int. J. Nanomed. 2019, 14, 4145–4155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkhodja, H.; Meddah, B.; Tirtouil, A.M.; Slimani, K.; Tou, A.; Hamza, B.; Boumediene, M.; Aicha, M.T.; Khaled, S.; Abdenacer, T. Radiographic and Histopathologic Analysis on Osteoarthritis Rat Model Treated with Essential Oils of Rosmarinus officinalis and Populus alba. Pharm. Sci. 2017, 23, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, S.M.; Lopes, D.M.; McMahon, S.B.; Dickenson, A.H. Characterisation of peripheral and central components of the rat monoiodoacetate model of Osteoarthritis. Osteoarthr. Cartil. 2019, 27, 712–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guingamp, C.; Gegout-Pottie, P.; Philippe, L.; Terlain, B.; Netter, P.; Gillet, P. Mono-iodoacetate-induced experimental osteoarthritis. A dose-response study of loss of mobility, morphology, and biochemistry. Arthritis Rheum. 1997, 40, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Park, S.-W.; Kang, J.-W.; Kim, Y.-J.; Lee, S.Y.; Shin, J.; Lee, S.; Lee, S.-M. Effect of GCSB-5, a Herbal Formulation, on Monosodium Iodoacetate-Induced Osteoarthritis in Rats. Evid. Based Complement. Altern. Med. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Valente, J.D.S. The Pharmacology of Pain Associated with the Monoiodoacetate Model of Osteoarthritis. Front. Pharmacol. 2019, 10, 974. [Google Scholar] [CrossRef]
- Henrotin, Y.; Pesesse, L.; Lambert, C. Targeting the synovial angiogenesis as a novel treatment approach to osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2014, 6, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Mobasheri, A.; Rayman, M.P.; Gualillo, O.; Sellam, J.; Van Der Kraan, P.; Fearon, U. The role of metabolism in the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2017, 13, 302–311. [Google Scholar] [CrossRef]
- Goldring, M.; Otero, M.; Plumb, D.; Dragomir, C.; Favero, M.; Hachem, K.E.; Hashimoto, K.; Roach, H.; Olivotto, E.; Borzì, R.; et al. Roles of inflammatory and anabolic cytokines in cartilage metabolism: Signals and multiple effectors converge upon MMP-13 regulation in osteoarthritis. Eur. Cells Mater. 2011, 21, 202–220. [Google Scholar] [CrossRef]
- Wei, F.-Y.; Lee, J.K.; Wei, L.; Qu, F.; Zhang, J.-Z. Correlation of insulin-like growth factor 1 and osteoarthritic cartilage degradation: A spontaneous osteoarthritis in guinea-pig. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 4493–4500. [Google Scholar]
- Rose, B.J.; Kooyman, D.L. A Tale of Two Joints: The Role of Matrix Metalloproteases in Cartilage Biology. Dis. Markers 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Choudhury, M.D.; Ghosh, P.; Palit, P. Discoidin domain receptor 2: An emerging pharmacological drug target for prospective therapy against osteoarthritis. Pharmacol. Rep. 2019, 71, 399–408. [Google Scholar] [CrossRef]
- Chen, J.; Crawford, R.; Xiao, Y. Vertical inhibition of the PI3K/Akt/mTOR pathway for the treatment of osteoarthritis. J. Cell. Biochem. 2013, 114, 245–249. [Google Scholar] [CrossRef]
- Takebe, K.; Nishiyama, T.; Hayashi, S.; Hashimoto, S.; Fujishiro, T.; Kanzaki, N.; Kawakita, K.; Iwasa, K.; Kuroda, R.; Kurosaka, M. Regulation of p38 MAPK phosphorylation inhibits chondrocyte apoptosis in response to heat stress or mechanical stress. Int. J. Mol. Med. 2011, 27, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Ding, H.; Peng, J.; Wang, X.; Pang, C.; Wei, J.; Wei, J.; Chen, H. Down-regulation of protease-activated receptor 2 ameliorated osteoarthritis in rats through regulation of MAPK/NF-κB signaling pathway in vivo and in vitro. Biosci. Rep. 2020, 40, BSR20192620. [Google Scholar] [CrossRef] [Green Version]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of Osteoarthritis of the Hand, Hip, and Knee. Arthritis Rheum. 2020, 72, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C.S.; Pearlman, A. Chemistry and Antihypertensive Effects of Tempol and Other Nitroxides. Pharmacol. Rev. 2008, 60, 418–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuzzocrea, S.; McDonald, M.C.; Mota-Filipe, H.; Mazzon, E.; Costantino, G.; Britti, D.; Mazzullo, G.; Caputi, A.P.; Thiemermann, C. Beneficial effects of tempol, a membrane-permeable radical scavenger, in a rodent model of collagen-induced arthritis. Arthritis Rheum. 2000, 43, 320–328. [Google Scholar] [CrossRef]
- Ali, S.M.; Okda, A.A.K.; Dessouky, I.S.; Hewedy, W.A.; Zahran, N.M.; Alamrani, B.A.-W. l-Carnitine ameliorates knee lesions in mono-iodoacetate induced osteoarthritis in rats. Alex. J. Med. 2017, 53, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.; Kothari, P.; Tripathi, A.K.; Singh, S.; Adhikary, S.; Ahmad, N.; Kumar, S.; Dev, K.; Mishra, V.K.; Shukla, S.; et al. Spinacia oleracea extract attenuates disease progression and sub-chondral bone changes in monosodium iodoacetate-induced osteoarthritis in rats. BMC Complement. Altern. Med. 2018, 18, 69. [Google Scholar] [CrossRef]
- Hasegawa, A.; Yonezawa, T.; Taniguchi, N.; Otabe, K.; Akasaki, Y.; Matsukawa, T.; Saito, M.; Neo, M.; Marmorstein, L.Y.; Lotz, M.K. Role of Fibulin 3 in Aging-Related Joint Changes and Osteoarthritis Pathogenesis in Human and Mouse Knee Cartilage. Arthritis Rheumatol. 2017, 69, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gögebakan, B.; İzmirli, M.; Okuyan, H.M.; Ataç, L. Biomarkers for Early Diagnosis of Osteoarthritis. In Osteoarthritis; SM Group: Dhaka, Bangladesh, 2016; pp. 1–15. Available online: www.smgebooks.com (accessed on 26 April 2020).
- Tsai, P.-W.; Lee, Y.-H.; Chen, L.-G.; Lee, C.-J.; Wang, C.-C. In Vitro and In Vivo Anti-Osteoarthritis Effects of 2,3,5,4′-Tetrahydroxystilbene-2-O-β-d-Glucoside from Polygonum Multiflorum. Molecules 2018, 23, 571. [Google Scholar] [CrossRef] [Green Version]
- Zahan, O.-M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Kogot-Levin, A.; Saada, A.; Leibowitz, G.; Soiferman, D.; Douiev, L.; Raz, I.; Weksler-Zangen, S. Upregulation of Mitochondrial Content in Cytochrome c Oxidase Deficient Fibroblasts. PLoS ONE 2016, 11, e0165417. [Google Scholar] [CrossRef]
- Pathak, N.N.; Balaganur, V.; Lingaraju, M.C.; Kant, V.; Kumar, D.; Kumar, D.; Sharma, A.K.; Tandan, S.K. Effect of atorvastatin, a HMG-CoA reductase inhibitor in monosodium iodoacetate-induced osteoarthritic pain: Implication for osteoarthritis therapy. Pharmacol. Rep. 2015, 67, 513–519. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, T.; Sun, H.; Wang, S.; Liu, M. Protective effects of dioscin against cartilage destruction in a monosodium iodoacetate (MIA)-indcued osteoarthritis rat model. Biomed. Pharmacother. 2018, 108, 1029–1038. [Google Scholar] [CrossRef]
- Izawa, T.; Hutami, I.R.; Mori, H.; Tanaka, E. Role of Smad3 and S1P Signaling in Mandibular Condylar Cartilage Homeostasis. J. Bone Res. 2017, 5, 1000178. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-de-Diego, C.; Valer, J.A.; Pimenta-Lopes, C.; Rosa, J.L.; Ventura, F. Interplay between BMPs and Reactive Oxygen Species in Cell Signaling and Pathology. Biomolecules 2019, 9, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavriilidis, C.; Miwa, S.; von Zglinicki, T.; Taylor, R.W.; Young, D.A. Mitochondrial dysfunction in osteoarthritis is associated with down-regulation of superoxide dismutase 2. Arthritis Rheum. 2013, 65, 378–387. [Google Scholar] [CrossRef]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive oxygen species, aging and articular cartilage homeostasis. Free. Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef]
- Wegner, A.M.; Campos, N.R.; Robbins, M.A.; Haddad, A.F.; Cunningham, H.C.; Yik, J.H.N.; Christiansen, B.A.; Haudenschild, D.R. Acute Changes in NADPH Oxidase 4 in Early Post-Traumatic Osteoarthritis. J. Orthop. Res. 2019, 37, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, L.; Xu, Z.; Yin, Y.; Su, J.; Niu, X.; Cao, X. Effects of estradiol on reduction of osteoarthritis in rabbits through effect on matrix metalloproteinase proteins. Iran. J. Basic Med. Sci. 2016, 19, 310–315. [Google Scholar]
- Aref-Eshghi, E.; Liu, M.; Harper, P.E.; Doré, J.; Martin, G.; Furey, A.; Green, R.; Rahman, P.; Zhai, G. Overexpression of MMP13 in human osteoarthritic cartilage is associated with the SMAD-independent TGF-β signalling pathway. Arthritis Res. Ther. 2015, 17, 264. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, L.A.; Shehata, N.I.; Abdelkader, N.; Khattab, M.M. Tempol, a Superoxide Dismutase Mimetic Agent, Ameliorates Cisplatin-Induced Nephrotoxicity through Alleviation of Mitochondrial Dysfunction in Mice. PLoS ONE 2014, 9, e108889. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.-H.; Tang, C.-C.; Chang, Y.-C.; Huang, S.-Y.; Hsieh, S.-P.; Lee, C.-H.; Huang, G.-S.; Ng, H.-F.; Neoh, C.-A.; Hsieh, C.-S.; et al. Glucosamine sulfate reduces experimental osteoarthritis and nociception in rats: Association with changes of mitogen-activated protein kinase in chondrocytes. Osteoarthr. Cartil. 2010, 18, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Yassin, N.; Yakoot, M.; El-Shenawy, S.; Abdel-Rahman, R.F.; Hassan, M.; Helmy, S. Effect of a topical copper indomethacin gel on inflammatory parameters in a rat model of osteoarthritis. Drug Des. Dev. Ther. 2015, 9, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.; Bae, K.; Kim, S.-G.; Kwak, D.; Moon, Y.-J.; Choi, C.-H.; Kim, Y.-R.; Na, C.-S.; Kim, S.-J. Anti-osteoarthritic effects of ChondroT in a rat model of collagenase-induced osteoarthritis. BMC Complement. Altern. Med. 2018, 18, 131. [Google Scholar] [CrossRef] [Green Version]
- Gundogdu, G.; Miloglu, F.D.; Gundogdu, K.; Tasci, S.Y.; Albayrak, M.; Demirci, T.; Cetin, M. Investigation of the efficacy of daidzein in experimental knee osteoarthritis-induced with monosodium iodoacetate in rats. Clin. Rheumatol. 2020, 39, 2399–2408. [Google Scholar] [CrossRef]
- Lee, Y.M.; Son, E.; Kim, S.-H.; Kim, O.S.; Kim, D.-S. Anti-inflammatory and anti-osteoarthritis effect of Mollugo pentaphylla extract. Pharm. Biol. 2019, 57, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Franz, A.; Joseph, L.; Mayer, C.; Harmsen, J.-F.; Schrumpf, H.; Fröbel, J.; Ostapczuk, M.S.; Krauspe, R.; Zilkens, C. The role of oxidative and nitrosative stress in the pathology of osteoarthritis: Novel candidate biomarkers for quantification of degenerative changes in the knee joint. Orthop. Rev. 2018, 10, 7460. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.-J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef]
- Yuan, J.; Ding, W.; Wu, N.; Jiang, S.; Li, W. Protective Effect of Genistein on Condylar Cartilage through Downregulating NF-κB Expression in Experimentally Created Osteoarthritis Rats. BioMed Res. Int. 2019, 2019, 2629791. [Google Scholar] [CrossRef] [Green Version]
- Olivotto, E.; Otero, M.; Marcu, K.B.; Goldring, M.B. Pathophysiology of osteoarthritis: Canonical NF-κB/IKKβ-dependent and kinase-independent effects of IKKα in cartilage degradation and chondrocyte differentiation. RMD Open 2015, 1, e000061. [Google Scholar] [CrossRef] [Green Version]
- di Paola, R.; Mazzon, E.; Zito, D.; Maiere, D.; Britti, D.; Genovese, T.; Cuzzocrea, S. Effects of Tempol, a membrane-permeable radical scavenger, in a rodent model periodontitis. J. Clin. Periodontol. 2005, 32, 1062–1068. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; Abdel-Rahman, R.F.; Abd-Elsalam, R.M. Neuroprotective effect of vildagliptin against cerebral ischemia in rats. Naunyn-Schmiedeberg′s Arch. Pharmacol. 2018, 391, 1133–1145. [Google Scholar] [CrossRef]
- Bowles, R.D.; Mata, B.A.; Bell, R.D.; Mwangi, T.K.; Huebner, J.L.; Kraus, V.B.; Setton, L.A. In vivo luminescence imaging of NF-κB activity and serum cytokine levels predict pain sensitivities in a rodent model of osteoarthritis. Arthritis Rheumatol. 2014, 66, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Pai, M.; Brederson, J.-D.; Wilcox, D.; Hsieh, G.; Jarvis, M.F.; Bitner, R.S. Monosodium Iodoacetate-Induced Joint Pain is Associated with Increased Phosphorylation of Mitogen Activated Protein Kinases in the Rat Spinal Cord. Mol. Pain 2011, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, D.; Wang, H.; Han, B.; Zhang, L.; Guo, J. Tempol Attenuates Neuropathic Pain by Inhibiting Nitric Oxide Production. Anal. Cell. Pathol. 2019, 2019, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.S.; Lee, I.; Chung, K.; Chung, J.M. Oxidative stress in the spinal cord is an important contributor in capsaicin-induced mechanical secondary hyperalgesia in mice. Pain 2008, 138, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.-W.; Lee, H.H.; Kim, J.; Choi, E.-O.; Hwang-Bo, H.; Kim, H.J.; Kim, M.Y.; Ahn, K.I.; Kim, G.-Y.; Lee, K.W.; et al. Mori Folium water extract alleviates articular cartilage damages and inflammatory responses in monosodium iodoacetate-induced osteoarthritis rats. Mol. Med. Rep. 2017, 16, 3841–3848. [Google Scholar] [CrossRef]
- Jeong, J.-H.; Moon, S.-J.; Jhun, J.-Y.; Yang, E.-J.; Cho, M.-L.; Min, J.-K. Eupatilin Exerts Antinociceptive and Chondroprotective Properties in a Rat Model of Osteoarthritis by Downregulating Oxidative Damage and Catabolic Activity in Chondrocytes. PLoS ONE 2015, 10, e0130882. [Google Scholar] [CrossRef]
- Tanito, M.; Li, F.; Elliott, M.H.; Dittmar, M.; Anderson, R.E. Protective Effect of TEMPOL Derivatives against Light-Induced Retinal Damage in Rats. Investig. Opthalmology Vis. Sci. 2007, 48, 1900–1905. [Google Scholar] [CrossRef] [Green Version]
- Karanovic, D.; Grujic-Milanovic, J.; Miloradović, Z.; Ivanov, M.; Jovovic, D.; Vajic, U.-J.; Zivotic, M.; Marković-Lipkovski, J.; Mihailovic-Stanojevic, N. Effects of Single and Combined Losartan and Tempol Treatments on Oxidative Stress, Kidney Structure and Function in Spontaneously Hypertensive Rats with Early Course of Proteinuric Nephropathy. PLoS ONE 2016, 11, e0161706. [Google Scholar] [CrossRef]
- Ranjbar, A.; Ghasemi, H.; Hatami, M.; Dadras, F.; Shayesteh, T.H.; Khoshjou, F. Tempol effects on diabetic nephropathy in male rats. J. Ren. Inj. Prev. 2016, 5, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-W.; Kim, J.; Choi, E.O.; Kwon, D.H.; Kong, G.M.; Choi, I.-W.; Kim, B.H.; Kim, G.-Y.; Lee, K.W.; Kim, K.Y.; et al. Schisandrae Fructus ethanol extract ameliorates inflammatory responses and articular cartilage damage in monosodium iodoacetate-induced osteoarthritis in rats. EXCLI J. 2017, 16, 265–277. [Google Scholar] [CrossRef]
- Tsai, H.-C.; Chen, T.-L.; Chen, Y.-P.; Chen, R.-M. Traumatic osteoarthritis-induced persistent mechanical hyperalgesia in a rat model of anterior cruciate ligament transection plus a medial meniscectomy. J. Pain Res. 2017, 11, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ji, L.; Yang, Y.; Wei, Y.; Zhang, X.; Gang, Y.; Lu, J.; Bai, L. The Therapeutic Effects of Treadmill Exercise on Osteoarthritis in Rats by Inhibiting the HDAC3/NF-KappaB Pathway in vivo and in vitro. Front. Physiol. 2019, 10, 1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellgren, J.H.; Lawrence, J.S. Radiological Assessment of Osteo-Arthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Culling, C.F.A. Handbook of Histopathological and Histochemical Techniques, 3rd ed.; Butterworths: London, UK, 2013. [Google Scholar]
- El-Gogary, R.I.; Khattab, M.A.; Abd-Allah, H. Intra-articular multifunctional celecoxib loaded hyaluronan nanocapsules for the suppression of inflammation in an osteoarthritic rat model. Int. J. Pharm. 2020, 583, 119378. [Google Scholar] [CrossRef]
- van der Sluijs, J.A.; Geesink, R.G.T.; van der Linden, A.J.; Bulstra, S.K.; Kuyer, R.; Drukker, J. The reliability of the mankin score for osteoarthritis. J. Orthop. Res. 1992, 10, 58–61. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA Species | Primer Sequence (5′‒3′) |
|---|---|
| chemokine (C-C motif) ligand 2 (CCL2) | F: AGCCAACTCTCACTGAAGC R: GTGAATTGAGTAGCAGCAGGT |
| Fibulin-3 (fib-3) | F: TGTGACCCAGGATATGAACTTGAG R: AGCCCCCTTGTAGATTGTAGCA |
| β-actin | F: CCAACCGCGAGAAGATGA R: CCAGAGGCGTACAGGGATAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-zalam, H.B.; Abdelsalam, R.M.; Abdel-Rahman, R.F.; Abd-Ellah, M.F.; Khattab, M.M. In Vivo Investigation of the Ameliorating Effect of Tempol against MIA-Induced Knee Osteoarthritis in Rats: Involvement of TGF-β1/SMAD3/NOX4 Cue. Molecules 2021, 26, 6993. https://doi.org/10.3390/molecules26226993
Abo-zalam HB, Abdelsalam RM, Abdel-Rahman RF, Abd-Ellah MF, Khattab MM. In Vivo Investigation of the Ameliorating Effect of Tempol against MIA-Induced Knee Osteoarthritis in Rats: Involvement of TGF-β1/SMAD3/NOX4 Cue. Molecules. 2021; 26(22):6993. https://doi.org/10.3390/molecules26226993
Chicago/Turabian StyleAbo-zalam, Hagar B., Rania M. Abdelsalam, Rehab F. Abdel-Rahman, Mohamed F. Abd-Ellah, and Mahmoud M. Khattab. 2021. "In Vivo Investigation of the Ameliorating Effect of Tempol against MIA-Induced Knee Osteoarthritis in Rats: Involvement of TGF-β1/SMAD3/NOX4 Cue" Molecules 26, no. 22: 6993. https://doi.org/10.3390/molecules26226993